
Berries and Their Polyphenols as a Potential Therapy for Coronary Microvascular Dysfunction: A Mini-Review

 , ,
, ,
Abstract
:1. Introduction
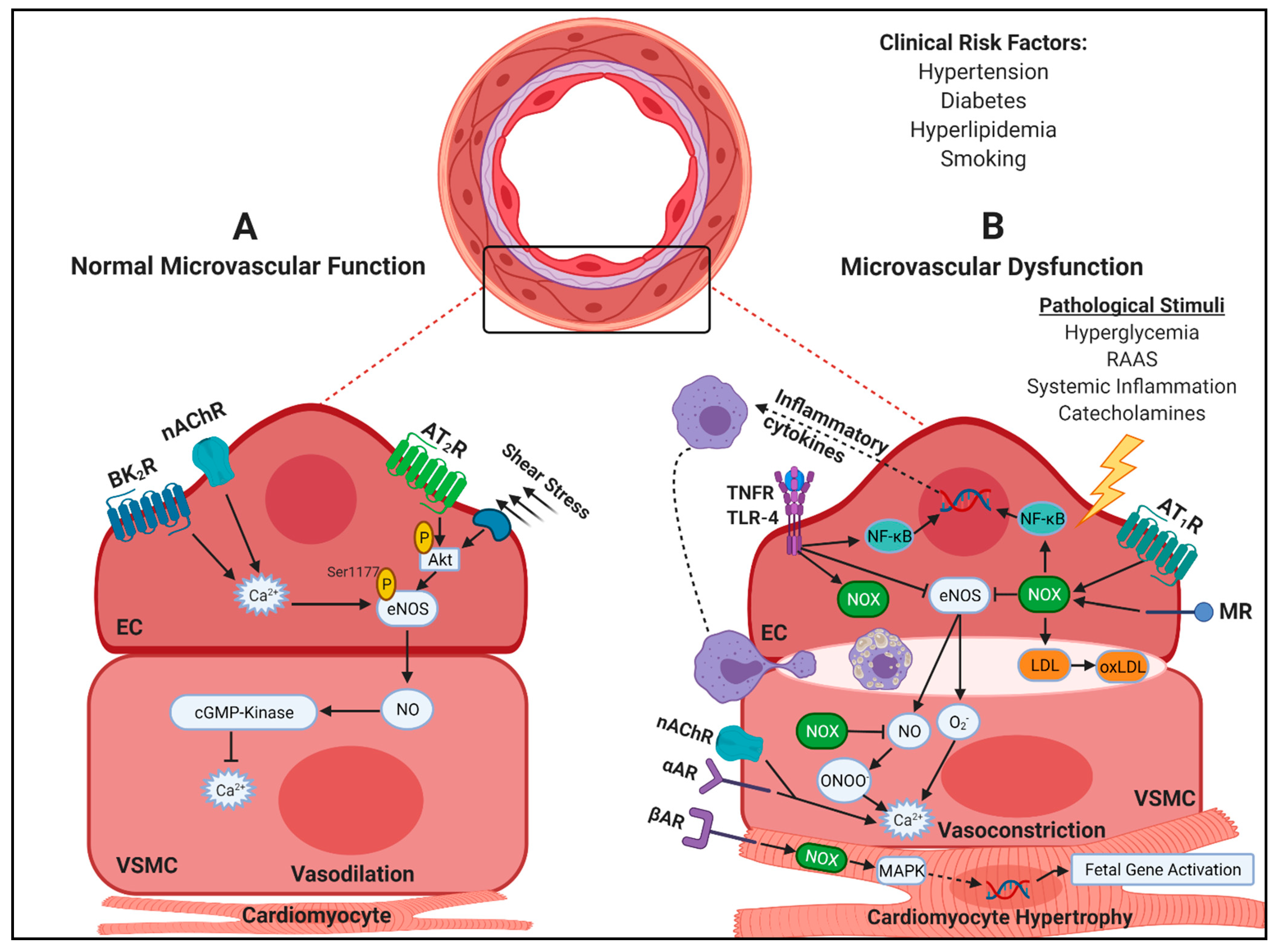
2. Coronary Microvascular Dysfunction
3. Mechanisms of CMD
3.1. Oxidative Stress and Inflammation
3.2. Endothelial-Dependent and -Independent Dysfunction
3.3. Pathological Remodeling
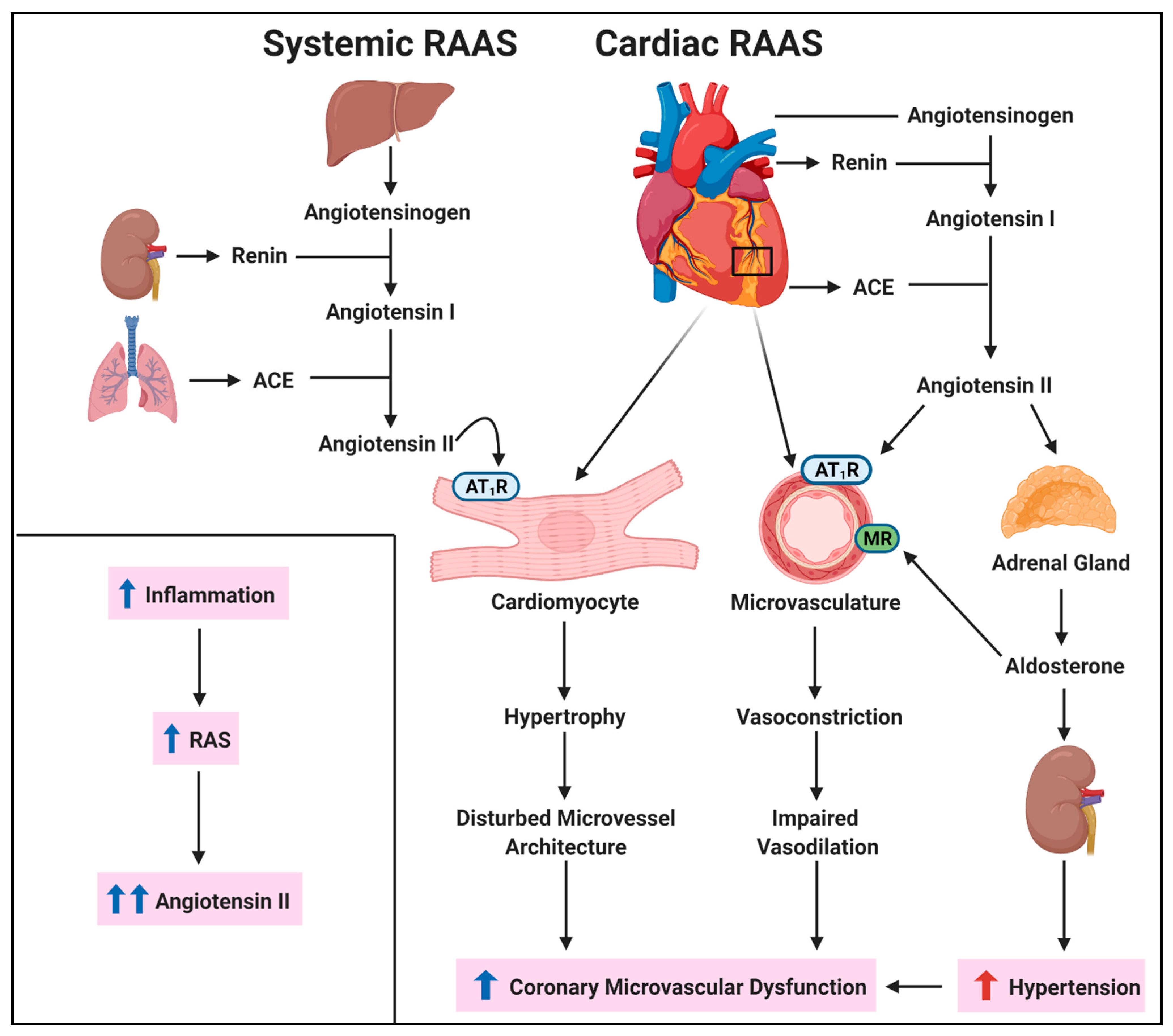
3.4. Renin-Angiotensin System
3.5. Adrenergic Receptors in CMD
4. Therapeutics in CMD
4.1. Polyphenols
4.1.1. Berry Polyphenols in Oxidative Stress and Inflammation
4.1.2. Regulation of RAAS by Berry Polyphenols
4.1.3. Berry Polyphenols as Adrenergic Receptor Inhibitors
5. Clinical Effects of Berries: Implications in CMD
5.1. Blueberries
5.2. Strawberries
5.3. Cranberries
5.4. Red and Black Raspberries
6. Limitations
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bugiardini, R.; Bairey Merz, C.N. Angina with “normal” coronary arteries: A changing philosophy. JAMA 2005, 293, 477–484. [Google Scholar] [CrossRef]
- Jespersen, L.; Hvelplund, A.; Abildstrøm, S.Z.; Pedersen, F.; Galatius, S.; Madsen, J.K.; Jørgensen, E.; Kelbaek, H.; Prescott, E. Stable angina pectoris with no obstructive coronary artery disease is associated with increased risks of major adverse cardiovascular events. Eur. Heart J. 2011, 33, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Bairey Merz, C.N.; Pepine, C.J.; Walsh, M.N.; Fleg, J.L. Ischemia and No Obstructive Coronary Artery Disease (INOCA): Developing Evidence-Based Therapies and Research Agenda for the Next Decade. Circulation 2017, 135, 1075–1092. [Google Scholar] [CrossRef]
- Gulati, M.; Cooper-DeHoff, R.M.; McClure, C.; Johnson, B.D.; Shaw, L.J.; Handberg, E.M.; Zineh, I.; Kelsey, S.F.; Arnsdorf, M.F.; Black, H.R.; et al. Adverse cardiovascular outcomes in women with nonobstructive coronary artery disease: A report from the Women’s Ischemia Syndrome Evaluation Study and the St James Women Take Heart Project. Arch. Intern. Med. 2009, 169, 843–850. [Google Scholar] [CrossRef]
- Sara, J.D.; Widmer, R.J.; Matsuzawa, Y.; Lennon, R.J.; Lerman, L.O.; Lerman, A. Prevalence of Coronary Microvascular Dysfunction Among Patients with Chest Pain and Nonobstructive Coronary Artery Disease. JACC Cardiovasc. Interv. 2015, 8, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.K.; Bess, C.; Elias-Smale, S.; Vaccarino, V.; Quyyumi, A.; Pepine, C.J.; Merz, C.N.B. Gender in cardiovascular medicine: Chest pain and coronary artery disease. Eur. Heart J. 2019, 40, 3819–3826. [Google Scholar] [CrossRef] [PubMed]
- Taqueti, V.R.; Di Carli, M.F. Coronary Microvascular Disease Pathogenic Mechanisms and Therapeutic Options: JACC State-of-the-Art Review. J. Am. Coll. Cardiol. 2018, 72, 2625–2641. [Google Scholar] [CrossRef] [PubMed]
- Ong, P.; Camici, P.G.; Beltrame, J.F.; Crea, F.; Shimokawa, H.; Sechtem, U.; Kaski, J.C.; Merz, C.N.B. International standardization of diagnostic criteria for microvascular angina. Int. J. Cardiol. 2018, 250, 16–20. [Google Scholar] [CrossRef]
- Rahman, H.; Demir, O.M.; Ryan, M.; McConkey, H.; Scannell, C.; Ellis, H.; Webb, A.; Chiribiri, A.; Perera, D. Optimal Use of Vasodilators for Diagnosis of Microvascular Angina in the Cardiac Catheterization Laboratory. Circ. Cardiovasc. Interv. 2020, 13, 009019. [Google Scholar] [CrossRef]
- Bravo, P.E.; Di Carli, M.F.; Dorbala, S. Role of PET to evaluate coronary microvascular dysfunction in non-ischemic cardiomyopathies. Heart Fail. Rev. 2017, 22, 455–464. [Google Scholar] [CrossRef]
- Schwartz, B.G.; Economides, C.; Mayeda, G.S.; Burstein, S.; Kloner, R.A. The endothelial cell in health and disease: Its function, dysfunction, measurement and therapy. Int. J. Impot. Res. 2009, 22, 77–90. [Google Scholar] [CrossRef]
- Ludmer, P.L.; Selwyn, A.P.; Shook, T.L.; Wayne, R.R.; Mudge, G.H.; Alexander, R.W.; Ganz, P. Paradoxical Vasoconstriction Induced by Acetylcholine in Atherosclerotic Coronary Arteries. N. Engl. J. Med. 1986, 315, 1046–1051. [Google Scholar] [CrossRef] [PubMed]
- Pries, A.R.; Badimon, L.; Bugiardini, R.; Camici, P.G.; Dorobantu, M.; Duncker, D.J.; Escaned, J.; Koller, A.; Piek, J.J.; De Wit, C. Coronary vascular regulation, remodelling, and collateralization: Mechanisms and clinical implications on behalf of the working group on coronary pathophysiology and microcirculation. Eur. Heart J. 2015, 36, 3134–3146. [Google Scholar] [CrossRef] [PubMed]
- Bravo, P.E.; Zimmerman, S.L.; Luo, H.-C.; Pozios, I.; Rajaram, M.; Pinheiro, A.; Steenbergen, C.; Kamel, I.R.; Wahl, R.L.; Bluemke, D.A.; et al. Relationship of Delayed Enhancement by Magnetic Resonance to Myocardial Perfusion by Positron Emission Tomography in Hypertrophic Cardiomyopathy. Circ. Cardiovasc. Imaging 2013, 6, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Cecchi, F.; Olivotto, I.; Gistri, R.; Lorenzoni, R.; Chiriatti, G.; Camici, P.G. Coronary Microvascular Dysfunction and Prognosis in Hypertrophic Cardiomyopathy. N. Engl. J. Med. 2003, 349, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Khuddus, M.A.; Pepine, C.J.; Handberg, E.M.; Bairey Merz, C.N.; Sopko, G.; Bavry, A.A.; Denardo, S.J.; McGorray, S.P.; Smith, K.M.; Sharaf, B.L.; et al. An intravascular ultrasound analysis in women experiencing chest pain in the absence of obstructive coronary artery disease: A substudy from the National Heart, Lung and Blood Institute-Sponsored Women’s Ischemia Syndrome Evaluation (WISE). J. Interv. Cardiol. 2010, 23, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Albadri, A.; Eshtehardi, P.; Hung, O.Y.; Bouchi, Y.; Khawaja, S.; Mercado, K.; Corban, M.T.; Mehta, P.K.; Shaw, L.J.; Samady, H. Coronary Microvascular Dysfunction Is Associated with Significant Plaque Burden and Diffuse Epicardial Atherosclerotic Disease. JACC Cardiovasc. Interv. 2019, 12, 1519–1520. [Google Scholar] [CrossRef]
- Yang, J.H.; Obokata, M.; Reddy, Y.N.; Redfield, M.M.; Lerman, A.; Borlaug, B.A. Endothelium-dependent and independent coronary microvascular dysfunction in patients with heart failure with preserved ejection fraction. Eur. J. Heart Fail. 2020, 22, 432–441. [Google Scholar] [CrossRef]
