
Hesperidin Promotes Osteogenesis and Modulates Collagen Matrix Organization and Mineralization In Vitro and In Vivo

 , and
, and 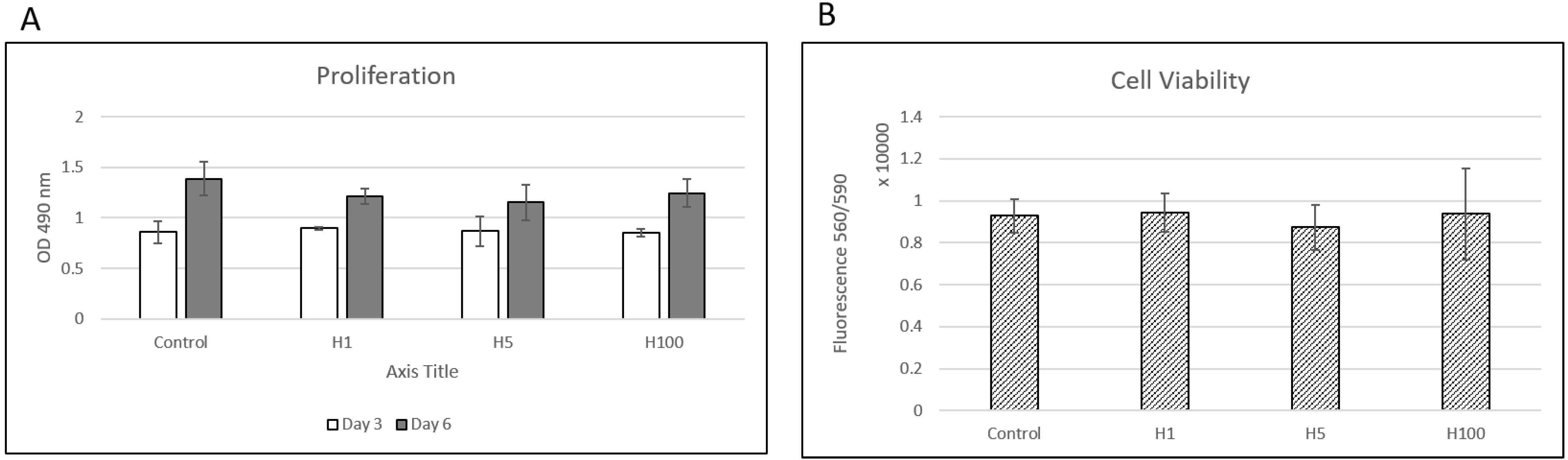{kind=link}
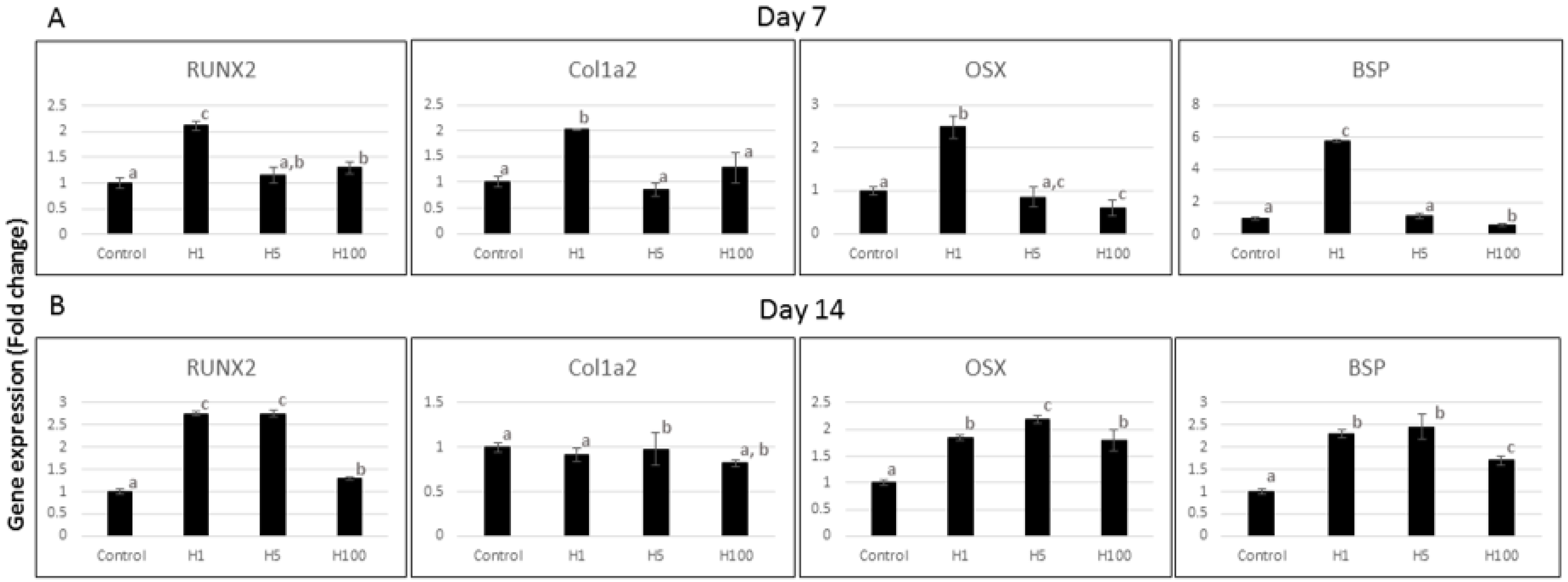{kind=link}
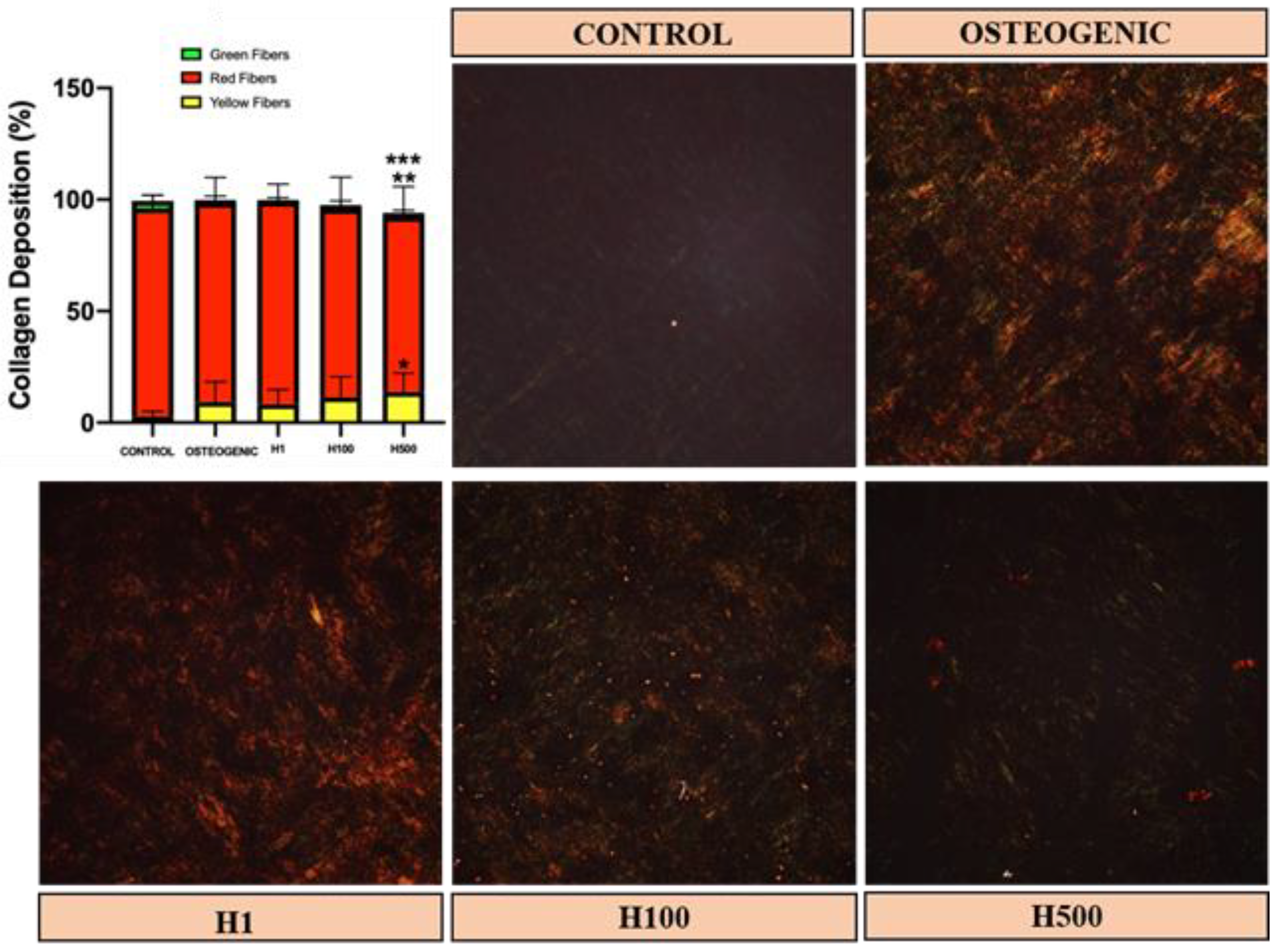{kind=link}
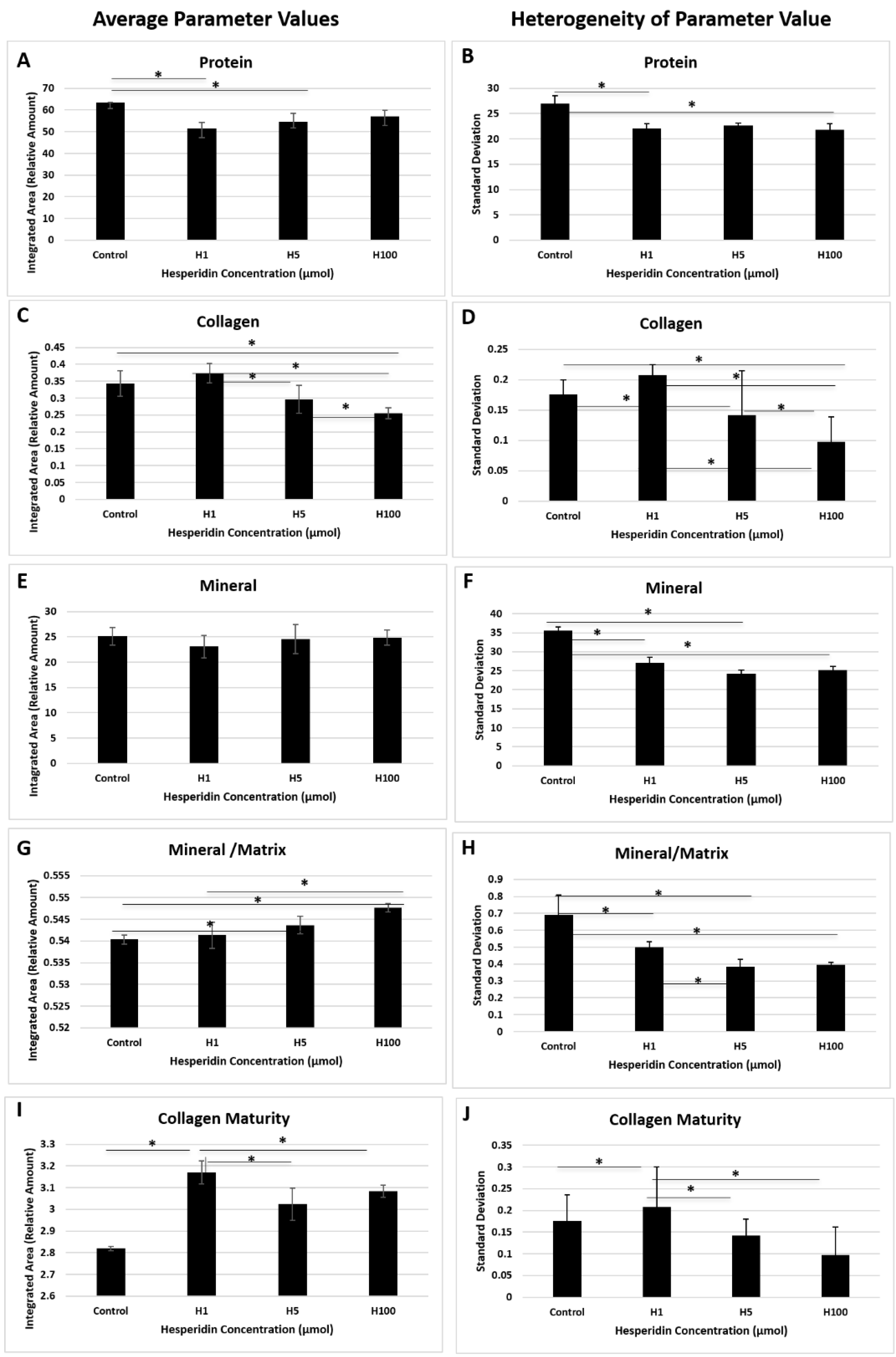{kind=link}
{kind=link}
Abstract
:1. Introduction
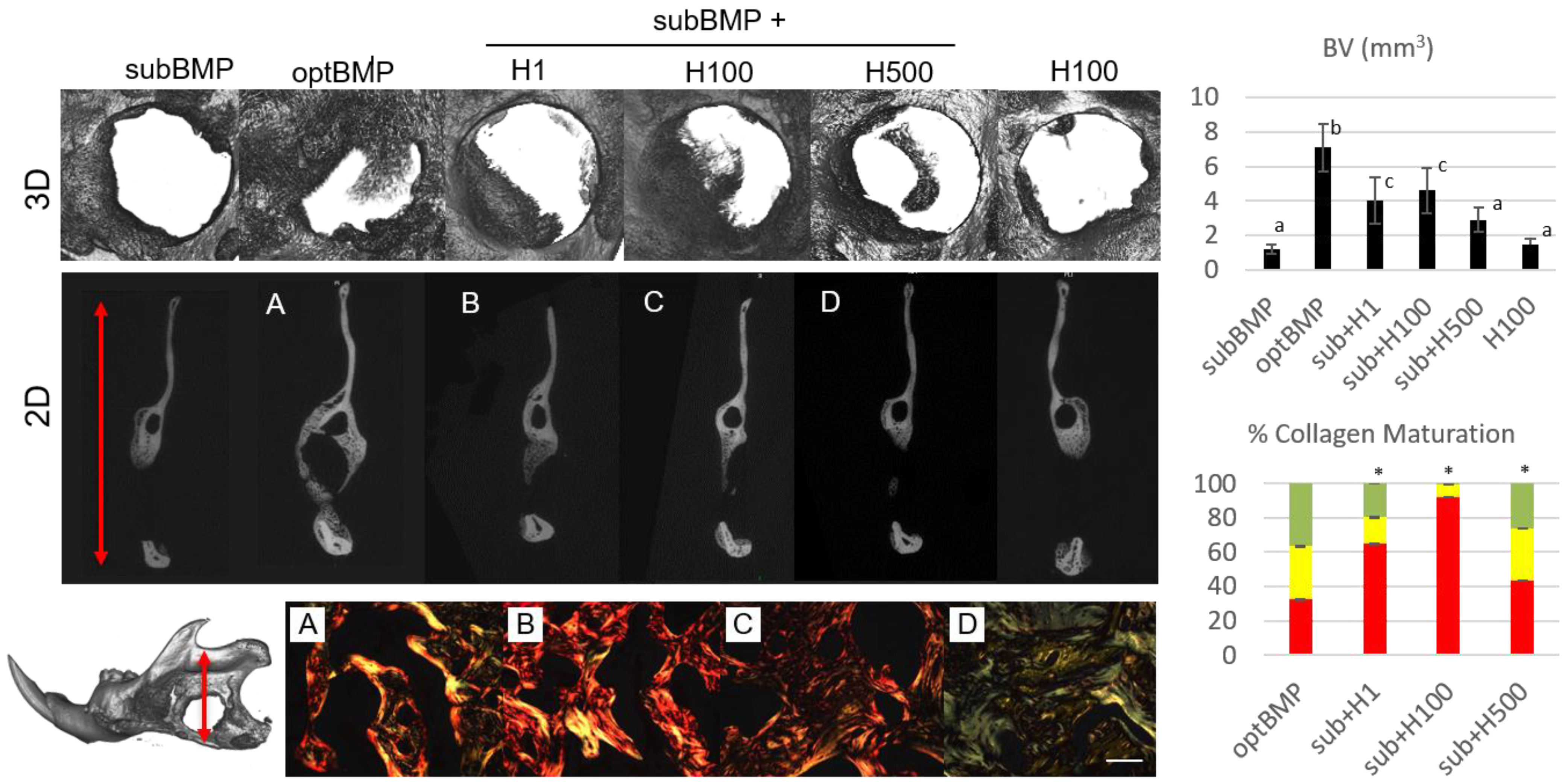
2. Results and Discussion
3. Materials and Methods
3.1. Cell Culture, Proliferation and Viability
3.2. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
3.3. Collagen Organization/Maturation In Vitro by Picrosirius Red (PSR) Staining
3.4. Analysis of Mineral and Protein by Fourier-Transform Infrared Spectroscopy Imaging (FT-IR)
3.5. Mandible Model and Post-Surgery Analyses
3.6. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bauer, T.W.; Muschler, G.F. Bone graft materials. An overview of the basic science. Clin. Orthop. Relat. Res. 2000, 371, 10–27. [Google Scholar] [CrossRef]
- Bhumiratana, S.; Vunjak-Novakovic, G. Concise Review: Personalized Human Bone Grafts for Reconstructing Head and Face. STEM CELLS Transl. Med. 2011, 1, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Kao, D.W.K.; Kubota, A.; Nevins, M.; Fiorellini, J.P. The negative effect of combining rhBMP-2 and Bio-Oss on bone formation for maxillary sinus augmentation. Int. J. Periodontics Restor. Dent. 2012, 32, 61–67. [Google Scholar]
