
Fabrication of Adipose-Derived Stem Cell-Based Self-Assembled Scaffold under Hypoxia and Mechanical Stimulation for Urethral Tissue Engineering

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
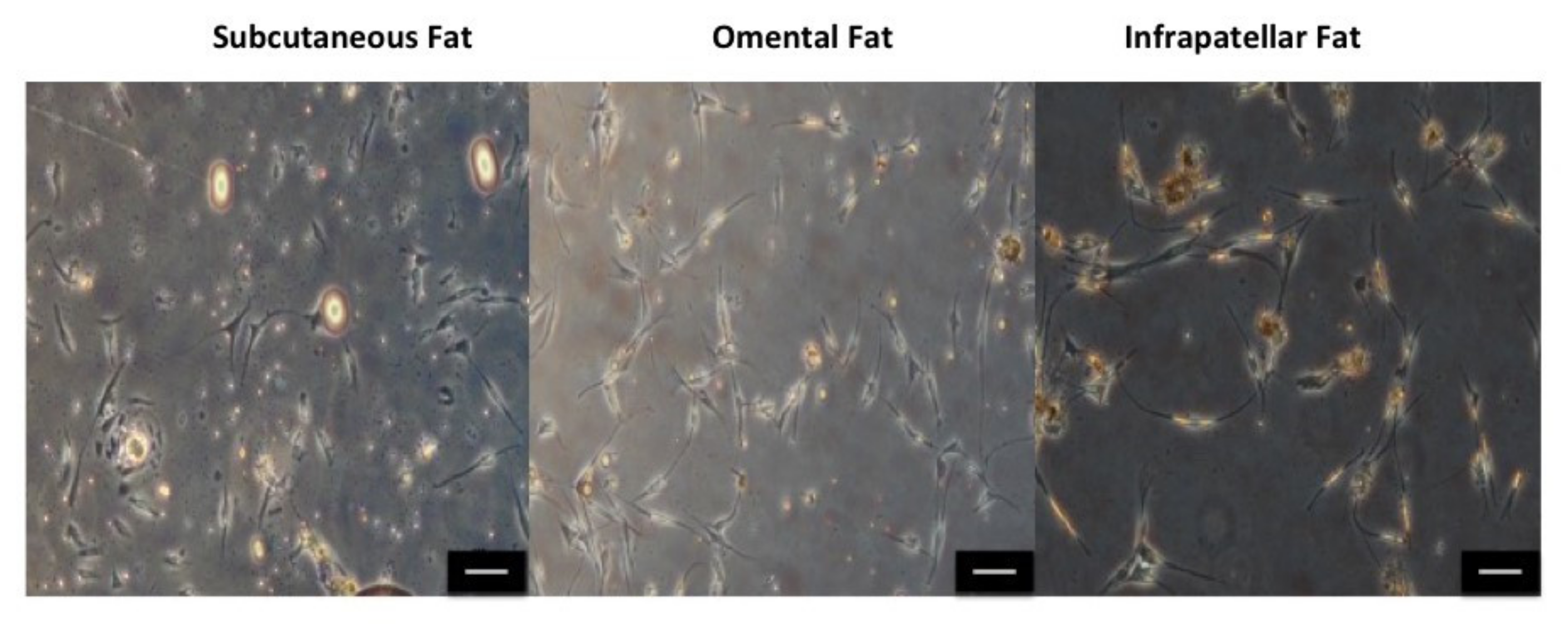
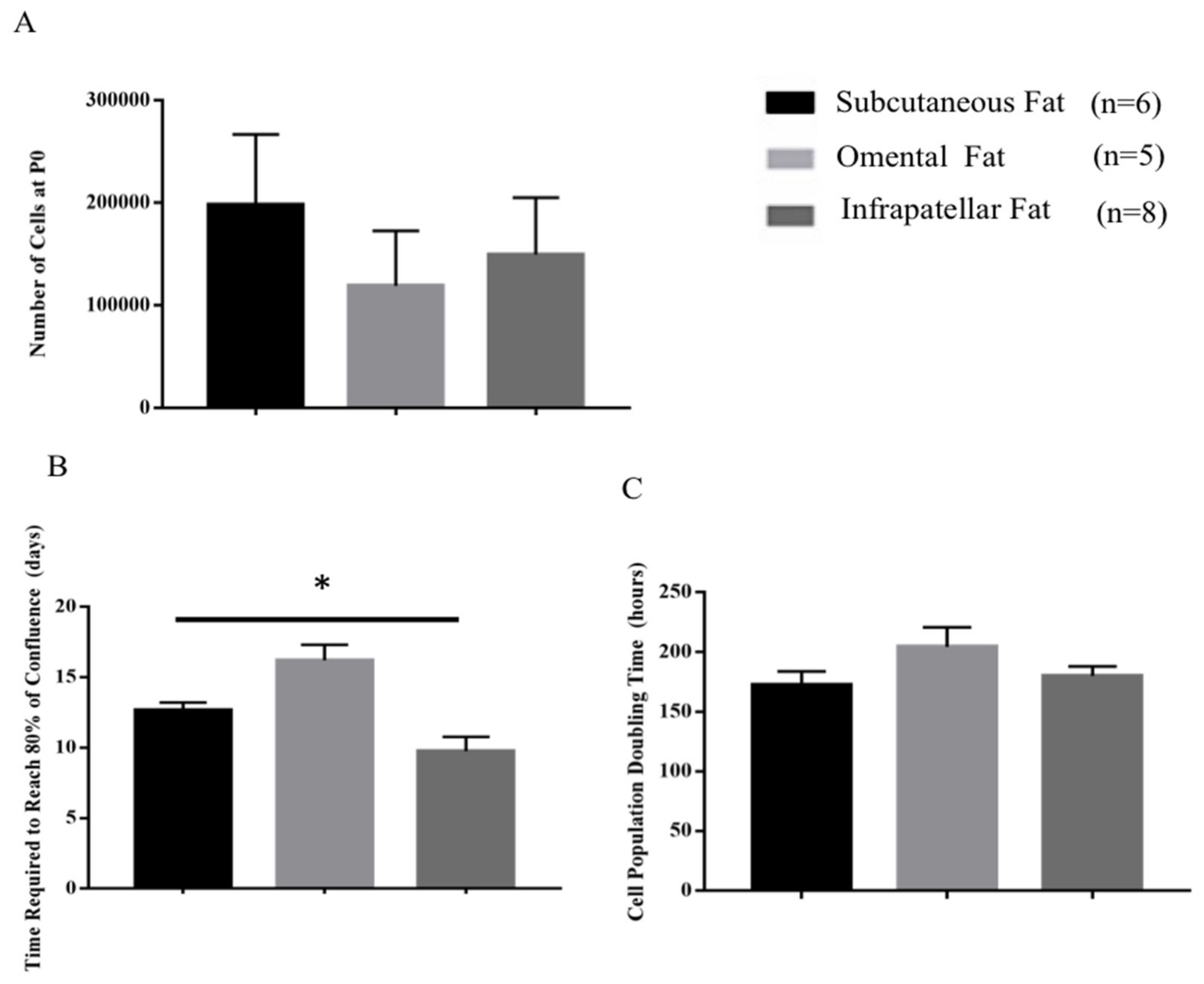
2.1. Cell Morphology, Cell Yield and Population Doubling Time
2.2. Thickness Optimization Of ASC-Based Self-Assembled Scaffold under Different Parameters
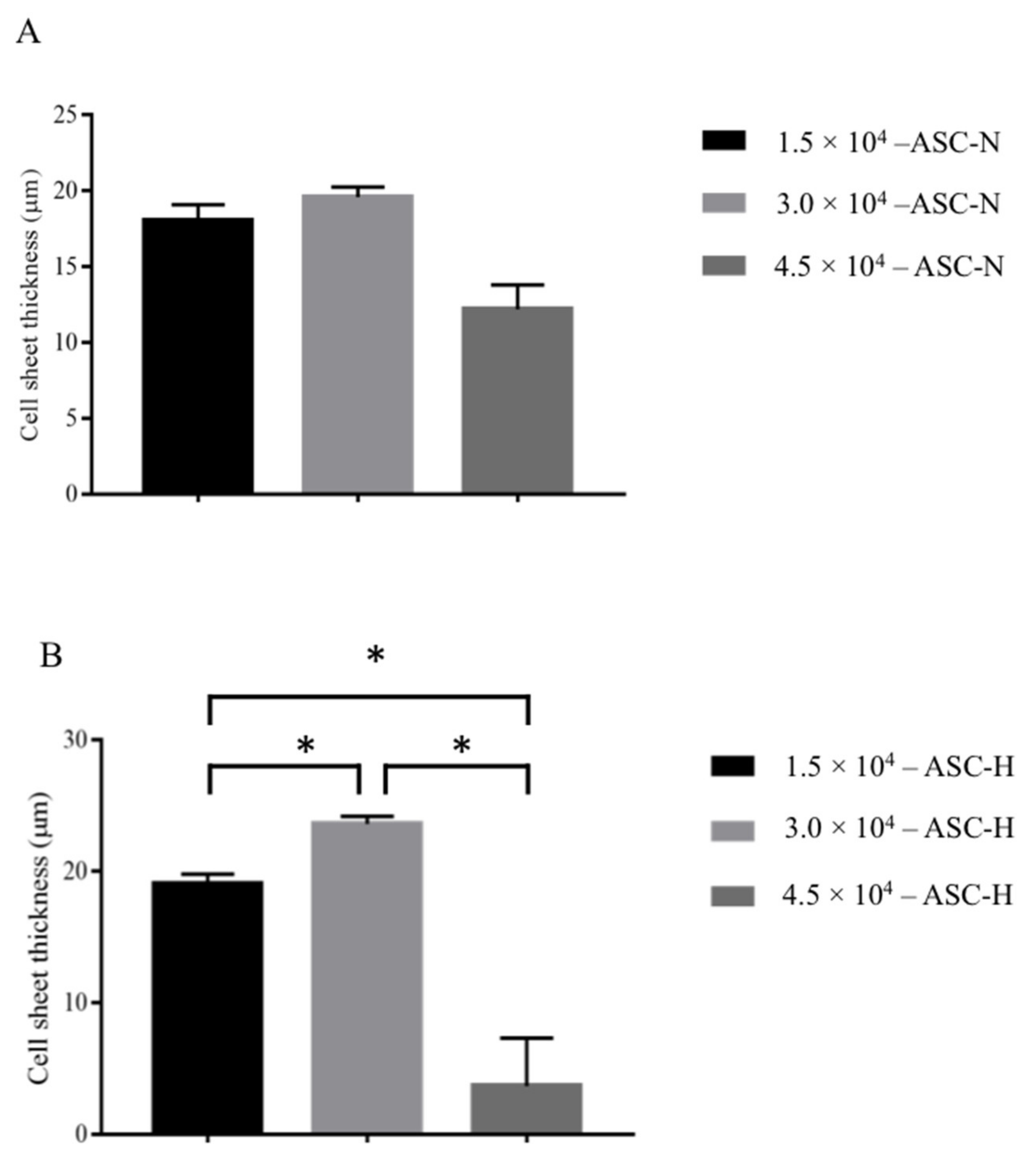
2.2.1. Different Seeding Densities and under Normoxic (21% O2) and Hypoxic (1% O2) Culture Conditions