- Löffler, A.I.; Bourque, J.M. Coronary Microvascular Dysfunction, Microvascular Angina, and Management. Curr. Cardiol. Rep. 2016, 18, 1–7. [Google Scholar] [CrossRef]
- Taqueti, V.R.; Shaw, L.J.; Cook, N.R.; Murthy, V.L.; Shah, N.R.; Foster, C.R.; Hainer, J.; Blankstein, R.; Dorbala, S.; Di Carli, M.F. Excess Cardiovascular Risk in Women Relative to Men Referred for Coronary Angiography Is Associated with Severely Impaired Coronary Flow Reserve, Not Obstructive Disease. Circulation 2017, 135, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Hiteshi, A.K.; Li, N.; Gao, Y.; Chen, A.; Flores, F.; Mao, S.S.; Budoff, M.J. Gender Differences in Coronary Artery Diameter Are Not Related to Body Habitus or Left Ventricular Mass. Clin. Cardiol. 2014, 37, 605–609. [Google Scholar] [CrossRef]
- Murthy, V.L.; Naya, M.; Taqueti, V.R.; Foster, C.R.; Gaber, M.; Hainer, J.; Dorbala, S.; Blankstein, R.; Rimoldi, O.; Camici, P.G.; et al. Effects of Sex on Coronary Microvascular Dysfunction and Cardiac Outcomes. Circulation 2014, 129, 2518–2527. [Google Scholar] [CrossRef]
- Chatzizisis, Y.S.; Coskun, A.U.; Jonas, M.; Edelman, E.R.; Feldman, C.L.; Stone, P.H. Role of endothelial shear stress in the natural history of coronary atherosclerosis and vascular remodeling: Molecular, cellular, and vascular behavior. J. Am. Coll. Cardiol. 2007, 49, 2379–2393. [Google Scholar] [CrossRef]
- Panza, A.J.; Laurienzo, J.M.; Curiel, R.V.; Unger, E.F.; Quyyumi, A.A.; Dilsizian, V.; Cannon, R.O. Investigation of the Mechanism of Chest Pain in Patients with Angiographically Normal Coronary Arteries Using Transesophageal Dobutamine Stress Echocardiography. J. Am. Coll. Cardiol. 1997, 29, 293–301. [Google Scholar] [CrossRef]
- Quyyumi, A.A.; Cannon, R.O.; Panza, J.A.; Diodati, J.G.; Epstein, S.E. Endothelial dysfunction in patients with chest pain and normal coronary arteries. Circulation 1992, 86, 1864–1871. [Google Scholar] [CrossRef] [PubMed]
- Kip, K.E.; Marroquin, O.C.; Shaw, L.J.; Arant, C.B.; Wessel, T.R.; Olson, M.B.; Johnson, B.D.; Mulukutla, S.; Sopko, G.; Merz, C.N.; et al. Global inflammation predicts cardiovascular risk in women: A report from the Women’s Ischemia Syndrome Evaluation (WISE) study. Am. Heart J. 2005, 150, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Marroquin, O.C.; Kip, K.E.; Mulukutla, S.R.; Ridker, P.M.; Pepine, C.J.; Tjandrawan, T.; Kelsey, S.F.; Mankad, S.; Rogers, W.J.; Merz, C.N.B.; et al. Inflammation, endothelial cell activation, and coronary microvascular dysfunction in women with chest pain and no obstructive coronary artery disease. Am. Heart J. 2005, 150, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Shaw, L.J.; Bugiardini, R.; Merz, C.N. Women and ischemic heart disease: Evolving knowledge. J. Am. Coll. Cardiol. 2009, 54, 1561–1575. [Google Scholar] [CrossRef]
- Wessel, T.R.; Arant, C.B.; McGorray, S.P.; Sharaf, B.L.; Reis, S.E.; Kerensky, R.A.; von Mering, G.O.; Smith, K.M.; Pauly, D.F.; Handberg, E.M.; et al. Coronary microvascular reactivity is only partially predicted by atherosclerosis risk factors or coronary artery disease in women evaluated for suspected ischemia: Results from the NHLBI Women’s Ischemia Syndrome Evaluation (WISE). Clin. Cardiol. 2007, 30, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Daiber, A.; Chlopicki, S. Revisiting pharmacology of oxidative stress and endothelial dysfunction in cardiovascular disease: Evidence for redox-based therapies. Free Radic. Biol. Med. 2020, 157, 15–37. [Google Scholar] [CrossRef]
- Lanza, G.A.; Crea, F. Response to Letter Regarding Article, Primary Coronary Microvascular Dysfunction: Clinical Presentation, Pathophysiology, and Management. Circulation 2011, 123, 2317–2325. [Google Scholar] [CrossRef]
- Higashi, Y.; Maruhashi, T.; Noma, K.; Kihara, Y. Oxidative stress and endothelial dysfunction: Clinical evidence and therapeutic implications. Trends Cardiovasc. Med. 2014, 24, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Faccini, A.; Kaski, J.C.; Camici, P.G. Coronary microvascular dysfunction in chronic inflammatory rheumatoid diseases. Eur. Heart J. 2016, 37, 1799–1806. [Google Scholar] [CrossRef]
- Altıparmak, I.H.; Erkuş, M.E.; Sezen, H.; Demirbag, R.; Gunebakmaz, O.; Kaya, Z.; Sezen, Y.; Asoglu, R.; Dedeoglu, I.H.; Neselioglu, S.; et al. The relation of serum thiol levels and thiol/disulphide homeostasis with the severity of coronary artery disease. Kardiol. Polska 2016, 74, 1346–1353. [Google Scholar] [CrossRef]
- Bilir, B.; Akkoyun, D.C.; Aydın, M.; Ozkaramanli-Gur, D.; Degirmenci, H.; Albayrak, N.; Akyuz, A.; Alpsoy, Ş.; Koca, C.; Erel, Ö. Association of coronary artery disease severity and disulphide/native thiol ratio. Electron. J. Gen. Med. 2017, 14, 30–33. [Google Scholar] [CrossRef]
- Schroder, J.; Mygind, N.D.; Frestad, D.; Michelsen, M.; Suhrs, H.E.; Bove, K.B.; Gustafsson, I.; Kastrup, J.; Prescott, E. Pro-inflammatory biomarkers in women with non-obstructive angina pectoris and coronary microvascular dysfunction. IJC Heart Vasc. 2019, 24, 100370. [Google Scholar] [CrossRef]
- Tona, F.; Serra, R.; Di Ascenzo, L.; Osto, E.; Scarda, A.; Fabris, R.; Montisci, R.; Famoso, G.; Tellatin, S.; Foletto, M.; et al. Systemic inflammation is related to coronary microvascular dysfunction in obese patients without obstructive coronary disease. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 447–453. [Google Scholar] [CrossRef]
- Piaserico, S.; Osto, E.; Famoso, G.; Zanetti, I.; Gregori, D.; Poretto, A.; Iliceto, S.; Peserico, A.; Tona, F. Treatment with tumor necrosis factor inhibitors restores coronary microvascular function in young patients with severe psoriasis. Atherosclerosis 2016, 251, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Cosín-Sales, J.; Pizzi, C.; Brown, S.; Kaski, J.C. C-reactive protein, clinical presentation, and ischemic activity in patients with chest pain and normal coronary angiograms. J. Am. Coll. Cardiol. 2003, 41, 1468–1474. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Santillo, M.; Colantuoni, A.; Mondola, P.; Guida, B.; Damiano, S. NOX signaling in molecular cardiovascular mechanisms involved in the blood pressure homeostasis. Front. Physiol. 2015, 6, 194. [Google Scholar] [CrossRef]
