
Revealing the Role of High-Density Lipoprotein in Colorectal Cancer

 ,
,
Abstract
:1. Introduction
2. HDL-C Levels and HDL Subclass Distribution in CRC
3. Functional Characteristics of HDL Particles in CRC
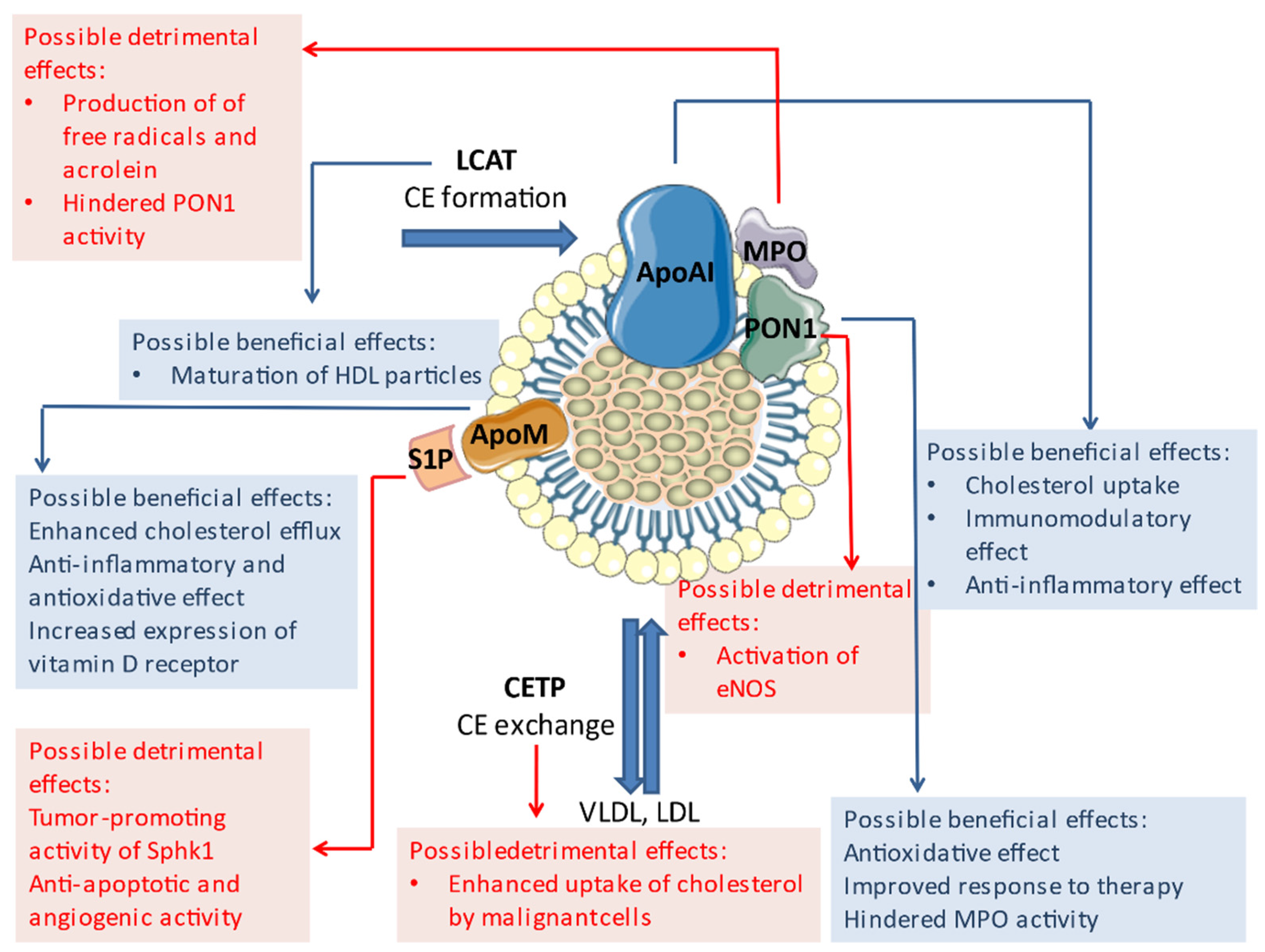
3.1. ApoAI
3.2. ApoM
3.3. PON1
3.4. LCAT
3.5. CETP
4. Genetic Regulation of HDL Metabolism in CRC
4.1. ABC Transporters
4.2. Scavenger Receptor Class B, Type 1

{kind=link}
{kind=link}
| Sources | Gene Expression/SNP Findings | Possible Clinical Significance |
|---|---|---|
| Hlavata et al. [133] | Decreased ATP-binding cassette transporter A1 (ABCA1) and ABCG1 mRNA levels in CRC tumor tissue No differences in ABCG5 and ABCG8 transcript levels between tumor and control tissues Decrease in ABCG5 and ABCG8 transcript levels from colon to rectum | Assumed prognostic potential of ABC transporters |
| Aguirre-Portolés et al. [65] | Increased ABCA1 gene expression in advanced stages of CRC | Marker of CRC invasiveness and overall survival |
| Vargas et al. [135] Fernández et al. [136] | Increased ABCA1 gene expression in a prognostic fingerprint (ColoLipidGene signature), primarily used for stage II of CRC and later extended to all stages of CRC Non-synonymous ABCA1 polymorphism (rs2230808) association with ABCA1 mRNA levels | Prognostic marker (emphasized as the main carrier of ColoLipidGene signature potential) |
| Namba et al. [137] | Increased ABCG1 gene expression in aggregative metastatic colon cancer cells Decreased viability of tumoroids caused by ABCG1 depletion | Possible therapeutic target Prognostic marker for patients with CRC |
| Po et al. [132] | Increased ABCG1 gene expression in CRC tissue Increased ABCG1 expression related to worse disease prognosis | Prognostic biomarker |
| Bi et al. [139] | MicroRNA (miR)-183 downregulating ABCA1 gene and protein expression in colon cancer cells Silencing of ABCA1 gene increasing proliferating capacity and inhibiting tumor cells’ apoptosis | ABCA1 singled out as a tumor-suppressive gene in colon cancer miR-183 named as a possible predictive biomarker of advanced aggressive forms of CRC |
| Hostettler et al. [145] | Poor survival time obtained in patients with ABCG5-positive tumor buds | Prognostic biomarker |
| Shahzad et al. [150] | Increased expression of scavenger receptor class B, type 1 (SR-B1) in breast, colorectal, pancreatic, and ovarian cancer cell lines | SR-B1 representing potential therapeutic targets for delivery of small interfering RNA (siRNA)-based therapeutic HDL’s payloads |
| Sharma et al. [151] | Inconsistent results regarding low-density lipoprotein receptor (LDLR), SCARB1, and ABCA1 mRNA levels between experimental and in silico analysis of colorectal tissue and adjacent/normal mucosa TCGA cohort: increased LDLR and SCARB1 expression but downregulated ABCA1 | Evaluation of diagnostic and prognostic potential of biomarkers included in total cholesterol homeostasis |
4.3. LCAT and CETP
5. HDL and Cholesterol Trafficking in CRC
6. Possible Therapeutic Implications
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, K. Colorectal cancer development and advances in screening. Clin. Interv. Aging 2016, 11, 967–976. [Google Scholar] [PubMed] [Green Version]
- Jochem, C.; Leitzmann, M. Obesity and Colorectal Cancer. Recent Results Cancer Res. 2016, 208, 17–41. [Google Scholar] [PubMed]
- Ganjali, S.; Banach, M.; Pirro, M.; Fras, Z.; Sahebkar, A. HDL and cancer-causality still needs to be confirmed? Update 2020. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- Schaefer, E.J.; Anthanont, P.; Asztalos, B.F. High-density lipoprotein metabolism, composition, function, and deficiency. Curr. Opin. Lipidol. 2014, 25, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Ertek, S. High-density Lipoprotein (HDL) Dysfunction and the Future of HDL. Curr. Vasc. Pharmacol. 2018, 16, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, S.T.; Charakida, M. High-Density Lipoprotein Function and Dysfunction in Health and Disease. Cardiovasc. Drugs Ther. 2019, 33, 207–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bains, Y.; Caccavello, R.; Kotani, K.; Gugliucci, A. Paraoxonase 1, HDL subclasses and post surgery acute inflammation: A pilot study. Antioxidants 2019, 8, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linton, M.R.F.; Yancey, P.G.; Davies, S.S.; Jerome, W.G.; Linton, E.F.; Song, W.L.; Doran, A.C.; Vickers, K.C. The Role of Lipids and Lipoproteins in Atherosclerosis. Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dungan, K., Grossman, A., Hershman, J.M., Hofland, J., Kaltsas, G., et al., Eds.; South Dartmouth, MA, USA, 2000. Available online: https://www.endotext.org/ (accessed on 15 March 2021).