- Nevins, M.; Camelo, M.; Nevins, M.L.; Schenk, R.K.; Lynch, S.E. Periodontal Regeneration in Humans Using Recombinant Human Platelet- Derived Growth Factor-BB (rhPDGF-BB) and Allogenic Bone. J. Periodontol. 2003, 74, 1282–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, M.P.; Kakarla, U.K.; Porter, R.W.; Sonntag, V.K. Formation of Painful Seroma and Edema After the Use of Recombinant Human Bone Morphogenetic Protein-2 in Posterolateral Lumbar Spine Fusions. Neurosurgery 2010, 66, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Selvig, K.A.; Sorensen, R.G.; Wozney, J.M.; Wikesjö, U.M. Bone Repair Following Recombinant Human Bone Morphogenetic Protein-2 Stimulated Periodontal Regeneration. J. Periodontol. 2002, 73, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Zara, J.N.; Siu, R.K.; Zhang, X.; Shen, J.; Ngo, R.; Lee, M.; Li, W.; Chiang, M.; Chung, J.; Kwak, J.; et al. High Doses of Bone Morphogenetic Protein 2 Induce Structurally Abnormal Bone and Inflammation In Vivo. Tissue Eng. Part A 2011, 17, 1389–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, T.C.; Black, L.I.; Stussman, B.J.; Barnes, P.M.; Nahin, R.L. Trends in the use of complementary health approaches among adults: United States, 2002–2012. Natl. Health Stat. Rep. 2015, 29, 178–179. [Google Scholar]
- Bagchi, D.; Sen, C.K.; Ray, S.D.; Das, D.K.; Bagchi, M.; Preuss, H.G.; Vinson, J.A. Molecular mechanisms of cardioprotection by a novel grape seed proanthocyanidin extract. Mutat. Res. Mol. Mech. Mutagen. 2003, 523–524, 87–97. [Google Scholar] [CrossRef]
- Jain, M.; Parmar, H.S. Evaluation of antioxidative and anti-inflammatory potential of hesperidin and naringin on the rat air pouch model of inflammation. Inflamm. Res. 2010, 60, 483–491. [Google Scholar] [CrossRef]