- Trebak, M.; Ginnan, R.; Singer, H.A.; Jourd’Heuil, D. Interplay between Calcium and Reactive Oxygen/Nitrogen Species: An Essential Paradigm for Vascular Smooth Muscle Signaling. Antioxid. Redox. Signal. 2010, 12, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Montezano, A.C.; Touyz, R.M. Reactive Oxygen Species and Endothelial Function—Role of Nitric Oxide Synthase Uncoupling and Nox Family Nicotinamide Adenine Dinucleotide Phosphate Oxidases. Basic Clin. Pharmacol. Toxicol. 2011, 110, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Alves-Lopes, R.; Rios, F.J.; Camargo, L.L.; Anagnostopoulou, A.; Arner, A.; Montezano, A.C. Vascular smooth muscle contraction in hypertension. Cardiovasc. Res. 2018, 114, 529–539. [Google Scholar] [CrossRef]
- Zhang, H.; Park, Y.; Wu, J.; Chen, X.P.; Lee, S.; Yang, J.; Dellsperger, K.C.; Zhang, C. Role of TNF-α in vascular dysfunction. Clin. Sci. 2009, 116, 219–230. [Google Scholar] [CrossRef]
- Camici, P.G.; Olivotto, I.; Rimoldi, O.E. The coronary circulation and blood flow in left ventricular hypertrophy. J. Mol. Cell. Cardiol. 2012, 52, 857–864. [Google Scholar] [CrossRef]
- Perrotta, I.; Aquila, S. The Role of Oxidative Stress and Autophagy in Atherosclerosis. Oxidative Med. Cell. Longev. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K. Inflammation, Atherosclerosis, and Coronary Artery Disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef]
- Aviram, M.; Rosenblat, M.; Etzioni, A.; Lévy, R. Activation of NADPH oxidase is required for macrophage-mediated oxidation of low-density lipoprotein. Metabolism 1996, 45, 1069–1079. [Google Scholar] [CrossRef]
- Schaub, M.C.; Hefti, M.A.; Harder, B.A.; Eppenberger, H.M. Various hypertrophic stimuli induce distinct phenotypes in cardiomyocytes. J. Mol. Med. 1997, 75, 901–920. [Google Scholar] [CrossRef]
- Wu, C.-H.; Mohammadmoradi, S.; Chen, J.Z.; Sawada, H.; Daugherty, A.; Lu, H.S. Renin-Angiotensin System and Cardiovascular Functions. Arter. Thromb. Vasc. Biol. 2018, 38, e108–e116. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.M.; Wang, Z.-Q.; Siragy, H.M. Role of the Angiotensin Type 2 Receptor in the Regulation of Blood Pressure and Renal Function. Hypertension 2000, 35, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Freel, E.M.; Connell, J.M. Mechanisms of hypertension: The expanding role of aldosterone. J. Am. Soc. Nephrol. 2004, 15, 1993–2001. [Google Scholar] [CrossRef] [PubMed]
- Usberti, M.; Federico, S.; Di Minno, G.; Ungaro, B.; Ardillo, G.; Pecoraro, C.; Cianciaruso, B.; Cerbone, A.M.; Cirillo, F.; Pannain, M.; et al. Effects of angiotensin II on plasma ADH, prostaglandin synthesis, and water excretion in normal humans. Am. J. Physiol. Physiol. 1985, 248, F254–F259. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, H.; Suzuki, H.; Ohtsu, H.; Chao, J.Y.; Utsunomiya, H.; Frank, G.D.; Eguchi, S. Angiotensin II Regulates Vascular and Endothelial Dysfunction: Recent Topics of Angiotensin II Type-1 Receptor Signaling in the Vasculature. Curr. Vasc. Pharmacol. 2006, 4, 67–78. [Google Scholar] [CrossRef]
- Satou, R.; Penrose, H.; Navar, L.G. Inflammation as a Regulator of the Renin-Angiotensin System and Blood Pressure. Curr. Hypertens. Rep. 2018, 20, 1–9. [Google Scholar] [CrossRef]
- Hanafy, S.; Tavasoli, M.; Jamali, F. Inflammation Alters Angiotensin Converting Enzymes (ACE and ACE-2) Balance in Rat Heart. Inflammation 2010, 34, 609–613. [Google Scholar] [CrossRef]
- Pauly, D.F.; Johnson, B.D.; Anderson, R.D.; Handberg, E.M.; Smith, K.M.; Cooper-DeHoff, R.M.; Sopko, G.; Sharaf, B.M.; Kelsey, S.F.; Merz, C.N.B.; et al. In women with symptoms of cardiac ischemia, nonobstructive coronary arteries, and microvascular dysfunction, angiotensin-converting enzyme inhibition is associated with improved microvascular function: A double-blind randomized study from the National Heart, Lung and Blood Institute Women’s Ischemia Syndrome Evaluation (WISE). Am. Heart J. 2011, 162, 678–684. [Google Scholar]
- Schwartzkopff, B.; Brehm, M.; Mundhenke, M.; Strauer, B.E. Repair of Coronary Arterioles After Treatment with Perindopril in Hypertensive Heart Disease. Hypertension 2000, 36, 220–225. [Google Scholar] [CrossRef]
- Serné, E.H.; de Jongh, R.T.; Eringa, E.C.; IJzerman, R.G.; Stehouwer, C.D. Microvascular dysfunction: A potential pathophysiological role in the metabolic syndrome. Hypertension 2007, 50, 204–211. [Google Scholar] [CrossRef]
- Woo, A.Y.; Xiao, R.P. beta-Adrenergic receptor subtype signaling in heart: From bench to bedside. Acta Pharmacol. Sin. 2012, 33, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Michelotti, A.G.; Price, D.T.; Schwinn, A.D. Alpha 1-adrenergic receptor regulation: Basic science and clinical implications. Pharmacol. Ther. 2000, 88, 281–309. [Google Scholar] [CrossRef]
- Pinilla-Vera, M.; Hahn, V.S.; Kass, D.A. Leveraging Signaling Pathways to Treat Heart Failure with Reduced Ejection Fraction. Circ. Res. 2019, 124, 1618–1632. [Google Scholar] [CrossRef]
- Gericke, A.; Martinka, P.; Nazarenko, I.; Persson, P.B.; Patzak, A. Impact of α1-adrenoceptor expression on contractile properties of vascular smooth muscle cells. Am. J. Physiol. Integr. Comp. Physiol. 2007, 293, R1215–R1221. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.K.; Nelson, M.; Thomson, L.; Friedman, J.; Hayes, S.; Hermel, D.; Slomka, P.; Swift, A.; Wei, J.; Cook-Wiens, G.; et al. Abnormal cardiac sympathetic activity detected by 123-I-meta-iodobenzylguanidine imaging in women with signs and symptoms of ischemia and no obstructive coronary artery disease. J. Am. Coll. Cardiol. 2016, 67 (Suppl. 13), 1619. [Google Scholar] [CrossRef]