- Wang, N.; Lan, D.; Chen, W.; Matsuura, F.; Tall, A.R. ATP-binding cassette transporters G1 and G4 mediate cellular cholesterol efflux to high-density lipoproteins. Proc. Natl. Acad. Sci. USA 2004, 101, 9774–9779. [Google Scholar] [CrossRef] [Green Version]
- Adorni, M.P.; Zimetti, F.; Billheimer, J.T.; Wang, N.; Rader, D.J.; Phillips, M.C.; Rothblat, G.H. The roles of different pathways in the release of cholesterol from macrophages. J. Lipid Res. 2007, 48, 2453–2462. [Google Scholar] [CrossRef] [Green Version]
- Riwanto, M.; Rohrer, L.; von Eckardstein, A.; Landmesser, U. Dysfunctional HDL: From structure-function-relationships to biomarkers. Handb. Exp. Pharmacol. 2015, 224, 337–366. [Google Scholar] [PubMed] [Green Version]
- Lee, M.K.; Moore, X.L.; Fu, Y.; Al-Sharea, A.; Dragoljevic, D.; Fernandez-Rojo, M.A.; Parton, R.; Sviridov, D.; Murphy, A.J.; Chin-Dusting, J.P. High-density lipoprotein inhibits human M1 macrophage polarization through redistribution of caveolin-1. Br. J. Pharmacol. 2016, 173, 741–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabet, F.; Vickers, K.C.; Cuesta Torres, L.F.; Wiese, C.B.; Shoucri, B.M.; Lambert, G.; Catherinet, C.; Prado-Lourenco, L.; Levin, M.G.; Thacker, S.; et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat. Commun. 2014, 5, 3292. [Google Scholar] [CrossRef] [Green Version]
- Ganjali, S.; Ricciuti, B.; Pirro, M.; Butler, A.E.; Atkin, S.L.; Banach, M.; Sahebkar, A. High-Density Lipoprotein Components and Functionality in Cancer: State-of-the-Art. Trends Endocrinol. Metab. 2019, 30, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Jafri, H.; Alsheikh-Ali, A.A.; Karas, R.H. Baseline and on-treatment high-density lipoprotein cholesterol and the risk of cancer in randomized controlled trials of lipid-altering therapy. J. Am. Coll. Cardiol. 2010, 55, 2846–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Duijnhoven, F.J.; Bueno-De-Mesquita, H.B.; Calligaro, M.; Jenab, M.; Pischon, T.; Jansen, E.H.; Frohlich, J.; Ayyobi, A.; Overvad, K.; Toft-Petersen, A.P.; et al. Blood lipid and lipoprotein concentrations and colorectal cancer risk in the European Prospective Investigation into Cancer and Nutrition. Gut 2011, 60, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Tian, Z. Dyslipidemia and colorectal cancer risk: A meta-analysis of prospective studies. Cancer Causes Control 2015, 26, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, X.W.; Liu, D.B.; Han, C.Z.; Du, L.L.; Jing, J.X.; Wang, Y. Lipid levels in serum and cancerous tissues of colorectal cancer patients. World J. Gastroenterol. 2014, 20, 8646–8652. [Google Scholar] [CrossRef] [PubMed]
- Dessì, S.; Batetta, B.; Pulisci, D.; Spano, O.; Anchisi, C.; Tessitore, L.; Costelli, P.; Baccino, F.M.; Aroasio, E.; Pani, P. Cholesterol content in tumor tissues is inversely associated with high-density lipoprotein cholesterol in serum in patients with gastrointestinal cancer. Cancer 1994, 73, 253–258. [Google Scholar] [CrossRef]
- Healy, L.A.; Howard, J.M.; Ryan, A.M.; Beddy, P.; Mehigan, B.; Stephens, R.; Reynolds, J.V. Metabolic syndrome and leptin are associated with adverse pathological features in male colorectal cancer patients. Colorectal. Dis. 2012, 14, 157–165. [Google Scholar] [CrossRef]
- Notarnicola, M.; Altomare, D.F.; Correale, M.; Ruggieri, E.; D’Attoma, B.; Mastrosimini, A.; Guerra, V.; Caruso, M.G. Serum lipid profile in colorectal cancer patients with and without synchronous distant metastases. Oncology 2005, 68, 371–374. [Google Scholar] [CrossRef]
- Zhu, P.W.; Gong, Y.X.; Min, Y.L.; Lin, Q.; Li, B.; Shi, W.Q.; Yuan, Q.; Ye, L.; Shao, Y. The predictive value of high-density lipoprotein for ocular metastases in colorectal cancer patients. Cancer Manag. Res. 2019, 11, 3511–3519. [Google Scholar] [CrossRef] [PubMed]
- Melichar, B.; Kalábová, H.; Krcmová, L.; Kasparová, M.; Malírová, E.; Melicharová, K.; Pecka, M.; Hyspler, R.; Solichová, D. Serum homocysteine, cholesterol, retinol, alpha-tocopherol, glycosylated hemoglobin and inflammatory response during therapy with bevacizumab, oxaliplatin, 5-fluorouracil and leucovorin. Anticancer Res. 2009, 29, 4813–4820. [Google Scholar] [PubMed]
- Wang, Y.; Wang, Z.Q.; Wang, F.H.; Lei, X.F.; Yan, S.M.; Wang, D.S.; Zhang, F.; Xu, R.H.; Wang, L.Y.; Li, Y.H. Predictive value of chemotherapy-related high-density lipoprotein cholesterol (HDL) elevation in patients with colorectal cancer receiving adjuvant chemotherapy: An exploratory analysis of 851 cases. Oncotarget 2016, 7, 57290–57300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, C.; Zhang, J.; Liu, Y.; Gong, G.; Mo, X.; Liu, P.; Li, B.; Yin, Y. Predictive value of blood lipid association with response to neoadjuvant chemoradiotherapy in colorectal cancer. Tumour Biol. 2016, 37, 4955–4961. [Google Scholar] [CrossRef]
- Benson, A.B.; Venook, A.P.; Al-Hawary, M.M.; Arain, M.A.; Chen, Y.J.; Ciombor, K.K.; Cohen, S.A.; Cooper, H.S.; Deming, D.A.; Garrido-Laguna, I.; et al. Small Bowel Adenocarcinoma, Version 1.2020, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2019, 17, 1109–1133. [Google Scholar] [CrossRef] [PubMed]
- Tabuso, M.; Christian, M.; Kimani, P.K.; Gopalakrishnan, K.; Arasaradnam, R.P. KRAS Status is Associated with Metabolic Parameters in Metastatic Colorectal Cancer According to Primary Tumour Location. Pathol. Oncol. Res. 2020, 26, 2537–2548. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, X.Q.; Lin, H.C.; Wang, D.S.; Wang, Z.Q.; Shao, Q.; Wang, F.H.; Yan, S.M.; Liang, J.Y.; Zeng, Z.L.; et al. Correlation between immune signature and high-density lipoprotein cholesterol level in stage II/III colorectal cancer. Cancer Med. 2019, 8, 1209–1217. [Google Scholar] [CrossRef] [Green Version]
- Hong, T.T.; Shen, D.; Chen, X.P.; Wu, X.H.; Hua, D. Preoperative serum lipid profile and outcome in nonmetastatic colorectal cancer. Chronic Dis. Transl. Med. 2016, 2, 241–249. [Google Scholar] [CrossRef]
- Brantley, K.D.; Riis, A.H.; Erichsen, R.; Thorlacius-Ussing, O.; Moller, H.J.; Lash, T.L. The association of serum lipid levels with colorectal cancer recurrence. Cancer Epidemiol. 2020, 66, 101725. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, G.; Shi, J.; Zhang, J. Increased blood lipid level is associated with cancer-specific mortality and all-cause mortality in patients with colorectal cancer (65 years): A population-based prospective cohort study. Risk Manag. Health Policy 2020, 13, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhang, L.; Chen, D.B.; Yu, X.G.; He, X.H.; Liu, M.; Chen, P.S. Validation of Prognosis Value of Cumulative Prognostic Scores Based on Serum High-Density Lipoprotein Cholesterol and Albumin Levels in Patients with Colorectal Cancer. J. Cancer 2019, 10, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Ben-Aicha, S.; Badimon, L.; Vilahur, G. Advances in HDL: Much more than lipid transporters. Int. J. Mol. Sci. 2020, 21, 732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kontush, A.; de Faria, E.C.; Chantepie, S.; Chapman, M.J. A normotriglyceridemic, low HDL-cholesterol phenotype is characterised by elevated oxidative stress and HDL particles with attenuated antioxidative activity. Atherosclerosis 2005, 82, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.J.; Skeggs, J.W.; Morton, R.E. Elevated triglyceride content diminishes the capacity of high density lipoprotein to deliver cholesteryl esters via the scavenger receptor class B type I (SR-BI). J. Biol. Chem. 2001, 276, 4804–4811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Yang, Y.; Liao, Z. Diabetes and cancer: Epidemiological and biological links. World J Diabetes 2020, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Pruzanski, W.; Stefanski, E.; de Beer, F.C.; de Beer, M.C.; Ravandi, A.; Kuksis, A. Comparative analysis of lipid composition of normal and acute-phase high density lipoproteins. J. Lipid Res. 2000, 41, 1035–1047. [Google Scholar] [CrossRef]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 117443. [Google Scholar] [CrossRef]
