
Advancing Drug Discovery for Neurological Disorders Using iPSC-Derived Neural Organoids



1
Dino Ferrari Centre, Department of Pathophysiology and Transplantation (DEPT), Neuroscience Section, University of Milan, 20122 Milan, Italy
2
IRCCS Foundation Ca’ Granda Ospedale Maggiore Policlinico, Neurology Unit, Via Francesco Sforza 35, 20122 Milan, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(5), 2659; https://doi.org/10.3390/ijms22052659
Submission received: 20 February 2021
/
Revised: 1 March 2021
/
Accepted: 3 March 2021
/
Published: 6 March 2021
(This article belongs to the Special Issue Drug Discovery and Development 2.0)
Abstract
:In the last decade, different research groups in the academic setting have developed induced pluripotent stem cell-based protocols to generate three-dimensional, multicellular, neural organoids. Their use to model brain biology, early neural development, and human diseases has provided new insights into the pathophysiology of neuropsychiatric and neurological disorders, including microcephaly, autism, Parkinson’s disease, and Alzheimer’s disease. However, the adoption of organoid technology for large-scale drug screening in the industry has been hampered by challenges with reproducibility, scalability, and translatability to human disease. Potential technical solutions to expand their use in drug discovery pipelines include Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) to create isogenic models, single-cell RNA sequencing to characterize the model at a cellular level, and machine learning to analyze complex data sets. In addition, high-content imaging, automated liquid handling, and standardized assays represent other valuable tools toward this goal. Though several open issues still hamper the full implementation of the organoid technology outside academia, rapid progress in this field will help to prompt its translation toward large-scale drug screening for neurological disorders.
1. Introduction
Organoids are stem cell-derived, three-dimensional (3D) cultures that are artificially generated. Organoids contain different cell types that self-organize through cell-sorting and spatially restricted lineage commitment, similarly to in vivo organs [1]. Different cell types have been used to generate organoids in vitro, including primary cultured cells from human tissues, embryonic stem cells (ESCs), and induced pluripotent stem cells (iPSCs) [2]. iPSC cultures have provided invaluable information for modeling neurological and neuromuscular disorders [3,4]. However, organoids present some advantages over traditional two-dimensional cultures. They exhibit near-physiologic cellular composition, and they can grow extensively in culture while maintaining genomic stability [5,6], making them potentially valuable for high-throughput screenings [7]. Compared with animal models, organoids can reduce experimental complexity and allow the study of human development features that may be difficult to investigate in animal models. Examples of 3D cultures recapitulating human organs in vitro include the peripheral nerve, [8] the spinal cord [9], and the brain [1]. These neural organoids proved to be useful platforms to model neurodevelopmental, neuropsychiatric [10], and neurodegenerative disorders [11], such as microcephaly [12], Miller-Dieker Syndrome [13], and Alzheimer’s disease [14].
Animal models represent the gold standard for in vivo modeling in drug development and their widespread adoption across studies has prompted invaluable progress in drug discovery (Figure 1).
The conventional drug discovery pipeline begins with compound screening and target discovery. Then, the efficacy and toxicity of drug candidates are validated in animal models. Based on the results of preclinical studies, clinical trials enable drug testing in humans. Finally, successful drugs are released onto the market (upper row). IPSC-based preclinical studies provide a model with human-relevant genetic backgrounds. In addition, iPSCs enable the generation of 3D cultures with physiologically improved modeling of disease-relevant phenotypes, ameliorating patients’ stratification. These culture systems can be coupled with bioengineering solutions, including organoids, organs (such as neuromuscular junction, NMJ), and a blood-brain barrier (BBB) on-a-chip (lower row), to provide more reliable data for clinical trials and final regulatory approval.
Unfortunately, many therapeutics successful in preclinical model trials fail in late-stage clinical studies due to reproducibility issues between animal models and humans. IPSC-derived organoids permit in vitro and in vivo investigations, representing a relevant innovation to recapitulate the physiological mechanisms of human organs [15]. Particularly, organoid technology could better mimic aspects of human physiology when compared with animal models [1]. Drugs (such as PTC-124 and Ataluren) that were successful in non-neurological animal models were not efficacious in human intestinal organoids modeling cystic fibrosis. These results turned out to be accurate in two-phase clinical studies [16], suggesting that organoids might have the potential to bridge preclinical and clinical trials [17]. However, some challenges still hamper the full application of organoid technology in drug discovery. For instance, the intra-organoid and inter-organoid cellular heterogeneity, the limited scalability, the lack of reproducibility across protocols, and the variable degree of maturity still represent important roadblocks.
Some recent technological advances could overcome these open issues. The advent of CRISPR-Cas9 (Clustered Regularly Interspaced Short Palindromic Repeats/Cas9) gene-editing tools represent a game-changer, opening new avenues in organoid research. Similarly, single-cell RNA (scRNAseq) sequencing could help to delve into the biology of neural organoids at a subcellular level. The adoption of automated systems, machine-learning-based strategies, and high-content imaging (HCI) tools in the generation and analysis of organoids could speed-up the processing of data derived from 3D cultures, potentially enabling the translation of neural organoids from the academic to the industrial setting.
Here, we focus on the early use of neural organoids in the industry, the current challenges, and the possible technical solutions that might support their implementation into drug discovery pipelines.
2. Neural Organoids for Drug Discovery in the Industry
The demand for commercial organoids will likely increase in the next few years [18]. In 2019, six companies were already active in the field, whereas at least 19 companies were interested in the business in 2020 [18]. Some of these (e.g., Organome and Hubrecht Organoid Technology) are involved in biobanking, while others are dedicated to manufacturing and commercializing organoids as well as organoid-related technologies. Particularly, different companies show an interest in developing renal, gut, cancer, and neural organoids [18].
Most of the early studies on neural organoids have been conducted in academic settings. However, in the last few years, the interest in neural organoids in the industry has raised, leading to industry-academia collaborations [19].
For instance, Novartis-based and Harvard-based groups collaborated to generate neural organoids to model Zika virus infection [20]. CRISPR-based approaches helped the researchers to knock out the putative entry receptor and to identify viral entry receptors. These organoids displayed an elevated number of viral copies within neural progenitors and exhibited increased cell death, leading to a severe microcephaly-like pattern resembling the infection in vivo [20]. This protocol set the groundwork for a successful collaboration with academia in defining the possible role of the viral attachment factor Tyrosine-protein kinase receptor UFO (AXL) as a potential drug target [19,20].
Stemonix is a company providing iPSC-derived micro-brain organoids for drug screening [21]. Though it is not clear how the process is standardized and reproducible, the company aims to provide neural organoids as a service for disease modeling and drug discovery. Particularly, the focus would be to use this technology as a supplement to the standard drug discovery process. Stemonix has planned to apply organoid technology to discover potential therapeutic approaches for the Rett syndrome [22].
System 1 biosciences is a company dedicated to neurotherapeutic development. They aim to adopt artificial intelligence and robotics-based phenotypic screening on iPSC-derived neural organoids for drug discovery in schizophrenia, autism, and epilepsy [23]. Particularly, they intend to integrate machine learning and high-scale dimensional phenotype analytics to create a comprehensive molecular description of the brain and to reveal paths to new disease-modifying therapies.
A:head bio represents an example of a mixed academia-industry collaboration between a newly formed start-up and the Institute of Molecular Biotechnology (IBMA) in Vienna, Austria. This collaboration aims to develop new therapeutics within central nervous system (CNS) disorders, focusing on Dravet syndrome (a severe epileptic syndrome due to a loss-of-function mutation in the sodium channel SCN1A) [24]. Similarly, in 2006, STEMCELL Technologies and the IBMA signed an agreement to commercialize cerebral organoid culture technologies [25].
Overall, the technology of neural organoids is still in its early stages. While this system may bring some value to the drug discovery pipeline, its use in the industry is limited to recent start-up companies or small teams in larger biotech companies.
3. Challenges for the Adoption of Neural Organoids for Drug Discovery
Basic research has benefited from the use of neural organoids, particularly in the field of disease modeling and target identification in the context of neuropsychiatric and neurological disorders [10] (Table 1).
Unfortunately, some obstacles still hamper the implementation of neural factors for large-scale drug screening in the industry. Their degree of heterogeneity and maturity as well their reproducibility and scalability represent some of these hurdles.
3.1. Heterogeneity
Sample heterogeneity refers to the inherent variability within biological samples. In the context of large-scale studies—such as in drug discovery research—scientists aim to reduce this variability to reach conclusive and generalizable results. Similar to primary patient samples, both environmental and genetic factors represent potential confounders in neural organoids. The two most important sources of variability include iPSC lines and organoid batch-to-batch variation [42]. The former refers to the variable ability of different iPSC lines to generate relevant architectures and cell types observed in their in vivo counterparts. The latter variability depends on the multitude of reagents and molecules as well as the lack of standardized protocols across experiments used to generate organoids. In addition to this, organoids exhibit a high degree of heterogeneity even within a single batch, reducing the comparability among samples [43,44].
A recent attempt to reduce brain organoid heterogeneity involved the generation of midbrain-like organoids modeling Parkinson’s disease-associated phenotypes using primitive neural stem cells (NSCs). Ha et al. generated NSC-derived organoids in two weeks using either an unspecific or a midbrain-specific medium. These 3D cultures exhibited homogenous cytoarchitectural and transcriptional features, as confirmed with immunochemistry and single-cell analysis [45]. Disease phenotypes were responsive to disease-specific therapies, supporting the potential role of this approach to large-scale disease modeling and drug testing. Similarly, Nickels et al. have recently optimized the generation of midbrain organoids by modifying the timing of maturation, the patterning strategy, and the starting number and type of cells [46]. Particularly, the optimization of timing and the use of a more committed cell line consistently increased the reproducibility of the model, reducing the variation among batches while maintaining the cellular complexity of midbrain organoids.