- Lanza, G.A.; Giordano, A.; Pristipino, C.; Calcagni, M.L.; Meduri, G.; Trani, C.; Franceschini, R.; Crea, F.; Troncone, L.; Maseri, A. Abnormal cardiac adrenergic nerve function in patients with syndrome X detected by [123I]metaiodobenzylguanidine myocardial scintigraphy. Circulation 1997, 96, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Galderisi, M.; D’Errico, A. Beta-blockers and coronary flow reserve: The importance of a vasodilatory action. Drugs 2008, 68, 579–590. [Google Scholar] [CrossRef]
- Pedersen, M.E.; Cockcroft, J.R. The vasodilatory beta-blockers. Curr. Hypertens. Rep. 2007, 9, 269–277. [Google Scholar] [CrossRef]
- Cohn, J.; Archibald, D.; Ziesche, S.; Franciosa, J.; Harston, W.; Tristani, F.; Dunkman, W.; Jacobs, W.; Francis, G.; Flohr, K.; et al. Effect of Vasodilator Therapy on Mortality in Chronic Congestive Heart Failure. Results of a Veterans Administration Cooperative Study. Surv. Anesthesiol. 1987, 31, 53. [Google Scholar] [CrossRef]
- Lymperopoulos, A.; Rengo, G.; Koch, W.J. Adrenergic nervous system in heart failure: Pathophysiology and therapy. Circ. Res. 2013, 113, 739–753. [Google Scholar] [CrossRef]
- Engelhardt, S.; Hein, L.; Wiesmann, F.; Lohse, M.J. Progressive hypertrophy and heart failure in beta1-adrenergic receptor transgenic mice. Proc. Natl. Acad. Sci. USA 1999, 96, 7059–7064. [Google Scholar] [CrossRef]
- Task Force, M.; Montalescot, G.; Sechtem, U.; Achenbach, S.; Andreotti, F.; Arden, C.; Budaj, A.; Bugiardini, R.; Crea, F.; Cuisset, T.; et al. 2013 ESC guidelines on the management of stable coronary artery disease: The Task Force on the management of stable coronary artery disease of the European Society of Cardiology. Eur. Heart J. 2013, 34, 2949–3003. [Google Scholar]
- Shaw, L.J.; Veledar, E.; Berman, D.S.; Hayes, S.W.; Friedman, J.; Slomka, P.; Germano, G.; Maron, D.J.; Mancini, G.B.J.; Hartigan, P.M.; et al. Response to Letters Regarding Article, “Optimal Medical Therapy with or Without Percutaneous Coronary Intervention to Reduce Ischemic Burden: Results from the Clinical Outcomes Utilizing Revascularization and Aggressive Drug Evaluation (COURAGE) Trial Nuclear Substudy”. Circulation 2008, 118, 1283–1291. [Google Scholar]
- Merz, C.N.B.; Pepine, C.J.; Shimokawa, H.; Berry, C. Treatment of coronary microvascular dysfunction. Cardiovasc. Res. 2020, 116, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.H.; Pedersen, L.R.; Jürs, A.; Snoer, M.; Haugaard, S.B.; Prescott, E. A randomised trial comparing the effect of exercise training and weight loss on microvascular function in coronary artery disease. Int. J. Cardiol. 2015, 185, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Najjar, R.; Turner, C.; Wong, B.; Feresin, R. Berry-Derived Polyphenols in Cardiovascular Pathologies: Mechanisms of Disease and the Role of Diet and Sex. Nutrients 2021, 13, 387. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox. Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Perezjimenez, J.; Neveu, V.; Vos, F.; Scalbert, A. Identification of the 100 richest dietary sources of polyphenols: An application of the Phenol-Explorer database. Eur. J. Clin. Nutr. 2010, 64 (Suppl. 3), S112–S120. [Google Scholar] [CrossRef]
- Olas, B. Berry Phenolic Antioxidants—Implications for Human Health? Front. Pharmacol. 2018, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Perez-Jimenez, J.; Neveu, V.; Medina-Remón, A.; M’Hiri, N.; García-Lobato, P.; Manach, C.; Knox, C.; Eisner, R.; Wishart, D.S.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef] [PubMed]
- Kimble, R.; Keane, K.M.; Lodge, J.K.; Howatson, G. Dietary intake of anthocyanins and risk of cardiovascular disease: A systematic review and meta-analysis of prospective cohort studies. Crit. Rev. Food Sci. Nutr. 2018, 59, 3032–3043. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Rabassa, M.; Cherubini, A.; Urpi-Sarda, M.; Bandinelli, S.; Ferrucci, L.; Andres-Lacueva, C. High Concentrations of a Urinary Biomarker of Polyphenol Intake Are Associated with Decreased Mortality in Older Adults. J. Nutr. 2013, 143, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Ivey, K.L.; Hodgson, J.M.; Croft, K.D.; Lewis, J.R.; Prince, R.L. Flavonoid intake and all-cause mortality. Am. J. Clin. Nutr. 2015, 101, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Piao, Z.H.; Sun, S.; Liu, B.; Kim, G.R.; Seok, Y.M.; Lin, M.Q.; Ryu, Y.; Choi, S.Y.; Kee, H.J.; et al. Gallic Acid Reduces Blood Pressure and Attenuates Oxidative Stress and Cardiac Hypertrophy in Spontaneously Hypertensive Rats. Sci. Rep. 2017, 7, 15607. [Google Scholar] [CrossRef]
- Rozentsvit, A.; Vinokur, K.; Samuel, S.; Li, Y.; Gerdes, A.M.; Carrillo-Sepulveda, M.A. Ellagic Acid Reduces High. Glucose-Induced Vascular Oxidative Stress Through ERK1/2/NOX4 Signaling Pathway. Cell Physiol. Biochem. 2017, 44, 1174–1187. [Google Scholar] [CrossRef] [PubMed]
- Vrhovsek, U.; Giongo, L.; Mattivi, F.; Viola, R. A survey of ellagitannin content in raspberry and blackberry cultivars grown in Trentino (Italy). Eur. Food Res. Technol. 2008, 226, 817–824. [Google Scholar] [CrossRef]
- Häkkinen, S.H.; Kärenlampi, S.O.; Mykkänen, H.M.; Heinonen, I.M.; Torronen, A. Ellagic acid content in berries: Influence of domestic processing and storage. Eur. Food Res. Technol. 2000, 212, 75–80. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox. Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- Truchado, P.; Larrosa, M.; García-Conesa, M.T.; Cerdá, B.; Vidal-Guevara, M.L.; Tomás-Barberán, F.A.; Espín, J.C. Strawberry Processing Does Not Affect the Production and Urinary Excretion of Urolithins, Ellagic Acid Metabolites, in Humans. J. Agric. Food Chem. 2011, 60, 5749–5754. [Google Scholar] [CrossRef]
- Istas, G.; Feliciano, R.P.; Weber, T.; Garcia-Villalba, R.; Tomas-Barberan, F.; Heiss, C.; Rodriguez-Mateos, A. Plasma urolithin metabolites correlate with improvements in endothelial function after red raspberry consumption: A double-blind randomized controlled trial. Arch. Biochem. Biophys. 2018, 651, 43–51. [Google Scholar] [CrossRef]