- Ahaneku, J.E.; Taylor, G.O.; Olubuyide, I.O.; Agbedana, E.O. Abnormal lipid and lipoprotein patterns in liver cirrhosis with and without hepatocellular carcinoma. J. Pak. Med. Assoc. 1992, 42, 260–263. [Google Scholar] [PubMed]
- Pan, B.; Ren, H.; Lv, X.; Zhao, Y.; Yu, B.; He, Y.; Ma, Y.; Niu, C.; Kong, J.; Yu, F.; et al. Hypochlorite-induced oxidative stress elevates the capability of HDL in promoting breast cancer metastasis. J. Transl. Med. 2012, 10, 65. [Google Scholar] [CrossRef] [Green Version]
- Zeljkovic, A.; Spasojevic-Kalimanovska, V.; Vekic, J.; Jelic-Ivanovic, Z.; Topic, A.; Bogavac-Stanojevic, N.; Spasic, S.; Vujovic, A.; Kalimanovska-Ostric, D. Does simultaneous determination of LDL and HDL particle size improve prediction of coronary artery disease risk? Clin. Exp. Med. 2008, 8, 109–116. [Google Scholar] [CrossRef]
- Zeljkovic, A.; Vekic, J.; Spasojevic-Kalimanovska, V.; Jelic-Ivanovic, Z.; Bogavac-Stanojevic, N.; Gulan, B.; Spasic, S. LDL and HDL subclasses in acute ischemic stroke: Prediction of risk and short-term mortality. Atherosclerosis 2010, 210, 548–554. [Google Scholar] [CrossRef]
- Vekic, J.; Zeljkovic, A.; Bogavac-Stanojevic, N.; Jelic-Ivanovic, Z.; Spasojevic-Kalimanovska, V.; Simic-Ogrizovic, S.; Dopsaj, V.; Spasic, S. Cox proportional hazard model analysis of survival in end-stage renal disease patients with small-sized high-density lipoprotein particles. Clin. Biochem. 2011, 44, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Stevanovic, M.; Vekic, J.; Bogavac-Stanojevic, N.; Janac, J.; Stjepanovic, Z.; Zeljkovic, D.; Trifunovic, B.; Spasojevic-Kalimanovska, V.; Zeljkovic, A. Significance of LDL and HDL subclasses characterization in the assessment of risk for colorectal cancer development. Biochem. Med. 2018, 28, 030703. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Broadbent, H.; Law, P.J.; Sud, A.; Palin, K.; Tuupanen, S.; Gylfe, A.; Hanninen, U.A.; Cajuso, T.; Tanskanen, T.; Kondelin, J.; et al. Mendelian randomisation implicates hyperlipidaemia as a risk factor for colorectal cancer. Int. J. Cancer 2017, 140, 2701–2708. [Google Scholar] [CrossRef] [PubMed]
- Ibanez-Sanz, G.; Diez-Villanueva, A.; Riera-Ponsati, M.; Fernandez-Villa, T.; Fernandez Navarro, P.; Bustamante, M.; Llorca, J.; Amiano, P.; Ascunce, N.; Fernandez-Tardon, G.; et al. Mendelian randomization analysis rules out disylipidaemia as colorectal cancer cause. Sci. Rep. 2019, 9, 13407. [Google Scholar] [CrossRef] [Green Version]
- Pirillo, A.; Catapano, A.L.; Norata, G.D. Biological Consequences of Dysfunctional HDL. Curr. Med. Chem. 2019, 26, 1644–1664. [Google Scholar] [CrossRef] [PubMed]
- Soran, H.; Hama, S.; Yadav, R.; Durrington, P.N. HDL functionality. Curr. Opin. Lipidol. 2012, 23, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, H.; Vincent, V.; Roy, A.; Singh, S.; Ramakrishnan, L.; Kalaivani, M.; Singh, A. HDL functions and their interaction in patients with ST elevation myocardial infarction: A case control study. Lipids Health Dis. 2020, 19, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardner, M.; Yalcinkaya, M.; Goetze, S.; Luca, E.; Balaz, M.; Hunjadi, M.; Hartung, J.; Shemet, A.; Krankel, N.; Radosavljevic, S.; et al. Structure-function relationships of HDL in diabetes and coronary heart disease. JCI Insight 2020, 5, e131491. [Google Scholar] [CrossRef] [Green Version]
- Hafiane, A.; Genest, J. High density lipoproteins: Measurement techniques and potential biomarkers of cardiovascular risk. BBA Clin. 2015, 3, 175–188. [Google Scholar] [CrossRef] [Green Version]
- Favari, E.; Thomas, M.J.; Sorci-Thomas, M.G. High-Density Lipoprotein Functionality as a New Pharmacological Target on Cardiovascular Disease: Unifying Mechanism That Explains High-Density Lipoprotein Protection Toward the Progression of Atherosclerosis. J. Cardiovasc. Pharmacol. 2018, 71, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Favari, E.; Chroni, A.; Tietge, U.J.; Zanotti, I.; Escola-Gil, J.C.; Bernini, F. Cholesterol efflux and reverse cholesterol transport. Handb. Exp. Pharmacol. 2015, 224, 181–206. [Google Scholar] [PubMed] [Green Version]
- Kosmas, C.E.; Martinez, I.; Sourlas, A.; Bouza, K.V.; Campos, F.N.; Torres, V.; Montan, P.D.; Guzman, E. High-density lipoprotein (HDL) functionality and its relevance to atherosclerotic cardiovascular disease. Drugs Context 2018, 7, 212525. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Jia, X.; Wang, Z.; Li, X.; Mu, L.; Zhang, B. The anti-inflammatory properties of HDLs are impaired in gout. Clin. Rheumatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Guirgis, F.W.; Dodani, S.; Leeuwenburgh, C.; Moldawer, L.; Bowman, J.; Kalynych, C.; Grijalva, V.; Reddy, S.T.; Jones, A.E.; Moore, F.A. HDL inflammatory index correlates with and predicts severity of organ failure in patients with sepsis and septic shock. PLoS ONE 2018, 13, e0203813. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, M.; Stefanovic, A.; Vekic, J.; Zeljkovic, A.; Gojkovic, T.; Simic-Ogrizovic, S.; Bogavac-Stanojevic, N.; Cerne, D.; Ilic, J.; Stefanovic, I.; et al. Activity of paraoxonase 1 (PON1) on HDL2 and HDL3 subclasses in renal disease. Clin. Biochem. 2018, 60, 52–58. [Google Scholar] [CrossRef]
- Bae, S.S.; Lee, Y.Y.; Shahbazian, A.; Wang, J.; Meriwether, D.; Golub, I.; Oganesian, B.; Dowd, T.; Reddy, S.T.; Charles-Schoeman, C. High- density lipoprotein function is abnormal in idiopathic inflammatory myopathies. Rheumatology 2020, 59, 3515–3525. [Google Scholar] [CrossRef] [PubMed]
- Kontush, A.; Lhomme, M.; Chapman, M.J. Unraveling the complexities of the HDL lipidome. J. Lipid Res. 2013, 54, 2950–2963. [Google Scholar] [CrossRef] [Green Version]
- Ronsein, G.E.; Vaisar, T. Deepening our understanding of HDL proteome. Expert Rev. Proteom. 2019, 16, 749–760. [Google Scholar] [CrossRef]
- Ye, J.; Luo, Q.Y.; Wang, X.P.; Liu, Z.Y.; Chen, M.X.; Huang, H.; Zhang, L. Serum Apolipoprotein A-I Combined With C-Reactive Protein Serves As A Novel Prognostic Stratification System For Colorectal Cancer. Cancer Manag. Res. 2019, 11, 9265–9276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirniö, P.; Väyrynen, J.P.; Klintrup, K.; Mäkelä, J.; Mäkinen, M.J.; Karttunen, T.J.; Tuomisto, A. Decreased serum apolipoprotein A1 levels are associated with poor survival and systemic inflammatory response in colorectal cancer. Sci. Rep. 2017, 7, 5374. [Google Scholar] [CrossRef] [PubMed]
