
Differential Therapeutic Effect of Extracellular Vesicles Derived by Bone Marrow and Adipose Mesenchymal Stem Cells on Wound Healing of Diabetic Ulcers and Correlation to Their Cargoes

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
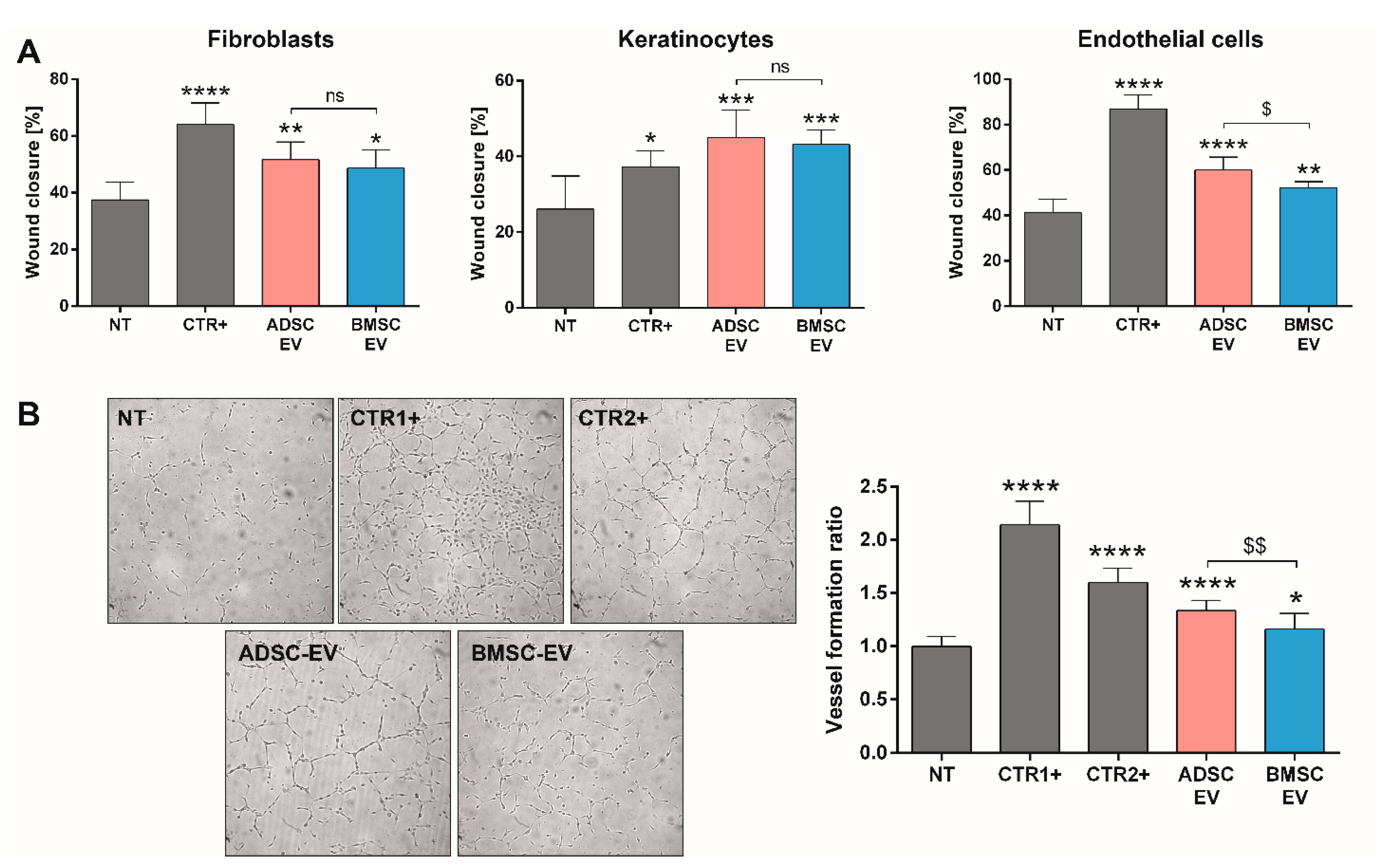
2.1. Experimental Results
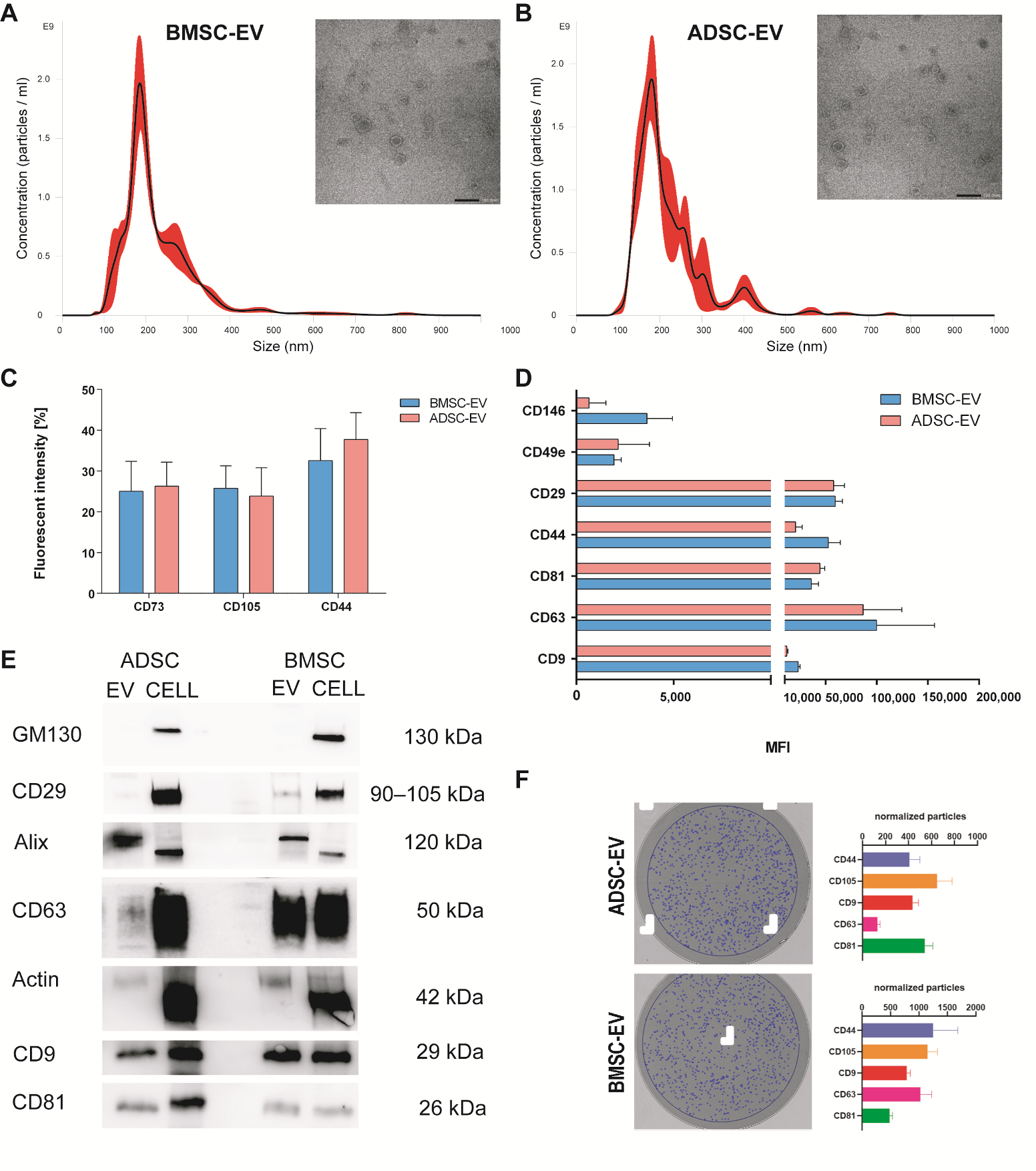
2.1.1. Characterization of BMSC and ADSC-EVs
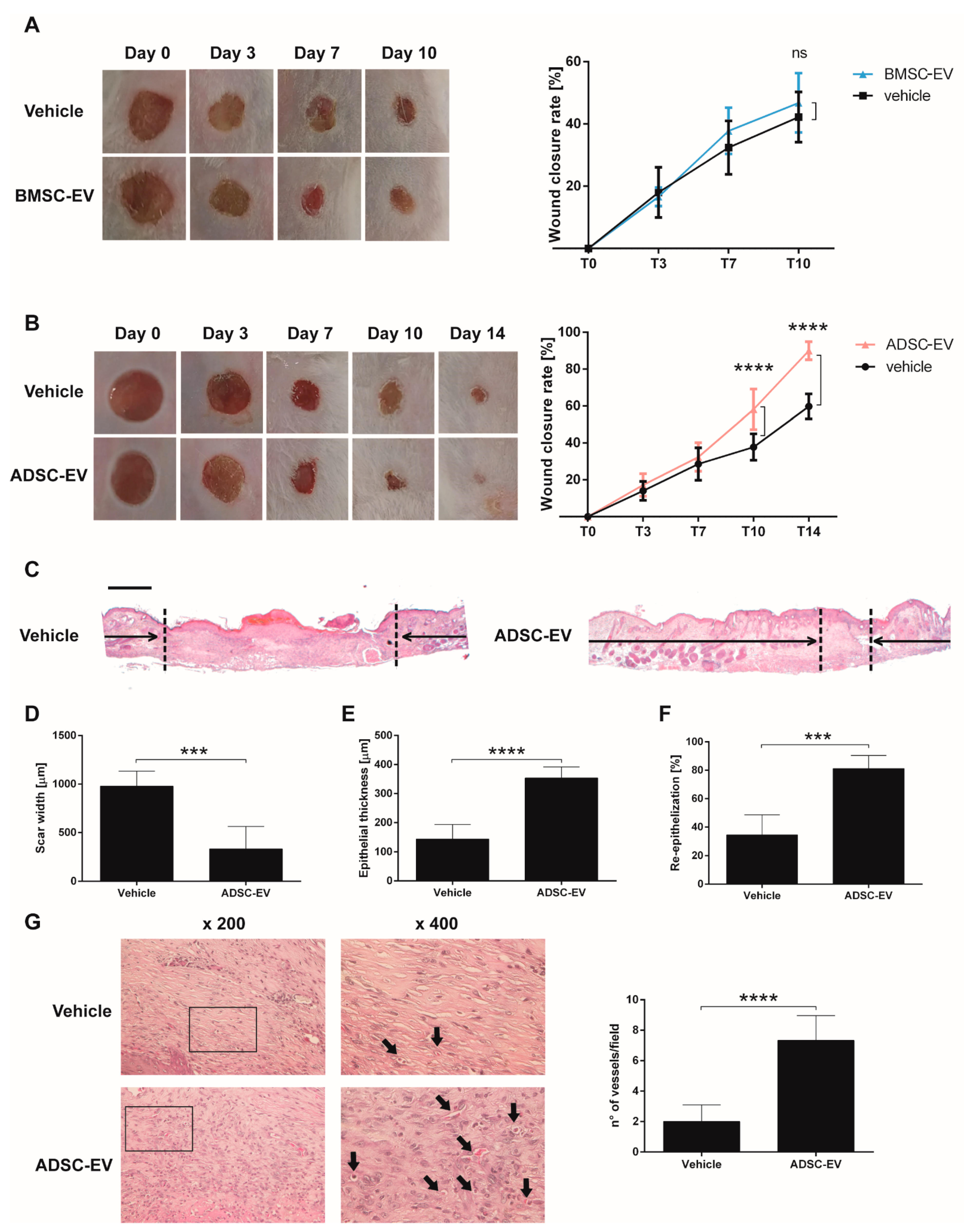
2.1.2. BMSC and ADSC-EVs Exerted Different Therapeutic Action on Wound Healing in a Mouse Model of Diabetic Ulcers
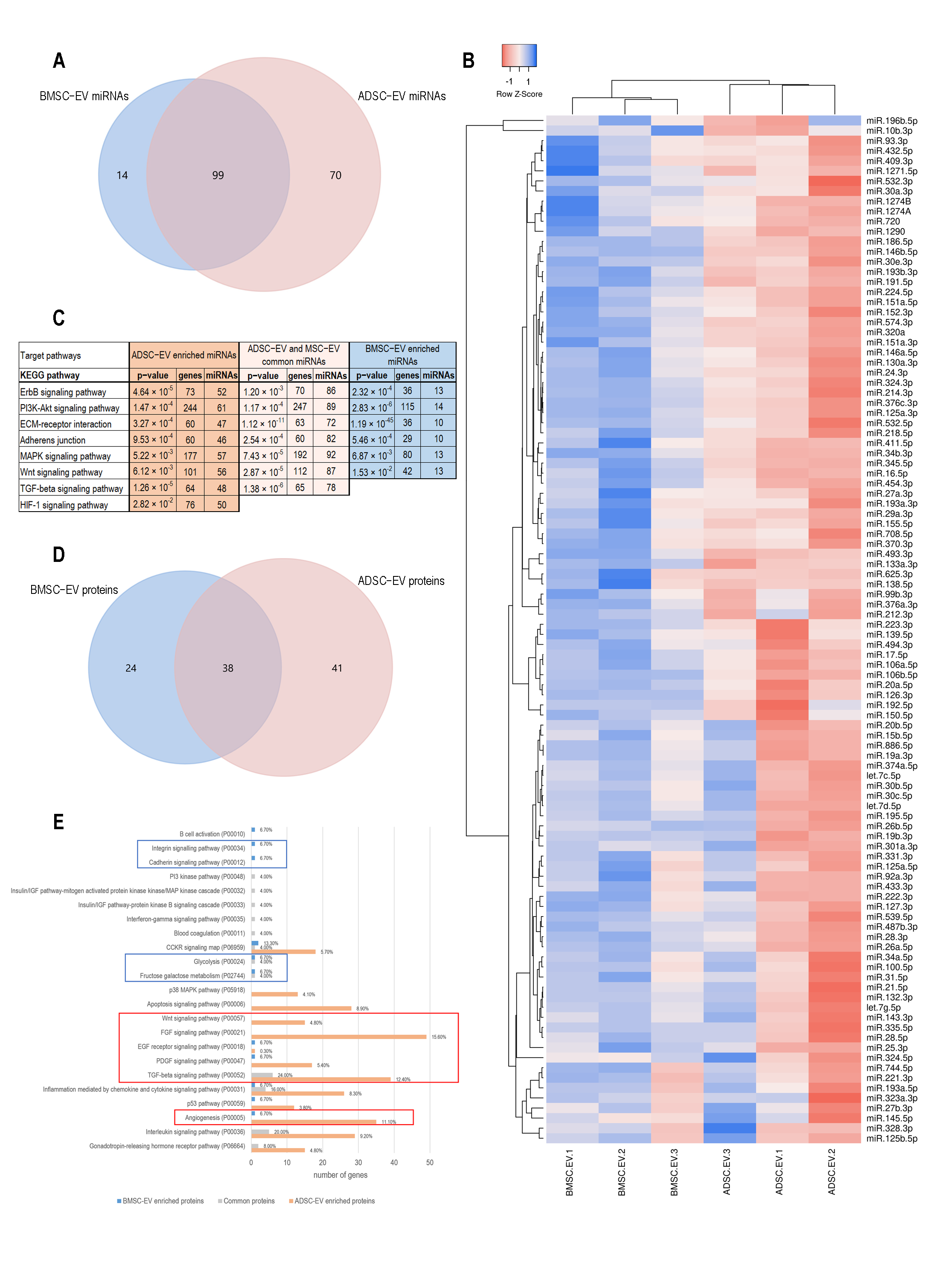
2.1.3. BMSC and ADSC-EV Cargo Analysis Revealed Different Target Pathways
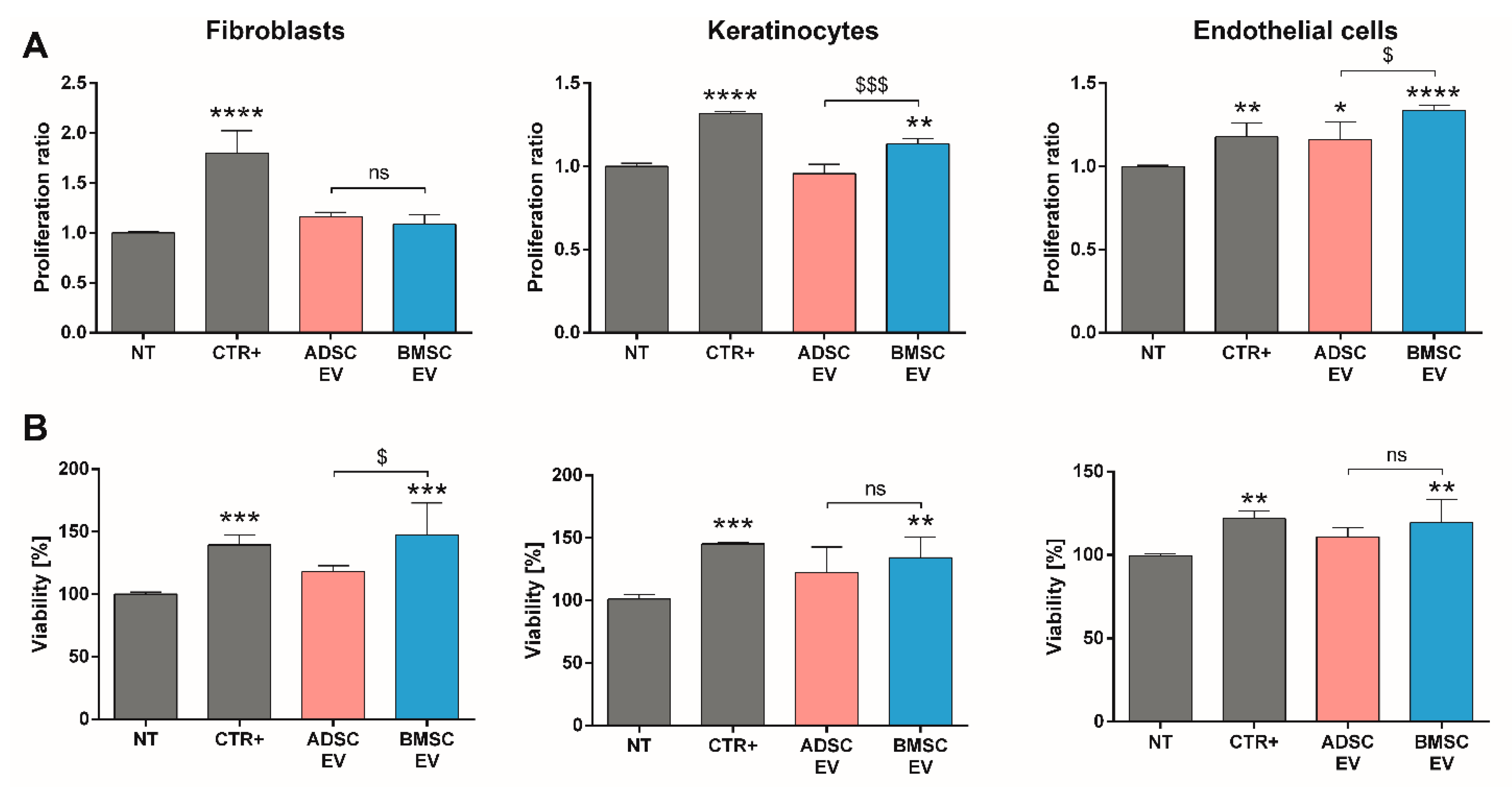
2.1.4. A Direct Comparison of BMSC and ADSC-EVs Revealed Different Beneficial Effects In Vitro
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Isolation of EVs
4.3. Nanosight Analysis of EVs
4.4. Transmission Electron Microscopy Analysis of EVs
4.5. FACS Characterization of EVs
4.6. Protein Measurement and Western Blot Analysis
4.7. Single Particle Interferometric Reflectance Imaging Sensor with ExoView®
4.8. Diabetic Wound Healing Mouse Model
4.9. In Vitro Proliferation Assay
4.10. In Vitro Viability Assay
4.11. In Vitro Scratch Test
4.12. In Vitro Formation of Capillary-Like Structures Assay
4.13. miRNA Expression Analysis
4.14. Target Pathway and Enrichment Analysis
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of Extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Gould, S.J.; Raposo, G. As we wait: Coping with an imperfect nomenclature for Extracellular Vesicles. J. Extracell. Vesicles 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other Extracellular Vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.; Kholia, S.; Deregibus, M.C.; Camussi, G. The Role of Extracellular Vesicles as Paracrine Effectors in Stem Cell-Based Therapies. Adv. Exp. Med. Biol. 2019, 1201, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Viejo, M. Mesenchymal stem cells from different sources and their derived exosomes: A pre-clinical perspective. World J. Stem Cells 2020, 12, 100–109. [Google Scholar] [CrossRef]
- Bochon, B.; Kozubska, M.; Surygała, G.; Witkowska, A.; Kuźniewicz, R.; Grzeszczak, W.; Wystrychowski, G. Mesenchymal Stem Cells-Potential Applications in Kidney Diseases. Int. J. Mol. Sci. 2019, 20, 2462. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.H.; Nguyen, T.D.; Nguyen, H.P.; Nguyen, X.H.; Do, P.; Dang, V.D.; Dam, P.; Bui, H.; Trinh, M.Q.; Vu, D.M.; et al. Differential Wound Healing Capacity of Mesenchymal Stem Cell-Derived Exosomes Originated from Bone Marrow, Adipose Tissue and Umbilical Cord Under Serum- and Xeno-Free Condition. Front. Mol. Biosci. 2020, 7, 119. [Google Scholar] [CrossRef]
- Bielefeld, K.A.; Amini-Nik, S.; Alman, B.A. Cutaneous wound healing: Recruiting developmental pathways for regeneration. Cell Mol. Life Sci. 2013, 70, 2059–2081. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Dipietro, L.A. Factors affecting wound healing. J. Dent. Res. 2010, 89, 219–229. [Google Scholar] [CrossRef]
- Hu, L.; Wang, J.; Zhou, X.; Xiong, Z.; Zhao, J.; Yu, R.; Huang, F.; Zhang, H.; Chen, L. Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Sci. Rep. 2016, 6, 32993. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Dong, Z.; Cao, Y.; Wang, H.; Liu, S.; Liao, L.; Jin, Y.; Yuan, L.; Li, B. MSC-Derived Exosome Promotes M2 Polarization and Enhances Cutaneous Wound Healing. Stem Cells Int. 2019, 2019, 7132708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, S.; Chen, J.; Duscher, D.; Liu, Y.; Guo, G.; Kang, Y.; Xiong, H.; Zhan, P.; Wang, Y.; Wang, C.; et al. Microvesicles from human adipose stem cells promote wound healing by optimizing cellular functions via AKT and ERK signaling pathways. Stem Cell Res. Ther. 2019, 10, 47. [Google Scholar] [CrossRef]