- Zheng, D.; Liu, Z.; Zhou, Y.; Hou, N.; Yan, W.; Qin, Y.; Ye, Q.; Cheng, X.; Xiao, Q.; Bao, Y.; et al. Urolithin B, a gut microbiota metabolite, protects against myocardial ischemia/reperfusion injury via p62/Keap1/Nrf2 signaling pathway. Pharmacol. Res. 2020, 153, 104655. [Google Scholar] [CrossRef] [PubMed]
- Häkkinen, S.H.; Kärenlampi, S.O.; Heinonen, I.M.; Mykkänen, H.M.; Törrönen, A.R. Content of the Flavonols Quercetin, Myricetin, and Kaempferol in 25 Edible Berries. J. Agric. Food Chem. 1999, 47, 2274–2279. [Google Scholar] [CrossRef]
- Liao, H.H.; Zhang, N.; Meng, Y.Y.; Feng, H.; Yang, J.J.; Li, W.J.; Chen, S.; Wu, H.M.; Deng, W.; Tang, Q.Z. Myricetin Alleviates Pathological Cardiac Hypertrophy via TRAF6/TAK1/MAPK and Nrf2 Signaling Pathway. Oxid. Med. Cell Longev. 2019, 2019, 6304058. [Google Scholar] [CrossRef]
- Zhang, C.; Lin, G.; Wan, W.; Li, X.; Zeng, B.; Yang, B.; Huang, C. Resveratrol, a polyphenol phytoalexin, protects cardiomyocytes against anoxia/reoxygenation injury via the TLR4/NF-kappaB signaling pathway. Int. J. Mol. Med. 2012, 29, 557–563. [Google Scholar] [CrossRef]
- Pantan, R.; Tocharus, J.; Suksamrarn, A.; Tocharus, C. Synergistic effect of atorvastatin and Cyanidin-3-glucoside on angiotensin II-induced inflammation in vascular smooth muscle cells. Exp. Cell Res. 2016, 342, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Sivasinprasasn, S.; Pantan, R.; Thummayot, S.; Tocharus, J.; Suksamrarn, A.; Tocharus, C. Cyanidin-3-glucoside attenuates angiotensin II-induced oxidative stress and inflammation in vascular endothelial cells. Chem. Interact. 2016, 260, 67–74. [Google Scholar] [CrossRef]
- Zhang, M.; Xue, Y.; Chen, H.; Meng, L.; Chen, B.; Gong, H.; Zhao, Y.; Qi, R. Resveratrol Inhibits MMP3 and MMP9 Expression and Secretion by Suppressing TLR4/NF-kappaB/STAT3 Activation in Ox-LDL-Treated HUVECs. Oxid. Med. Cell Longev. 2019, 2019, 9013169. [Google Scholar]
- Li, T.; Li, F.; Liu, X.; Liu, J.; Li, D. Synergistic anti-inflammatory effects of quercetin and catechin via inhibiting activation of TLR4-MyD88-mediated NF-kappaB and MAPK signaling pathways. Phytother. Res. 2019, 33, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, S.; Sudhakaran, P.R.; Helen, A. Quercetin attenuates atherosclerotic inflammation and adhesion molecule expression by modulating TLR-NF-kappaB signaling pathway. Cell Immunol. 2016, 310, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Feresin, R.G.; Huang, J.; Klarich, D.S.; Zhao, Y.; Pourafshar, S.; Arjmandi, B.H.; Salazar, G. Blackberry, raspberry and black raspberry polyphenol extracts attenuate angiotensin II-induced senescence in vascular smooth muscle cells. Food Funct. 2016, 7, 4175–4187. [Google Scholar] [CrossRef]
- Luo, X.; Fang, S.; Xiao, Y.; Song, F.; Zou, T.; Wang, M.; Xia, M.; Ling, W. Cyanidin-3-glucoside suppresses TNF-α-induced cell proliferation through the repression of Nox activator 1 in mouse vascular smooth muscle cells: Involvement of the STAT3 signaling. Mol. Cell. Biochem. 2011, 362, 211–218. [Google Scholar] [CrossRef]
- Parichatikanond, W.; Pinthong, D.; Mangmool, S. Blockade of the Renin-Angiotensin System with Delphinidin, Cyanin, and Quercetin. Planta Med. 2012, 78, 1626–1632. [Google Scholar] [CrossRef]
- Neto-Neves, E.M.; Montenegro, M.F.; Dias-Junior, C.A.; Spiller, F.; Kanashiro, A.; Tanus-Santos, J.E. Chronic Treatment with Quercetin does not Inhibit Angiotensin-Converting Enzyme In Vivo or In Vitro. Basic Clin. Pharmacol. Toxicol. 2010, 107, 825–829. [Google Scholar] [CrossRef]
- Paul, M.; Wagner, J.; Dzau, V.J. Gene expression of the renin-angiotensin system in human tissues. Quantitative analysis by the polymerase chain reaction. J. Clin. Investig. 1993, 91, 2058–2064. [Google Scholar] [CrossRef]
- Jain, S.; Li, Y.; Patil, S.; Kumar, A. HNF-1α plays an important role in IL-6-induced expression of the human angiotensinogen gene. Am. J. Physiol. Physiol. 2007, 293, C401–C410. [Google Scholar] [CrossRef]
- Brasier, A.R.; Ron, D.; Tate, J.E.; Habener, J.F. A family of constitutive C/EBP-like DNA binding proteins attenuate the IL-1 alpha induced, NF kappa B mediated trans-activation of the angiotensinogen gene acute-phase response element. EMBO J. 1990, 9, 3933–3944. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, R.; Penrose, H.; Miyata, K.; Satou, R. Macrophage-derived IL-6 contributes to ANG II-mediated angiotensinogen stimulation in renal proximal tubular cells. Am. J. Physiol. Renal Physiol. 2016, 310, F1000–F1007. [Google Scholar] [CrossRef] [PubMed]
- Flesch, M.; Höper, A.; Italia, L.D.; Evans, K.; Bond, R.; Peshock, R.; Diwan, A.; Brinsa, T.A.; Wei, C.-C.; Sivasubramanian, N.; et al. Activation and Functional Significance of the Renin-Angiotensin System in Mice with Cardiac Restricted Overexpression of Tumor Necrosis Factor. Circulation 2003, 108, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Hutabarat, R.P.; Chai, Z.; Zheng, T.; Zhang, W.; Li, D. Antioxidant Blueberry Anthocyanins Induce Vasodilation via PI3K/Akt Signaling Pathway in High-Glucose-Induced Human Umbilical Vein Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 1575. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, W.; Egan, J.M.; Slemmer, J.E.; Shaughnessy, K.S.; Ballem, K.; Gottschall-Pass, K.T.; Sweeney, M.I. Feeding blueberry diets inhibits angiotensin II-converting enzyme (ACE) activity in spontaneously hypertensive stroke-prone rats. Can. J. Physiol. Pharmacol. 2011, 89, 67–71. [Google Scholar] [CrossRef]
- Miyazaki, R.; Ichiki, T.; Hashimoto, T.; Inanaga, K.; Imayama, I.; Sadoshima, J.; Sunagawa, K. SIRT1, a Longevity Gene, Downregulates Angiotensin II Type 1 Receptor Expression in Vascular Smooth Muscle Cells. Arter. Thromb. Vasc. Biol. 2008, 28, 1263–1269. [Google Scholar] [CrossRef]
- D’Onofrio, N.; Servillo, L.; Balestrieri, M.L. SIRT1 and SIRT6 Signaling Pathways in Cardiovascular Disease Protection. Antioxid. Redox. Signal. 2018, 28, 711–732. [Google Scholar] [CrossRef]