3.2. Scalability
The scalability of the organoid technology represents another challenge. When used for drug screening, the assays are generally performed in 384 or 1536-well plates, leading to adequate control of the study conditions to explore the vast chemical space. However, organoids are typically generated in 96-well plates, growing too big for 1536-well plates, and contain too many cells to be properly fed using 100 μL of media in 384-well plates. The implementation of spinning bioreactors to feed a higher number of organoids has partially overcome this roadblock [47,48], but limitations regarding drug screening remain. The adoption of engineering solutions for micro-fluidic and milli-fluidic systems, automated liquid handling (see “Automation and high-throughput screening”), and robotic manipulations may overcome this issue, enhancing scalability for drug screening approaches.
3.3. Reproducibility
Available protocols typically use Matrigel, which is the gelatinous protein mixture secreted by mouse sarcoma cells, as an extracellular matrix (ECM)-like structure to support proper organoid formation [1]. Physiologically, different cell types contribute to ECM formation and maintenance, including microglia and vascular endothelial cells, which may lack in current neural organoids. Though Matrigel is extensively adopted, its composition is generic and largely uncharacterized, increasing variability and lacking the physiologically-relevant biochemical signals driving organoid generation.
Some recent developments have tried to recreate a more physiological microenvironment for organoids and to increase their reproducibility. For instance, the use of ECM derived from fetal porcine brains combined with silk scaffolding improves the functional maturation of neural populations, better supporting their long-term growth [49]. In another case, Lancaster et al. used polylactic-co-glycolic acid (PLGA) molecules, which are synthetic biocompatible microfilaments, as a basement membrane-like structure for self-patterning organoids, generating the microfilament-engineered cerebral organoids (enCORs) [50]. This scaffolding supported the cytoarchitectural organization of enCORs, improving radial organization and highlighting the role of tailored structures in improving organoid development. Different studies have applied bioprinting technologies to support regulated, cellular patterning at specific locations within the cultures, providing a spatial control of cell development [51,52], thus, increasing the reproducibility of organoids during differentiation.
3.4. Maturity
The degree of maturity may limit the potential of neural organoids to model adult-onset disease in vitro. Organoids may lack neuro-endothelial-glial-immune inter-lineage signaling if compared with their in vivo counterparts [42]. In addition to this, spontaneous vascularization lacks neural organoids, leading to aberrant maturation. As a result, late-stage organoids have inadequate nutrient delivery, accumulate waste, and develop central necrotic cores. This process may have two consequences. First, the hypoxia-induced release of necrosis-associated cytokines may lead to paracrine effects disrupting the function of neighboring vital cells, ultimately, impairing the sample analysis. Second, the limited diffusion of nutrients may account for the lack of stereotypical anatomy in neural organoids [53].
Integrating a functional vascular network within neural organoids is crucial to accurately model later stages of brain maturity. Different groups have adopted various methods toward this goal. Some laboratories have generated vascularized neural organoids using patients’ iPSC-derived endothelial cells [54] or by in-vivo transplantation in mice [55], creating the chimeras [56]. Ham et al. treated early brain organoids with vascular endothelial growth factor (VEGF) and WNT7b to favor the differentiation of endothelial and pericyte-like cells without altering neuronal populations [57]. This protocol led to the generation of open-circle vascular structures containing CD31-positive cells and to the modulation of gene networks associated with brain embryogenesis in neuronal populations, suggesting a cross-talk between neurons and endothelial cells [57]. Wörsdörfer et al. applied a co-culture method using mesodermal progenitors integrated into brain organoid cultures, generating a vessel network with a blood vessel-like ultrastructure after eight weeks in vitro [58].
Vascularized organoids display a blood-brain barrier (BBB)-like characteristics [59]—such as expression of tight junctions, nutrient transporters, and trans-endothelial electrical resistance—offering invaluable advancements for drug discovery pipelines, as the BBB represents a key limiting factor for drugs targeting the CNS. Other organoid-based studies showed the possibility of generating complex BBB models containing various cell types, including pericytes, astrocytes, microglia, and oligodendrocytes, which are susceptible to functional assessment testing and potentially valuable for toxicity testing and drug discovery [60,61].
Other approaches to bypass diffusion limitations include the generation of sliced neocortical organoids (SNs), allowing improved oxygenation into organoid cores and promoting long-term neurogenesis, sustained progenitor expansion, and separation of cortical layers [62]. Similarly, Giandomenico et al. have cultured sliced, cortical organoids at the air/liquid interface [63]. These organoids exhibited improved neuronal survival compared with whole organoids as well as mature morphological and functional features, such as long-range axons. These axons could elicit coordinated muscle contractions in co-cultured mouse spinal cord–muscle explants.
The time needed to reach organoid maturity may represent another roadblock to widespread adoption. Currently, though novel protocols are available [47,64], most cerebral organoids might require months [1] to reach full development. Innovative strategies to speed up growth or processing [65] might facilitate their use for drug discovery.
In the following section, we will discuss the possible technical solutions to improve the implementation of neural organoids in drug discovery. These include CRISPR-Cas-9 to generate isogenic models and facilitate disease modeling of genetic disorders, advanced imaging, and single-cell sequencing approaches to characterize the diseased phenotypes more accurately, automated systems to accelerate the production, and machine learning to speed up the analysis of these complex data sets. Engineering solutions such as automated liquid handling and optimized plate systems represent other important approaches as well.
4. Overcoming Challenges with Possible Technical Solutions
4.1. CRISPR
CRISPR/Cas 9 (Clustered Regularly Interspaced Short Palindromic Repeats/Cas9) technology has evolved the field of functional genomics, enabling the generation of isogenic iPSC lines at an unprecedented speed, precision, and flexibility. This has enabled the interrogation of the role of a specific mutation in the same genetic background. The CRISPR/Cas9 system is based on a guide RNA to target the nuclease Cas9 to a specific genome locus, causing a double-strand break and activating DNA repair mechanisms, such as homology-directed repair (HDR) and non-homologous end joining (NHEJ) repair pathways [66] (Figure 2).
Methods harnessing the CRISPR-Cas9 technology in organoids involve dissociation into single cells. Then, liposomal transfection, electroporation, and lentiviruses can be used to genetically engineer organoids. Non-homologous end joining (NHEJ) facilitates gene knock-out, while homology-directed repair (HDR) enables gene knock-in. Finally, single organoid clones can be selected and expanded to obtain isogenic cell populations.
NHEJ represents the gold standard to create gene knockouts by CRISPR. Cas cleavage followed by NHEJ-mediated repair at the break site is repeated until an error—usually a small indel—occurs. As shown in seminal studies on mice, these indels are valuable to introduce frameshift mutations for the generation of gene knockouts [67,68].
Considering in vitro disease modeling, CRISPR-Cas9 is typically performed on neural organoid founder cells, such as ESCs and iPSCs [13,69,70]. Then, neural organoids can be grown from selected cells carrying the mutation, with the advantage that all forming cells contain the desired genetic change. Alternative methods to introduce CRISPR/Cas9-mediated genetic changes in growing organoids exist. Plasmid vectors containing Cas9 and one or more guide RNAs [71] can be electroporated in early organoids, which are known as embryoid bodies, inducing loss-of-function mutations, as seen in organoid models of CNS tumors [34]. Another example includes the combination of CRISPR-Cas9-directed tumor suppressor (TP53) knock-out with oncogene Kirsten rat sarcoma (KRAS) knock-in in a four-month-old neural organoid-based glioblastoma model [72]. In this case, the proliferative potential of several glioblastoma-like cells is sufficient to provide a persistent genetic modification within the organoids.
Though CRISPR has not been used for high-throughput screening using neural organoids yet, this technology has provided new tools for disease modeling. Some examples include neuropsychiatric disorders, such as Miller-Dieker Syndrome [13,73], autism [74], macrocephaly [70], and neurological diseases, such as tumors, including glioblastoma [72] and retinoblastoma [69], viral infections, Zika Virus (ZV) [19], and neurodegenerative disorders, including Alzheimer’s disease and frontotemporal dementia [14,75]. Particularly, gene editing on neural organoids infected with ZV has allowed the study of ZV entry receptors. This system relied on a doxycycline-inducible Cas9 and the introduction of a single gRNA via lentivirus, promoting the efficient knock-out of the AXL receptor, demonstrating that this receptor was unnecessary to drive ZV infection [19].
CRISPR-Cas technology applied to model neuro-ophthalmic conditions has driven an important advancement in the field. This technology used in patient-derived iPSCs [76] has paved the way to a recent CRISPR-based clinical trial in patients with Leber congenital amaurosis (NCT03872479) and congenital retinal dystrophy leading to severe vision loss at an early age. A construct containing an adenovirus vector AAV5 with dual single-guide RNA (sgRNA) mediated the gene correction of the IVS26 cryptic-splice mutation in Centrosomal Protein 290 (CEP 290) in the retina [77]. Correction of retinitis pigmentosa GTPase regulator (RPGR) mutations in iPSCs followed by the generation of retinal organoids proved to be a valuable approach for testing new potential therapies in retinitis pigmentosa [78]. In parallel to therapy testing, neuroophthalmological disease modeling profited from CRISPR-Cas as well. The introduction of the E50K mutation in the optineurin (OPTN) gene enabled the modeling of glaucoma in retinal organoids [79] and the knockout of RB1 demonstrated its crucial role in retinal development [80].
Similarly, Magil et al. have recently applied a CRISPR-Cas interference-based approach in brain cancer organoids [81]. Particularly, they generated meningioma cells expressing a specific Cas9 from patients’ tumor samples and co-cultured them with brain organoids. Then, they repressed two key genes involved in meningioma proliferation (CHD2 and PTPRZ1) by transducing cells with lentiviruses harboring the single guide RNA (sgRNA) targeting the genes. This CRISPR-induced repression attenuated tumor cell proliferation and was valuable in defining the potential role of chromodomain helicase DNA binding protein 2 (CHD2) and Protein Tyrosine Phosphatase Receptor Type Z1 (PTPRZ1) as novel targets for molecular therapy to treat meningioma patients.
Overall, these findings suggest the valuable role of CRISPR in disease modeling and therapy testing using disease neural organoids and their isogenic counterparts.