- Borgquist, S.; Butt, T.; Almgren, P.; Shiffman, D.; Stocks, T.; Orho-Melander, M.; Manjer, J.; Melander, O. Apolipoproteins, lipids and risk of cancer. Int. J. Cancer 2016, 138, 2648–2656. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Portolés, C.; Feliu, J.; Reglero, G.; Ramírez de Molina, A. ABCA1 overexpression worsens colorectal cancer prognosis by facilitating tumour growth and caveolin-1-dependent invasiveness, and these effects can be ameliorated using the BET inhibitor apabetalone. Mol. Oncol. 2018, 12, 1735–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgila, K.; Vyrla, D.; Drakos, E. Apolipoprotein A-I (ApoA-I), Immunity, Inflammation and Cancer. Cancers 2019, 11, 1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, F.; Grijalva, V.; Navab, K.; Ganapathy, E.; Meriwether, D.; Imaizumi, S.; Navab, M.; Fogelman, A.M.; Reddy, S.T.; Farias-Eisner, R. HDL mimetics inhibit tumor development in both induced and spontaneous mouse models of colon cancer. Mol. Cancer Ther. 2012, 11, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Gkouskou, K.K.; Ioannou, M.; Pavlopoulos, G.A.; Georgila, K.; Siganou, A.; Nikolaidis, G.; Kanellis, D.C.; Moore, S.; Papadakis, K.A.; Kardassis, D.; et al. Apolipoprotein A-I inhibits experimental colitis and colitis-propelled carcinogenesis. Oncogene 2016, 35, 2496–2505. [Google Scholar] [CrossRef] [PubMed]
- Meriwether, D.; Sulaiman, D.; Volpe, C.; Dorfman, A.; Grijalva, V.; Dorreh, N.; Solorzano-Vargas, R.S.; Wang, J.; O’Connor, E.; Papesh, J.; et al. Apolipoprotein A-I mimetics mitigate intestinal inflammation in COX2-dependent inflammatory bowel disease model. J. Clin. Investig. 2019, 129, 3670–3685. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Q.; Wang, Y.; Wang, J.; Su, Y.; Wang, F.; Wang, G. AIBP and APOA-I synergistically inhibit intestinal tumor growth and metastasis by promoting cholesterol efflux. J. Transl. Med. 2019, 17, 161. [Google Scholar] [CrossRef]
- Huang, L.Z.; Gao, J.L.; Pu, C.; Zhang, P.H.; Wang, L.Z.; Feng, G.; Zhang, Y. Apolipoprotein M: Research progress, regulation and metabolic functions (Review). Mol. Med. Rep. 2015, 12, 1617–1624. [Google Scholar] [CrossRef] [Green Version]
- Wolfrum, C.; Poy, M.N.; Stoffel, M. Apolipoprotein M is required for prebeta-HDL formation and cholesterol efflux to HDL and protects against atherosclerosis. Nat. Med. 2005, 11, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Elsøe, S.; Ahnström, J.; Christoffersen, C.; Hoofnagle, A.N.; Plomgaard, P.; Heinecke, J.W.; Binder, C.J.; Björkbacka, H.; Dahlback, B.; Nielsen, L.B. Apolipoprotein M binds oxidized phospholipids and increases the antioxidant effect of HDL. Atherosclerosis 2012, 221, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, H.; Liu, H.; Yu, Y.; Zhang, J.; Li, Y.; Luo, G.; Zhang, X.; Xu, N. Increased expression levels of inflammatory cytokines and adhesion molecules in lipopolysaccharideinduced acute inflammatory apoM/ mice. Mol. Med. Rep. 2020, 22, 3117–3126. [Google Scholar] [PubMed]
- Li, T.; Yang, L.; Zhao, S.; Zhang, S. Correlation between Apolipoprotein M and Inflammatory Factors in Obese Patients. Med. Sci. Monit. 2018, 24, 5698–5703. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Yu, Y.; Feng, Y.; Zhang, J.; Jiang, Q.; Zheng, L.; Zhang, X.; Xu, N.; Luo, G. Negative Correlation Between Serum Levels of Homocysteine and Apolipoprotein M. Curr. Mol. Med. 2019, 19, 120–126. [Google Scholar] [CrossRef]
- Vekic, J.; Zeljkovic, A.; Stefanovic, A.; Jelic-Ivanovic, Z.; Spasojevic-Kalimanovska, V. Obesity and dyslipidemia. Metabolism 2019, 92, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Guo, Y.; Zhang, C.; Fan, F.; Yang, W. Sphingosine Kinase 1 and Sphingosine-1-Phosphate Signaling in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feuerborn, R.; Becker, S.; Poti, F.; Nagel, P.; Brodde, M.; Schmidt, H.; Christoffersen, C.; Ceglarek, U.; Burkhardt, R.; Nofer, J.R. High density lipoprotein (HDL)-associated sphingosine 1-phosphate (S1P) inhibits macrophage apoptosis by stimulating STAT3 activity and survivin expression. Atherosclerosis 2017, 257, 29–37. [Google Scholar] [CrossRef]
- Gu, X.; Jiang, Y.; Xue, W.; Song, C.; Wang, Y.; Liu, Y.; Cui, B. SPNS2 promotes the malignancy of colorectal cancer cells via regulating Akt and ERK pathway. Clin. Exp. Pharmacol. Physiol. 2019, 46, 861–871. [Google Scholar] [CrossRef]
- Parham, K.A.; Zebol, J.R.; Tooley, K.L.; Sun, W.Y.; Moldenhauer, L.M.; Cockshell, M.P.; Gliddon, B.L.; Moretti, P.A.; Tigyi, G.; Pitson, S.M.; et al. Sphingosine 1-phosphate is a ligand for peroxisome proliferator-activated receptor-gamma that regulates neoangiogenesis. FASEB J. 2015, 29, 3638–3653. [Google Scholar] [CrossRef]
- Ogretmen, B. Sphingolipid metabolism in cancer signalling and therapy. Nat. Rev. Cancer 2018, 18, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Nagahashi, M.; Kim, E.Y.; Harikumar, K.B.; Yamada, A.; Huang, W.C.; Hait, N.C.; Allegood, J.C.; Price, M.M.; Avni, D.; et al. Sphingosine-1-phosphate links persistent STAT3 activation, chronic intestinal inflammation, and development of colitis-associated cancer. Cancer Cell 2013, 23, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machala, M.; Prochazkova, J.; Hofmanova, J.; Kralikova, L.; Slavik, J.; Tylichova, Z.; Ovesna, P.; Kozubik, A.; Vondracek, J. Colon Cancer and Perturbations of the Sphingolipid Metabolism. Int. J. Mol. Sci. 2019, 20, 6051. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, X.; Zuo, Z.; Hao, C.; Ma, Y. Sphingosine kinase 2 promotes colorectal cancer cell proliferation and invasion by enhancing MYC expression. Tumour Biol. 2016, 37, 8455–8460. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, N.; Omori, Y.; Tanaka, K.; Ito, H.; Takagi, A.; Kojima, T.; Nakatochi, M.; Ogiso, H.; Kawamoto, Y.; Nakamura, M.; et al. Increased SPHK2 Transcription of Human Colon Cancer Cells in Serum-Depleted Culture: The Involvement of CREB Transcription Factor. J. Cell. Biochem. 2015, 116, 2227–2238. [Google Scholar] [CrossRef] [PubMed]
- Kurano, M.; Yatomi, Y. Sphingosine 1-Phosphate and Atherosclerosis. J. Atheroscler. Thromb. 2018, 25, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Xue, H.; Yu, M.; Zhou, Y.; Zhang, J.; Mu, Q.; Chen, T.; Luo, G.; Liu, J. Apolipoprotein M inhibits proliferation and migration of larynx carcinoma cells. Sci. Rep. 2020, 10, 19424. [Google Scholar] [CrossRef]
- Yu, M.; Pan, L.; Sang, C.; Mu, Q.; Zheng, L.; Luo, G.; Xu, N. Apolipoprotein M could inhibit growth and metastasis of SMMC7721 cells via vitamin D receptor signaling. Cancer Manag. Res. 2019, 11, 3691–3701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.M.; Yao, S.; Luo, K.M.; Mu, Q.F.; Yu, Y.; Luo, G.H.; Xu, N. Apolipoprotein M increases the expression of vitamin D receptor mRNA in colorectal cancer cells detected with duplex fluorescence reverse transcription-quantitative polymerase chain reaction. Mol. Med. Rep. 2017, 16, 1167–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umar, M.; Sastry, K.S.; Chouchane, A.I. Role of Vitamin D beyond the Skeletal Function: A Review of the Molecular and Clinical Studies. Int. J. Mol. Sci. 2018, 19, 1618. [Google Scholar] [CrossRef] [Green Version]