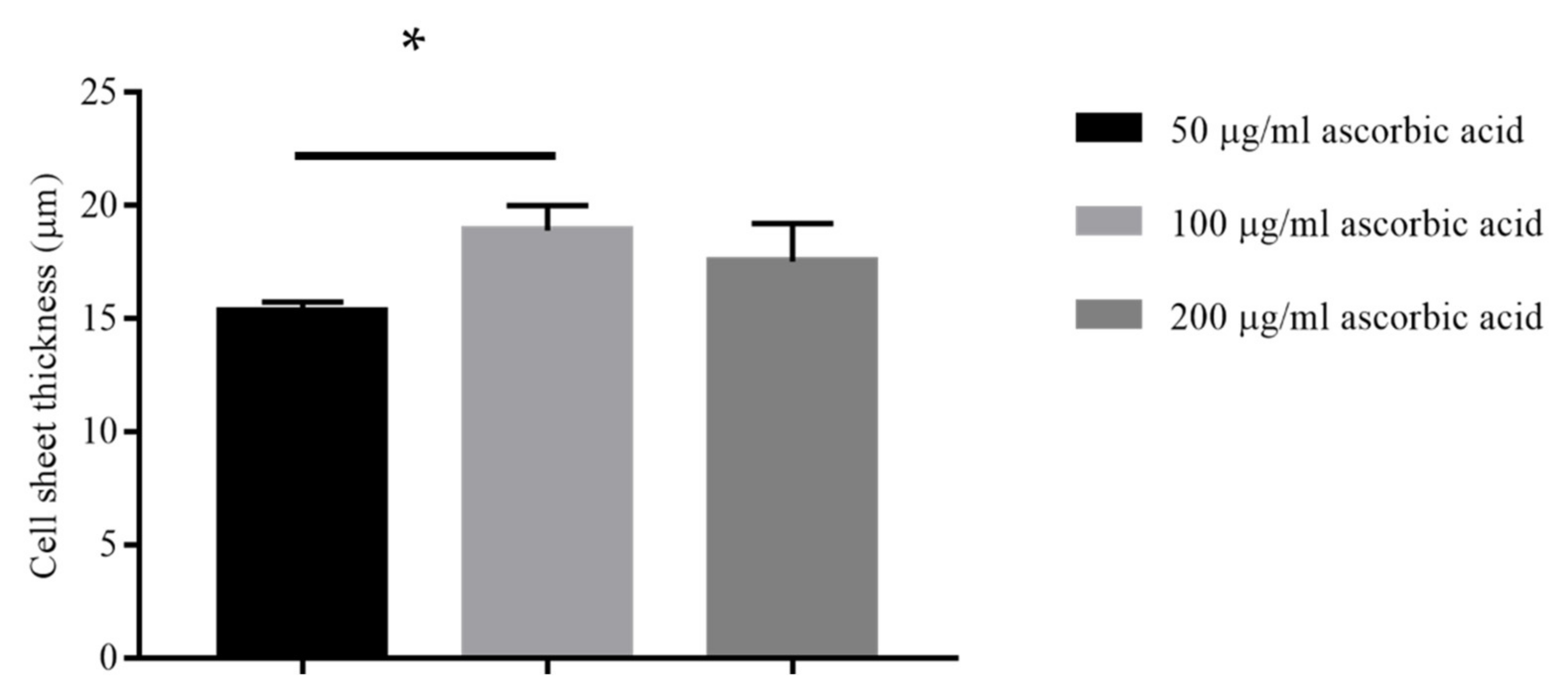
2.2.2. Different Concentrations of Ascorbic Acid
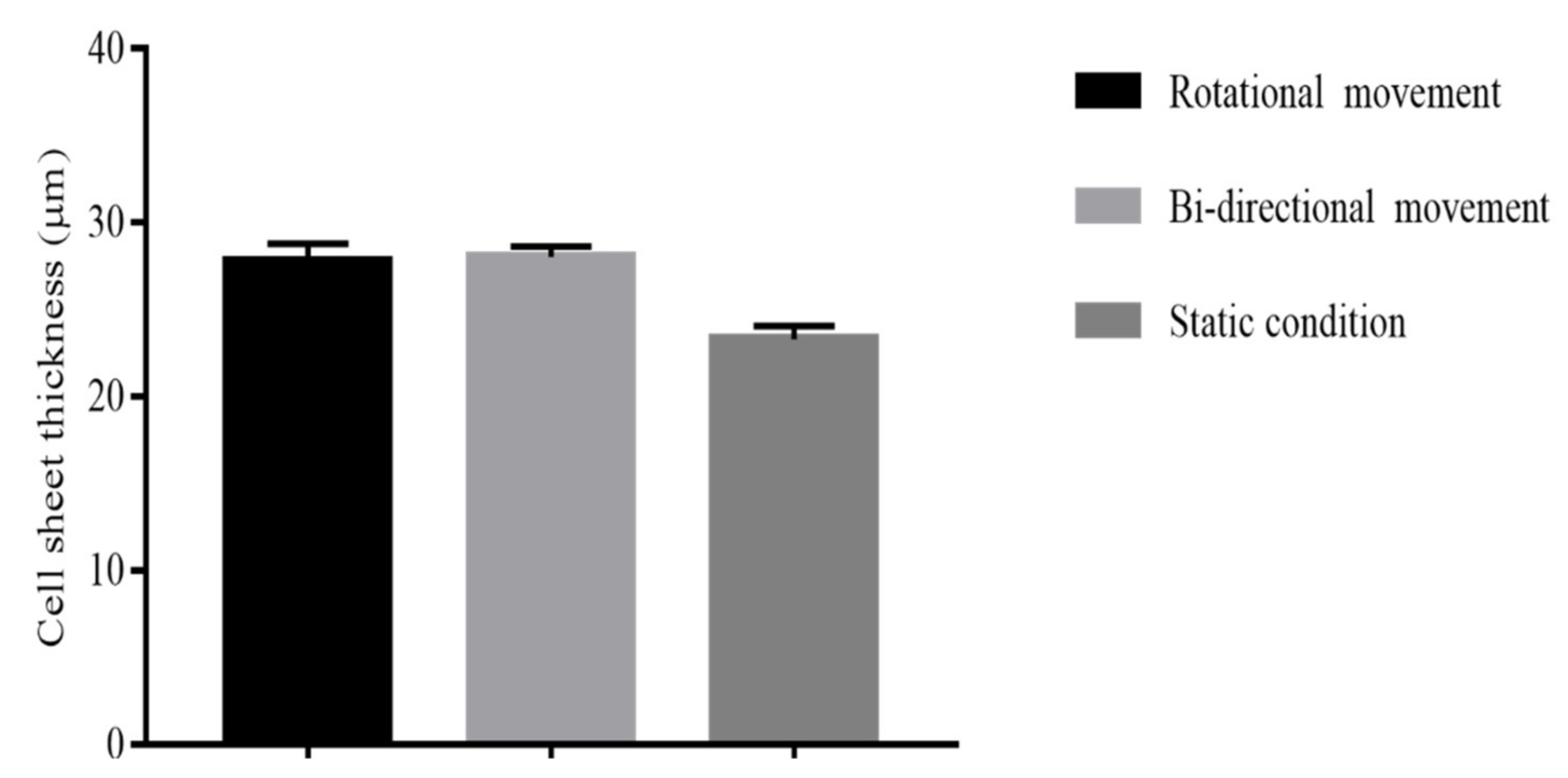
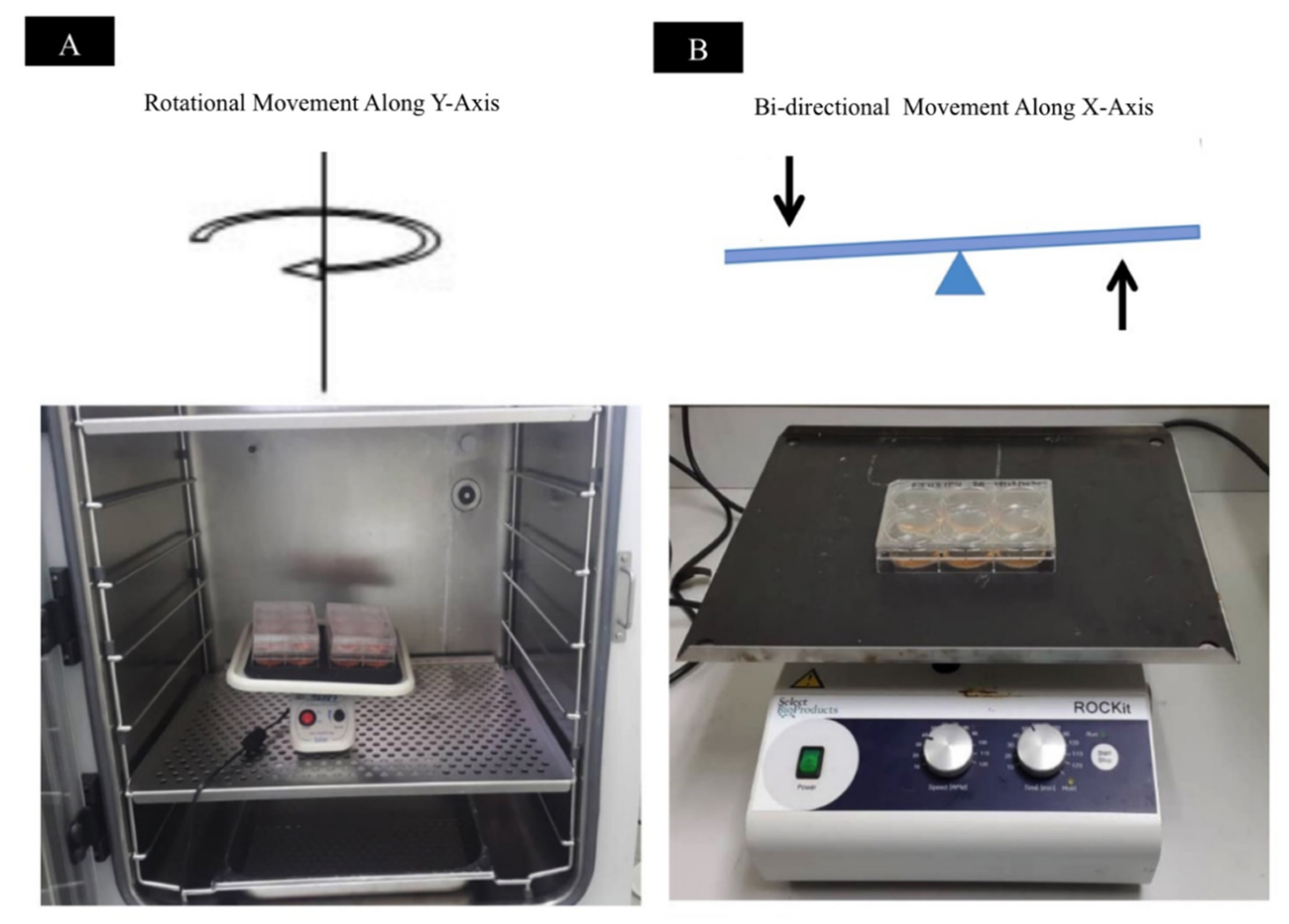
2.2.3. Different Mechanical Stimulation
2.3. Characterization Of ASC-Based Self-Assembled Scaffold
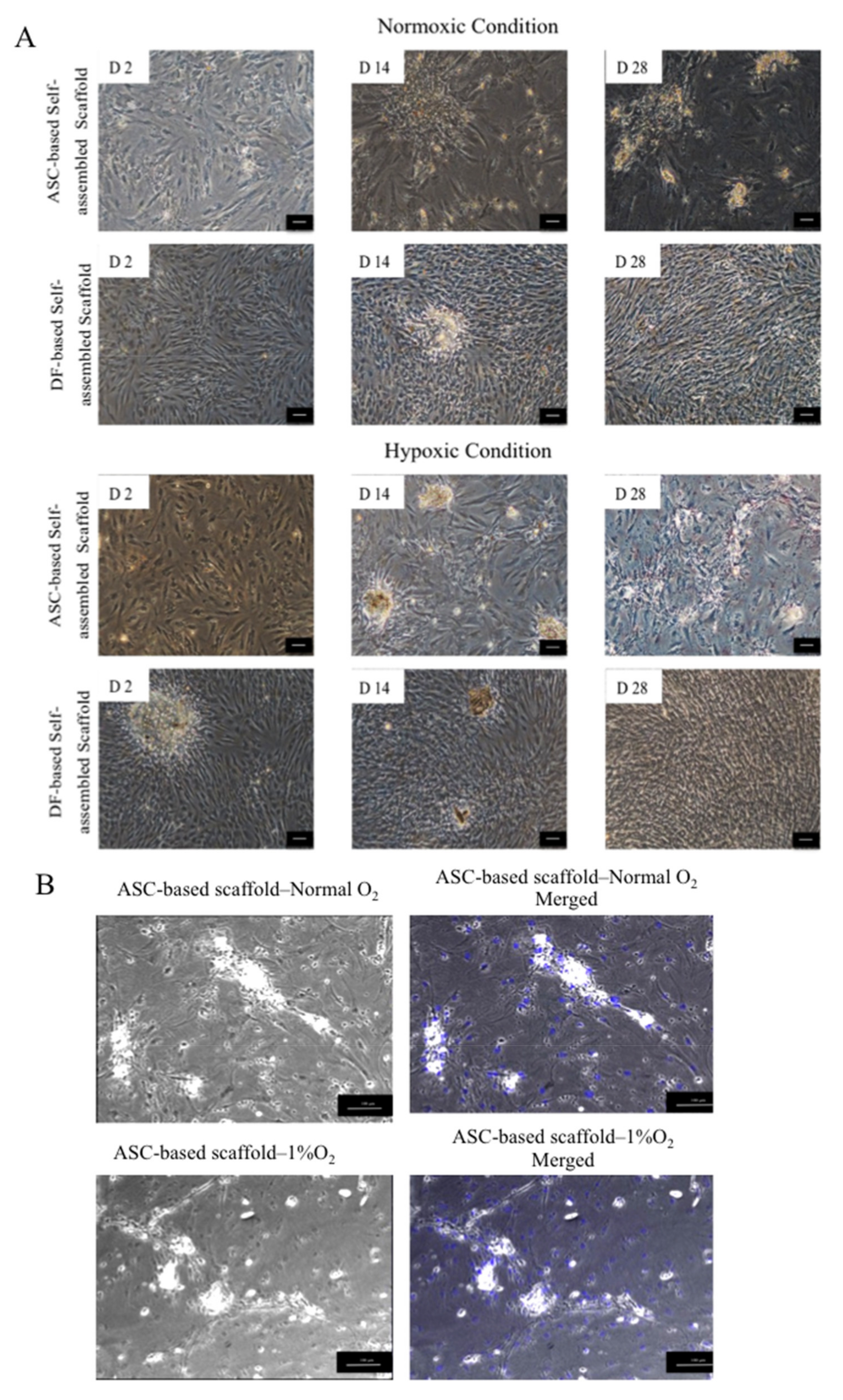
2.3.1. Phase Contrast Imaging
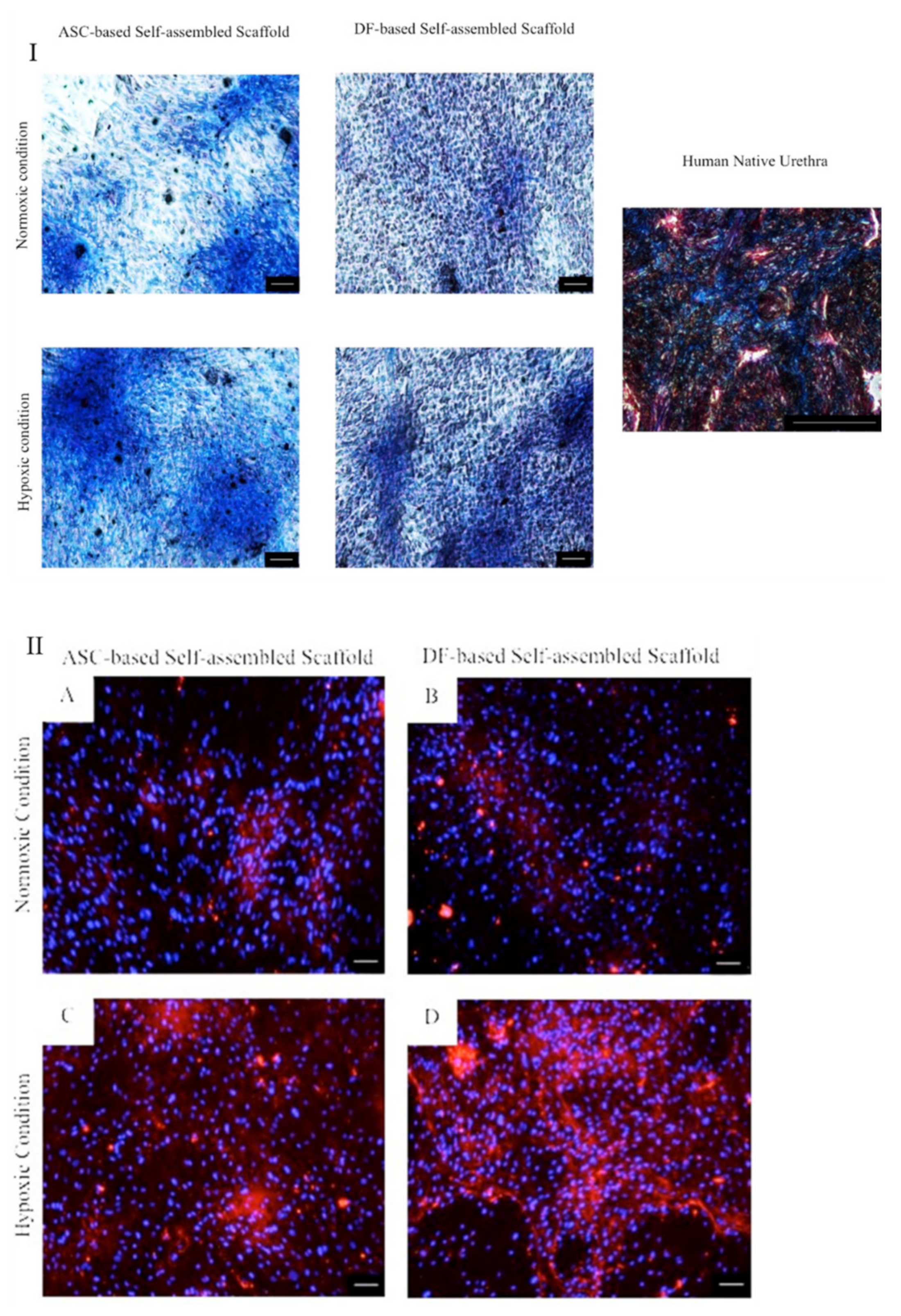
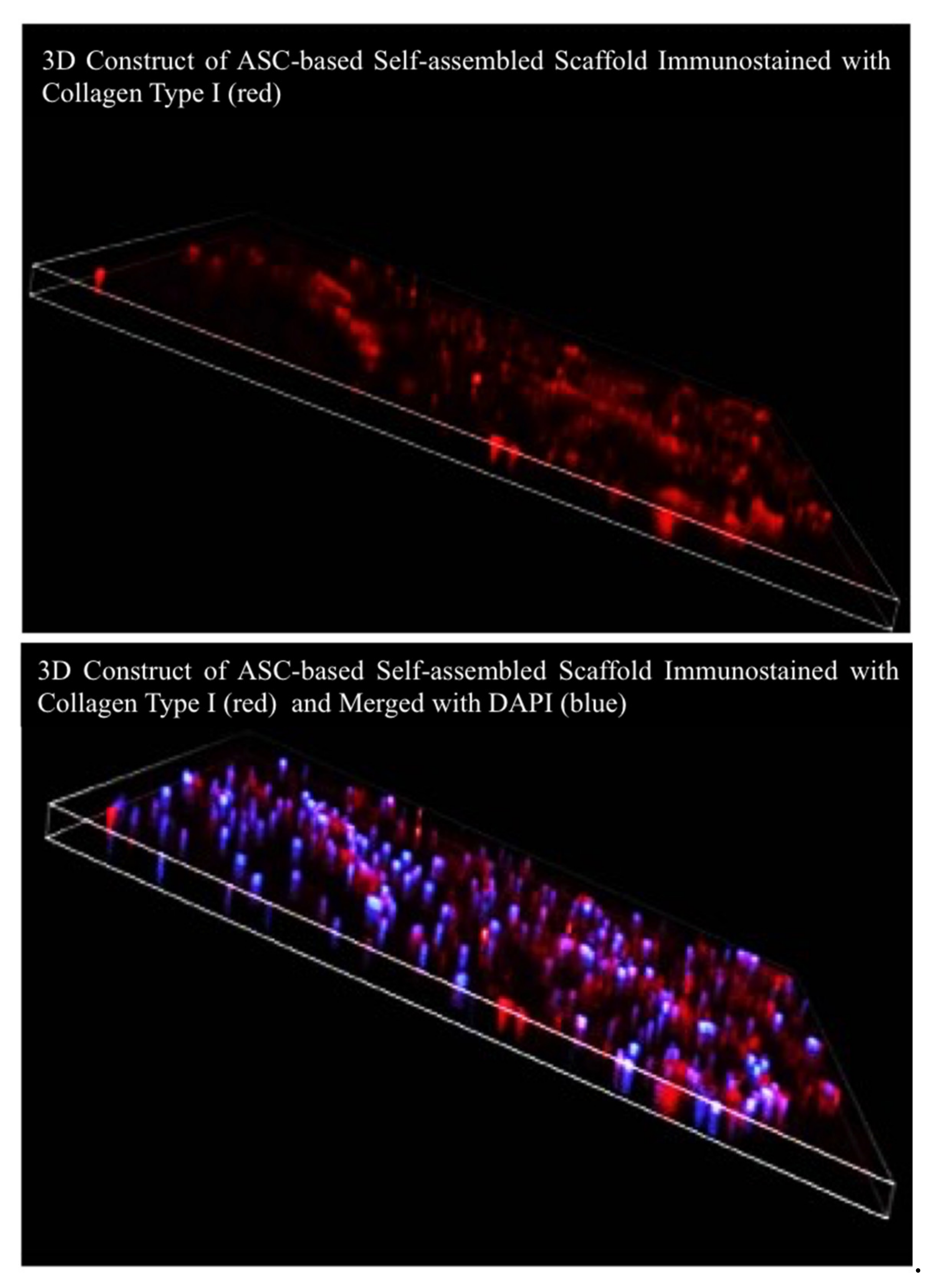
2.3.2. Masson’s Trichrome and Immunocytochemical Staining