- Shafei, S.; Khanmohammadi, M.; Heidari, R.; Ghanbari, H.; Taghdiri Nooshabadi, V.; Farzamfar, S.; Akbariqomi, M.; Sanikhani, N.S.; Absalan, M.; Tavoosidana, G. Exosome loaded alginate hydrogel promotes tissue regeneration in full-thickness skin wounds: An in vivo study. J. Biomed. Mater. Res. A 2020, 108, 545–556. [Google Scholar] [CrossRef]
- Zhang, W.; Bai, X.; Zhao, B.; Li, Y.; Zhang, Y.; Li, Z.; Wang, X.; Luo, L.; Han, F.; Zhang, J.; et al. Cell-free therapy based on adipose tissue stem cell-derived exosomes promotes wound healing via the PI3K/Akt signaling pathway. Exp. Cell Res. 2018, 370, 333–342. [Google Scholar] [CrossRef]
- Yager, D.R.; Kulina, R.A.; Gilman, L.A. Wound fluids: A window into the wound environment? Int. J. Low Extrem Wounds 2007, 6, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, X.; Lian, W.; Shi, R.; Han, S.; Zhang, H.; Lu, L.; Li, M. Exosomes from adipose-derived stem cells overexpressing Nrf2 accelerate cutaneous wound healing by promoting vascularization in a diabetic foot ulcer rat model. Exp. Mol. Med. 2018, 50, 29. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wang, M.; Xu, T.; Zhang, X.; Lin, C.; Gao, W.; Xu, H.; Lei, B.; Mao, C. Engineering Bioactive Self-Healing Antibacterial Exosomes Hydrogel for Promoting Chronic Diabetic Wound Healing and Complete Skin Regeneration. Theranostics 2019, 9, 65–76. [Google Scholar] [CrossRef]
- Ding, J.; Wang, X.; Chen, B.; Zhang, J.; Xu, J. Exosomes Derived from Human Bone Marrow Mesenchymal Stem Cells Stimulated by Deferoxamine Accelerate Cutaneous Wound Healing by Promoting Angiogenesis. Biomed. Res. Int. 2019, 2019, 9742765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Yu, M.; Xie, D.; Wang, L.; Ye, C.; Zhu, Q.; Liu, F.; Yang, L. Melatonin-stimulated MSC-derived exosomes improve diabetic wound healing through regulating macrophage M1 and M2 polarization by targeting the PTEN/AKT pathway. Stem Cell Res. Ther. 2020, 11, 259. [Google Scholar] [CrossRef]
- Yu, M.; Liu, W.; Li, J.; Lu, J.; Lu, H.; Jia, W.; Liu, F. Exosomes derived from atorvastatin-pretreated MSC accelerate diabetic wound repair by enhancing angiogenesis via AKT/eNOS pathway. Stem Cell Res. Ther. 2020, 11, 350. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Luan, S.; Chen, J.; Zhou, Y.; Wang, T.; Li, Z.; Fu, Y.; Zhai, A.; Bi, C. The MSC-Derived Exosomal lncRNA H19 Promotes Wound Healing in Diabetic Foot Ulcers by Upregulating PTEN via MicroRNA-152-3p. Mol. Ther. Nucleic Acids 2020, 19, 814–826. [Google Scholar] [CrossRef] [PubMed]
- Assunção-Silva, R.C.; Mendes-Pinheiro, B.; Patrício, P.; Behie, L.A.; Teixeira, F.G.; Pinto, L.; Salgado, A.J. Exploiting the impact of the secretome of MSCs isolated from different tissue sources on neuronal differentiation and axonal growth. Biochimie 2018, 155, 83–91. [Google Scholar] [CrossRef]
- Pires, A.O.; Mendes-Pinheiro, B.; Teixeira, F.G.; Kruminis-Kaszkiel, E.; Osowski, A.; Bejer-Oleńska, E.; Dziekoński, M.; Wojtkiewicz, J. Unveiling the Differences of Secretome of Human Bone Marrow Mesenchymal Stem Cells, Adipose Tissue-Derived Stem Cells, and Human Umbilical Cord Perivascular Cells: A Proteomic Analysis. Stem Cells Dev. 2016, 25, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, C.; Nagaya, N.; Ohnishi, S.; Yamahara, K.; Takabatake, S.; Konno, T.; Hayashi, K.; Kawashiri, M.A.; Tsubokawa, T.; Yamagishi, M. Gene and protein expression analysis of mesenchymal stem cells derived from rat adipose tissue and bone marrow. Circ. J. 2011, 75, 2260–2268. [Google Scholar] [CrossRef] [Green Version]