4.2. Automation and High-Throughput Screening
Compared with organoid-based studies for cancer [82,83], automation in the generation and analysis of neural organoids is still in the early stages. Established protocols tend to lack a focus on scalable, homogenous organoids with a predictable morphology, cellular composition, and organization. Extensive manual handling includes complicated matrix embedding steps, rendering organoids painstaking to culture as well as time-consuming and resource-consuming as limiting the scale-up [84]. In addition, common analysis methods (e.g., tissue sectioning, immunohistochemical analysis, RNA sequence) do not perfectly fit high-throughput screening (HTS) requirements.
As already demonstrated using kidney organoids, liquid-handling robots can perform all steps of cell culturing, including plating, differentiation, and fixation [85]. The use of robots coupled with software trained to recognize organoids in microwells and to perform quantitative imaging analysis that enabled organoid phenotyping, providing an attractive starting point for high-quality HTS and possible application as a therapeutic screening tool. Recently, Renner et al. have used a similar system to generate a fully-automated HTS workflow to generate and analyze midbrain organoids [86]. The authors used a liquid-handling robot that could perform different steps, such as seeding, maintenance, fixation, whole-mount staining, and clearing in a fully scalable automated fashion in standard 96-well-plates. The resulting midbrain organoids showed small intra-batch and inter-batch variability in size distribution, morphology, and cellular composition as well as comparable morphological features compared with other studies [86]. Further characterization using RNA sequencing and quantitative whole mount HCI confirmed the reproducibility of these organoids.
Though preliminary, these automated methods from seeding to analysis in standard plates could represent a feasible scale-up and implementation into existing, screening facilities.
4.3. Imaging
Morphological analysis and microscopic dissection of human organoids are invaluable to investigate their cytoarchitecture. Fluorescence microscopy theoretically enables the study of intact organoids with high spatiotemporal resolution (nanometer and millisecond), providing a potential tool for spatiotemporal reconstructions known as 4D imaging. Unfortunately, different roadblocks have hampered their use in this context. Both traditional fluorescence microscopy and computational methods for 3D deconvolution are not ideal for studying organoids. In particular, the first struggle to sample 3D structures thicker than 10 micrometers, whereas the second faces challenges in properly deconvolving complex, multilayered architectures. To address these limitations, early organoid-based studies have implemented thin-sectioning methods, such as cryo-sectioning to obtain suitable samples for microscopy [1]. Though useful, these techniques are labor-intensive, impair the living tissue, and dismantle the 3D cytoarchitecture. Recently, various new approaches have allowed a quicker and a more traumatic observation of organoids.
The most common method for in vivo optical section is laser point scanning, as applied in confocal and two-photon microscopy [87,88]. Laser point scanning is based on a small laser point moved sequentially across a sample to different 3D positions, while an image is created by assigning the signal detected at each point with its position in 3D space. 3D microscopy is available in various commercial HCI systems. While many different confocal implementations exist, all these approaches share the same principle. Out-of-focus light is excluded from the microscope’s camera by using a pinhole. Several different flavors of confocal microscopy are commonly employed, from point scanning systems to spinning disc microscopy, compromising between Z resolution (axial) and speed. While point scanning devices offer sharper and deeper images in the sample, they are slower than spinning disc microscopes. Despite enabling 4D observations, they cause phototoxicity due to out-of-focus light, reducing their applicability to study living organoids. Multi-photon microscopy represents a noninvasive optical sectioning method to explore thick tissue samples and fine cellular details with high depth and low phototoxicity [89], but its commercial use is still limited.
Light-sheet fluorescence microscopy (LSFM) has emerged as a particularly fast and gentle technology, enabling both high-resolution imaging and live-cell microscopy. Unlike point-scanning, LSFM illuminates the regions of the sample being actively imaged by the device. This reduces laser power and phototoxicity while increasing the image dimension and rate of acquisition [90]. Low phototoxicity permits the imaging of a sample for hours or days, with clear advantages in terms of temporal analysis. Traditional LSFM requires specific sample mounting, making it unsuitable for automation or multi-well plates. Recent advances might enable 4D imaging in multi-well plates providing quicker analysis. The use of single objective light-sheet techniques [91,92,93] (instead of multiple objective lenses) has enabled low-resolution imaging of transparent multi-well plates. In parallel, improvements in optics [94] have increased the resolution of this technique. The commercialization of these systems will likely enable 4D imaging using available automation technologies.
Pitfalls in Organoid Imaging
Elevated scattering of light represents a common challenge in imaging complex biological tissues, including organoids. This phenomenon derives from the high structural complexity of these 3D cultures, particularly due to deposits of proteins, carbohydrates, and lipids [93]. Recent potential solutions to reduce tissue scattering include the use of adaptive optics compensating for the specific scattering of each sample, as shown in studies with drosophila and zebrafish [95,96]. However, the diffusion of these approaches is still limited and not widely commercialized.
Fixed organoids can be chemically modified to render them transparent and reduce light scattering. Early tissue clearing aims to reduce the scattering of tissue by replacing water with oils matching the refractive index of lipids’ membranes [97]. Different methods have been used, including solvent denaturation, hyperhydration, and tissue embedding [98]. The most common approach is clear lipid-exchanged acrylamide-hybridized rigid imaging/immunostaining/in situ hybridization-compatible tissue-hydrogel (CLARITY) [99]. This system is based on porous hydrogel tissue embedding and electrophoresis to remove lipids while preserving the structure and native biomolecules such as protein and nucleic acids. Its application on neural organoids has already enabled 3D imaging of whole cerebral organoids using LSFM [100]. However, none of the previous tools has seen widespread use in neural organoid-based studies yet. Future high-content screening platforms should focus on the compatibility of the clearing methods with fluorophores, dyes, and antibodies used for the characterization of organoids.
Organoids can be challenging for dyes and antibodies to diffuse through, limiting the application of traditional methods. Even well diffusive dyes such as 4′,6-diamidino-2-phenylindole (DAPI) may encounter difficulties in staining DNA in the cells toward the center of an organoid. Some solutions include the use of genetically encoded fluorescent proteins—especially red-shifted labels—coupled with tissue clearing methods. Fluorescence In Situ Hybridization (FISH) probes and antibodies seem to diffuse particularly well through cleared tissues, enabling the detection of proteins and nucleic acids [89,101]. However, many fluorescent dyes have scarce compatibility with clearing techniques [102,103], which is an aspect to properly evaluate before staining.
Prospectively, organoid imaging techniques might include highly multiplexed approaches, such as transcriptome mapping [104] and proteome imaging [105,106,107], using small nucleic acid probes and antibodies diffusing through cleared tissue. Transcriptome technologies based on in situ hybridization fluorescence with confocal microscopy can image up to 10,000 genes in single cells with high accuracy and resolution [108], enabling both morphological and transcriptomic phenotyping. Proteome imaging techniques achieve comprehensive profiling of intracellular protein maps with a quantitative description of compartmentalized intracellular proteins [106]. These maps help to identify cellular states and to define the intracellular organization between single cells in different cell cycle states and therapeutic contexts.
Altogether, these findings show that, despite some current technical challenges related to organoid imaging, recent progress in high-resolution imaging coupled with multiplexed approaches will likely enable valuable biological insights for drug discovery.
4.4. Single-Cell RNA Sequencing
Scientists have used different emerging techniques to delve into the biology of neural organoids, including scRNAseq. Historically, researchers have performed gene expression studies on “bulk” tissue samples composed of many cells of different types and functions. This approach has yielded valuable results in comparing general characteristics of tissues—such as pre-treatment and post-treatment conditions—though presenting a low-resolution for elucidating cellular composition within a tissue [109]. Advances in sgRNAseq have enabled us to profile gene expression in individual cells on a large scale, helping to investigate the heterogeneity across cell types, including those in complex neural organoids [110].
Although scRNAseq has not reached a full stride in neural organoid technology for drug discovery—differently from the preliminary results on cancer organoids [111], including glioblastoma [112,113,114]—this approach has opened up new possibilities in exploring neurodevelopment and cell phenotyping [42]. For instance, Quadrato et al. have highlighted a considerable variability of cellular populations within organoids [43]. Using a high-throughput droplet-based microfluidic approach known as Drop-seq [115], they analyzed 80,000 cells from 31 organoids grown over an extended period, providing the first molecular map of the variety of cells generated in neural organoids. The authors detected a large diversity of cell classes from different regions of the brain and retina, including glial cells and mature neurons. In addition, this cellular composition diversified over time, showing a progressive level of maturity.
The use of RNAseq has enabled researchers to bypass some of this variability by filtering out cells of no interest, increasing the granularity of the analysis (Figure 3).
Different high-throughput methods based on droplets microfluidic, combinatorial barcoding, and micro-well technologies have expanded the number of cells that can be investigated in each experiment. These approaches enable the quantitative comparisons of differential cell states within organoids.
However, the most frequently applied systems become labor-intensive and cost-prohibitive when processing more than 20 samples at a time [98]. Emerging “cell-hashing” approaches, including multiplexing for single-cell RNA sequencing (MULTI-seq) and Cellular Indexing of Transcriptomes and Epitopes by Sequencing (CITE-seq), might be more efficient in multiplexing single cells for scaled projects [116,117]. The MULTI-seq approach refers to the use of lipid-modified and cholesterol-modified oligonucleotides as sample barcodes in live cells and nuclei, regardless of species and genetic background, improving scRNA-seq data quality. By integrating this method with automated liquid handling, MULTI-seq might serve as a valuable technology to perform “screen-by-sequencing” (such as the DRUG-seq method [118]) in multicellular systems, including organoids [116]. The CITE-seq is based on oligonucleotide-tagged antibodies targeting cell surface proteins that uniquely label different experimental samples. This barcoded antibody signal—known as “cell hashing”—serves as a reference for assigning each multiplexed cell to its original sample. With this tool, different cellular samples can be labeled, pooled, and finally run simultaneously in a single-droplet scRNA-seq. This technology might enable a consistent sample multiplexing, precise multiplet identification, and a cost reduction of commercial scRNA-seq platforms [117].
Some limitations still hamper the widespread application of this method for organoids. Though rapidly decreasing, the cost of sequencing is still considerable. The challenges related to cell dissociation of organoids (e.g., dissociation of mature neurons from older organoids and batch effects introduced by dissociation [119]) still exist.