- Hung, C.-H.; Chan, S.-H.; Chu, P.-M.; Tsai, K.-L. Quercetin is a potent anti-atherosclerotic compound by activation of SIRT1 signaling under oxLDL stimulation. Mol. Nutr. Food Res. 2015, 59, 1905–1917. [Google Scholar] [CrossRef]
- de Boer, V.C.; de Goffau, M.C.; Arts, I.C.; Hollman, P.C.; Keijer, J. SIRT1 stimulation by polyphenols is affected by their stability and metabolism. Mech. Ageing Dev. 2006, 127, 618–627. [Google Scholar] [CrossRef]
- Jin, X.; Chen, M.; Yi, L.; Chang, H.; Zhang, T.; Wang, L.; Ma, W.; Peng, X.; Zhou, Y.; Mi, M. Delphinidin-3-glucoside protects human umbilical vein endothelial cells against oxidized low-density lipoprotein-induced injury by autophagy upregulation via the AMPK/SIRT1 signaling pathway. Mol. Nutr. Food Res. 2014, 58, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, K.S.; Lassalle-Claux, G.; Touaibia, M.; Rupasinghe, H.V. Antihypertensive effect of caffeic acid and its analogs through dual renin–angiotensin–aldosterone system inhibition. Eur. J. Pharmacol. 2014, 730, 125–132. [Google Scholar] [CrossRef]
- Fujimoto, S.-I.; Niisato, N.; Sugimoto, T.; Marunaka, Y. Quercetin and NPPB-induced diminution of aldosterone action on Na+ absorption and ENaC expression in renal epithelium. Biochem. Biophys. Res. Commun. 2005, 336, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Maron, B.A.; Leopold, J.A. Mineralocorticoid receptor antagonists and endothelial function. Curr. Opin. Investig. Drugs 2008, 9, 963–969. [Google Scholar] [PubMed]
- Xiao, H.B.; Lu, X.Y.; Liu, Z.K.; Luo, Z.F. Kaempferol inhibits the production of ROS to modulate OPN-alphavbeta3 integrin pathway in HUVECs. J. Physiol. Biochem. 2016, 72, 303–313. [Google Scholar] [CrossRef]
- Liu, Z.-K.; Xiao, H.-B.; Fang, J. Anti-inflammatory properties of kaempferol via its inhibition of aldosterone signaling and aldosterone-induced gene expression. Can. J. Physiol. Pharmacol. 2014, 92, 117–123. [Google Scholar] [CrossRef]
- Norton, C.; Kalea, A.Z.; Harris, P.D.; Klimis-Zacas, D.J. Wild Blueberry-Rich Diets Affect the Contractile Machinery of the Vascular Smooth Muscle in the Sprague–Dawley Rat. J. Med. Food 2005, 8, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Kristo, A.S.; Kalea, A.Z.; Schuschke, D.A.; Klimis-Zacas, D.J. A Wild Blueberry-Enriched Diet (Vaccinium angustifolium) Improves Vascular Tone in the Adult Spontaneously Hypertensive Rat. J. Agric. Food Chem. 2010, 58, 11600–11605. [Google Scholar] [CrossRef]
- Rodriguez-Mateos, A.; Ishisaka, A.; Mawatari, K.; Vidal-Diez, A.; Spencer, J.P.E.; Terao, J. Blueberry intervention improves vascular reactivity and lowers blood pressure in high-fat-, high-cholesterol-fed rats. Br. J. Nutr. 2013, 109, 1746–1754. [Google Scholar] [CrossRef] [PubMed]
- Vendrame, S.; Kristo, A.; Schuschke, D.A.; Klimis-Zacas, D. Wild blueberry consumption affects aortic vascular function in the obese Zucker rat. Appl. Physiol. Nutr. Metab. 2014, 39, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Louis, X.L.; Thandapilly, S.J.; Kalt, W.; Vinqvist-Tymchuk, M.; Aloud, B.M.; Raj, P.; Yu, L.; Le, H.; Netticadan, T. Blueberry polyphenols prevent cardiomyocyte death by preventing calpain activation and oxidative stress. Food Funct. 2014, 5, 1785–1794. [Google Scholar] [CrossRef]
- Wei, H.; Li, H.; Wan, S.-P.; Zeng, Q.-T.; Cheng, L.-X.; Jiang, L.-L.; Peng, Y.-D. Cardioprotective Effects of Malvidin Against Isoproterenol-Induced Myocardial Infarction in Rats: A Mechanistic Study. Med. Sci. Monit. 2017, 23, 2007–2016. [Google Scholar] [CrossRef]
- Ryu, Y.; Jin, L.; Kee, H.J.; Piao, Z.H.; Cho, J.Y.; Kim, G.R.; Choi, S.Y.; Lin, M.Q.; Jeong, M.H. Gallic acid prevents isoproterenol-induced cardiac hypertrophy and fibrosis through regulation of JNK2 signaling and Smad3 binding activity. Sci. Rep. 2016, 6, 34790. [Google Scholar] [CrossRef]
- De Lacerda Alexandre, J.V.D.L.; Viana, Y.I.P.; David, C.E.B.; Cunha, P.L.O.; Albuquerque, A.C.; Varela, A.L.N.; Kowaltowski, A.J.; Facundo, H.T. Quercetin treatment increases H2O2 removal by restoration of endogenous antioxidant activity and blocks isoproterenol-induced cardiac hypertrophy. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2021, 394, 217–226. [Google Scholar] [CrossRef]
- Manjunatha, S.; Shaik, A.H.; E., M.P.; Al Omar, S.Y.; Mohammad, A.; Kodidhela, L.D. Combined cardio-protective ability of syringic acid and resveratrol against isoproterenol induced cardio-toxicity in rats via attenuating NF-kB and TNF-alpha pathways. Sci. Rep. 2020, 10, 3426. [Google Scholar]
- Van Der Vaart, H.; Postma, D.S.; Timens, W.; Hacken, N.H.T.T. Acute effects of cigarette smoke on inflammation and oxidative stress: A review. Thorax 2004, 59, 713–721. [Google Scholar] [CrossRef]
- Gullu, H.; Caliskan, M.; Ciftci, O.; Erdogan, D.; Topcu, S.; Yildirim, E.; Yildirir, A.; Muderrisoglu, H. Light cigarette smoking impairs coronary microvascular functions as severely as smoking regular cigarettes. Heart 2007, 93, 1274–1277. [Google Scholar] [CrossRef]
- Del Bo’, C.; Porrini, M.; Fracassetti, D.; Campolo, J.; Klimis-Zacas, D.; Riso, P. A single serving of blueberry (V. corymbosum) modulates peripheral arterial dysfunction induced by acute cigarette smoking in young volunteers: A randomized-controlled trial. Food Funct. 2014, 5, 3107–3116. [Google Scholar] [CrossRef]
- Riso, P.; Klimis-Zacas, D.; Del Bo’, C.; Martini, D.; Campolo, J.; Vendrame, S.; Møller, P.; Loft, S.; De Maria, R.; Porrini, M. Effect of a wild blueberry (Vaccinium angustifolium) drink intervention on markers of oxidative stress, inflammation and endothelial function in humans with cardiovascular risk factors. Eur. J. Nutr. 2012, 52, 949–961. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Mateos, A.M.R.; Rendeiro, C.; Bergillos-Meca, T.; Tabatabaee, S.; George, T.W.; Heiss, C.; Spencer, J.P. Intake and time dependence of blueberry flavonoid–induced improvements in vascular function: A randomized, controlled, double-blind, crossover intervention study with mechanistic insights into biological activity. Am. J. Clin. Nutr. 2013, 98, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Del Bo’, C.; Porrini, M.; Campolo, J.; Parolini, M.; Lanti, C.; Klimis-Zacas, D.; Riso, P. A single blueberry (Vaccinium corymbosum) portion does not affect markers of antioxidant defence and oxidative stress in healthy volunteers following cigarette smoking. Mutagenesis 2016, 31, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Yaginuma, K.; Hashimoto, S.; Toki, T.; Nakamura, I.; Fukuchi, S. Acute effect of cigarette smoking on serum angiotensin-converting enzyme activity in normal man. Tohoku J. Exp. Med. 1982, 137, 113–114. [Google Scholar] [CrossRef]