- Del Fattore, A.; Luciano, R.; Saracino, R.; Battafarano, G.; Rizzo, C.; Pascucci, L.; Alessandri, G.; Pessina, A.; Perrotta, A.; Fierabracci, A.; et al. Differential effects of Extracellular vesicles secreted by mesenchymal stem cells from different sources on glioblastoma cells. Expert Opin. Biol. Ther. 2015, 15, 495–504. [Google Scholar] [CrossRef]
- Xu, H.; Wang, Z.; Liu, L.; Zhang, B.; Li, B. Exosomes derived from adipose tissue, bone marrow, and umbilical cord blood for cardioprotection after myocardial infarction. J. Cell Biochem. 2020, 121, 2089–2102. [Google Scholar] [CrossRef] [PubMed]
- Katsuda, T.; Tsuchiya, R.; Kosaka, N.; Yoshioka, Y.; Takagaki, K.; Oki, K.; Takeshita, F.; Sakai, Y.; Kuroda, M.; Ochiyaa, T. Human adipose tissue-derived mesenchymal stem cells secrete functional neprilysin-bound exosomes. Sci. Rep. 2013, 3, 1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baglio, S.R.; Rooijers, K.; Koppers-Lalic, D.; Verweij, F.J.; Pérez Lanzón, M.; Zini, N.; Naaijkens, B.; Perut, F.; Niessen, H.W.; Baldini, N.; et al. Human bone marrow- and adipose-mesenchymal stem cells secrete exosomes enriched in distinctive miRNA and tRNA species. Stem Cell Res. Ther. 2015, 6, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Wang, X.; Zhao, Z.; Chen, J.; Li, C.; Zhao, G. Paeoniflorin accelerates foot wound healing in diabetic rats though activating the Nrf2 pathway. Acta Histochem. 2020, 122, 151649. [Google Scholar] [CrossRef]
- Collino, F.; Pomatto, M.; Bruno, S.; Lindoso, R.S.; Tapparo, M.; Sicheng, W.; Quesenberry, P.; Camussi, G. Exosome and Microvesicle-Enriched Fractions Isolated from Mesenchymal Stem Cells by Gradient Separation Showed Different Molecular Signatures and Functions on Renal Tubular Epithelial Cells. Stem Cell Rev. Rep. 2017, 13, 226–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figliolini, F.; Ranghino, A.; Grange, C.; Cedrino, M.; Tapparo, M.; Cavallari, C.; Rossi, A.; Togliatto, G.; Femminò, S.; Gugliuzza, M.V.; et al. Extracellular Vesicles from Adipose Stem Cells Prevent Muscle Damage and Inflammation in a Mouse Model of Hind Limb Ischemia: Role of Neuregulin-1. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 239–254. [Google Scholar] [CrossRef]
- Lopatina, T.; Favaro, E.; Grange, C.; Cedrino, M.; Ranghino, A.; Occhipinti, S.; Fallo, S.; Buffolo, F.; Gaykalova, D.A.; Zanone, M.M.; et al. PDGF enhances the protective effect of adipose stem cell-derived Extracellular vesicles in a model of acute hindlimb ischemia. Sci. Rep. 2018, 8, 17458. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Li, L.; He, Y.; Zhao, M.; Jiang, J. Collective cell migration: Implications for wound healing and cancer invasion. Burns Trauma 2013, 1, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lioupis, C. Effects of diabetes mellitus on wound healing: An update. J. Wound Care 2005, 14, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Pelizzo, G.; Avanzini, M.A.; Icaro Cornaglia, A.; De Silvestri, A.; Mantelli, M.; Travaglino, P.; Croce, S.; Romano, P.; Avolio, L.; Iacob, G.; et al. Extracellular vesicles derived from mesenchymal cells: Perspective treatment for cutaneous wound healing in pediatrics. Regen. Med. 2018, 13, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, C.; Zhao, J. Human endothelial progenitor cells-derived exosomes accelerate cutaneous wound healing in diabetic rats by promoting endothelial function. J. Diabetes Complicat. 2016, 30, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.J.; Kuan, C.H.; Wu, H.C.; Tsai, J.C.; Chen, T.M.; Hsieh, D.J.; Wang, T.W. Tailored design of electrospun composite nanofibers with staged release of multiple angiogenic growth factors for chronic wound healing. Acta Biomater. 2014, 10, 4156–4166. [Google Scholar] [CrossRef]