Overall, these findings suggest that the use of scRNA-seq for drug discovery in neural organoids is still in its formative years. However, its future application might provide valuable advancements in the field.
4.5. Machine Learning
Machine learning refers to a branch of artificial intelligence regarding computers that can improve their performance through experience, i.e., training. It is a rapidly growing technical field lying at the intersection between computer science and statistics. In cell biology, machine learning teaches computers to recognize phenotypes. Particularly, machine learning aims to learn processing rules from examples rather than relying on manual adjustments of parameters or pre-defined processing steps [120]. In this way, computational data analysis reduces the workload for the experimentalists and permit objectivity and consistency in the processing of large datasets. Machine learning typically proceeds in two phases. In the training phase, data samples are used to build or improve a computer system by learning from inherent structures and relationships within this data. In the second phase, this computer system is used to infer the properties of the data samples. The ultimate goal is to generalize from a few training examples to make predictions on larger data sets not observed during training [121].
4.5.1. Machine Learning in Drug Discovery Using Neural Organoids
Machine learning has proved to be useful in various stages of drug discovery, including analysis of digital pathology data in clinical trials, identification of prognostic biomarkers, and target validation [122]. Different machine learning methods, including the support vector machine [123], K nearest neighbors [124], naïve Bayes [125], random forest [126], and many others [120] have provided valuable insights in this context. In addition to these approaches, recent developments in deep neural networks [127] have catalyzed the interest and have sparked further research in the field. Machine learning approaches in the pharmaceutical industry are commonly used for virtual screening of compounds, performing low to medium-throughput screening [128]. Similar machine learning tools have been used in toxicological research [129].
Machine learning methods for drug discovery in neural organoids are still in their infancy. Most of the studies using organoids have focused on the phenotypical characterization of the model using machine learning tools. A study based on iPSC-derived cortical organoids grown over several months documented a similarity between the electrophysiological activity pattern of cortical organoids and human preterm neonatal electroencephalography using supervised machine learning [130]. In another study, iPSC-derived retinal organoids adopted a deep-learning computer algorithm based on bright-field imaging to recognize and predict retinal differentiation [131]. Particularly, they applied a transfer learning approach using convolutional neural network (CNN) algorithms that are loosely modeled on the network of a human brain agglomerated into multiple functional layers [87]. CNN processing consisted of two phases: a pretraining on the ImageNet classification Dataset [132] and the actual prediction of retinal differentiation in organoids, showing the ability of CNN-based approaches to classify stem-cell-derived tissues in vitro.
Regarding machine learning for drug discovery in neural organoids, the available literature is scant. A study by Monzel et al. developed a machine learning-based HCI analysis using Parkinson’s disease (PD) human midbrain organoids (hMO) exposed to toxic compounds. The researchers analyzed in vitro toxicity mediated by 6-hydroxydopamine (6-OHDA), a toxic molecule specifically targeting dopaminergic neurons, using HCI coupled with random forest classification. This approach discriminated between controls and treated organoids, predicting neurotoxin-induced cellular perturbations with a 75% accuracy [133].
Despite a widespread application of machine learning in drug discovery with organoid platforms still lacking, rapid progress in this field might advance their use in the future.
4.5.2. Future Perspectives on Machine Learning Applied to Organoid Technology
Culturing cerebral organoids has enabled high-throughput screening to test genetic and chemical modifications on cellular phenotypes [2]. Quantification of these phenotypes has historically been a laborious process, involving painstaking manual work. However, recent advances in microscopy and single-cell sequencing coupled with novel computing systems and data processing might support the description of phenotypes at an unprecedented speed.
High-content microscopy is fundamental to provide a characterization of cell components. In many instances, scientists rely on manual measurements of organoid phenotypes [134]. Some workflows permit a more rapid nucleus identification, neurite segmentation, and nucleus-to-cytoplasmic ratio calculation, limiting time-consuming work [135,136]. Recently, the use of CNN on microscopy imaging has improved cell phenotyping under various conditions [137,138], increasing imaging resolution [139], synapse quantification [140], and neurite measurements [141].
ScRNA-seq platforms can characterize cell types and states and they have offered new insights into the phenotyping of cellular systems (see single-cell RNA sequencing). However, as the scale of single-cell studies continues to grow, the increasing number of cells and the batch effect represent computational challenges. Emerging deep learning methods might represent a valuable tool to unfold complex cellular heterogeneous samples—such as organoids—using scRNA-seq [142,143]. Deep-learning refers to representation-learning methods with multiple layers of representation derived from the combination of single modules. Each module transforms the representation at one level (starting with the raw data) into a representation at a higher level [144]. The key aspect of this approach is that these layers of features are not human-driven but learn from data using a general-purpose learning process. Some examples include the Deep Embedding for Single-cell Clustering (DESC) method and scDeepCluster tool. The DESC approach proposed by Li and colleagues clusters scRNA-seq data through iterative clustering and self-learning [142], removing complex batch effects among cellular samples while preserving biological variations. ScDeepCluster refers to a clustering method that simultaneously learns future representation and clustering through modeling of scRNA-seq data generation [143], providing a valuable algorithm for large-scale scRNA-seq data. Other computational models that might help to analyze the amount of data from scRNA-seq of heterogeneous cellular samples include models of RNA splicing dynamics, enabling the study of the cell state and development [145,146,147].
Altogether, these findings suggest that high-content microscopy and scRNA-seq methods coupled with advances in computational tools might provide more reliable and cost-effective analysis of heterogeneous cellular models, including neural organoids.
5. Conclusions
The organoid technology is potentially revolutionary and might bridge the gap between preclinical and clinical trials, enabling more reliable testing models for precision medicine and drug discovery. So far, organoid-based research has been mainly conducted in academic environments, with only a few companies actively investing in the technology for scalable manufacturing. The target customers of these development companies include primarily academic researchers and larger pharmaceutical companies. Some of them provide media, reagents, and products for organoid development, while only a handful of them are commercializing whole organoids as products. However, many companies have pending patents [18] for organoid development and culturing, aiming to ramp up the production when the technology will be ripe for commercialization on a larger scale.
The integration of innovative bioengineering solutions could accelerate organoid manufacturing. The application of CRISPR-Cas technologies will help to provide more reliable disease models. The implementation of HCI and scRNA sequencing coupled with machine learning tools might enable the processing of larger datasets more rapidly. In addition, the use of automated systems will help to accelerate the generation of organoids. Microfluidic organoid-on-a-chip platforms [148] could provide an accurate system for consistent organoid perfusion, mimicking a more physiological nutrient supply. The confluence of different disciplines, including tissue engineering, artificial intelligence, and automation, will likely help to achieve a better reproducibility, scalability, and efficiency of the organoid model and to prompt its translation toward large-scale studies for drug discovery of neurological disorders.
Author Contributions
Conceptualization, G.C. and S.C. Investigation, G.C. Writing—original draft preparation, G.C. Writing—review and editing, G.C. and S.C. Visualization, G.C. Supervision, G.P.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Italian Ministry of Health, Foundation IRCCS Ca’ Granda Ospedale Maggiore Policlinico Ricerca Corrente 2021 to GPC and by Italian Ministry of Health, RF, RF-2018-12366357 to S.C.
Acknowledgments
The authors would like to thank Associazione Amici del Centro Dino Ferrari for its support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lancaster, M.A.; Knoblich, J.A. Generation of Cerebral Organoids from Human Pluripotent Stem Cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhibber, T.; Bagchi, S.; Lahooti, B.; Verma, A.; Al-Ahmad, A.; Paul, M.K.; Pendyala, G.; Jayant, R.D. CNS Organoids: An Innovative Tool for Neurological Disease Modeling and Drug Neurotoxicity Screening. Drug Discov. Today 2020, 25, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Dolmetsch, R.; Geschwind, D.H. The Human Brain in a Dish: The Promise of IPSC-Derived Neurons. Cell 2011, 145, 831–834. [Google Scholar] [CrossRef] [Green Version]
- Salani, S.; Donadoni, C.; Rizzo, F.; Bresolin, N.; Comi, G.P.; Corti, S. Generation of Skeletal Muscle Cells from Embryonic and Induced Pluripotent Stem Cells as an in Vitro Model and for Therapy of Muscular Dystrophies. J. Cell. Mol. Med. 2012, 16, 1353–1364. [Google Scholar] [CrossRef]
- Duarte, A.A.; Gogola, E.; Sachs, N.; Barazas, M.; Annunziato, S.; de Ruiter, J.R.; Velds, A.; Blatter, S.; Houthuijzen, J.M.; van de Ven, M.; et al. BRCA-Deficient Mouse Mammary Tumor Organoids to Study Cancer-Drug Resistance. Nat. Methods 2018, 15, 134–140. [Google Scholar] [CrossRef]
- Huch, M.; Gehart, H.; van Boxtel, R.; Hamer, K.; Blokzijl, F.; Verstegen, M.M.A.; Ellis, E.; van Wenum, M.; Fuchs, S.A.; de Ligt, J.; et al. Long-Term Culture of Genome-Stable Bipotent Stem Cells from Adult Human Liver. Cell 2015, 160, 299–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Wetering, M.; Francies, H.E.; Francis, J.M.; Bounova, G.; Iorio, F.; Pronk, A.; van Houdt, W.; van Gorp, J.; Taylor-Weiner, A.; Kester, L.; et al. Prospective Derivation of a Living Organoid Biobank of Colorectal Cancer Patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef] [Green Version]
- Kawada, J.; Kaneda, S.; Kirihara, T.; Maroof, A.; Levi, T.; Eggan, K.; Fujii, T.; Ikeuchi, Y. Generation of a Motor Nerve Organoid with Human Stem Cell-Derived Neurons. Stem Cell Rep. 2017, 9, 1441–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hor, J.H.; Soh, E.S.-Y.; Tan, L.Y.; Lim, V.J.W.; Santosa, M.M.; Winanto; Ho, B.X.; Fan, Y.; Soh, B.-S.; Ng, S.-Y. Cell Cycle Inhibitors Protect Motor Neurons in an Organoid Model of Spinal Muscular Atrophy. Cell Death Dis. 2018, 9, 1100. [Google Scholar] [CrossRef]
- Amin, N.D.; Paşca, S.P. Building Models of Brain Disorders with Three-Dimensional Organoids. Neuron 2018, 100, 389–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wray, S. Modelling Neurodegenerative Disease Using Brain Organoids. Semin. Cell Dev. Biol. 2020, 111. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral Organoids Model Human Brain Development and Microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Iefremova, V.; Manikakis, G.; Krefft, O.; Jabali, A.; Weynans, K.; Wilkens, R.; Marsoner, F.; Brändl, B.; Müller, F.-J.; Koch, P.; et al. An Organoid-Based Model of Cortical Development Identifies Non-Cell-Autonomous Defects in Wnt Signaling Contributing to Miller-Dieker Syndrome. Cell Rep. 2017, 19, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-T.; Seo, J.; Gao, F.; Feldman, H.M.; Wen, H.-L.; Penney, J.; Cam, H.P.; Gjoneska, E.; Raja, W.K.; Cheng, J.; et al. APOE4 Causes Widespread Molecular and Cellular Alterations Associated with Alzheimer’s Disease Phenotypes in Human IPSC-Derived Brain Cell Types. Neuron 2018, 98, 1294. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Izpisua Belmonte, J.C. Organoids—Preclinical Models of Human Disease. N. Engl. J. Med. 2019, 380, 569–579. [Google Scholar] [CrossRef]
- Abidin, N.Z.; Haq, I.J.; Gardner, A.I.; Brodlie, M. Ataluren in Cystic Fibrosis: Development, Clinical Studies and Where Are We Now? Expert Opin. Pharmacother. 2017, 18, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, G.; Ponnusamy, M.P.; Batra, S.K. Concise Review: Current Status of Three-Dimensional Organoids as Preclinical Models. Stem Cells 2018, 36, 1329–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, D.; Ashok, A.; Naing, M.W. Commercialization of Organoids. Trends Mol. Med. 2020, 26, 245–249. [Google Scholar] [CrossRef]
- Wells, M.F.; Salick, M.R.; Wiskow, O.; Ho, D.J.; Worringer, K.A.; Ihry, R.J.; Kommineni, S.; Bilican, B.; Klim, J.R.; Hill, E.J.; et al. Genetic Ablation of AXL Does Not Protect Human Neural Progenitor Cells and Cerebral Organoids from Zika Virus Infection. Cell Stem Cell 2016, 19, 703–708. [Google Scholar] [CrossRef] [Green Version]
- Salick, M.R.; Wells, M.F.; Eggan, K.; Kaykas, A. Modelling Zika Virus Infection of the Developing Human Brain In Vitro Using Stem Cell Derived Cerebral Organoids. J. Vis. Exp. 2017, 56404. [Google Scholar] [CrossRef] [Green Version]
- Technology-Microbrain. Available online: https://www.stemonix.com (accessed on 5 March 2021).