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What are their functions? Chem. Biol. Interact. 2016, 259, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotur-Stevuljevic, J.; Vekic, J.; Stefanovic, A.; Zeljkovic, A.; Ninic, A.; Ivanisevic, J.; Miljkovic, M.; Sopic, M.; Munjas, J.; Mihajlovic, M.; et al. Paraoxonase 1 and atherosclerosis-related diseases. Biofactors 2020, 46, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Bacchetti, T.; Ferretti, G.; Sahebkar, A. The role of paraoxonase in cancer. Semin. Cancer Biol. 2019, 56, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Afsar, C.U.; Gunaldi, M.; Okuturlar, Y.; Gedikbasi, A.; Tiken, E.E.; Kahraman, S.; Karaca, F.; Ercolak, V.; Karabulut, M. Paraoxonase-1 and arylesterase activities in patients with colorectal cancer. Int. J. Clin. Exp. Med. 2015, 8, 21599–21604. [Google Scholar] [PubMed]
- Mihajlovic, M.; Gojkovic, T.; Vladimirov, S.; Miljkovic, M.; Stefanovic, A.; Vekic, J.; Zeljkovic, D.; Trifunovic, B.; Kotur-Stevuljevic, J.; Spasojevic-Kalimanovska, V.; et al. Changes in lecithin: Cholesterol acyltransferase, cholesteryl ester transfer protein and paraoxonase-1 activities in patients with colorectal cancer. Clin. Biochem. 2019, 63, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Marín, M.; Moya, C.; Máñez, S. Mutual Influences between Nitric Oxide and Paraoxonase 1. Antioxidants 2019, 8, 619. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Zhou, S.; Xu, Y.; Sheng, S.; Qian, S.Y.; Huo, X. Nitric oxide synthase inhibitors 1400W and L-NIO inhibit angiogenesis pathway of colorectal cancer. Nitric Oxide 2019, 83, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Penarando, J.; Lopez-Sanchez, L.M.; Mena, R.; Guil-Luna, S.; Conde, F.; Hernandez, V.; Toledano, M.; Gudino, V.; Raponi, M.; Billard, C.; et al. A role for endothelial nitric oxide synthase in intestinal stem cell proliferation and mesenchymal colorectal cancer. BMC Biol. 2018, 16, 3. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.S.; Shafik, N.M.; Elraheem, O.A.; Abou-Elnoeman, S.E. Association of paraoxonase-1(Q192R and L55M) gene polymorphisms and activity with colorectal cancer and effect of surgical intervention. Asian Pac. J. Cancer Prev. 2015, 16, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Abudayyak, M.; Boran, T.; Tukel, R.; Oztas, E.; Ozhan, G. The Role of PON1 Variants in Disease Susceptibility in a Turkish Population. Glob. Med. Genet. 2020, 7, 41–46. [Google Scholar] [CrossRef]
- Van Der Logt, E.M.; Janssen, C.H.; Van Hooijdonk, Z.; Roelofs, H.M.; Wobbes, T.; Nagengast, F.M.; Peters, W.H. No association between genetic polymorphisms in NAD(P)H oxidase p22phox and paraoxonase 1 and colorectal cancer risk. Anticancer Res. 2005, 25, 1465–1470. [Google Scholar]
- Ose, J.; Botma, A.; Balavarca, Y.; Buck, K.; Scherer, D.; Habermann, N.; Beyerle, J.; Pfutze, K.; Seibold, P.; Kap, E.J.; et al. Pathway analysis of genetic variants in folate-mediated one-carbon metabolism-related genes and survival in a prospectively followed cohort of colorectal cancer patients. Cancer Med. 2018, 7, 2797–2807. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, X.; Wei, L.; Tan, H.; Chen, J.; Li, W.; Chan, K.; Su, Y.; Zhao, L.; Hu, S.; et al. Improved diagnostic value by combining plasma PON1 level with tumor biomarkers in Colorectal Cancer patients. J. Cancer 2020, 11, 6491–6496. [Google Scholar] [CrossRef]
- Ahn, S.B.; Sharma, S.; Mohamedali, A.; Mahboob, S.; Redmond, W.J.; Pascovici, D.; Wu, J.X.; Zaw, T.; Adhikari, S.; Vaibhav, V.; et al. Potential early clinical stage colorectal cancer diagnosis using a proteomics blood test panel. Clin. Proteom. 2019, 16, 34. [Google Scholar] [CrossRef]
- Gugliucci, A.; Menini, T. Paraoxonase 1 and HDL maturation. Clin. Chim. Acta 2015, 439, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Roncucci, L.; Mora, E.; Mariani, F.; Bursi, S.; Pezzi, A.; Rossi, G.; Pedroni, M.; Luppi, D.; Santoro, L.; Monni, S.; et al. Myeloperoxidase-positive cell infiltration in colorectal carcinogenesis as indicator of colorectal cancer risk. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2291–2297. [Google Scholar] [CrossRef] [Green Version]
- Mancini, S.; Mariani, F.; Sena, P.; Benincasa, M.; Roncucci, L. Myeloperoxidase expression in human colonic mucosa is related to systemic oxidative balance in healthy subjects. Redox Rep. 2017, 22, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarkovic, K.; Uchida, K.; Kolenc, D.; Hlupic, L.; Zarkovic, N. Tissue distribution of lipid peroxidation product acrolein in human colon carcinogenesis. Free Radic. Res. 2006, 40, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Ossoli, A.; Simonelli, S.; Vitali, C.; Franceschini, G.; Calabresi, L. Role of LCAT in Atherosclerosis. J. Atheroscler. Thromb. 2016, 23, 119–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, G.; Yi, B.; Pan, G.; Chen, X. A robust twelve-gene signature for prognosis prediction of hepatocellular carcinoma. Cancer Cell Int. 2020, 20, 207. [Google Scholar] [CrossRef]
- Long, J.; Chen, P.; Lin, J.; Bai, Y.; Yang, X.; Bian, J.; Lin, Y.; Wang, D.; Yang, X.; Zheng, Y.; et al. DNA methylation-driven genes for constructing diagnostic, prognostic, and recurrence models for hepatocellular carcinoma. Theranostics 2019, 9, 7251–7267. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Yang, X.B.; Sang, X.T. Construction of a lipid metabolism-related and immune-associated prognostic signature for hepatocellular carcinoma. Cancer Med. 2020, 9, 7646–7662. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.R.; Graham, C.; D’Amato, A.; Gentry-Maharaj, A.; Ryan, A.; Kalsi, J.K.; Ainley, C.; Whetton, A.D.; Menon, U.; Jacobs, I.; et al. A combined biomarker panel shows improved sensitivity for the early detection of ovarian cancer allowing the identification of the most aggressive type II tumours. Br. J. Cancer 2017, 117, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Simonelli, S.; Gianazza, E.; Mombelli, G.; Bondioli, A.; Ferraro, G.; Penco, S.; Sirtori, C.R.; Franceschini, G.; Calabresi, L. Severe high-density lipoprotein deficiency associated with autoantibodies against lecithin:cholesterol acyltransferase in non-Hodgkin lymphoma. Arch. Intern. Med. 2012, 172, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Cooke, A.L.; Morris, J.; Melchior, J.T.; Street, S.E.; Jerome, W.G.; Huang, R.; Herr, A.B.; Smith, L.E.; Segrest, J.P.; Remaley, A.T.; et al. A thumbwheel mechanism for APOA1 activation of LCAT activity in HDL. J. Lipid Res. 2018, 59, 1244–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engstrand, J.; Nilsson, H.; Strömberg, C.; Jonas, E.; Freedman, J. Colorectal cancer liver metastases—A population-based study on incidence, management and survival. BMC Cancer 2018, 18, 78. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R. Introduction to Lipids and Lipoproteins. Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dungan, K., Grossman, A., Hershman, J.M., Hofland, J., Kaltsas, G., et al., Eds.; South Dartmouth, MA, USA, 2000. Available online: https://www.endotext.org/ (accessed on 15 March 2021).