- Kota, S.K.; Meher, L.K.; Jammula, S.; Kota, S.K.; Krishna, S.V.; Modi, K.D. Aberrant angiogenesis: The gateway to diabetic complications. Indian J. Endocrinol Metab. 2012, 16, 918–930. [Google Scholar] [CrossRef]
- Shi, R.; Jin, Y.; Hu, W.; Lian, W.; Cao, C.; Han, S.; Zhao, S.; Yuan, H.; Yang, X.; Shi, J.; et al. Exosomes derived from mmu_circ_0000250-modified adipose-derived mesenchymal stem cells promote wound healing in diabetic mice by inducing miR-128-3p/SIRT1-mediated autophagy. Am. J. Physiol. Cell Physiol. 2020, 318, C848–C856. [Google Scholar] [CrossRef] [PubMed]
- Gai, C.; Gomez, Y.; Tetta, C.; Brizzi, M.F.; Camussi, G. Protective Role of Stem Cell Derived Extracellular Vesicles in an In Vitro Model of Hyperglycemia-Induced Endothelial Injury. J. Cell Sci. Ther. 2017, 8, 2. [Google Scholar] [CrossRef]
- Okonkwo, U.A.; Chen, L.; Ma, D.; Haywood, V.A.; Barakat, M.; Urao, N.; DiPietro, L.A. Compromised angiogenesis and vascular Integrity in impaired diabetic wound healing. PLoS ONE 2020, 15, e0231962. [Google Scholar] [CrossRef]
- Shiojima, I.; Walsh, K. Role of Akt signaling in vascular homeostasis and angiogenesis. Circ. Res. 2002, 90, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, X.; Ma, K.; Zhang, C.; Fu, X. Therapeutic angiogenesis using stem cell-derived Extracellular vesicles: An emerging approach for treatment of ischemic diseases. Stem Cell Res. Ther. 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Wang, S.; Han, Q.; Zhao, R.C. Exosomes secreted by mesenchymal stem cells promote endothelial cell angiogenesis by transferring miR-125a. J. Cell Sci. 2016, 129, 2182–2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.C.; Guo, S.C.; Li, M.; Ke, Q.F.; Guo, Y.P.; Zhang, C.Q. Chitosan Wound Dressings Incorporating Exosomes Derived from MicroRNA-126-Overexpressing Synovium Mesenchymal Stem Cells Provide Sustained Release of Exosomes and Heal Full-Thickness Skin Defects in a Diabetic Rat Model. Stem Cells Transl. Med. 2017, 6, 736–747. [Google Scholar] [CrossRef] [PubMed]
- Cavallari, C.; Figliolini, F.; Tapparo, M.; Cedrino, M.; Trevisan, A.; Positello, L.; Rispoli, P.; Solini, A.; Migliaretti, G.; Camussi, G.; et al. miR-130a and Tgfβ Content in Extracellular Vesicles Derived from the Serum of Subjects at High Cardiovascular Risk Predicts their In-Vivo Angiogenic Potential. Sci. Rep. 2020, 10, 706. [Google Scholar] [CrossRef]
- Geiger, A.; Walker, A.; Nissen, E. Human fibrocyte-derived exosomes accelerate wound healing in genetically diabetic mice. Biochem. Biophys. Res. Commun. 2015, 467, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Togliatto, G.; Dentelli, P.; Gili, M.; Gallo, S.; Deregibus, C.; Biglieri, E.; Iavello, A.; Santini, E.; Rossi, C.; Solini, A.; et al. Obesity reduces the pro-angiogenic potential of adipose tissue stem cell-derived Extracellular vesicles (EVs) by impairing miR-126 content: Impact on clinical applications. Int. J. Obes. 2016, 40, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, X.; Zhou, L.; Liu, Y.; Gu, J.; Mi, Q. MicroRNA-146a deficiency delays wound healing in normal and diabetic mice. Adv. Wound Care 2021. [Google Scholar] [CrossRef]