- Inacio, P. 3D Microbrains Can Accelerate Development of Rett Syndrome Therapies. Available online: https://rettsyndromenews.com/2019/02/21/stemonix-develops-3d-human-microbrains-of-rett-syndrome-to-accelerate-therapy-development/ (accessed on 5 March 2021).
- System1 Biosciences. Available online: https://system1.bio/ (accessed on 8 December 2020).
- A:Head. Available online: https://www.viennabiocenter.org/companies/biotech-companies/ahead/ (accessed on 8 December 2020).
- STEMCELL Technologies Signs Exclusive Agreement with the Institute of Molecular Biotechnology to Commercialize Cerebral Organoid Culture Systems. Available online: https://www.Stemcell.Com/about-Us/News/Stemcell-Technologies-Signs-Exclusive-Agreement-with-the-Institute-of-Molecular-Biotechnology-to-Commercialize-Cerebral-Organoid-Culture-Systems.Html (accessed on 5 March 2021).
- Li, R.; Sun, L.; Fang, A.; Li, P.; Wu, Q.; Wang, X. Recapitulating Cortical Development with Organoid Culture in Vitro and Modeling Abnormal Spindle-like (ASPM Related Primary) Microcephaly Disease. Protein Cell 2017, 8, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, E.; Wason, A.; Ramani, A.; Gooi, L.M.; Keller, P.; Pozniakovsky, A.; Poser, I.; Noack, F.; Telugu, N.S.; Calegari, F.; et al. CPAP Promotes Timely Cilium Disassembly to Maintain Neural Progenitor Pool. EMBO J. 2016, 35, 803–819. [Google Scholar] [CrossRef]
- Klaus, J.; Kanton, S.; Kyrousi, C.; Ayo-Martin, A.C.; Di Giaimo, R.; Riesenberg, S.; O’Neill, A.C.; Camp, J.G.; Tocco, C.; Santel, M.; et al. Altered Neuronal Migratory Trajectories in Human Cerebral Organoids Derived from Individuals with Neuronal Heterotopia. Nat. Med. 2019, 25, 561–568. [Google Scholar] [CrossRef]
- Pașca, A.M.; Park, J.-Y.; Shin, H.-W.; Qi, Q.; Revah, O.; Krasnoff, R.; O’Hara, R.; Willsey, A.J.; Palmer, T.D.; Pașca, S.P. Human 3D Cellular Model of Hypoxic Brain Injury of Prematurity. Nat. Med. 2019, 25, 784–791. [Google Scholar] [CrossRef]
- Xu, R.; Brawner, A.T.; Li, S.; Liu, J.-J.; Kim, H.; Xue, H.; Pang, Z.P.; Kim, W.-Y.; Hart, R.P.; Liu, Y.; et al. OLIG2 Drives Abnormal Neurodevelopmental Phenotypes in Human IPSC-Based Organoid and Chimeric Mouse Models of Down Syndrome. Cell Stem Cell 2019, 24, 908–926.e8. [Google Scholar] [CrossRef]
- Cairns, D.M.; Rouleau, N.; Parker, R.N.; Walsh, K.G.; Gehrke, L.; Kaplan, D.L. A 3D Human Brain-like Tissue Model of Herpes-Induced Alzheimer’s Disease. Sci. Adv. 2020, 6, eaay8828. [Google Scholar] [CrossRef]
- Kim, H.; Park, H.J.; Choi, H.; Chang, Y.; Park, H.; Shin, J.; Kim, J.; Lengner, C.J.; Lee, Y.K.; Kim, J. Modeling G2019S-LRRK2 Sporadic Parkinson’s Disease in 3D Midbrain Organoids. Stem Cell Rep. 2019, 12, 518–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groveman, B.R.; Foliaki, S.T.; Orru, C.D.; Zanusso, G.; Carroll, J.A.; Race, B.; Haigh, C.L. Sporadic Creutzfeldt-Jakob Disease Prion Infection of Human Cerebral Organoids. Acta Neuropathol. Commun. 2019, 7, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, S.; Repic, M.; Guo, Z.; Kavirayani, A.; Burkard, T.; Bagley, J.A.; Krauditsch, C.; Knoblich, J.A. Genetically Engineered Cerebral Organoids Model Brain Tumour Formation. Nat. Methods 2018, 15, 631–639. [Google Scholar] [CrossRef] [PubMed]
- D’Aiuto, L.; Bloom, D.C.; Naciri, J.N.; Smith, A.; Edwards, T.G.; McClain, L.; Callio, J.A.; Jessup, M.; Wood, J.; Chowdari, K.; et al. Modeling Herpes Simplex Virus 1 Infections in Human Central Nervous System Neuronal Cells Using Two- and Three-Dimensional Cultures Derived from Induced Pluripotent Stem Cells. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, R.M.; Rana, P.S.J.B.; Jaeger, H.K.; O’Dowd, J.M.; Balemba, O.B.; Fortunato, E.A. Human Cytomegalovirus Compromises Development of Cerebral Organoids. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-P.; Qiu, Y.; Zhang, B.; Chen, G.; Chen, Q.; Wang, M.; Mo, F.; Xu, J.; Wu, J.; Zhang, R.-R.; et al. Zika Virus Infection Induces RNAi-Mediated Antiviral Immunity in Human Neural Progenitors and Brain Organoids. Cell Res. 2019, 29, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Stachowiak, E.K.; Benson, C.A.; Narla, S.T.; Dimitri, A.; Chuye, L.E.B.; Dhiman, S.; Harikrishnan, K.; Elahi, S.; Freedman, D.; Brennand, K.J.; et al. Cerebral Organoids Reveal Early Cortical Maldevelopment in Schizophrenia—Computational Anatomy and Genomics, Role of FGFR1. Transl. Psychiatry 2017, 7, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, L.; Zhu, Y.; Qin, J. Human Brain Organoid-on-a-Chip to Model Prenatal Nicotine Exposure. Lab Chip 2018, 18, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Dang, J.; Tiwari, S.K.; Agrawal, K.; Hui, H.; Qin, Y.; Rana, T.M. Glial Cell Diversity and Methamphetamine-Induced Neuroinflammation in Human Cerebral Organoids. Mol. Psychiatry 2020, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ao, Z.; Cai, H.; Havert, D.J.; Wu, Z.; Gong, Z.; Beggs, J.M.; Mackie, K.; Guo, F. One-Stop Microfluidic Assembly of Human Brain Organoids To Model Prenatal Cannabis Exposure. Anal. Chem. 2020, 92, 4630–4638. [Google Scholar] [CrossRef]
- Brancati, G.; Treutlein, B.; Camp, J.G. Resolving Neurodevelopmental and Vision Disorders Using Organoid Single-Cell Multi-Omics. Neuron 2020, 107, 1000–1013. [Google Scholar] [CrossRef]
- Quadrato, G.; Nguyen, T.; Macosko, E.Z.; Sherwood, J.L.; Min Yang, S.; Berger, D.R.; Maria, N.; Scholvin, J.; Goldman, M.; Kinney, J.P.; et al. Cell Diversity and Network Dynamics in Photosensitive Human Brain Organoids. Nature 2017, 545, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Pollen, A.A.; Bhaduri, A.; Andrews, M.G.; Nowakowski, T.J.; Meyerson, O.S.; Mostajo-Radji, M.A.; Di Lullo, E.; Alvarado, B.; Bedolli, M.; Dougherty, M.L.; et al. Establishing Cerebral Organoids as Models of Human-Specific Brain Evolution. Cell 2019, 176, 743–756.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, J.; Kang, J.S.; Lee, M.; Baek, A.; Kim, S.; Chung, S.-K.; Lee, M.-O.; Kim, J. Simplified Brain Organoids for Rapid and Robust Modeling of Brain Disease. Front. Cell Dev. Biol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Nickels, S.L.; Modamio, J.; Mendes-Pinheiro, B.; Monzel, A.S.; Betsou, F.; Schwamborn, J.C. Reproducible Generation of Human Midbrain Organoids for in Vitro Modeling of Parkinson’s Disease. Stem Cell Res. 2020, 46, 101870. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.P.; Fernandes, T.G.; Nogueira, D.E.S.; Rodrigues, C.A.V.; Bekman, E.P.; Hashimura, Y.; Jung, S.; Lee, B.; Carmo-Fonseca, M.; Cabral, J.M.S. Scalable Generation of Mature Cerebellar Organoids from Human Pluripotent Stem Cells and Characterization by Immunostaining. JoVE 2020, e61143. [Google Scholar] [CrossRef]
- Romero-Morales, A.I.; O’Grady, B.J.; Balotin, K.M.; Bellan, L.M.; Lippmann, E.S.; Gama, V. Spin∞: An Updated Miniaturized Spinning Bioreactor Design for the Generation of Human Cerebral Organoids from Pluripotent Stem Cells. HardwareX 2019, 6, e00084. [Google Scholar] [CrossRef] [PubMed]