- Gauthier, A.; Lau, P.; Zha, X.; Milne, R.; McPherson, R. Cholesteryl ester transfer protein directly mediates selective uptake of high density lipoprotein cholesteryl esters by the liver. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2177–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izem, L.; Morton, R.E. Possible role for intracellular cholesteryl ester transfer protein in adipocyte lipid metabolism and storage. J. Biol. Chem. 2007, 282, 21856–21865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granot, E.; Tabas, I.; Tall, A.R. Human plasma cholesteryl ester transfer protein enhances the transfer of cholesteryl ester from high density lipoproteins into cultured HepG2 cells. J. Biol. Chem. 1987, 262, 3482–3487. [Google Scholar] [CrossRef]
- Huang, Z.; Inazu, A.; Kawashiri, M.A.; Nohara, A.; Higashikata, T.; Mabuchi, H. Dual effects on HDL metabolism by cholesteryl ester transfer protein inhibition in HepG2 cells. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E1210–E1219. [Google Scholar] [CrossRef] [Green Version]
- Esau, L.; Sagar, S.; Bangarusamy, D.; Kaur, M. Identification of CETP as a molecular target for estrogen positive breast cancer cell death by cholesterol depleting agents. Genes Cancer 2016, 7, 309–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.E.; Siewert, K.M.; Klarin, D.; Damrauer, S.M.; Program, V.A.M.V.; Chang, K.M.; Tsao, P.S.; Assimes, T.L.; Maxwell, K.N.; Voight, B.F. The relationship between circulating lipids and breast cancer risk: A Mendelian randomization study. PLoS Med. 2020, 17, e1003302. [Google Scholar] [CrossRef]
- Nowak, C.; Arnlov, J. A Mendelian randomization study of the effects of blood lipids on breast cancer risk. Nat. Commun. 2018, 9, 3957. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Duan, J.; Wang, H.; Zhang, Y.; Qiao, K.; Wang, J. Cholesterol depletion sensitizes gallbladder cancer to cisplatin by impairing DNA damage response. Cell Cycle 2019, 18, 3337–3350. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, I.; Mastalier, B. Genetics of colorectal cancer. J. Med. Life 2014, 7, 507. [Google Scholar]
- Demina, E.P.; Miroshnikova, V.V.; Schwarzman, A.L. Role of the ABC transporters A1 and G1, key reverse cholesterol transport proteins, in atherosclerosis. Mol. Biol. Mosk 2016, 50, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Ogata, M.; Tsujita, M.; Hossain, M.A.; Akita, N.; Gonzalez, F.J.; Staels, B.; Suzuki, S.; Fukutomi, T.; Kimura, G.; Yokoyama, S. On the mechanism for PPAR agonists to enhance ABCA1 gene expression. Atherosclerosis 2009, 205, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, S.; Born, E.; Mathur, S.N.; Field, F.J. LXR/RXR activation enhances basolateral efflux of cholesterol in CaCo-2 cells. J. Lipid Res. 2002, 43, 1054–1064. [Google Scholar] [CrossRef] [Green Version]
- Miroshnikova, V.V.; Panteleeva, A.A.; Bazhenova, E.A.; Demina, E.P.; Usenko, T.S.; Nikolaev, M.A.; Semenova, I.A.; Neimark, A.E.; He, J.; Belyaeva, O.D.; et al. Regulation of ABCA1 and ABCG1 gene expression in the intraabdominal adipose tissue. Biomed. Khim. 2016, 62, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Po, A.; Citarella, A.; Catanzaro, G.; Besharat, Z.M.; Trocchianesi, S.; Gianno, F.; Sabato, C.; Moretti, M.; De Smaele, E.; Vacca, A.; et al. Hedgehog-GLI signalling promotes chemoresistance through the regulation of ABC transporters in colorectal cancer cells. Sci. Rep. 2020, 10, 13988. [Google Scholar] [CrossRef] [PubMed]
- Hlavata, I.; Mohelnikova-Duchonova, B.; Vaclavikova, R.; Liska, V.; Pitule, P.; Novak, P.; Bruha, J.; Vycital, O.; Holubec, L.; Treska, V.; et al. The role of ABC transporters in progression and clinical outcome of colorectal cancer. Mutagenesis 2012, 27, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeljkovic, A.; Mihajlovic, M.; Stefanovic, A.; Zeljkovic, D.; Trifunovic, B.; Miljkovic, M.; Spasojevic-Kalimanovska, V.; Vekic, J. Potential use of serum insulin-like growth factor 1 and E-cadherin as biomarkers of colorectal cancer. Colorectal Dis. 2020. [Google Scholar] [CrossRef]
- Vargas, T.; Moreno-Rubio, J.; Herranz, J.; Cejas, P.; Molina, S.; Gonzalez-Vallinas, M.; Mendiola, M.; Burgos, E.; Aguayo, C.; Custodio, A.B.; et al. ColoLipidGene: Signature of lipid metabolism-related genes to predict prognosis in stage-II colon cancer patients. Oncotarget 2015, 6, 7348–7363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, L.P.; Ramos-Ruiz, R.; Herranz, J.; Martin-Hernandez, R.; Vargas, T.; Mendiola, M.; Guerra, L.; Reglero, G.; Feliu, J.; Ramirez de Molina, A. The transcriptional and mutational landscapes of lipid metabolism-related genes in colon cancer. Oncotarget 2018, 9, 5919–5930. [Google Scholar] [CrossRef]
- Namba, Y.; Sogawa, C.; Okusha, Y.; Kawai, H.; Itagaki, M.; Ono, K.; Murakami, J.; Aoyama, E.; Ohyama, K.; Asaumi, J.I.; et al. Depletion of Lipid Efflux Pump ABCG1 Triggers the Intracellular Accumulation of Extracellular Vesicles and Reduces Aggregation and Tumorigenesis of Metastatic Cancer Cells. Front. Oncol. 2018, 8, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchino, K.; Ochiya, T.; Takeshita, F. RNAi therapeutics and applications of microRNAs in cancer treatment. JPN J. Clin. Oncol. 2013, 43, 596–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, D.P.; Yin, C.H.; Zhang, X.Y.; Yang, N.N.; Xu, J.Y. MiR-183 functions as an oncogene by targeting ABCA1 in colon cancer. Oncol. Rep. 2016, 35, 2873–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klett, E.L.; Lee, M.H.; Adams, D.B.; Chavin, K.D.; Patel, S.B. Localization of ABCG5 and ABCG8 proteins in human liver, gall bladder and intestine. BMC Gastroenterol. 2004, 4, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repa, J.J.; Berge, K.E.; Pomajzl, C.; Richardson, J.A.; Hobbs, H.; Mangelsdorf, D.J. Regulation of ATP-binding cassette sterol transporters ABCG5 and ABCG8 by the liver X receptors alpha and beta. J. Biol. Chem. 2002, 277, 18793–18800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumi, K.; Tanaka, T.; Uchida, A.; Magoori, K.; Urashima, Y.; Ohashi, R.; Ohguchi, H.; Okamura, M.; Kudo, H.; Daigo, K.; et al. Cooperative interaction between hepatocyte nuclear factor 4 alpha and GATA transcription factors regulates ATP-binding cassette sterol transporters ABCG5 and ABCG8. Mol. Cell. Biol. 2007, 27, 4248–4260. [Google Scholar] [CrossRef] [Green Version]
- Aulchenko, Y.S.; Ripatti, S.; Lindqvist, I.; Boomsma, D.; Heid, I.M.; Pramstaller, P.P.; Penninx, B.W.; Janssens, A.C.; Wilson, J.F.; Spector, T.; et al. Loci influencing lipid levels and coronary heart disease risk in 16 European population cohorts. Nat. Genet. 2009, 41, 47–55. [Google Scholar] [PubMed]
- Junyent, M.; Tucker, K.L.; Smith, C.E.; Garcia-Rios, A.; Mattei, J.; Lai, C.Q.; Parnell, L.D.; Ordovas, J.M. The effects of ABCG5/G8 polymorphisms on plasma HDL cholesterol concentrations depend on smoking habit in the Boston Puerto Rican Health Study. J. Lipid Res. 2009, 50, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Hostettler, L.; Zlobec, I.; Terracciano, L.; Lugli, A. ABCG5-positivity in tumor buds is an indicator of poor prognosis in node-negative colorectal cancer patients. World J. Gastroenterol. 2010, 16, 732–739. [Google Scholar] [CrossRef]