- von Scholten, B.J.; Hasbak, P.; Christensen, T.E.; Ghotbi, A.A.; Kjaer, A.; Rossing, P.; Hansen, T.W. Cardiac (82)Rb PET/CT for fast and non-invasive assessment of microvascular function and structure in asymptomatic patients with type 2 diabetes. Diabetologia 2016, 59, 371–378. [Google Scholar] [CrossRef]
- Picchi, A.; Capobianco, S.; Qiu, T.; Focardi, M.; Zou, X.; Cao, J.-M.; Zhang, C. Coronary microvascular dysfunction in diabetes mellitus: A review. World J. Cardiol. 2010, 2, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Curtis, P.J.; Van Der Velpen, V.; Berends, L.; Jennings, A.; Feelisch, M.; Umpleby, A.M.; Evans, M.; Fernandez, B.O.; Meiss, M.S.; Minnion, M.; et al. Blueberries improve biomarkers of cardiometabolic function in participants with metabolic syndrome—results from a 6-month, double-blind, randomized controlled trial. Am. J. Clin. Nutr. 2019, 109, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Stull, A.J.; Cash, K.C.; Champagne, C.M.; Gupta, A.K.; Boston, R.; Beyl, R.A.; Johnson, W.D.; Cefalu, W.T. Blueberries Improve Endothelial Function, but Not Blood Pressure, in Adults with Metabolic Syndrome: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients 2015, 7, 4107–4123. [Google Scholar] [CrossRef]
- Basu, A.; Du, M.; Leyva, M.J.; Sanchez, K.; Betts, N.M.; Wu, M.; Aston, C.E.; Lyons, T.J. Blueberries Decrease Cardiovascular Risk Factors in Obese Men and Women with Metabolic Syndrome. J. Nutr. 2010, 140, 1582–1587. [Google Scholar] [CrossRef]
- Nair, A.R.; Mariappan, N.; Stull, A.J.; Francis, J. Blueberry supplementation attenuates oxidative stress within monocytes and modulates immune cell levels in adults with metabolic syndrome: A randomized, double-blind, placebo-controlled trial. Food Funct. 2017, 8, 4118–4128. [Google Scholar] [CrossRef]
- Stote, K.S.; Sweeney, M.I.; Kean, T.; Baer, D.J.; Novotny, J.A.; Shakerley, N.L.; Chandrasekaran, A.; Carrico, P.M.; Melendez, J.A.; Gottschall-Pass, K.T. The effects of 100% wild blueberry (Vaccinium angustifolium) juice consumption on cardiometablic biomarkers: A randomized, placebo-controlled, crossover trial in adults with increased risk for type 2 diabetes. BMC Nutr. 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Johnson, S.A.; Figueroa, A.; Navaei, N.; Wong, A.; Kalfon, R.; Ormsbee, L.T.; Feresin, R.G.; Elam, M.L.; Hooshmand, S.; Payton, M.E.; et al. Daily Blueberry Consumption Improves Blood Pressure and Arterial Stiffness in Postmenopausal Women with Pre- and Stage 1-Hypertension: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J. Acad. Nutr. Diet. 2015, 115, 369–377. [Google Scholar] [CrossRef]
- Burton-Freeman, B.; Linares, A.; Hyson, D.; Kappagoda, T. Strawberry Modulates LDL Oxidation and Postprandial Lipemia in Response to High-Fat Meal in Overweight Hyperlipidemic Men and Women. J. Am. Coll. Nutr. 2010, 29, 46–54. [Google Scholar] [CrossRef]
- Basu, A.; Betts, N.M.; Nguyen, A.; Newman, E.D.; Fu, D.; Lyons, T.J. Freeze-Dried Strawberries Lower Serum Cholesterol and Lipid Peroxidation in Adults with Abdominal Adiposity and Elevated Serum Lipids. J. Nutr. 2014, 144, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Djurica, D.; Holt, R.R.; Ren, J.; Shindel, A.W.; Hackman, R.M.; Keen, C.L. Effects of a dietary strawberry powder on parameters of vascular health in adolescent males. Br. J. Nutr. 2016, 116, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Feresin, R.G.; Johnson, S.A.; Pourafshar, S.; Campbell, J.C.; Jaime, S.J.; Navaei, N.; Elam, M.L.; Akhavan, N.S.; Alvarez-Alvarado, S.; Tenenbaum, G.; et al. Impact of daily strawberry consumption on blood pressure and arterial stiffness in pre- and stage 1-hypertensive postmenopausal women: A randomized controlled trial. Food Funct. 2017, 8, 4139–4149. [Google Scholar] [CrossRef]
- Rodriguez-Mateos, A.; Feliciano, R.P.; Boeres, A.; Weber, T.; Dos Santos, C.N.; Ventura, M.R.; Heiss, C. Cranberry (poly)phenol metabolites correlate with improvements in vascular function: A double-blind, randomized, controlled, dose-response, crossover study. Mol. Nutr. Food Res. 2016, 60, 2130–2140. [Google Scholar] [CrossRef] [PubMed]
- Dohadwala, M.M.; Holbrook, M.; Hamburg, N.M.; Shenouda, S.M.; Chung, W.B.; Titas, M.; Kluge, M.A.; Wang, N.; Palmisano, J.; Milbury, P.E.; et al. Effects of cranberry juice consumption on vascular function in patients with coronary artery disease. Am. J. Clin. Nutr. 2011, 93, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Ruel, G.; Lapointe, A.; Pomerleau, S.; Couture, P.; Lemieux, S.; Lamarche, B.; Couillard, C. Evidence that cranberry juice may improve augmentation index in overweight men. Nutr. Res. 2013, 33, 41–49. [Google Scholar] [CrossRef]
- Basu, A.; Betts, N.M.; Ortiz, J.; Simmons, B.; Wu, M.; Lyons, T.J. Low-energy cranberry juice decreases lipid oxidation and increases plasma antioxidant capacity in women with metabolic syndrome. Nutr. Res. 2011, 31, 190–196. [Google Scholar] [CrossRef]
- Novotny, J.A.; Baer, D.J.; Khoo, C.; Gebauer, S.K.; Charron, C.S. Cranberry Juice Consumption Lowers Markers of Cardiometabolic Risk, Including Blood Pressure and Circulating C-Reactive Protein, Triglyceride, and Glucose Concentrations in Adults. J. Nutr. 2015, 145, 1185–1193. [Google Scholar] [CrossRef]
- Ruel, G.; Pomerleau, S.; Couture, P.; Lemieux, S.; Lamarche, B.; Couillard, C. Low-calorie cranberry juice supplementation reduces plasma oxidized LDL and cell adhesion molecule concentrations in men. Br. J. Nutr. 2008, 99, 352–359. [Google Scholar] [CrossRef]