2.3.3. Gene Expression Data Analysis
2.4. Biocompatibility of ASC-Based Self-Assembled Scaffold with Urothelial Cells and Smooth Muscle Cells
2.4.1. Cell Morphology Evaluation
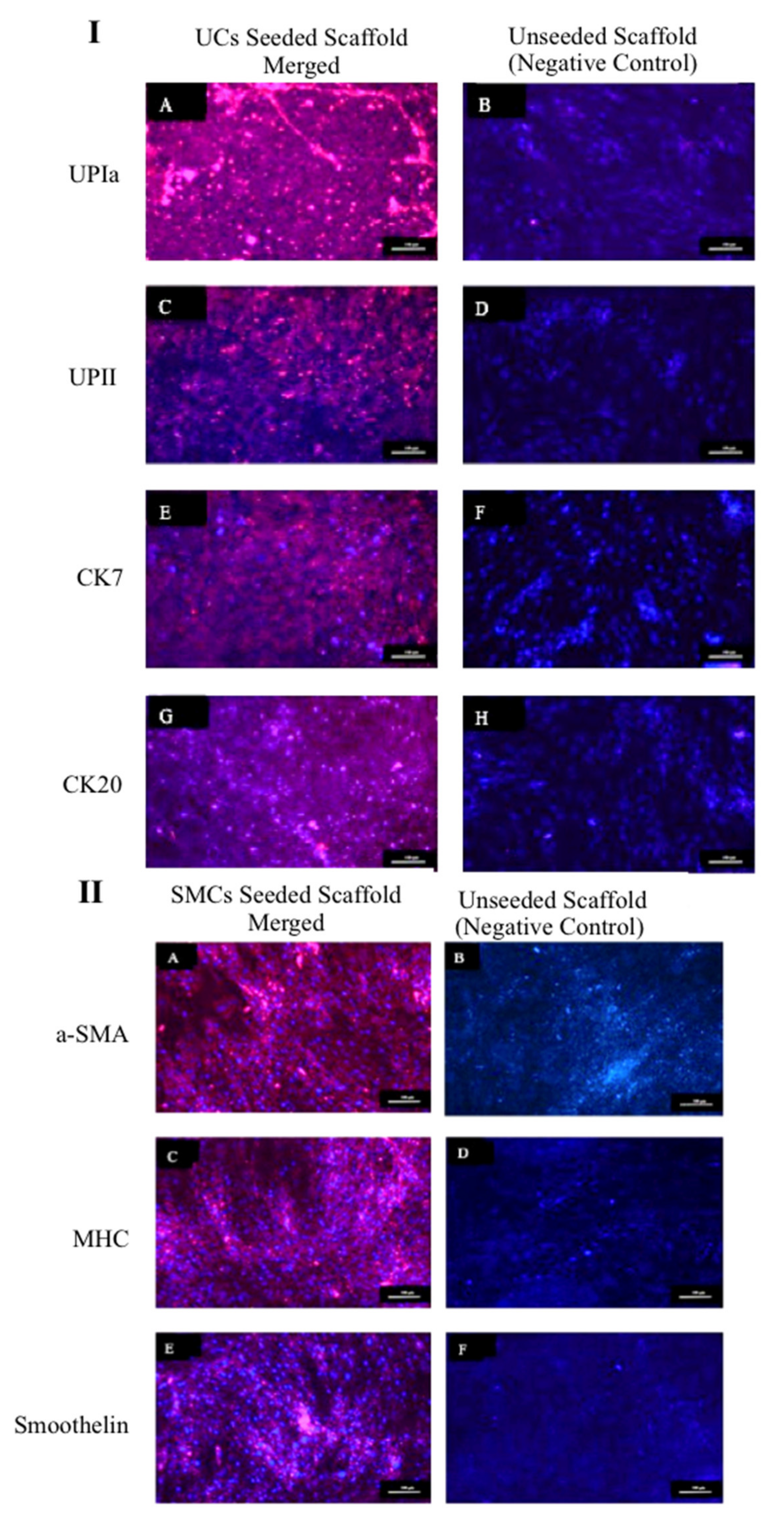
2.4.2. Urothelial and Smooth Muscle Cells Markers Expression
3. Discussion
3.1. Identifying the Ideal Source of Adipose-Derived Stem Cells
3.2. ASC-Based Self-Assembled Scaffold Thickness Optimization
3.2.1. Effect of Cell Seeding Density and Oxygen Concentration on Scaffold Thickness
3.2.2. Effect of Ascorbic Acid Concentration on Scaffold Thickness