- Shen, W.J.; Azhar, S.; Kraemer, F.B. SR-B1: A Unique Multifunctional Receptor for Cholesterol Influx and Efflux. Annu. Rev. Physiol. 2018, 80, 95–116. [Google Scholar] [CrossRef]
- Malerød, L.; Juvet, K.; Gjøen, T.; Berg, T. The expression of scavenger receptor class B, type I (SR-BI) and caveolin-1 in parenchymal and nonparenchymal liver cells. Cell Tissue Res. 2002, 307, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Pajares, J.L.; Ben Hassen, C.; Chevalier, S.; Frank, P.G. SR-BI: Linking Cholesterol and Lipoprotein Metabolism with Breast and Prostate Cancer. Front. Pharmacol. 2016, 7, 338. [Google Scholar] [CrossRef] [Green Version]
- Mooberry, L.K.; Sabnis, N.A.; Panchoo, M.; Nagarajan, B.; Lacko, A.G. Targeting the SR-B1 Receptor as a Gateway for Cancer Therapy and Imaging. Front. Pharmacol. 2016, 7, 466. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, M.M.; Mangala, L.S.; Han, H.D.; Lu, C.; Bottsford-Miller, J.; Nishimura, M.; Mora, E.M.; Lee, J.W.; Stone, R.L.; Pecot, C.V.; et al. Targeted delivery of small interfering RNA using reconstituted high-density lipoprotein nanoparticles. Neoplasia 2011, 13, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Sharma, B.; Gupta, V.; Dahiya, D.; Kumar, H.; Vaiphei, K.; Agnihotri, N. Clinical relevance of cholesterol homeostasis genes in colorectal cancer. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 1314–1327. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, Y.; Zhao, S.; Zheng, Z.; Shen, C.; An, L.; Yuan, Y. Large-scale analysis reveals a novel risk score to predict overall survival in hepatocellular carcinoma. Cancer Manag. Res. 2018, 10, 6079–6096. [Google Scholar] [CrossRef] [Green Version]
- Dachet, C.; Poirier, O.; Cambien, F.; Chapman, J.; Rouis, M. New functional promoter polymorphism, CETP/-629, in cholesteryl ester transfer protein (CETP) gene related to CETP mass and high density lipoprotein cholesterol levels: Role of Sp1/Sp3 in transcriptional regulation. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhofer, A.; Tatarczyk, T.; Laimer, M.; Ritsch, A.; Kaser, S.; Paulweber, B.; Ebenbichler, C.F.; Patsch, J.R. The Taq1B-variant in the cholesteryl ester-transfer protein gene and the risk of metabolic syndrome. Obesity 2008, 16, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Ma, R.; Guo, H.; He, J.; Hu, Y.; Mu, L.; Yan, Y.; Ma, J.; Li, S.; Zhang, J.; et al. Association between Six CETP Polymorphisms and Metabolic Syndrome in Uyghur Adults from Xinjiang, China. Int. J. Environ. Res. Public Health 2017, 14, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dizaji, B.F.; Rivandi, M.; Javandoost, A.; Karimian, M.S.; Raei, A.; Sahebkar, A.; Ferns, G.; Mobarhan, M.G.; Pasdar, A. Association of genetic polymorphisms of PON1 and CETP with the presence of metabolic syndrome; the effects of genotypes on their serum activity and concentrations. Egypt. J. Med Hum. Genet. 2018, 19, 43–48. [Google Scholar] [CrossRef]
- Milano, A.; Bianco, M.A.; Buri, L.; Cipolletta, L.; Grossi, E.; Rotondano, G.; Tessari, F.; Efthymakis, K.; Neri, M. Metabolic syndrome is a risk factor for colorectal adenoma and cancer: A study in a White population using the harmonized criteria. Therap. Adv. Gastroenterol. 2019, 12, 1756284819867839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saetang, J.; Sangkhathat, S. Diets link metabolic syndrome and colorectal cancer development (Review). Oncol. Rep. 2017, 37, 1312–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Báez, S.; Tsuchiya, Y.; Calvo, A.; Pruyas, M.; Nakamura, K.; Kiyohara, C.; Oyama, M.; Yamamoto, M. Genetic variants involved in gallstone formation and capsaicin metabolism, and the risk of gallbladder cancer in Chilean women. World J. Gastroenterol. 2010, 16, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Sidaraite, A.; Liutkeviciene, R.; Glebauskiene, B.; Vilkeviciute, A.; Kriauciuniene, L. Associations of cholesteryl ester transfer protein (CETP) gene variants with pituitary adenoma. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech Repub. 2020, 164, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Guo, H.; Li, Y.; Wang, H.; He, J.; Mu, L.; Hu, Y.; Ma, J.; Yan, Y.; Li, S.; et al. Interactions among genes involved in reverse cholesterol transport and in the response to environmental factors in dyslipidemia in subjects from the Xinjiang rural area. PLoS ONE 2018, 13, e0196042. [Google Scholar] [CrossRef]
- Bathaie, S.Z.; Ashrafi, M.; Azizian, M.; Tamanoi, F. Mevalonate Pathway and Human Cancers. Curr. Mol. Pharmacol. 2017, 10, 77–85. [Google Scholar]
- Hopkins, R.J.; Young, R.P. Mevalonate signaling, COPD and cancer: The statins and beyond. J. Investig. Med. 2019, 67, 711–714. [Google Scholar] [CrossRef]
- Long, J.; Zhang, C.J.; Zhu, N.; Du, K.; Yin, Y.F.; Tan, X.; Liao, D.F.; Qin, L. Lipid metabolism and carcinogenesis, cancer development. Am. J. Cancer Res. 2018, 8, 778–791. [Google Scholar]
- Broitman, S.A.; Cerda, S.; Wilkinson, J., 4th. Cholesterol metabolism and colon cancer. Prog. Food Nutr. Sci. 1993, 17, 1–40. [Google Scholar] [PubMed]
- Chimento, A.; Casaburi, I.; Avena, P.; Trotta, F.; De Luca, A.; Rago, V.; Pezzi, V.; Sirianni, R. Cholesterol and Its Metabolites in Tumor Growth: Therapeutic Potential of Statins in Cancer Treatment. Front. Endocrinol. 2018, 9, 807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajora, M.A.; Zheng, G. Targeting SR-BI for Cancer Diagnostics, Imaging and Therapy. Front. Pharmacol. 2016, 7, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilo, C.; Frank, P.G. Cholesterol and breast cancer development. Curr. Opin. Pharmacol. 2012, 12, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Danilo, C.; Gutierrez-Pajares, J.L.; Mainieri, M.A.; Mercier, I.; Lisanti, M.P.; Frank, P.G. Scavenger receptor class B type I regulates cellular cholesterol metabolism and cell signaling associated with breast cancer development. Breast Cancer Res. 2013, 15, R87. [Google Scholar] [CrossRef] [PubMed]
- Sekine, Y.; Demosky, S.J.; Stonik, J.A.; Furuya, Y.; Koike, H.; Suzuki, K.; Remaley, A.T. High-density lipoprotein induces proliferation and migration of human prostate androgen-independent cancer cells by an ABCA1-dependent mechanism. Mol. Cancer Res. 2010, 8, 1284–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llaverias, G.; Danilo, C.; Mercier, I.; Daumer, K.; Capozza, F.; Williams, T.M.; Sotgia, F.; Lisanti, M.P.; Frank, P.G. Role of cholesterol in the development and progression of breast cancer. Am. J. Pathol. 2011, 178, 402–412. [Google Scholar] [CrossRef]
- Samadi, S.; Ghayour-Mobarhan, M.; Mohammadpour, A.; Farjami, Z.; Tabadkani, M.; Hosseinnia, M.; Miri, M.; Heydari-Majd, M.; Mehramiz, M.; Rezayi, M.; et al. High-density lipoprotein functionality and breast cancer: A potential therapeutic target. J. Cell Biochem. 2019, 120, 5756–5765. [Google Scholar] [CrossRef]