- Neto, C.C. Cranberry and Its Phytochemicals: A Review of In Vitro Anticancer Studies. J. Nutr. 2007, 137, 186S–193S. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Blumberg, J.B. Phytochemical composition of nuts. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. 1), 329–332. [Google Scholar] [PubMed]
- Walter, R.; Miguez, P.A.; Arnold, R.R.; Pereira, P.N.R.; Duarte, W.R.; Yamauchi, M. Effects of Natural Cross-Linkers on the Stability of Dentin Collagen and the Inhibition of Root Caries in vitro. Caries Res. 2008, 42, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Xie, Q.; Bedran-Russo, A.K.; Pan, S.; Ling, J.; Wu, C.D. The preventive effect of grape seed extract on artificial enamel caries progression in a microbial biofilm-induced caries model. J. Dent. 2014, 42, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.H.; Hsu, C.W.; Chang, W.-T.; Waypa, G.B.; Li, J.; Li, D.; Li, C.Q.; Anderson, T.; Qin, Y.; Schumacker, P.T.; et al. Cytotoxicity induced by grape seed proanthocyanidins: Role of nitric oxide. Cell Biol. Toxicol. 2006, 22, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Trzeciakiewicz, A.; Habauzit, V.; Mercier, S.; Lebecque, P.; Davicco, M.-J.; Coxam, V.; Demigne, C.; Horcajada, M.-N. Hesperetin stimulates differentiation of primary rat osteoblasts involving the BMP signalling pathway. J. Nutr. Biochem. 2010, 21, 424–431. [Google Scholar] [CrossRef]
- Habauzit, V.; Sacco, S.M.; Gil-Izquierdo, A.; Trzeciakiewicz, A.; Morand, C.; Barron, D.; Pinaud, S.; Offord, E.; Horcajada, M.-N. Differential effects of two citrus flavanones on bone quality in senescent male rats in relation to their bioavailability and metabolism. Bone 2011, 49, 1108–1116. [Google Scholar] [CrossRef]