- Cheng, S.; Xi, Z.; Chen, G.; Liu, K.; Ma, R.; Zhou, C. Extracellular vesicle-carried microRNA-27b derived from mesenchymal stem cells accelerates cutaneous wound healing via E3 ubiquitin ligase ITCH. J. Cell Mol. Med. 2020, 24, 11254–11271. [Google Scholar] [CrossRef]
- Hettich, B.F.; Ben-Yehuda Greenwald, M.; Werner, S.; Leroux, J.C. Exosomes for Wound Healing: Purification Optimization and Identification of Bioactive Components. Adv. Sci. 2020, 7, 2002596. [Google Scholar] [CrossRef] [PubMed]
- Chance, T.C.; Herzig, M.C.; Christy, B.A.; Delavan, C.; Rathbone, C.R.; Cap, A.P.; Bynum, J.A. Human mesenchymal stromal cell source and culture conditions influence Extracellular vesicle angiogenic and metabolic effects on human endothelial cells in vitro. J. Trauma Acute Care Surg. 2020, 89 (Suppl. 2), S100–S108. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Rao, S.S.; Wang, Z.X.; Cao, J.; Tan, Y.J.; Luo, J.; Li, H.M.; Zhang, W.S.; Chen, C.Y.; Xie, H. Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function. Theranostics 2018, 8, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, J.; Sen, C.K. microRNA and Wound Healing. Adv. Exp. Med. Biol. 2015, 888, 291–305. [Google Scholar] [CrossRef] [Green Version]
- Mahdi, A.; Darvishi, B.; Majidzadeh-A, K.; Salehi, M.; Farahmand, L. Challenges facing antiangiogenesis therapy: The significant role of hypoxia-inducible factor and MET in development of resistance to anti-vascular endothelial growth factor-targeted therapies. J. Cell Physiol. 2019, 234, 5655–5663. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, B.H.; Lee, H.; Jeon, B.; Lee, Y.S.; Kwon, M.J.; Kim, T.Y. Regulation of skin inflammation and angiogenesis by EC-SOD via HIF-1α and NF-κB pathways. Free Radic. Biol Med. 2011, 51, 1985–1995. [Google Scholar] [CrossRef]
- Li, Y.; Sun, R.; Zou, J.; Ying, Y.; Luo, Z. Dual Roles of the AMP-Activated Protein Kinase Pathway in Angiogenesis. Cells 2019, 8, 752. [Google Scholar] [CrossRef] [Green Version]
- Cavallari, C.; Ranghino, A.; Tapparo, M.; Cedrino, M.; Figliolini, F.; Grange, C.; Giannachi, V.; Garneri, P.; Deregibus, M.C.; Collino, F.; et al. Serum-derived Extracellular vesicles (EVs) impact on vascular remodeling and prevent muscle damage in acute hind limb ischemia. Sci. Rep. 2017, 7, 8180. [Google Scholar] [CrossRef]
- Casado-Díaz, A.; Quesada-Gómez, J.M.; Dorado, G. Extracellular Vesicles Derived from Mesenchymal Stem Cells (MSC) in Regenerative Medicine: Applications in Skin Wound Healing. Front. Bioeng. Biotechnol. 2020, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wu, X.; Zhang, X.; Sun, Y.; Yan, Y.; Shi, H.; Zhu, Y.; Wu, L.; Pan, Z.; Zhu, W.; et al. Human umbilical cord mesenchymal stem cell exosomes enhance angiogenesis through the Wnt4/β-catenin pathway. Stem Cells Transl. Med. 2015, 4, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Golchin, A.; Hosseinzadeh, S.; Ardeshirylajimi, A. The exosomes released from different cell types and their effects in wound healing. J. Cell Biochem. 2018, 119, 5043–5052. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Park, J.; Liu, A.; Lee, J.; Zhang, X.; Hao, Q.; Lee, J.W. Mesenchymal Stem Cell Microvesicles Restore Protein Permeability Across Primary Cultures of Injured Human Lung Microvascular Endothelial Cells. Stem Cells Transl. Med. 2018, 7, 615–624. [Google Scholar] [CrossRef]