- Sood, D.; Tang-Schomer, M.; Pouli, D.; Mizzoni, C.; Raia, N.; Tai, A.; Arkun, K.; Wu, J.; Black, L.D.; Scheffler, B.; et al. 3D Extracellular Matrix Microenvironment in Bioengineered Tissue Models of Primary Pediatric and Adult Brain Tumors. Nat. Commun. 2019, 10, 4529. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, M.A.; Corsini, N.S.; Wolfinger, S.; Gustafson, E.H.; Phillips, A.W.; Burkard, T.R.; Otani, T.; Livesey, F.J.; Knoblich, J.A. Guided Self-Organization and Cortical Plate Formation in Human Brain Organoids. Nat. Biotechnol. 2017, 35, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, G.T.; Lundin, B.F.; Iyer, N.; Ashton, L.M.; Sethares, W.A.; Willett, R.M.; Ashton, R.S. Engineering Induction of Singular Neural Rosette Emergence within HPSC-Derived Tissues. eLife 2018, 7, e37549. [Google Scholar] [CrossRef] [PubMed]
- Haremaki, T.; Metzger, J.J.; Rito, T.; Ozair, M.Z.; Etoc, F.; Brivanlou, A.H. Self-Organizing Neuruloids Model Developmental Aspects of Huntington’s Disease in the Ectodermal Compartment. Nat. Biotechnol. 2019, 37, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Dosso, A.D.; Urenda, J.-P.; Nguyen, T.; Quadrato, G. Upgrading the Physiological Relevance of Human Brain Organoids. Neuron 2020, 107, 1014–1028. [Google Scholar] [CrossRef]
- Pham, M.T.; Pollock, K.M.; Rose, M.D.; Cary, W.A.; Stewart, H.R.; Zhou, P.; Nolta, J.A.; Waldau, B. Generation of Human Vascularized Brain Organoids. Neuroreport 2018, 29, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.A.; Gonçalves, J.T.; Bloyd, C.W.; Li, H.; Fernandes, S.; Quang, D.; Johnston, S.; Parylak, S.L.; Jin, X.; Gage, F.H. An in Vivo Model of Functional and Vascularized Human Brain Organoids. Nat. Biotechnol. 2018, 36, 432–441. [Google Scholar] [CrossRef]
- Costamagna, G.; Andreoli, L.; Corti, S.; Faravelli, I. IPSCs-Based Neural 3D Systems: A Multidimensional Approach for Disease Modeling and Drug Discovery. Cells 2019, 8, 1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, O.; Jin, Y.B.; Kim, J.; Lee, M.-O. Blood Vessel Formation in Cerebral Organoids Formed from Human Embryonic Stem Cells. Biochem. Biophys. Res. Commun. 2020, 521, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Wörsdörfer, P.; Dalda, N.; Kern, A.; Krüger, S.; Wagner, N.; Kwok, C.K.; Henke, E.; Ergün, S. Generation of Complex Human Organoid Models Including Vascular Networks by Incorporation of Mesodermal Progenitor Cells. Sci. Rep. 2019, 9, 15663. [Google Scholar] [CrossRef] [Green Version]
- Cakir, B.; Xiang, Y.; Tanaka, Y.; Kural, M.H.; Parent, M.; Kang, Y.-J.; Chapeton, K.; Patterson, B.; Yuan, Y.; He, C.-S.; et al. Engineering of Human Brain Organoids with a Functional Vascular-like System. Nat. Methods 2019, 16, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Nzou, G.; Wicks, R.T.; Wicks, E.E.; Seale, S.A.; Sane, C.H.; Chen, A.; Murphy, S.V.; Jackson, J.D.; Atala, A.J. Human Cortex Spheroid with a Functional Blood Brain Barrier for High-Throughput Neurotoxicity Screening and Disease Modeling. Sci. Rep. 2018, 8, 7413. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, S.; Lawler, S.E.; Qu, Y.; Fadzen, C.M.; Wolfe, J.M.; Regan, M.S.; Pentelute, B.L.; Agar, N.Y.R.; Cho, C.-F. Blood–Brain-Barrier Organoids for Investigating the Permeability of CNS Therapeutics. Nat. Protoc. 2018, 13, 2827–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.; Su, Y.; Adam, C.D.; Deutschmann, A.U.; Pather, S.R.; Goldberg, E.M.; Su, K.; Li, S.; Lu, L.; Jacob, F.; et al. Sliced Human Cortical Organoids for Modeling Distinct Cortical Layer Formation. Cell Stem Cell 2020, 26, 766–781.e9. [Google Scholar] [CrossRef]
- Giandomenico, S.L.; Mierau, S.B.; Gibbons, G.M.; Wenger, L.M.D.; Masullo, L.; Sit, T.; Sutcliffe, M.; Boulanger, J.; Tripodi, M.; Derivery, E.; et al. Cerebral Organoids at the Air–Liquid Interface Generate Diverse Nerve Tracts with Functional Output. Nat. Neurosci. 2019, 22, 669–679. [Google Scholar] [CrossRef]
- Tanaka, Y.; Cakir, B.; Xiang, Y.; Sullivan, G.J.; Park, I.-H. Synthetic Analyses of Single-Cell Transcriptomes from Multiple Brain Organoids and Fetal Brain. Cell Rep. 2020, 30, 1682–1689.e3. [Google Scholar] [CrossRef] [Green Version]
- Durens, M.; Nestor, J.; Williams, M.; Herold, K.; Niescier, R.F.; Lunden, J.W.; Phillips, A.W.; Lin, Y.-C.; Dykxhoorn, D.M.; Nestor, M.W. High-Throughput Screening of Human Induced Pluripotent Stem Cell-Derived Brain Organoids. J. Neurosci. Methods 2020, 335, 108627. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, D.; Clevers, H.; Artegiani, B. CRISPR-Cas Tools and Their Application in Genetic Engineering of Human Stem Cells and Organoids. Cell Stem Cell 2020, 27, 705–731. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Zhang, J.; Wu, H.; Wang, J.; Ma, K.; Li, Z.; Zhang, X.; Zhang, P.; Huang, X. Generation of Gene-Modified Mice via Cas9/RNA-Mediated Gene Targeting. Cell Res. 2013, 23, 720–723. [Google Scholar] [CrossRef]
- Li, D.; Qiu, Z.; Shao, Y.; Chen, Y.; Guan, Y.; Liu, M.; Li, Y.; Gao, N.; Wang, L.; Lu, X.; et al. Heritable Gene Targeting in the Mouse and Rat Using a CRISPR-Cas System. Nat. Biotechnol. 2013, 31, 681–683. [Google Scholar] [CrossRef]
- Matsui, T.; Nieto-Estévez, V.; Kyrychenko, S.; Schneider, J.W.; Hsieh, J. Retinoblastoma Protein Controls Growth, Survival and Neuronal Migration in Human Cerebral Organoids. Development 2017, 144, 1025–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Muffat, J.; Omer, A.; Bosch, I.; Lancaster, M.A.; Sur, M.; Gehrke, L.; Knoblich, J.A.; Jaenisch, R. Induction of Expansion and Folding in Human Cerebral Organoids. Cell Stem Cell 2017, 20, 385–396.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalebic, N.; Taverna, E.; Tavano, S.; Wong, F.K.; Suchold, D.; Winkler, S.; Huttner, W.B.; Sarov, M. CRISPR/Cas9-Induced Disruption of Gene Expression in Mouse Embryonic Brain and Single Neural Stem Cells in Vivo. EMBO Rep. 2016, 17, 338–348. [Google Scholar] [CrossRef]
- Ogawa, J.; Pao, G.M.; Shokhirev, M.N.; Verma, I.M. Glioblastoma Model Using Human Cerebral Organoids. Cell Rep. 2018, 23, 1220–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bershteyn, M.; Nowakowski, T.J.; Pollen, A.A.; Di Lullo, E.; Nene, A.; Wynshaw-Boris, A.; Kriegstein, A.R. Human IPSC-Derived Cerebral Organoids Model Cellular Features of Lissencephaly and Reveal Prolonged Mitosis of Outer Radial Glia. Cell Stem Cell 2017, 20, 435–449.e4. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Mokhtari, R.; Pedrosa, E.; Kirschenbaum, M.; Bayrak, C.; Zheng, D.; Lachman, H.M. CRISPR/Cas9-Mediated Heterozygous Knockout of the Autism Gene CHD8 and Characterization of Its Transcriptional Networks in Cerebral Organoids Derived from IPS Cells. Mol. Autism 2017, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Knupp, A.; Mishra, S.; Martinez, R.; Braggin, J.E.; Szabo, M.; Kinoshita, C.; Hailey, D.W.; Small, S.A.; Jayadev, S.; Young, J.E. Depletion of the AD Risk Gene SORL1 Selectively Impairs Neuronal Endosomal Traffic Independent of Amyloidogenic APP Processing. Cell Rep. 2020, 31, 107719. [Google Scholar] [CrossRef] [PubMed]
- Burnight, E.R.; Gupta, M.; Wiley, L.A.; Anfinson, K.R.; Tran, A.; Triboulet, R.; Hoffmann, J.M.; Klaahsen, D.L.; Andorf, J.L.; Jiao, C.; et al. Using CRISPR-Cas9 to Generate Gene-Corrected Autologous IPSCs for the Treatment of Inherited Retinal Degeneration. Mol. Ther. 2017, 25, 1999–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- First CRISPR Therapy Dosed. Nat. Biotechnol. 2020, 38, 382. [CrossRef] [PubMed] [Green Version]
- Deng, W.-L.; Gao, M.-L.; Lei, X.-L.; Lv, J.-N.; Zhao, H.; He, K.-W.; Xia, X.-X.; Li, L.-Y.; Chen, Y.-C.; Li, Y.-P.; et al. Gene Correction Reverses Ciliopathy and Photoreceptor Loss in IPSC-Derived Retinal Organoids from Retinitis Pigmentosa Patients. Stem Cell Rep. 2018, 10, 1267–1281. [Google Scholar] [CrossRef] [Green Version]