- Schell, J.; Betts, N.M.; Lyons, T.J.; Basu, A. Raspberries Improve Postprandial Glucose and Acute and Chronic Inflammation in Adults with Type 2 Diabetes. Ann. Nutr. Metab. 2019, 74, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Franck, M.; De Toro-Martín, J.; Garneau, V.; Guay, V.; Kearney, M.; Pilon, G.; Roy, D.; Couture, P.; Couillard, C.; Marette, A.; et al. Effects of Daily Raspberry Consumption on Immune-Metabolic Health in Subjects at Risk of Metabolic Syndrome: A Randomized Controlled Trial. Nutrients 2020, 12, 3858. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.S.; Kim, S.; Hong, S.J.; Choi, S.C.; Choi, J.-H.; Kim, J.-H.; Park, C.-Y.; Cho, J.Y.; Lee, T.-B.; Kwon, J.-W.; et al. Black Raspberry Extract Increased Circulating Endothelial Progenitor Cells and Improved Arterial Stiffness in Patients with Metabolic Syndrome: A Randomized Controlled Trial. J. Med. Food 2016, 19, 346–352. [Google Scholar] [CrossRef]
- Jeong, H.S.; Hong, S.J.; Lee, T.-B.; Kwon, J.-W.; Jeong, J.T.; Joo, H.J.; Park, J.H.; Ahn, C.-M.; Yu, C.W.; Lim, D.-S. Effects of Black Raspberry on Lipid Profiles and Vascular Endothelial Function in Patients with Metabolic Syndrome. Phytother. Res. 2014, 28, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.S.; Hong, S.J.; Cho, J.Y.; Lee, T.-B.; Kwon, J.-W.; Joo, H.J.; Park, J.H.; Yu, C.W.; Lim, D.-S. Effects of Rubus occidentalis extract on blood pressure in patients with prehypertension: Randomized, double-blinded, placebo-controlled clinical trial. Nutrients 2016, 32, 461–467. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cellular Target | Berry/Polyphenol(s) | Cellular Effect and Physiological Consequence |
|---|---|---|
| αAR ↓ [121,122,123,124] | Blueberries | Decreased phenylephrine-induced αAR signaling in the vasculature with reduced VSMC-mediated vasoconstriction. Thus, epicardial microvascular blood flow can potentially be improved in the presence of classical αAR agonists, epinephrine, and norepinephrine. |
| βAR ↓ [125,126,127,128,129] | Blueberries, malvidin, gallic acid, quercetin, resveratrol and syringic acid. | Decreased isoproterenol-, epinephrine- and norepinephrine-induced βAR signaling in the heart, reducing LVH, aberrant Ca2+ handling, and cardiac ROS. These cumulative effects preserve myocardial architecture, thus, maintaining adequate microvascular flow. |
| ACE ↓ [84,109,110] | Gallic acid and blueberries | Decreased ACE expression throughout the cardiovascular system, reducing the cleavage of Ang I to Ang II and reducing hypertension. ACE inhibitors are classically used in CMD treatment; thus, berry polyphenols may target ACE in a pharmacological fashion. |
| Aldosterone ↓ [116,117,119,120] | Caffeic acid, quercetin and kaempferol | May decrease the synthesis of aldosterone in the adrenal cortex and decrease the action of aldosterone in the kidneys, resulting in reduced blood pressure, decreasing microcirculatory resistance. Additionally, polyphenols may decrease MR activity in the endothelium, resulting in reduced endothelial dysfunction. |
| AT1R ↓ [84,111] | Gallic acid and resveratrol | Decreased AT1R expression in the heart and endothelium, reducing downstream AT1R signaling, potentially preserving endothelial function, and reducing LVH. Angiotensin receptor blockers are common medications prescribed to patients with CMD, thus, polyphenols may act in a similar pharmacological fashion. |
| NF-κB ↓ [93,94,95,96,97,98,99] | Myricetin, resveratrol, cyandin-3-glucoside, quercetin and catechin | Decreased NF-κB phosphorylation and nuclear translocation leading to a decrease in inflammatory cytokine expression in ECs, VSMCs, and cardiomyocytes. In the endothelium, this leads to a decrease in leukocyte infiltration, decreasing LDL phagocytosis and diffuse atherosclerosis in the sub endothelial space. |
| MAPK ↓ [85,93,98,100] | Blackberry, raspberry and black raspberry polyphenol extracts, ellagic acid, myricetin, quercetin and catechin | Polyphenols decrease cardiomyocyte MAPK signaling, reducing fetal gene activation and a subsequent attenuation of cardiomyocyte hypertrophic growth. Decreased myocardial hypertrophy prevents an impediment of lumenal space of arteriole and pre-arterioles of epicardial microvasculature. |
| NOX ↓ [84,85,100,101] | Blackberry, raspberry and black raspberry polyphenol extracts, gallic acid, cyanidin-3-glucoside | Decreased NOX protein expression in ECs, VSMCs, and cardiomyocytes, which reduces ROS, thereby reducing redox sensitive kinases upstream from MAPK and NF-κB. In ECs, reduced ROS from NOX prevents eNOS uncoupling and increases NO bioavailability. In VSMCs, reduced NOX-derived ROS prevent aberrant intracellular Ca2+ fluctuations, thus, endothelial-independent and -dependent dysfunction is attenuated. |
| NRF2 ↑ [91,93,95,96] | Urolithin, B, myricetin and cyanidin-3-glucoside | Increased nuclear translocation of NRF2, increasing transcription of antioxidant enzymes, leading to the neutralization of ROS. Thus, upregulated NRF2 can lead to quenching of excessive ROS produced from NOX and other potential ROS sources. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Najjar, R.S.; Schwartz, A.M.; Wong, B.J.; Mehta, P.K.; Feresin, R.G. Berries and Their Polyphenols as a Potential Therapy for Coronary Microvascular Dysfunction: A Mini-Review. Int. J. Mol. Sci. 2021, 22, 3373. https://doi.org/10.3390/ijms22073373
Najjar RS, Schwartz AM, Wong BJ, Mehta PK, Feresin RG. Berries and Their Polyphenols as a Potential Therapy for Coronary Microvascular Dysfunction: A Mini-Review. International Journal of Molecular Sciences. 2021; 22(7):3373. https://doi.org/10.3390/ijms22073373
Chicago/Turabian StyleNajjar, Rami S., Arielle M. Schwartz, Brett J. Wong, Puja K. Mehta, and Rafaela G. Feresin. 2021. "Berries and Their Polyphenols as a Potential Therapy for Coronary Microvascular Dysfunction: A Mini-Review" International Journal of Molecular Sciences 22, no. 7: 3373. https://doi.org/10.3390/ijms22073373