- Kou, X.; Xu, X.; Chen, C.; Sanmillan, M.L.; Cai, T.; Zhou, Y.; Giraudo, C.; Le, A.; Shi, S. The Fas/Fap-1/Cav-1 complex regulates IL-1RA secretion in mesenchymal stem cells to accelerate wound healing. Sci. Transl. Med. 2018, 10, eaai8524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.Y.; Suh, H.N.; Choi, G.E.; Lee, H.J.; Jung, Y.H.; Ko, S.H.; Kim, J.S.; Chae, C.W.; Lee, C.K.; Han, H.J. Modulation of sonic hedgehog-induced mouse embryonic stem cell behaviours through E-cadherin expression and integrin β1-dependent F-actin formation. Br. J. Pharmacol. 2018, 175, 3548–3562. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E. Skin stem cells: Rising to the surface. J. Cell Biol. 2008, 180, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Longmate, W.M.; Varney, S.; Power, D.; Miskin, R.P.; Anderson, K.E.; DeFreest, L.; Van De Water, L.; DiPersio, C.M. Integrin α3β1 on Tumor Keratinocytes Is Essential to Maintain Tumor Growth and Promotes a Tumor-Supportive Keratinocyte Secretome. J. Investig. Dermatol. 2021, 141, 142–151.e6. [Google Scholar] [CrossRef]
- Henning, C.; Branopolski, A.; Follert, P.; Lewandowska, O.; Ayhan, A.; Benkhoff, M.; Flögel, U.; Kelm, M.; Heiss, C.; Lammert, E. Endothelial β1 Integrin-Mediated Adaptation to Myocardial Ischemia. Thromb. Haemost. 2021. [Google Scholar] [CrossRef]
- Coelho, A.I.; Berry, G.T.; Rubio-Gozalbo, M.E. Galactose metabolism and health. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 422–427. [Google Scholar] [CrossRef]
- Lunt, S.Y.; Vander Heiden, M.G. Aerobic glycolysis: Meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Rao, S.S.; Ren, L.; Hu, X.K.; Tan, Y.J.; Hu, Y.; Luo, J.; Liu, Y.W.; Yin, H.; Huang, J.; et al. Exosomal DMBT1 from human urine-derived stem cells facilitates diabetic wound repair by promoting angiogenesis. Theranostics 2018, 8, 1607–1623. [Google Scholar] [CrossRef]
- Ha, D.H.; Kim, H.K.; Lee, J.; Kwon, H.H.; Park, G.H.; Yang, S.H.; Jung, J.Y.; Choi, H.; Lee, J.H.; Sung, S.; et al. Mesenchymal Stem/Stromal Cell-Derived Exosomes for Immunomodulatory Therapeutics and Skin Regeneration. Cells 2020, 9, 1157. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, T.; Bruno, S.; Tetta, C.; Kalinina, N.; Porta, M.; Camussi, G. Platelet-derived growth factor regulates the secretion of Extracellular vesicles by adipose mesenchymal stem cells and enhances their angiogenic potential. Cell Commun. Signal. 2014, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Collino, F.; Lopes, J.A.; Corrêa, S.; Abdelhay, E.; Takiya, C.M.; Wendt, C.; de Miranda, K.R.; Vieyra, A.; Lindoso, R.S. Adipose-Derived Mesenchymal Stromal Cells Under Hypoxia: Changes in Extracellular Vesicles Secretion and Improvement of Renal Recovery after Ischemic Injury. Cell Physiol. Biochem. 2019, 52, 1463–1483. [Google Scholar] [CrossRef]
- Deregibus, M.C.; Figliolini, F.; D’Antico, S.; Manzini, P.M.; Pasquino, C.; De Lena, M.; Tetta, C.; Brizzi, M.F.; Camussi, G. Charge-based precipitation of Extracellular Vesicles. Int. J. Mol. Med. 2016, 38, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- Bruno, S.; Tapparo, M.; Collino, F.; Chiabotto, G.; Deregibus, M.C.; Soares Lindoso, R.; Neri, F.; Kholia, S.; Giunti, S.; Wen, S.; et al. Renal Regenerative Potential of Different Extracellular Vesicle Populations Derived from Bone Marrow Mesenchymal Stromal Cells. Tissue Eng. Part A 2017, 23, 1262–1273. [Google Scholar] [CrossRef] [PubMed]
- Kholia, S.; Herrera Sanchez, M.B.; Cedrino, M.; Papadimitriou, E.; Tapparo, M.; Deregibus, M.C.; Bruno, S.; Antico, F.; Brizzi, M.F.; Quesenberry, P.J.; et al. Mesenchymal Stem Cell Derived Extracellular Vesicles Ameliorate Kidney Injury in Aristolochic Acid Nephropathy. Front. Cell Dev. Biol. 2020, 8, 188. [Google Scholar] [CrossRef] [PubMed]