- VanderWall, K.B.; Huang, K.-C.; Pan, Y.; Lavekar, S.S.; Fligor, C.M.; Allsop, A.R.; Lentsch, K.A.; Dang, P.; Zhang, C.; Tseng, H.C.; et al. Retinal Ganglion Cells with a Glaucoma OPTN(E50K) Mutation Exhibit Neurodegenerative Phenotypes When Derived from Three-Dimensional Retinal Organoids. Stem Cell Rep. 2020, 15, 52–66. [Google Scholar] [CrossRef]
- Zheng, C.; Schneider, J.W.; Hsieh, J. Role of RB1 in Human Embryonic Stem Cell-Derived Retinal Organoids. Dev. Biol. 2020, 462, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Magill, S.T.; Vasudevan, H.N.; Seo, K.; Villanueva-Meyer, J.E.; Choudhury, A.; John Liu, S.; Pekmezci, M.; Findakly, S.; Hilz, S.; Lastella, S.; et al. Multiplatform Genomic Profiling and Magnetic Resonance Imaging Identify Mechanisms Underlying Intratumor Heterogeneity in Meningioma. Nat. Commun. 2020, 11, 4803. [Google Scholar] [CrossRef] [PubMed]
- Brandenberg, N.; Hoehnel, S.; Kuttler, F.; Homicsko, K.; Ceroni, C.; Ringel, T.; Gjorevski, N.; Schwank, G.; Coukos, G.; Turcatti, G.; et al. High-Throughput Automated Organoid Culture via Stem-Cell Aggregation in Microcavity Arrays. Nat. Biomed. Eng. 2020, 4, 863–874. [Google Scholar] [CrossRef]
- Driehuis, E.; Kretzschmar, K.; Clevers, H. Establishment of Patient-Derived Cancer Organoids for Drug-Screening Applications. Nat. Protoc. 2020, 15, 3380–3409. [Google Scholar] [CrossRef]
- Tong, Z.; Martyn, K.; Yang, A.; Yin, X.; Mead, B.E.; Joshi, N.; Sherman, N.E.; Langer, R.S.; Karp, J.M. Towards a Defined ECM and Small Molecule Based Monolayer Culture System for the Expansion of Mouse and Human Intestinal Stem Cells. Biomaterials 2018, 154, 60–73. [Google Scholar] [CrossRef]
- Czerniecki, S.M.; Cruz, N.M.; Harder, J.L.; Menon, R.; Annis, J.; Otto, E.A.; Gulieva, R.E.; Islas, L.V.; Kim, Y.K.; Tran, L.M.; et al. High-Throughput Automation Enhances Kidney Organoid Differentiation from Human Pluripotent Stem Cells and Enables Multidimensional Phenotypic Screening. Cell Stem Cell 2018, 22, 929–940.e4. [Google Scholar] [CrossRef] [Green Version]
- Renner, H.; Grabos, M.; Becker, K.J.; Kagermeier, T.E.; Wu, J.; Otto, M.; Peischard, S.; Zeuschner, D.; TsyTsyura, Y.; Disse, P.; et al. A Fully Automated High-Throughput Workflow for 3D-Based Chemical Screening in Human Midbrain Organoids. eLife 2020, 9. [Google Scholar] [CrossRef]
- White, J.G.; Amos, W.B.; Fordham, M. An Evaluation of Confocal versus Conventional Imaging of Biological Structures by Fluorescence Light Microscopy. J. Cell Biol. 1987, 105, 41–48. [Google Scholar] [CrossRef]
- Denk, W.; Strickler, J.H.; Webb, W.W. Two-Photon Laser Scanning Fluorescence Microscopy. Science 1990, 248, 73–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekkers, J.F.; Alieva, M.; Wellens, L.M.; Ariese, H.C.R.; Jamieson, P.R.; Vonk, A.M.; Amatngalim, G.D.; Hu, H.; Oost, K.C.; Snippert, H.J.G.; et al. High-Resolution 3D Imaging of Fixed and Cleared Organoids. Nat. Protoc. 2019, 14, 1756–1771. [Google Scholar] [CrossRef] [PubMed]
- Rios, A.C.; Clevers, H. Imaging Organoids: A Bright Future Ahead. Nat. Methods 2018, 15, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, M.B.; Voleti, V.; Mendes, C.S.; Lacefield, C.; Grueber, W.B.; Mann, R.S.; Bruno, R.M.; Hillman, E.M.C. Swept Confocally-Aligned Planar Excitation (SCAPE) Microscopy for High-Speed Volumetric Imaging of Behaving Organisms. Nat. Photonics 2015, 9, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakotoson, I.; Delhomme, B.; Djian, P.; Deeg, A.; Brunstein, M.; Seebacher, C.; Uhl, R.; Ricard, C.; Oheim, M. Fast 3-D Imaging of Brain Organoids With a New Single-Objective Planar-Illumination Two-Photon Microscope. Front. Neuroanat. 2019, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillman, E.M.C.; Voleti, V.; Li, W.; Yu, H. Light-Sheet Microscopy in Neuroscience. Annu. Rev. Neurosci. 2019, 42, 295–313. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Q.; Huang, F.; Bai, K.; Guo, Y.; Zhang, Y.; Li, N.; Cui, Y.; Sun, P.; Zeng, S.; et al. High-Throughput Light Sheet Tomography Platform for Automated Fast Imaging of Whole Mouse Brain. J. Biophotonics 2018, 11, e201800047. [Google Scholar] [CrossRef] [PubMed]
- Legant, W.R.; Shao, L.; Grimm, J.B.; Brown, T.A.; Milkie, D.E.; Avants, B.B.; Lavis, L.D.; Betzig, E. High-Density Three-Dimensional Localization Microscopy across Large Volumes. Nat. Methods 2016, 13, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Royer, L.A.; Lemon, W.C.; Chhetri, R.K.; Wan, Y.; Coleman, M.; Myers, E.W.; Keller, P.J. Adaptive Light-Sheet Microscopy for Long-Term, High-Resolution Imaging in Living Organisms. Nat. Biotechnol. 2016, 34, 1267–1278. [Google Scholar] [CrossRef]
- Dodt, H.-U.; Leischner, U.; Schierloh, A.; Jährling, N.; Mauch, C.P.; Deininger, K.; Deussing, J.M.; Eder, M.; Zieglgänsberger, W.; Becker, K. Ultramicroscopy: Three-Dimensional Visualization of Neuronal Networks in the Whole Mouse Brain. Nat. Methods 2007, 4, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Salick, M.R.; Lubeck, E.; Riesselman, A.; Kaykas, A. The Future of Cerebral Organoids in Drug Discovery. Semin. Cell Dev. Biol. 2020, 67–73. [Google Scholar] [CrossRef]
- Chung, K.; Deisseroth, K. CLARITY for Mapping the Nervous System. Nat. Methods 2013, 10, 508–513. [Google Scholar] [CrossRef]
- Sakaguchi, H.; Ozaki, Y.; Ashida, T.; Matsubara, T.; Oishi, N.; Kihara, S.; Takahashi, J. Self-Organized Synchronous Calcium Transients in a Cultured Human Neural Network Derived from Cerebral Organoids. Stem Cell Rep. 2019, 13, 458–473. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.; Lubeck, E.; Schwarzkopf, M.; He, T.-F.; Greenbaum, A.; Sohn, C.H.; Lignell, A.; Choi, H.M.T.; Gradinaru, V.; Pierce, N.A.; et al. Single-Molecule RNA Detection at Depth by Hybridization Chain Reaction and Tissue Hydrogel Embedding and Clearing. Development 2016, 143, 2862–2867. [Google Scholar] [CrossRef] [Green Version]
- Jensen, K.H.R.; Berg, R.W. CLARITY-Compatible Lipophilic Dyes for Electrode Marking and Neuronal Tracing. Sci. Rep. 2016, 6, 32674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hama, H.; Hioki, H.; Namiki, K.; Hoshida, T.; Kurokawa, H.; Ishidate, F.; Kaneko, T.; Akagi, T.; Saito, T.; Saido, T.; et al. ScaleS: An Optical Clearing Palette for Biological Imaging. Nat. Neurosci. 2015, 18, 1518–1529. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Allen, W.E.; Wright, M.A.; Sylwestrak, E.L.; Samusik, N.; Vesuna, S.; Evans, K.; Liu, C.; Ramakrishnan, C.; Liu, J.; et al. Three-Dimensional Intact-Tissue Sequencing of Single-Cell Transcriptional States. Science 2018, 361. [Google Scholar] [CrossRef] [Green Version]
- Goltsev, Y.; Samusik, N.; Kennedy-Darling, J.; Bhate, S.; Hale, M.; Vazquez, G.; Black, S.; Nolan, G.P. Deep Profiling of Mouse Splenic Architecture with CODEX Multiplexed Imaging. Cell 2018, 174, 968–981.e15. [Google Scholar] [CrossRef] [Green Version]
- Gut, G.; Herrmann, M.D.; Pelkmans, L. Multiplexed Protein Maps Link Subcellular Organization to Cellular States. Science 2018, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, C.; Lewis, E.E.L.; Flint, L.; Clench, M.R. Mass Spectrometry Imaging of 3D Tissue Models. Proteomics 2018, 18, e1700462. [Google Scholar] [CrossRef]