- Horcajada, M.N.; Habauzit, V.; Trzeciakiewicz, A.; Morand, C.; Gil-Izquierdo, A.; Mardon, J.; Lebecque, P.; Davicco, M.J.; Chee, W.S.S.; Coxam, V.; et al. Hesperidin inhibits ovariectomized-induced osteopenia and shows differential effects on bone mass and strength in young and adult intact rats. J. Appl. Physiol. 2008, 104, 648–654. [Google Scholar] [CrossRef] [Green Version]
- Habauzit, V.; Nielsen, I.-L.; Gil-Izquierdo, A.; Trzeciakiewicz, A.; Morand, C.; Chee, W.; Barron, D.; Lebecque, P.; Davicco, M.-J.; Williamson, G.; et al. Increased bioavailability of hesperetin-7-glucoside compared with hesperidin results in more efficient prevention of bone loss in adult ovariectomised rats. Br. J. Nutr. 2009, 102, 976–984. [Google Scholar] [CrossRef] [Green Version]
- Chiba, H.; Kim, H.; Matsumoto, A.; Akiyama, S.; Ishimi, Y.; Suzuki, K.; Uehara, M. Hesperidin Prevents Androgen Deficiency-induced Bone Loss in Male Mice. Phytother. Res. 2014, 28, 289–295. [Google Scholar] [CrossRef]
- Souza, J.J.M.; Tuin, S.A.; Robinson, A.G.; De Souza, J.G.O.; Bianchini, M.A.; Miguez, P.A. Effect of Flavonoid Supplementation on Alveolar Bone Healing—A Randomized Pilot Trial. Dent. J. 2020, 8, 86. [Google Scholar] [CrossRef]
- Shahi, M.; Peymani, A.; Sahmani, M. Regulation of Bone Metabolism. Rep. Biochem. Mol. Biol. 2017, 5, 73–82. [Google Scholar] [PubMed]
- Nakashima, K.; Zhou, X.; Kunkel, G.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The Novel Zinc Finger-Containing Transcription Factor Osterix Is Required for Osteoblast Differentiation and Bone Formation. Cell 2002, 108, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Xue, D.; Chen, E.; Zhang, W.; Gao, X.; Wang, S.; Zheng, Q.; Pan, Z.; Li, H.; Liu, L. The role of hesperetin on osteogenesis of human