- Vladimirov, S.; Zeljkovic, A.; Gojkovic, T.; Miljkovic, M.; Stefanovic, A.; Zeljkovic, D.; Trifunovic, B.; Spasojevic-Kalimanovska, V. Associations of cholesterol and vitamin D metabolites with the risk for development of high grade colorectal cancer. J. Med. Biochem. 2020, 39, 318–327. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Yokoyama, S.; Chang, T.Y. Methods for Monitoring ABCA1-Dependent Sterol Release. Methods Mol. Biol. 2017, 1583, 257–273. [Google Scholar]
- Gomaraschi, M. Role of Lipoproteins in the Microenvironment of Hormone-Dependent Cancers. Trends Endocrinol. Metab. 2020, 31, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Warns, J.; Marwarha, G.; Freking, N.; Ghribi, O. 27-hydroxycholesterol decreases cell proliferation in colon cancer cell lines. Biochimie 2018, 153, 171–180. [Google Scholar] [CrossRef]
- Rossin, D.; Calfapietra, S.; Sottero, B.; Poli, G.; Biasi, F. HNE and cholesterol oxidation products in colorectal inflammation and carcinogenesis. Free Radic. Biol. Med. 2017, 111, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Traversari, C.; Sozzani, S.; Steffensen, K.R.; Russo, V. LXR-dependent and -independent effects of oxysterols on immunity and tumor growth. Eur. J. Immunol. 2014, 44, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- Swan, R.; Alnabulsi, A.; Cash, B.; Alnabulsi, A.; Murray, G.I. Characterisation of the oxysterol metabolising enzyme pathway in mismatch repair proficient and deficient colorectal cancer. Oncotarget 2016, 7, 46509–46527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossin, D.; Dias, I.H.K.; Solej, M.; Milic, I.; Pitt, A.R.; Iaia, N.; Scoppapietra, L.; Devitt, A.; Nano, M.; Degiuli, M.; et al. Increased production of 27-hydroxycholesterol in human colorectal cancer advanced stage: Possible contribution to cancer cell survival and infiltration. Free Radic. Biol. Med. 2019, 136, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; You, N.Y.; Kim, Y.; Kim, Y.; Kim, J.; Kang, H.T. Statin use and site-specific risk of colorectal cancer in individuals with hypercholesterolemia from the National Health Insurance Service-National Health Screening Cohort (NHIS-HEALS). Nutr. Metab. Cardiovasc. Dis. 2019, 29, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.C.; Glass, J.; Giustino, G.; Hove, J.R.T.; Castaneda, D.; Torres, J.; Kumar, A.; Elman, J.; Ullman, T.A.; Itzkowitz, S.H. Statin Exposure Is Not Associated with Reduced Prevalence of Colorectal Neoplasia in Patients with Inflammatory Bowel Disease. Gut Liver 2019, 13, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Dobrzycka, M.; Spychalski, P.; Lachinski, A.J.; Kobiela, P.; Jedrusik, P.; Kobiela, J. Statins and Colorectal Cancer—A Systematic Review. Exp. Clin. Endocrinol. Diabetes 2020, 128, 255–262. [Google Scholar] [CrossRef]
- Hryniewicz-Jankowska, A.; Augoff, K.; Biernatowska, A.; Podkalicka, J.; Sikorski, A.F. Membrane rafts as a novel target in cancer therapy. Biochim. Biophys. Acta 2014, 1845, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Meng, S.; Gu, Q.; Araujo-Gutierrez, R.; Kumar, S.; Yan, Q.; Almazan, F.; Youker, K.A.; Fu, Y.; Pownall, H.J.; et al. AIBP Limits Angiogenesis Through gamma-Secretase-Mediated Upregulation of Notch Signaling. Circ. Res. 2017, 120, 1727–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Vakoc, C.R. Targeting Cancer Cells with BET Bromodomain Inhibitors. Cold Spring Harb. Perspect. Med. 2017, 7, a026674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilham, D.; Wasiak, S.; Tsujikawa, L.M.; Halliday, C.; Norek, K.; Patel, R.G.; Kulikowski, E.; Johansson, J.; Sweeney, M.; Wong, N.C.; et al. RVX-208, a BET-inhibitor for treating atherosclerotic cardiovascular disease, raises ApoA-I/HDL and represses pathways that contribute to cardiovascular disease. Atherosclerosis 2016, 247, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell. Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Sherman, C.B.; Peterson, S.J.; Frishman, W.H. Apolipoprotein A-I mimetic peptides: A potential new therapy for the prevention of atherosclerosis. Cardiol. Rev. 2010, 18, 141–147. [Google Scholar] [CrossRef]
- Reddy, S.T.; Navab, M.; Anantharamaiah, G.M.; Fogelman, A.M. Apolipoprotein A-I mimetics. Curr. Opin. Lipidol. 2014, 25, 304–308. [Google Scholar] [CrossRef] [Green Version]
- Cedó, L.; García-León, A.; Baila-Rueda, L.; Santos, D.; Grijalva, V.; Martinez-Cignoni, M.R.; Carbó, J.M.; Metso, J.; López-Vilaró, L.; Zorzano, A.; et al. ApoA-I mimetic administration, but not increased apoA-I-containing HDL, inhibits tumour growth in a mouse model of inherited breast cancer. Sci. Rep. 2016, 6, 36387. [Google Scholar] [CrossRef]
- Delk, S.C.; Chattopadhyay, A.; Escola-Gil, J.C.; Fogelman, A.M.; Reddy, S.T. Apolipoprotein mimetics in cancer. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Raut, S.; Mooberry, L.; Sabnis, N.; Garud, A.; Dossou, A.S.; Lacko, A. Reconstituted HDL: Drug Delivery Platform for Overcoming Biological Barriers to Cancer Therapy. Front. Pharmacol. 2018, 9, 1154. [Google Scholar] [CrossRef] [PubMed]
- Aghamiri, S.; Jafarpour, A.; Malekshahi, Z.V.; Mahmoudi Gomari, M.; Negahdari, B. Targeting siRNA in colorectal cancer therapy: Nanotechnology comes into view. J. Cell Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Damiano, M.G.; Zhang, H.; Tripathy, S.; Luthi, A.J.; Rink, J.S.; Ugolkov, A.V.; Singh, A.T.; Dave, S.S.; Gordon, L.I.; et al. Biomimetic, synthetic HDL nanostructures for lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 2511–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Chen, Z.; Zeng, X.; Chen, X.; Gu, Z. Advances in nanomedicine for cancer starvation therapy. Theranostics 2019, 9, 8026–8047. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Zhang, T.; He, W.; Jin, H.; Liu, C.; Yang, Z.; Ren, J. Methotrexate-Loaded Extracellular Vesicles Functionalized with Therapeutic and Targeted Peptides for the Treatment of Glioblastoma Multiforme. ACS Appl. Mater. Interfaces 2018, 10, 12341–12350. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeljkovic, A.; Vekic, J.; Mihajlovic, M.; Gojkovic, T.; Vladimirov, S.; Zeljkovic, D.; Spasojevic-Kalimanovska, V.; Trifunovic, B. Revealing the Role of High-Density Lipoprotein in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 3352. https://doi.org/10.3390/ijms22073352
Zeljkovic A, Vekic J, Mihajlovic M, Gojkovic T, Vladimirov S, Zeljkovic D, Spasojevic-Kalimanovska V, Trifunovic B. Revealing the Role of High-Density Lipoprotein in Colorectal Cancer. International Journal of Molecular Sciences. 2021; 22(7):3352. https://doi.org/10.3390/ijms22073352
Chicago/Turabian StyleZeljkovic, Aleksandra, Jelena Vekic, Marija Mihajlovic, Tamara Gojkovic, Sandra Vladimirov, Dejan Zeljkovic, Vesna Spasojevic-Kalimanovska, and Bratislav Trifunovic. 2021. "Revealing the Role of High-Density Lipoprotein in Colorectal Cancer" International Journal of Molecular Sciences 22, no. 7: 3352. https://doi.org/10.3390/ijms22073352