3.2.3. Effect of Mechanical Stimulation on Scaffold Thickness
3.3. Characterization of ASC-Based Self-Assembled Scaffold
3.4. Biocompatibility of the ASC-Based Self-Assembled Scaffold with Urothelial and Smooth Muscle Cells
4. Materials and Methods
4.1. Cell Isolation and Culture
4.2. Cell Growth Profile Assessment
4.3. Production of ASC-Based Self-Assembled Scaffold under Different Conditions
4.3.1. Different Seeding Densities and under Normoxic (21% O2) and Hypoxic (1% O2) Culture Conditions
4.3.2. Different Concentrations of Ascorbic Acid
4.3.3. Different Mechanical Stimulation
4.4. Self-Assembled Scaffold Thickness Measurement
4.4.1. Immunocytochemical Staining
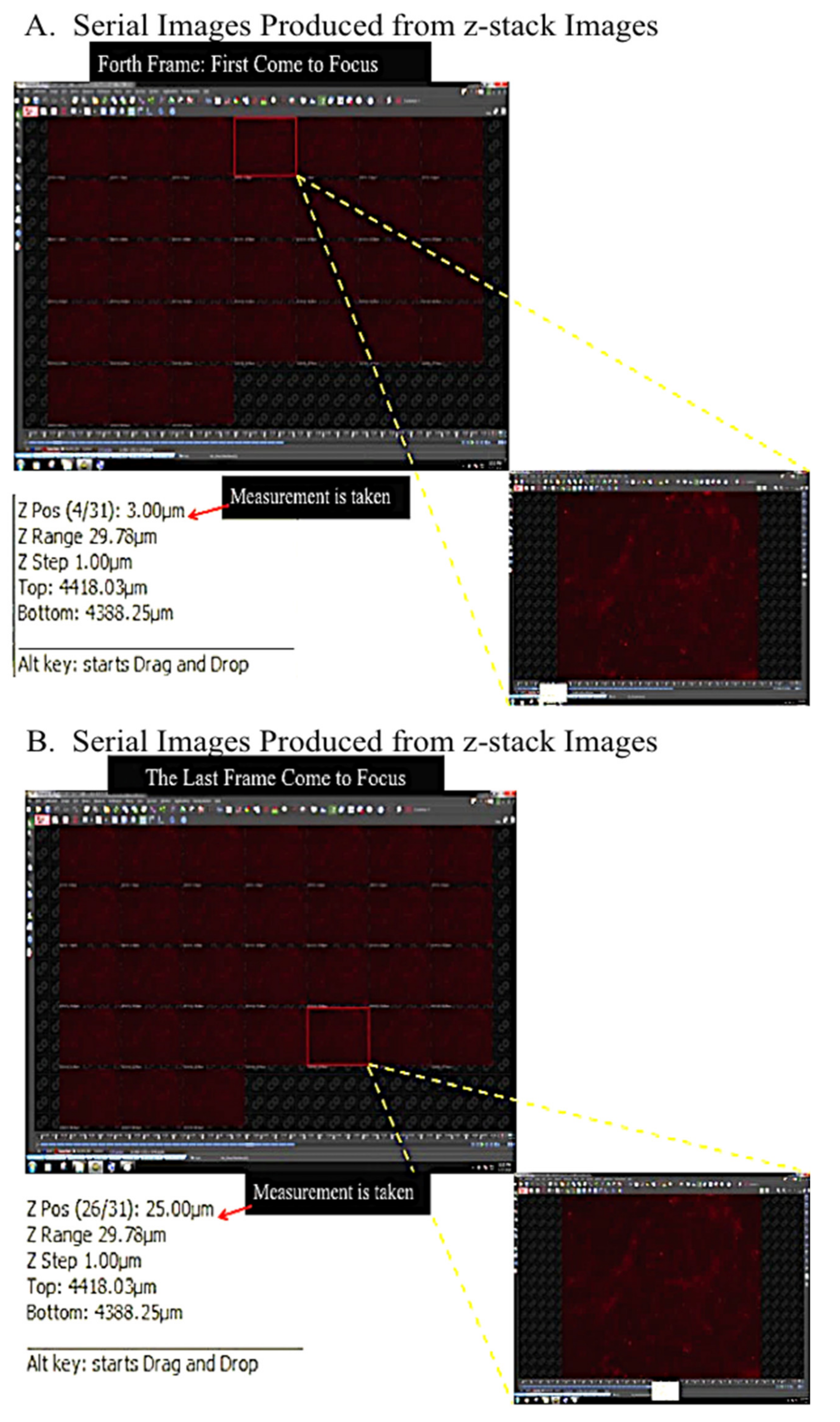
4.4.2. Three-Dimensional Image Reconstruction and Thickness Measurement
4.5. Characterization of ASC-Based Self-Assembled Scaffold
4.5.1. Morphological Characterization
4.5.2. Histological and Immunocytochemical Characterization
4.5.3. Gene Expression Profile
- a.
- Total RNA extraction
- b.
- Complementary DNA synthesis
- c.