mesenchymal stem cells and its function in bone regeneration. Oncotarget 2017, 8, 21031–21043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piérard, G.E. Sirius Red Polarization Method is Useful to Visualize the Organization of Connective Tissues but not the Molecular Composition of their Fibrous Polymers. Matrix 1989, 9, 68–71. [Google Scholar] [CrossRef]
- Smith, L.R.; Barton, E.R. Collagen content does not alter the passive mechanical properties of fibrotic skeletal muscle inmdxmice. Am. J. Physiol. Physiol. 2014, 306, C889–C898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraishi, N.; Maruno, T.; Tochio, N.; Sono, R.; Otsuki, M.; Takatsuka, T.; Tagami, J.; Kobayashi, Y. Hesperidin interaction to collagen detected by physico-chemical techniques. Dent. Mater. 2017, 33, 33–42. [Google Scholar] [CrossRef]
- Van Strijp, A.J.P.; Takatsuka, T.; Sono, R.; Iijima, Y. Inhibition of dentine collagen degradation by hesperidin: An in situ study. Eur. J. Oral Sci. 2015, 123, 447–452. [Google Scholar] [CrossRef]
- Hiraishi, N.; Sono, R.; Sofiqul, I.; Yiu, C.; Nakamura, H.; Otsuki, M.; Takatsuka, T.; Tagami, J. In vitro evaluation of plant-derived agents to preserve dentin collagen. Dent. Mater. 2013, 29, 1048–1054. [Google Scholar] [CrossRef]
- Islam, S.M.; Hiraishi, N.; Nassar, M.; Sono, R.; Otsuki, M.; Takatsura, T.; Yiu, C.; Tagami, J. In vitro effect of hesperidin on root dentin collagen and de/re-mineralization. Dent. Mater. J. 2012, 31, 362–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arosarena, O.; Collins, W. Comparison of BMP-2 and -4 for rat mandibular bone regeneration at various doses. Orthod. Craniofacial Res. 2005, 8, 267–276. [Google Scholar] [CrossRef]