- Daaboul, G.G.; Gagni, P.; Benussi, L.; Bettotti, P.; Ciani, M.; Cretich, M.; Freedman, D.S.; Ghidoni, R.; Ozkumur, A.Y.; Piotto, C.; et al. Digital Detection of Exosomes by Interferometric Imaging. Sci. Rep. 2016, 6, 37246. [Google Scholar] [CrossRef] [PubMed]
- Grange, C.; Tritta, S.; Tapparo, M.; Cedrino, M.; Tetta, C.; Camussi, G.; Brizzi, M.F. Stem cell-derived Extracellular vesicles inhibit and revert fibrosis progression in a mouse model of diabetic nephropathy. Sci. Rep. 2019, 9, 4468. [Google Scholar] [CrossRef] [Green Version]
- Pathan, M.; Keerthikumar, S.; Chisanga, D.; Alessandro, R.; Ang, C.S.; Askenase, P.; Batagov, A.O.; Benito-Martin, A.; Camussi, G.; Clayton, A.; et al. A novel community driven software for functional enrichment analysis of Extracellular vesicles data. J. Extracell. Vesicles 2017, 6, 1321455. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.D.; Kejariwal, A.; Guo, N.; Mi, H.; Campbell, M.J.; Muruganujan, A.; Lazareva-Ulitsky, B. Applications for protein sequence-function evolution data: mRNA/protein expression analysis and coding SNP scoring tools. Nucleic Acids Res. 2006, 34, W645–W650. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common | BMSC-EV | ADSC-EV |
|---|---|---|
| hsa-let-7c-5p | hsa-let-7a-5p | hsa-miR-10a-5p |
| hsa-let-7d-5p | hsa-let-7e-5p | hsa-miR-1226-5p |
| hsa-let-7g-5p | hsa-miR-10b-5p | hsa-miR-125b-1-3p |
| hsa-miR-100-5p | hsa-miR-130b-3p | hsa-miR-126-5p |
| hsa-miR-106a-5p | hsa-miR-197-3p | hsa-miR-1270 |
| hsa-miR-106b-5p | hsa-miR-199a-3p | hsa-miR-1291 |
| hsa-miR-10b-3p | hsa-miR-29b-3p | hsa-miR-129-2-3p |
| hsa-miR-125a-3p | hsa-miR-342-3p | hsa-miR-136-3p |
| hsa-miR-125a-5p | hsa-miR-365a-3p | hsa-miR-137 |
| hsa-miR-125b-5p | hsa-miR-365b-3p | hsa-miR-140-5p |
| hsa-miR-126-3p | hsa-miR-483-5p | hsa-miR-142-3p |
| hsa-miR-1271-5p | hsa-miR-484 | hsa-miR-144-5p |
| hsa-miR-127-3p | hsa-miR-485-3p | hsa-miR-145-3p |
| hsa-miR-1290 | hsa-miR-99a-5p | hsa-miR-148a-3p |
| hsa-miR-130a-3p | hsa-miR-148b-5p | |
| hsa-miR-132-3p | hsa-miR-149-5p | |
| hsa-miR-133a-3p | hsa-miR-181a-2-3p | |
| hsa-miR-138-5p | hsa-miR-184 | |
| hsa-miR-139-5p | hsa-miR-18a-3p | |
| hsa-miR-143-3p | hsa-miR-18a-5p | |
| hsa-miR-145-5p | hsa-miR-193b-5p | |
| hsa-miR-146a-5p | hsa-miR-199-3p | |
| hsa-miR-146b-5p | hsa-miR-19b-1-5p | |
| hsa-miR-150-5p | hsa-miR-203a-3p | |
| hsa-miR-151a-3p | hsa-miR-204-5p | |
| hsa-miR-151a-5p | hsa-miR-210-3p | |
| hsa-miR-152-3p | hsa-miR-21-3p | |
| hsa-miR-155-5p | hsa-miR-214-5p | |
| hsa-miR-15b-5p | hsa-miR-222-5p | |
| hsa-miR-16-5p | hsa-miR-26a-1-3p | |
| hsa-miR-17-5p | hsa-miR-27a-5p | |
| hsa-miR-186-5p | hsa-miR-30a-5p | |
| hsa-miR-191-5p | hsa-miR-31-3p | |
| hsa-miR-192-5p | hsa-miR-320b | |
| hsa-miR-193a-3p | hsa-miR-335-3p | |
| hsa-miR-193a-5p | hsa-miR-339-3p | |
| hsa-miR-193b-3p | hsa-miR-34a-3p | |
| hsa-miR-195-5p | hsa-miR-362-5p | |
| hsa-miR-196b-5p | hsa-miR-374b-5p | |
| hsa-miR-19a-3p | hsa-miR-378 | |
| hsa-miR-19b-3p | hsa-miR-379-5p | |
| hsa-miR-20a-5p | hsa-miR-382-5p | |
| hsa-miR-20b-5p | hsa-miR-410-3p | |
| hsa-miR-212-3p | hsa-miR-424-3p | |
| hsa-miR-214-3p | hsa-miR-424-5p | |
| hsa-miR-21-5p | hsa-miR-425-3p | |
| hsa-miR-218-5p | hsa-miR-451a | |
| hsa-miR-221-3p | hsa-miR-452-5p | |
| hsa-miR-222-3p | hsa-miR-455-5p | |
| hsa-miR-223-3p | hsa-miR-495-3p | |
| hsa-miR-224-5p | hsa-miR-505-5p | |
| hsa-miR-24-3p | hsa-miR-542-5p | |
| hsa-miR-25-3p | hsa-miR-543 | |