- Real-time quantitative polymerase chain reaction
4.6. Biocompatibility Assessment of ASC-Based Self-Assembled Scaffold with Human Urothelial Cells and Smooth Muscle Cells
4.6.1. Growth Pattern Assessment
4.6.2. Urothelial Cells and Smooth Muscle Cells Specific Markers Expression
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Horiguchi, A. Substitution urethroplasty using oral mucosa graft for male anterior urethral stricture disease: Current topics and reviews. Int. J. Urol. 2017, 24, 493–503. [Google Scholar] [CrossRef]
- Rogovaya, O.S.; Fayzulin, A.K.; Vasiliev, A.V.; Kononov, A.V.; Terskikh, V.V. Reconstruction of rabbit urethral epithelium with skin keratinocytes. Acta Nat. 2015, 7, 70–77. [Google Scholar] [CrossRef]
- Mangera, A.; Chapple, C.R. Tissue engineering in urethral reconstruction—An update. Asian J. Androl. 2012, 15, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Orabi, H.; Bouhout, S.; Morissette, A.; Rousseau, A.; Chabaud, S.; Bolduc, S. Tissue engineering of urinary bladder and urethra: Advances from bench to patients. Sci. World J. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favorito, L.A. Urethral stricture: The oldest urologic disease in 2017. Int. Braz. J. Urol. 2017, 43, 4–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashidbenam, Z.; Jasman, M.H.; Hafez, P.; Tan, G.H.; Goh, E.H.; Fam, X.I.; Ho, C.C.; Zainuddin, Z.M.; Rajan, R.; Nor, F.M.; et al. Overview of Urethral Reconstruction by Tissue Engineering: Current Strategies, Clinical Status and Future Direction. Tissue Eng. Regen. Med. 2019, 16, 365–384. [Google Scholar] [CrossRef]
- Raya-Rivera, A.; Esquiliano, D.R.; Yoo, J.J.; Lopez-Bayghen, E.; Soker, S.; Atala, A. Tissue-engineered autologous urethras for patients who need reconstruction: An observational study. Lancet 2011, 377, 1175–1182. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xu, Y.; Xie, H.; Li, C.; Song, L.; Feng, C.; Zhang, Q.; Xie, M.; Wang, Y.; Lv, X. Epithelial-differentiated adipose-derived stem cells seeded bladder acellular matrix grafts for urethral reconstruction: An animal model. Tissue Eng. Part A 2014, 20, 774–784. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Koyanagi-Aoi, M.; Uehara, K.; Hinata, N.; Fujisawa, M.; Aoi, T. Directed differentiation of human induced pluripotent stem cells into mature stratified bladder urothelium. Sci. Rep. 2019, 9, 10506. [Google Scholar] [CrossRef] [Green Version]
- Chun, S.Y.; Kim, B.S.; Kwon, S.Y.; Park, S.I.; Song, P.H.; Yoo, E.S.; Kim, B.W.; Kwon, T.G.; Kim, H.T. Urethroplasty using autologous urethral tissue-embedded acellular porcine bladder submucosa matrix grafts for the management of long-segment urethral stricture in a rabbit model. J. Korean Med. Sci. 2015, 30, 301–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, S.A.; Rashidbenam, Z.; Jasman, M.H.; Ho, C.C.; Sagap, I.; Singh, R.; Yusof, M.R.; Zainuddin, Z.M.; Idrus, R.B.; Ng, M.H. Incorporation of Smooth Muscle Cells Derived from Human Adipose Stem Cells on Poly(Lactic-co-Glycolic Acid) Scaffold for the Reconstruction of Subtotally Resected Urinary Bladder in Athymic Rats. Tissue Eng. Regen. Med. 2020, 17, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Xu, Y.M.; Fu, Q.; Zhu, W.D.; Cui, L. Reconstruction of three dimensional neo-urethra using lingual keratinocytes & corporal smooth muscle cells seeded acellular corporal spongiosum. J. Urol. 2011, 185, e7. [Google Scholar]
- King, N.M.P.; Perrin, J. Ethical issues in stem cell research and therapy. Stem Cell Res. Ther. 2014, 5, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- L’heureux, N.; Paquet, S.; Labbé, R.; Germain, L.; Auger, F.A. A completely biological tissue-engineered human blood vessel. FASEB J. 1998, 12, 47–56. [Google Scholar] [PubMed]
- Vallières, K.; Laterreur, V.; Tondreau, M.Y.; Ruel, J.; Germain, L.; Fradette, J.; Auger, F.A. Human adipose-derived stromal cells for the production of completely autologous self-assembled tissue-engineered vascular substitutes. Acta Biomater. 2015, 24, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Magnan, M.; Lévesque, P.; Gauvin, R.; Dubé, J.; Barrieras, D.; El-Hakim, A.; Bolduc, S. Tissue Engineering of a Genitourinary Tubular Tissue Graft Resistant to Suturing and High Internal Pressures. Tissue Eng. Part. A 2009, 15, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Cattan, V.; Bernard, G.; Rousseau, A.; Bouhout, S.; Chabaud, S.; Auger, F.A.; Bolduc, S. Mechanical stimuli-induced urothelial differentiation in a human tissue-engineered tubular genitourinary graft. Eur. Urol. 2011, 60, 1291–1298. [Google Scholar] [CrossRef]
- Bouhout, S.; Gauvin, R.; Gibot, L.; Aubé, D.; Bolduc, S. Bladder substitute reconstructed in a physiological pressure environment. J. Pediatr. Urol. 2011, 7, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Rousseau, A.; Fradette, J.; Bernard, G.; Gauvin, R.; Laterreur, V.; Bolduc, S. Adipose-derived stromal cells for the reconstruction of a human vesical equivalent. J. Tissue Eng. Regen. Med. 2013, 9, E135–E143. [Google Scholar] [CrossRef]
- Salem, S.A.; Hwie, A.N.; Aminuddin, S.A.; Kong, C.H.; Sagap, I.; Singh, R.; Yusof, M.R.; Zainuddin, Z.M.; Idrus, R.H. Brief Communication Human Adipose Tissue Derived Stem Cells as a Source of Smooth Muscle Cells in the Regeneration of Muscular Layer of Urinary Bladder Wall. Malays. J. Med. Sci. 2013, 20, 80–87. [Google Scholar]
- Zhang, M.; Xu, M.X.; Zhou, Z.; Zhang, K.; Zhou, J.; Zhao, Y.; Wang, Z.; Lu, M.J. The differentiation of human adipose-derived stem cells towards a urothelium-like phenotype in vitro and the dynamic temporal changes of related cytokines by both paracrine and autocrine signal regulation. PLoS ONE 2014, 9, e95583. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Yu, H.; Fan, C.; Kong, Q.; Liu, D.; Meng, L. Differentiate into urothelium and smooth muscle cells from adipose tissue-derived stem cells for ureter reconstruction in a rabbit model. Am. J. Transl. Res. 2016, 8, 3757–3768. [Google Scholar] [PubMed]
- Leto Barone, A.A.; Khalifian, S.; Lee, W.P.; Brandacher, G. Immunomodulatory effects of adipose-derived stem cells: Fact or fiction? Biomed. Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Distler, J.H.; Jüngel, A.; Pileckyte, M.; Zwerina, J.; Michel, B.A.; Gay, R.E.; Kowal-Bielecka, O.; Matucci-Cerinic, M.; Schett, G.; Marti, H.H.; et al. Hypoxia-induced increase in the production of extracellular matrix proteins in systemic sclerosis. Arthritis Rheum. 2007, 56, 4203–4215. [Google Scholar] [CrossRef] [PubMed]
- Baskin, L.S.; Macarak, E.J. Biochemical Characterization and Quantitation of the Collagenous Components of Urethral Stricture Tissue. J. Urol. 1993, 150, 642–647. [Google Scholar] [CrossRef]
- Rashidbenam, Z.; Jasman, M.H.; Rajan, R.; Nor, F.M.; Tan, G.H.; Goh, E.H.; Fam, X.I.; Ho, C.C.; Shuhaili, M.A.; Kosai, N.R.; et al. Hypoxic Preconditioning of Adipose-derive Mesenchymal Stem Cells Upregulates Expression of Extracellular Matrix Proteins. Regen. Res. 2018, 7, 154–156. [Google Scholar]
- Khurana, J.S. Bone Pathology, 2nd ed.; Humana Press: New York, NY, USA, 2009; Available online: https://books.google.com.my/books?id=XlLKuZmxyiAC (accessed on 23 August 2019).