- Arosarena, O.A.; Collins, W.L. Bone Regeneration in the Rat Mandible with Bone Morphogenetic Protein-2: A Comparison of Two Carriers. Otolaryngol. Neck Surg. 2005, 132, 592–597. [Google Scholar] [CrossRef]
- Miguez, P.; Terajima, M.; Nagaoka, H.; Mochida, Y.; Yamauchi, M. Role of glycosaminoglycans of biglycan in BMP-2 signaling. Biochem. Biophys. Res. Commun. 2011, 405, 262–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miguez, P.; Terajima, M.; Nagaoka, H.; Ferreira, J.; Braswell, K.; Ko, C.; Yamauchi, M. Recombinant Biglycan Promotes Bone Morphogenetic Protein-induced Osteogenesis. J. Dent. Res. 2014, 93, 406–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreira, A.; Lojudice, F.; Halcsik, E.; Navarro, R.; Sogayar, M.; Granjeiro, J. Bone Morphogenetic Proteins. J. Dent. Res. 2014, 93, 335–345. [Google Scholar] [CrossRef]
- Kuzu, M.; Kandemir, F.M.; Yıldırım, S.; Çağlayan, C.; Küçükler, S. Attenuation of sodium arsenite-induced cardiotoxicity and neurotoxicity with the antioxidant, anti-inflammatory, and antiapoptotic effects of hesperidin. Environ. Sci. Pollut. Res. 2021, 28, 10818–10831. [Google Scholar] [CrossRef] [PubMed]
- Kaku, M.; Mochida, Y.; Atsawasuwan, P.; Parisuthiman, D.; Yamauchi, M. Post-translational modifications of collagen upon BMP-induced osteoblast differentiation. Biochem. Biophys. Res. Commun. 2007, 359, 463–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisuthiman, D.; Mochida, Y.; Duarte, W.R.; Yamauchi, M. Biglycan Modulates Osteoblast Differentiation and Matrix Mineralization. J. Bone Miner. Res. 2005, 20, 1878–1886. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mochida, Y.; Parisuthiman, D.; Pornprasertsuk-Damrongsri, S.; Atsawasuwan, P.; Sricholpech, M.; Boskey, A.L.; Yamauchi, M. Decorin modulates collagen matrix assembly and mineralization. Matrix Biol. 2009, 28, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongwattanapisan, P.; Terajima, M.; Miguez, P.A.; Querido, W.; Nagaoka, H.; Sumida, N.; Gurysh, E.G.; Ainslie, K.M.; Pleshko, N.; Perera, L.; et al. Identification of the effector domain of biglycan that facilitates BMP-2 osteogenic function. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pleshko, N.; Boskey, A.; Mendelsohn, R. Novel infrared spectroscopic method for the determination of crystallinity of hydroxyapatite minerals. Biophys. J. 1991, 60, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Khanarian, N.T.; Boushell, M.K.; Spalazzi, J.P.; Pleshko, N.; Boskey, A.L.; Lu, H.H. FTIR-I Compositional Mapping of the Cartilage-to-Bone Interface as a Function of Tissue Region and Age. J. Bone Miner. Res. 2014, 29, 2643–2652. [Google Scholar] [CrossRef]
- Faillace, M.E.; Phipps, R.J.; Miller, L.M. Fourier Transform Infrared Imaging as a Tool to Chemically and Spatially Characterize Matrix-Mineral Deposition in Osteoblasts. Calcif. Tissue Int. 2012, 92, 50–58. [Google Scholar] [CrossRef]
- Boskey, A.; Camacho, N.P. FT-IR imaging of native and tissue-engineered bone and cartilage. Biomaterials 2007, 28, 2465–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zellin, G.; Linde, A. Importance of delivery systems for growth-stimulatory factors in combination with osteopromotive membranes. An experimental study using rhBMP-2 in rat mandibular defects. J. Biomed. Mater. Res. 1997, 35, 181–190. [Google Scholar] [CrossRef]