| hsa-miR-26a-5p | hsa-miR-548c-3p | |
| hsa-miR-26b-5p | hsa-miR-590-3p | |
| hsa-miR-27a-3p | hsa-miR-590-5p | |
| hsa-miR-27b-3p | hsa-miR-597-5p | |
| hsa-miR-28-3p | hsa-miR-603 | |
| hsa-miR-28-5p | hsa-miR-628-3p | |
| hsa-miR-29a-3p | hsa-miR-628-5p | |
| hsa-miR-301a-3p | hsa-miR-629-3p | |
| hsa-miR-30a-3p | hsa-miR-652-3p | |
| hsa-miR-30b-5p | hsa-miR-655-3p | |
| hsa-miR-30c-5p | hsa-miR-660-5p | |
| hsa-miR-30e-3p | hsa-miR-664a-3p | |
| hsa-miR-31-5p | hsa-miR-7-1-3p | |
| hsa-miR-320a | hsa-miR-766-3p | |
| hsa-miR-323a-3p | hsa-miR-886-3p | |
| hsa-miR-324-3p | hsa-miR-889-3p | |
| hsa-miR-324-5p | hsa-miR-93-5p | |
| hsa-miR-328-3p | ||
| hsa-miR-331-3p | ||
| hsa-miR-335-5p | ||
| hsa-miR-345-5p | ||
| hsa-miR-34a-5p | ||
| hsa-miR-34b-3p | ||
| hsa-miR-370-3p | ||
| hsa-miR-374a-5p | ||
| hsa-miR-376a-3p | ||
| hsa-miR-376c-3p | ||
| hsa-miR-409-3p | ||
| hsa-miR-411-5p | ||
| hsa-miR-432-5p | ||
| hsa-miR-433-3p | ||
| hsa-miR-454-3p | ||
| hsa-miR-487b-3p | ||
| hsa-miR-493-3p | ||
| hsa-miR-494-3p | ||
| hsa-miR-532-3p | ||
| hsa-miR-532-5p | ||
| hsa-miR-539-5p | ||
| hsa-miR-574-3p | ||
| hsa-miR-625-3p | ||
| hsa-miR-708-5p | ||
| hsa-miR-744-5p | ||
| hsa-miR-886-5p | ||
| hsa-miR-92a-3p | ||
| hsa-miR-93-3p | ||
| hsa-miR-99b-3p |
| Common | BMSC-EV | ADSC-EV |
|---|---|---|
| ALDOC | A1BG | ADGRB1 |
| APOC3 | ALDOA | ALPP |
| CNTF | APOA4 | ANGPT1 |
| CNTFR | CCL2 | BMP5 |
| CSF2 | CKM | BMP7 |
| CXCL8 | CLUL1 | C2 |
| DEFB1 | CSF3 | CCHCR1 |
| ENPP2 | CXCL1 | CCL19 |
| F13A1 | CXCL9 | CCL28 |
| GDF11 | EPO | CCL4 |
| GDF3 | FN1 | CHI3L1 |
| GDF5 | IL10 | CKB |
| GDF9 | LYN | CLU |
| GH1 | MAN2B1 | CSF2RA |
| GREM1 | MINA | CXCL2 |
| GZMA | MUSK | DKK4 |
| IFNG | NPTX1 | EPX |
| IL13 | NRG3 | FGF10 |
| IL15 | PDGFRA | FGF16 |
| IL1A | POMC | FGF18 |
| IL1RL2 | RET | GDF1 |
| IL2 | S100A8 | GFRA3 |
| IL21 | TEC | HRG |
| IL5 | THBS1 | IAPP |
| IL6 | IL1R1 | |
| IL7 | IL1RL1 | |
| INSR | IL2RB | |
| KRT19 | IL9 | |
| LALBA | LAG3 | |
| MET | LHCGR | |
| MSTN | LTA | |
| S1PR1 | MMP20 | |
| SERPING1 | MSX1 | |
| SLC2A5 | MUC16 | |
| TFRC | PDGFRB | |
| TGFB1 | SCGB2A2 | |
| TNK2 | TNF | |
| TXNIP | TNFRSF13C | |
| TNFRSF8 | ||
| TNNC1 | ||
| UBB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pomatto, M.; Gai, C.; Negro, F.; Cedrino, M.; Grange, C.; Ceccotti, E.; Togliatto, G.; Collino, F.; Tapparo, M.; Figliolini, F.; et al. Differential Therapeutic Effect of Extracellular Vesicles Derived by Bone Marrow and Adipose Mesenchymal Stem Cells on Wound Healing of Diabetic Ulcers and Correlation to Their Cargoes. Int. J. Mol. Sci. 2021, 22, 3851. https://doi.org/10.3390/ijms22083851
Pomatto M, Gai C, Negro F, Cedrino M, Grange C, Ceccotti E, Togliatto G, Collino F, Tapparo M, Figliolini F, et al. Differential Therapeutic Effect of Extracellular Vesicles Derived by Bone Marrow and Adipose Mesenchymal Stem Cells on Wound Healing of Diabetic Ulcers and Correlation to Their Cargoes. International Journal of Molecular Sciences. 2021; 22(8):3851. https://doi.org/10.3390/ijms22083851
Chicago/Turabian StylePomatto, Margherita, Chiara Gai, Federica Negro, Massimo Cedrino, Cristina Grange, Elena Ceccotti, Gabriele Togliatto, Federica Collino, Marta Tapparo, Federico Figliolini, and et al. 2021. "Differential Therapeutic Effect of Extracellular Vesicles Derived by Bone Marrow and Adipose Mesenchymal Stem Cells on Wound Healing of Diabetic Ulcers and Correlation to Their Cargoes" International Journal of Molecular Sciences 22, no. 8: 3851. https://doi.org/10.3390/ijms22083851