- Kyllönen, L.; Haimi, S.; Mannerström, B.; Huhtala, H.; Rajala, K.M.; Skottman, H.; Sándor, G.K.; Miettinen, S. Effects of different serum conditions on osteogenic differentiation of human adipose stem cells in vitro. Stem Cell Res. Ther. 2013, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Puperi, D.S.; Grande-Allen, K.J.; West, J.L. Ascorbic acid promotes extracellular matrix deposition while preserving valve interstitial cell quiescence within 3D hydrogel scaffolds Yan. J. Tissue Eng. Regen. Med. 2017, 11, 1963–1973. [Google Scholar] [CrossRef]
- Fisher, A.E.; Naughton, D.P. Iron supplements:the quick fix with long-term consequences. Nutr. J. 2004, 3, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrazzo, P.; O’Leary, C. Repositioning natural antioxidants for therapeutic applications in tissue engineering. Bioengineering 2020, 7, 104. [Google Scholar] [CrossRef] [PubMed]
- Sthijns, M.M.J.P.E.; van Blitterswijk, C.A.; LaPointe, V.L.S. Redox regulation in regenerative medicine and tissue engineering: The paradox of oxygen. J. Tissue Eng. Regen. Med. 2018, 12, 2013–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigoni, O.; De Tullio, M.C. Ascorbic acid: Much more than just an antioxidant. Biochim. Biophys. Acta Gen. Subj. 2002, 1569, 1–9. [Google Scholar] [CrossRef]
- Solan, A.; Mitchell, S.; Moses, M.; Niklason, L. Effect of pulse rate on collagen deposition in the tissue-engineered blood vessel. Tissue Eng. 2003, 9, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Elhadj, S.; Mousa, S.A.; Forsten-Williams, K. Chronic pulsatile shear stress impacts synthesis of proteoglycans by endothelial cells: Effect on platelet aggregation and coagulation. J. Cell. Biochem. 2002, 86, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Fortier, G.M.; Gauvin, R.; Proulx, M.; Vallee, M.; Fradette, J. Dynamic culture induces a cell type-dependent response impacting on the thickness of engineered connective tissues. J. Tissue Eng. Regen. Med. 2013, 7, 292–301. [Google Scholar] [CrossRef]
- Da Silva, E.A.; Sampaio, F.J.; Ortiz, V.; Cardoso, L.E. Regional differences in the extracellular matrix of the human spongy urethra as evidenced by the composition of glycosaminoglycans. J. Urol. 2002, 167, 2183–2187. [Google Scholar] [CrossRef]
- Bastos, A.L.; Silva, E.A.; Silva Costa, W.; Sampaio, F.J. The concentration of elastic fibres in the male urethra during human fetal development. BJU Int. 2004, 94, 620–623. [Google Scholar] [CrossRef]
- Lertkiatmongkol, P.; Liao, D.; Mei, H.; Hu, Y.; Newman, P.J. Endothelial functions of PECAM-1 (CD31). Curr. Opin. Hematol. 2016, 23, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Geng, J.G. P-selectin mediates adhesion of leukocytes, platelets, and cancer cells in inflammation, thrombosis, and cancer growth and metastasis. Arch. Immunol. Ther. Exp. 2006, 54, 75–84. [Google Scholar] [CrossRef]
- Zhang, L. The αMß2 integrin and its role in neutrophil function. Cell Res. 1999, 9, 171–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feinberg, T.Y.; Rowe, R.G.; Saunders, T.L.; Weiss, S.J. Functional roles of mmp14 and mmp15 in early postnatal mammary gland development. Development 2016, 143, 3956–3968. [Google Scholar] [CrossRef] [Green Version]
- De Castro, F.; Seal, R.; Maggi, R. ANOS1: A unified nomenclature for Kallmann syndrome 1 gene (KAL1) and anosmin-1. Brief. Funct. Genom. 2017, 16, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Kreft, M.E.; Kreft, M.; Jezernik, K. Endocytotic activity of bladder superficial urothelial cells is inversely related to their differentiation stage. Differentiation 2009, 77, 48–59. [Google Scholar] [CrossRef]
- Calderon, R.O.; Glocker, M.; Eynard, A.R. Lipid and fatty acid composition of different fractions from rat urinary transitional epithelium. Lipids 1998, 33, 1017–1022. [Google Scholar] [CrossRef]
- Jones, R.; Jacobson, M.; Steudel, W. α-Smooth-Muscle Actin and Microvascular Precursor Smooth-Muscle Cells in Pulmonary Hypertension. Am. J. Respir. Cell Mol. Biol. 1999, 20, 582–594. [Google Scholar] [CrossRef]
- Owens, G.K. Regulation of differentiation of vascular smooth muscle cells. Physiol. Rev. 1995, 75, 487–517. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zeng, G.; Qi, Y.; Tang, X.; Zhang, J.; Wu, Z.; Liang, J.; Shi, L.; Liu, H.; Zhang, P. Xenotransplantation of human adipose-derived stem cells in zebrafish embryos. PLoS ONE 2015, 10, e0123264. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, S.; Pontecorvi, P.; Anastasiadou, E.; Napoli, C.; Marchese, C. Immunomodulatory Effect of Adipose-Derived Stem Cells: The Cutting Edge of Clinical Application. Front. Cell Dev. Biol. 2020, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fauzi, M.B.; Rashidbenam, Z.; Bin Saim, A.; Binti Hj Idrus, R. Preliminary study of in vitro three-dimensional skin model using an ovine collagen type i sponge seeded with co-culture skin cells: Submerged versus air-liquid interface conditions. Polymers 2020, 12, 2784. [Google Scholar] [CrossRef]
- Maarof, M.; Lokanathan, Y.; Ruszymah, I.; Saim, A.; Chowdhury, S.R. Proteomic Analysis of Human Dermal Fibroblast Conditioned Medium (DFCM). Protein J. 2018, 37, 589–607. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Symbol | Gene Name | Fold Change | Fold Regulation |
|---|---|---|---|---|
| Genes Upregulated in ASC-Based Self-Assembled Scaffold vs. DF-Based Self-Assembled Scaffold | ||||
| A01 | ADAMTS1 | ADAM metallopeptidase with thrombospondin type 1 motif, 1 | 9.84 | 9.84 |
| A02 | ADAMTS13 | ADAM metallopeptidase with thrombospondin type 1 motif, 13 | 66.87 | 66.87 |
| A03 | ADAMTS8 | ADAM metallopeptidase with thrombospondin type 1 motif, 8 | 21.55 | 21.55 |
| A04 | CD44 | CD44 molecule (Indian blood group) | 13.88 | 13.88 |
| A05 | CDH1 | Cadherin 1, type 1, E-cadherin (epithelial) | 2.82 | 2.82 |
| A06 | CLEC3B | C-type lectin domain family 3, member B | 42.51 | 42.51 |
| A10 | COL14A1 | Collagen, type XIV, alpha 1 | 34.77 | 34.77 |
| A12 | COL16A1 | Collagen, type XVI, alpha 1 | 1.67 | 1.67 |
| B02 | COL4A2 | Collagen, type IV, alpha 2 | 5.51 | 5.51 |
| B03 | COL5A1 | Collagen, type V, alpha 1 | 2.19 | 2.19 |
| B06 | COL7A1 | Collagen, type VII, alpha 1 | 8.05 | 8.05 |
| B09 | CTNNA1 | Catenin (cadherin-associated protein), alpha 1, 102kDa | 8.05 | 8.05 |
| B10 | CTNNB1 | Catenin (cadherin-associated protein), beta 1, 88kDa | 2.36 | 2.36 |
| B11 | CTNND1 | Catenin (cadherin-associated protein), delta 1 | 2.18 | 2.18 |
| B12 | CTNND2 | Catenin (cadherin-associated protein), delta 2 (neural plakophilin-related arm-repeat protein) | 1323.36 | 1323.36 |
| C03 | HAS1 | Hyaluronan synthase 1 | 494.55 | 494.55 |
| C05 | ITGA1 | Integrin, alpha 1 | 1.52 | 1. 52 |
| C06 | ITGA2 | Integrin, alpha 2 (CD49B, alpha 2 subunit of VLA-2 receptor) | 14.38 | 14.38 |
| C07 | ITGA3 | Integrin, alpha 3 (antigen CD49C, alpha 3 subunit of VLA-3 receptor) | 1.26 | 1.26 |
| C11 | ITGA7 | Integrin, alpha 7 | 37.44 | 37.44 |
| D05 | ITGB2 | Integrin, beta 2 (complement component 3 receptor 3 and 4 subunit) | 31.26 | 31.26 |
| D07 | ITGB4 | Integrin, beta 4 | 8.67 | 8.67 |