- Arosarena, O.A.; Falk, A.; Malmgren, L.; Bookman, L.; Allen, M.J.; Schoonmaker, J.; Tatum, S.; Kellman, R. Defect Repair in the Rat Mandible With Bone Morphogenic Proteins and Marrow Cells. Arch. Facial Plast. Surg. 2003, 5, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, N.; Nagaoka, H.; Yamauchi, S.; Teixeira, F.B.; Miguez, P.; Yamauchi, M. Immunohistological Characterization of Newly Formed Tissues after Regenerative Procedure in Immature Dog Teeth. J. Endod. 2011, 37, 1636–1641. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miguez, P.A.; Tuin, S.A.; Robinson, A.G.; Belcher, J.; Jongwattanapisan, P.; Perley, K.; de Paiva Gonҫalves, V.; Hanifi, A.; Pleshko, N.; Barton, E.R. Hesperidin Promotes Osteogenesis and Modulates Collagen Matrix Organization and Mineralization In Vitro and In Vivo. Int. J. Mol. Sci. 2021, 22, 3223. https://doi.org/10.3390/ijms22063223
Miguez PA, Tuin SA, Robinson AG, Belcher J, Jongwattanapisan P, Perley K, de Paiva Gonҫalves V, Hanifi A, Pleshko N, Barton ER. Hesperidin Promotes Osteogenesis and Modulates Collagen Matrix Organization and Mineralization In Vitro and In Vivo. International Journal of Molecular Sciences. 2021; 22(6):3223. https://doi.org/10.3390/ijms22063223
Chicago/Turabian StyleMiguez, Patricia A., Stephen A. Tuin, Adam G. Robinson, Joyce Belcher, Prapaporn Jongwattanapisan, Kimberly Perley, Vinicius de Paiva Gonҫalves, Arash Hanifi, Nancy Pleshko, and Elisabeth R. Barton. 2021. "Hesperidin Promotes Osteogenesis and Modulates Collagen Matrix Organization and Mineralization In Vitro and In Vivo" International Journal of Molecular Sciences 22, no. 6: 3223. https://doi.org/10.3390/ijms22063223