| D09 | ANOS1 | Kallmann syndrome 1 sequence | 72.84 | 72.84 |
| D10 | LAMA1 | Laminin, alpha 1 | 1.27 | 1.27 |
| D11 | LAMA2 | Laminin, alpha 2 | 1.93 | 1.93 |
| D12 | LAMA3 | Laminin, alpha 3 | 2.89 | 2.89 |
| E02 | LAMB3 | Laminin, beta 3 | 4.42 | 4.42 |
| E09 | MMP14 | Matrix metallopeptidase 14 (membrane-inserted) | 1.45 | 1.45 |
| E10 | MMP15 | Matrix metallopeptidase 15 (membrane-inserted) | 1.11 | 1.11 |
| F04 | MMP9 | Matrix metallopeptidase 9 (gelatinase B, 92kDa gelatinase, 92kDa type IV collagenase) | 5.40 | 5.40 |
| F06 | PECAM1 | Platelet/endothelial cell adhesion molecule | 160.52 | 160.52 |
| F07 | SELE | Selectin E | 144.50 | 144.50 |
| F08 | SELL | Selectin L | 17.2 | 17.2 |
| F09 | SELP | Selectin P (granule membrane protein 140kDa, antigen CD62) | 7.2 | 7.2 |
| F12 | SPG7 | Spastic paraplegia 7 (pure and complicated autosomal recessive) | 238.85 | 238.85 |
| G01 | SPP1 | Secreted phosphoprotein 1 | 3.69 | 3.69 |
| G03 | THBS1 | Thrombospondin 1 | 1.22 | 1.22 |
| G04 | THBS2 | Thrombospondin 2 | 5.85 | 5.85 |
| G05 | THBS3 | Thrombospondin 3 | 82.90 | 82.90 |
| G07 | TIMP2 | TIMP metallopeptidase inhibitor 2 | 2.66 | 2.66 |
| G10 | VCAM1 | Vascular cell adhesion molecule 1 | 6.42 | 6.42 |
| G11 | VCAN | Versican | 1.95 | 1.95 |
| G12 | VTN | Vitronectin | 29.65 | 29.65 |
| Total | 43 | |||
| Genes Downregulated in ASC-Based Self-Assembled Scaffold vs. DF-Based Self-Assembled Scaffold | ||||
| A07 | CNTN1 | Contactin 1 | 0.93 | −1.06 |
| A08 | COL11A1 | Collagen, type XI, alpha 1 | 0.37 | −2.70 |
| A09 | COL12A1 | Collagen, type XII, alpha 1 | 0.09 | −10.35 |
| A11 | COL15A1 | Collagen, type XV, alpha 1 | 0.27 | −3.61 |
| B01 | COL1A1 | Collagen, type I, alpha 1 | 0.20 | −4.81 |
| B04 | COL6A1 | Collagen, type VI, alpha 1 | 0.52 | −1.90 |
| B05 | COL6A2 | Collagen, type VI, alpha 2 | 0.29 | −3.40 |
| B07 | COL8A1 | Collagen, type VIII, alpha 1 | 0.41 | −2.38 |
| B08 | CTGF | Connective tissue growth factor | 0.01 | −70.93 |
| C01 | ECM1 | Extracellular matrix protein 1 | 0.12 | −8.11 |
| C02 | FN1 | Fibronectin 1 | 0.05 | −17.56 |
| C04 | ICAM1 | Intercellular adhesion molecule 1 | 0.60 | −1.66 |
| C08 | ITGA4 | Integrin, alpha 4 (antigen CD49D, alpha 4 subunit of VLA-4 receptor) | 0.44 | −2.26 |
| C09 | ITGA5 | Integrin, alpha 5 (fibronectin receptor, alpha polypeptide) | 0.10 | −9.71 |
| C10 | ITGA6 | Integrin, alpha 6 | 0.57 | −1.72 |
| C12 | ITGA8 | Integrin, alpha 8 | 0.45 | −2.19 |
| D01 | ITGAL | Integrin, alpha L (antigen CD11A (p180), lymphocyte function-associated antigen 1; alpha polypeptide) | 0.37 | −2.64 |
| D02 | ITGAM | Integrin, alpha M (complement component 3 receptor 3 subunit) | 0.002 | −384.89 |
| D03 | ITGAV | Integrin, alpha V (vitronectin receptor, alpha polypeptide, antigen CD51) | 0.74 | −1.33 |
| D04 | ITGB1 | Integrin, beta 1 (fibronectin receptor, beta polypeptide, antigen CD29 includes MDF2, MSK12) | 0.42 | −2.36 |
| D06 | ITGB3 | Integrin, beta 3 (platelet glycoprotein IIIa, antigen CD61) | 0.08 | −12.46 |
| D08 | ITGB5 | Integrin, beta 5 | 0.22 | −4.51 |
| E01 | LAMB1 | Laminin, beta 1 | 0.04 | −20.11 |
| E03 | LAMC1 | Laminin, gamma 1 (formerly LAMB2) | 0.35 | −2.80 |
| E04 | MMP1 | Matrix metallopeptidase 1 (interstitial collagenase) | 0.36 | −2.75 |
| E05 | MMP10 | Matrix metallopeptidase 10 (stromelysin 2) | 0.36 | −2.71 |
| E06 | MMP11 | Matrix metallopeptidase 11 (stromelysin 3) | 0.03 | −27.09 |
| E07 | MMP12 | Matrix metallopeptidase 12 (macrophage elastase) | 0.06 | −15.41 |
| E08 | MMP13 | Matrix metallopeptidase 13 (collagenase 3) | 0.73 | −1.35 |
| E11 | MMP16 | Matrix metallopeptidase 16 (membrane-inserted) | 0.12 | −8.03 |
| E12 | MMP2 | Matrix metallopeptidase 2 (gelatinase A, 72kDa gelatinase, 72kDa type IV collagenase) | 0.45 | −2.20 |
| F01 | MMP3 | Matrix metallopeptidase 3 (stromelysin 1, progelatinase) | 0.16 | −5.99 |
| F02 | MMP7 | Matrix metallopeptidase 7 (matrilysin, uterine) | 0.003 | −273.10 |
| F03 | MMP8 | Matrix metallopeptidase 8 (neutrophil collagenase) | 0.12 | −8.03 |
| F05 | NCAM1 | Neural cell adhesion molecule 1 | 0.42 | −2.33 |
| F10 | SGCE | Sarcoglycan, epsilon | 0.31 | −3.16 |
| F11 | SPARC | Secreted protein, acidic, cysteine-rich (osteonectin) | 0.67 | −1.49 |
| G02 | TGFBI | Transforming growth factor, beta-induced, 68kDa | 0.05 | −18.80 |
| G06 | TIMP1 | TIMP metallopeptidase inhibitor 1 | 0.23 | −4.31 |
| G08 | TIMP3 | TIMP metallopeptidase inhibitor 3 | 0.34 | −2.86 |
| G09 | TNC | Tenascin C | 0.67 | −1.47 |
| Total | 41 | |||
| Position | Symbol | Gene Name | p Value | |
|---|---|---|---|---|
| ASC-Based Self-Assembled Scaffold | ||||
| Not Expressed (Ct ≥ 35) | E08 | MMP13 | Matrix metallopeptidase 13 (collagenase 3) | 0.1179 |
| F02 | MMP7 | Matrix metallopeptidase 7 (matrilysin, uterine) | 0.4480 | |
| Total | 2 | |||
| Weakly Expressed (33 ≤ Ct < 35) | D02 | ITGAM | Integrin, alpha M (complement component 3 receptor 3 subunit) | 0.1161 |
| E10 | MMP15 | Matrix metallopeptidase 15 (membrane-inserted) | 0.3720 | |
| F09 | SELP | Selectin P (granule membrane protein 140kDa, antigen CD62) | 0.3630 | |
| Total | 3 | |||
| DF-Based Self-Assembled Scaffold | ||||
| Not Expressed (Ct≥35) | A02 | ADAMTS13 | ADAM metallopeptidase with thrombospondin type 1 motif, 13 | 0.3750 |
| A03 | ADAMTS8 | ADAM metallopeptidase with thrombospondin type 1 motif, 8 | 0.4810 | |
| B12 | CTNND2 | Catenin (cadherin-associated protein), delta 2 (neural plakophilin-related arm-repeat protein) | 0.1145 | |
| C03 | HAS1 | Hyaluronan synthase 1 | 0.3334 | |
| E08 | MMP13 | Matrix metallopeptidase 13 (collagenase 3) | 0.1179 | |
| F06 | PECAM1 | Platelet/endothelial cell adhesion molecule | 0.0003 | |
| F07 | SELE | Selectin E | 0.2760 | |
| F08 | SELL | Selectin L | ||
| F09 | SELP | Selectin P (granule membrane protein 140kDa, antigen CD62) | 0.4390 | |
| Total | 9 | |||
| Weakly Expressed (33 ≤ Ct < 35) | D09 | ANOS1 | Kallmann syndrome 1 sequence | 0.3661 |
| Total | 1 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rashidbenam, Z.; Jasman, M.H.; Tan, G.H.; Goh, E.H.; Fam, X.I.; Ho, C.C.K.; Zainuddin, Z.M.; Rajan, R.; Rani, R.A.; Nor, F.M.; et al. Fabrication of Adipose-Derived Stem Cell-Based Self-Assembled Scaffold under Hypoxia and Mechanical Stimulation for Urethral Tissue Engineering. Int. J. Mol. Sci. 2021, 22, 3350. https://doi.org/10.3390/ijms22073350
Rashidbenam Z, Jasman MH, Tan GH, Goh EH, Fam XI, Ho CCK, Zainuddin ZM, Rajan R, Rani RA, Nor FM, et al. Fabrication of Adipose-Derived Stem Cell-Based Self-Assembled Scaffold under Hypoxia and Mechanical Stimulation for Urethral Tissue Engineering. International Journal of Molecular Sciences. 2021; 22(7):3350. https://doi.org/10.3390/ijms22073350
Chicago/Turabian StyleRashidbenam, Zahra, Mohd Hafidzul Jasman, Guan Hee Tan, Eng Hong Goh, Xeng Inn Fam, Christopher Chee Kong Ho, Zulkifli Md Zainuddin, Reynu Rajan, Rizal Abdul Rani, Fatimah Mohd Nor, and et al. 2021. "Fabrication of Adipose-Derived Stem Cell-Based Self-Assembled Scaffold under Hypoxia and Mechanical Stimulation for Urethral Tissue Engineering" International Journal of Molecular Sciences 22, no. 7: 3350. https://doi.org/10.3390/ijms22073350