- Eng, C.-H.L.; Lawson, M.; Zhu, Q.; Dries, R.; Koulena, N.; Takei, Y.; Yun, J.; Cronin, C.; Karp, C.; Yuan, G.-C.; et al. Transcriptome-Scale Super-Resolved Imaging in Tissues by RNA SeqFISH+. Nature 2019, 568, 235–239. [Google Scholar] [CrossRef]
- Camp, J.G.; Treutlein, B. Human Organomics: A Fresh Approach to Understanding Human Development Using Single-Cell Transcriptomics. Development 2017, 144, 1584–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, J.R.; Ribas, A.; Mischel, P.S. Single-Cell Analysis Tools for Drug Discovery and Development. Nat. Rev. Drug Discov. 2015, 15, 204–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weeber, F.; Ooft, S.N.; Dijkstra, K.K.; Voest, E.E. Tumor Organoids as a Pre-Clinical Cancer Model for Drug Discovery. Cell Chem. Biol. 2017, 24, 1092–1100. [Google Scholar] [CrossRef]
- Chadwick, M.; Yang, C.; Liu, L.; Gamboa, C.M.; Jara, K.; Lee, H.; Sabaawy, H.E. Rapid Processing and Drug Evaluation in Glioblastoma Patient-Derived Organoid Models with 4D Bioprinted Arrays. iScience 2020, 23, 101365. [Google Scholar] [CrossRef]
- Jacob, F.; Salinas, R.D.; Zhang, D.Y.; Nguyen, P.T.T.; Schnoll, J.G.; Wong, S.Z.H.; Thokala, R.; Sheikh, S.; Saxena, D.; Prokop, S.; et al. A Patient-Derived Glioblastoma Organoid Model and Biobank Recapitulates Inter- and Intra-Tumoral Heterogeneity. Cell 2020, 180, 188–204.e22. [Google Scholar] [CrossRef]
- Zhang, C.; Jin, M.; Zhao, J.; Chen, J.; Jin, W. Organoid Models of Glioblastoma: Advances, Applications and Challenges. Am. J. Cancer Res. 2020, 10, 2242–2257. [Google Scholar]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-Wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- McGinnis, C.S.; Patterson, D.M.; Winkler, J.; Conrad, D.N.; Hein, M.Y.; Srivastava, V.; Hu, J.L.; Murrow, L.M.; Weissman, J.S.; Werb, Z.; et al. MULTI-Seq: Sample Multiplexing for Single-Cell RNA Sequencing Using Lipid-Tagged Indices. Nat. Methods 2019, 16, 619–626. [Google Scholar] [CrossRef]
- Stoeckius, M.; Zheng, S.; Houck-Loomis, B.; Hao, S.; Yeung, B.Z.; Mauck, W.M.; Smibert, P.; Satija, R. Cell Hashing with Barcoded Antibodies Enables Multiplexing and Doublet Detection for Single Cell Genomics. Genome Biol. 2018, 19, 224. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Ho, D.J.; Neri, M.; Yang, C.; Kulkarni, T.; Randhawa, R.; Henault, M.; Mostacci, N.; Farmer, P.; Renner, S.; et al. DRUG-Seq for Miniaturized High-Throughput Transcriptome Profiling in Drug Discovery. Nat. Commun. 2018, 9, 4307. [Google Scholar] [CrossRef]
- Kanton, S.; Treutlein, B.; Camp, J.G. Chapter 10—Single-Cell genomic analysis of human cerebral organoids. In Methods in Cell Biology; Spence, J.R., Ed.; Human Pluripotent Stem Cell Derived Organoid Models; Academic Press: Cambridge, MA, USA, 2020; Volume 159, pp. 229–256. [Google Scholar]
- Jordan, M.I.; Mitchell, T.M. Machine Learning: Trends, Perspectives, and Prospects. Science 2015, 349, 255–260. [Google Scholar] [CrossRef] [PubMed]
- De Ridder, D.; de Ridder, J.; Reinders, M.J.T. Pattern Recognition in Bioinformatics. Brief. Bioinform. 2013, 14, 633–647. [Google Scholar] [CrossRef] [Green Version]
- Ekins, S.; Puhl, A.C.; Zorn, K.M.; Lane, T.R.; Russo, D.P.; Klein, J.J.; Hickey, A.J.; Clark, A.M. Exploiting Machine Learning for End-to-End Drug Discovery and Development. Nat. Mater. 2019, 18, 435–441. [Google Scholar] [CrossRef]
- Cristianini, N.; Shawe-Taylor, J. An Introduction to Support Vector Machines and Other Kernel-Based Learning Methods; Cambridge University Press: Cambridge, UK, 2000; ISBN 978-0-521-78019-3. [Google Scholar]
- Shen, M.; Xiao, Y.; Golbraikh, A.; Gombar, V.K.; Tropsha, A. Development and Validation of K-Nearest-Neighbor QSPR Models of Metabolic Stability of Drug Candidates. J. Med. Chem. 2003, 46, 3013–3020. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.; Scheiber, J.; Glick, M.; Davies, J.W.; Azzaoui, K.; Hamon, J.; Urban, L.; Whitebread, S.; Jenkins, J.L. Analysis of Pharmacology Data and the Prediction of Adverse Drug Reactions and Off-Target Effects from Chemical Structure. ChemMedChem 2007, 2, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Susnow, R.G.; Dixon, S.L. Use of Robust Classification Techniques for the Prediction of Human Cytochrome P450 2D6 Inhibition. J. Chem. Inf. Comput. Sci. 2003, 43, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Aliper, A.; Plis, S.; Artemov, A.; Ulloa, A.; Mamoshina, P.; Zhavoronkov, A. Deep Learning Applications for Predicting Pharmacological Properties of Drugs and Drug Repurposing Using Transcriptomic Data. Mol. Pharm. 2016, 13, 2524–2530. [Google Scholar] [CrossRef] [Green Version]
- McGaughey, G.B.; Sheridan, R.P.; Bayly, C.I.; Culberson, J.C.; Kreatsoulas, C.; Lindsley, S.; Maiorov, V.; Truchon, J.-F.; Cornell, W.D. Comparison of Topological, Shape, and Docking Methods in Virtual Screening. J. Chem. Inf. Model. 2007, 47, 1504–1519. [Google Scholar] [CrossRef]
- Ekins, S. The Next Era: Deep Learning in Pharmaceutical Research. Pharm. Res. 2016, 33, 2594–2603. [Google Scholar] [CrossRef]
- Trujillo, C.A.; Gao, R.; Negraes, P.D.; Gu, J.; Buchanan, J.; Preissl, S.; Wang, A.; Wu, W.; Haddad, G.G.; Chaim, I.A.; et al. Complex Oscillatory Waves Emerging from Cortical Organoids Model Early Human Brain Network Development. Cell Stem Cell 2019, 25, 558–569.e7. [Google Scholar] [CrossRef] [PubMed]
- Kegeles, E.; Naumov, A.; Karpulevich, E.A.; Volchkov, P.; Baranov, P. Convolutional Neural Networks Can Predict Retinal Differentiation in Retinal Organoids. Front. Cell. Neurosci. 2020, 14. [Google Scholar] [CrossRef]
- Deng, J.; Dong, W.; Socher, R.; Li, L.; Kai, L.; Li, F.-F. ImageNet: A large-scale hierarchical image database. In Proceedings of the 2009 IEEE Conference on Computer Vision and Pattern Recognition, Miami, FL, USA, 20–25 June 2009; pp. 248–255. [Google Scholar]
- Monzel, A.S.; Hemmer, K.; Kaoma, T.; Smits, L.M.; Bolognin, S.; Lucarelli, P.; Rosety, I.; Zagare, A.; Antony, P.; Nickels, S.L.; et al. Machine Learning-Assisted Neurotoxicity Prediction in Human Midbrain Organoids. Parkinsonism Relat. Disord. 2020, 75, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehmelt, L.; Poplawski, G.; Hwang, E.; Halpain, S. NeuriteQuant: An Open Source Toolkit for High Content Screens of Neuronal Morphogenesis. BMC Neurosci. 2011, 12, 100. [Google Scholar] [CrossRef] [Green Version]
- Pani, G.; Vos, W.H.D.; Samari, N.; de Saint-Georges, L.; Baatout, S.; Oostveldt, P.V.; Benotmane, M.A. MorphoNeuroNet: An Automated Method for Dense Neurite Network Analysis. Cytom. Part A 2014, 85, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Godinez, W.J.; Hossain, I.; Lazic, S.E.; Davies, J.W.; Zhang, X. A Multi-Scale Convolutional Neural Network for Phenotyping High-Content Cellular Images. Bioinformatics 2017, 33, 2010–2019. [Google Scholar] [CrossRef]
- Berryman, S.; Matthews, K.; Lee, J.H.; Duffy, S.P.; Ma, H. Image-Based Phenotyping of Disaggregated Cells Using Deep Learning. Commun. Biol. 2020, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Belthangady, C.; Royer, L.A. Applications, Promises, and Pitfalls of Deep Learning for Fluorescence Image Reconstruction. Nat. Methods 2019, 16, 1215–1225. [Google Scholar] [CrossRef]
- Kulikov, V.; Guo, S.-M.; Stone, M.; Goodman, A.; Carpenter, A.; Bathe, M.; Lempitsky, V. DoGNet: A Deep Architecture for Synapse Detection in Multiplexed Fluorescence Images. PLoS Comput. Biol. 2019, 15, e1007012. [Google Scholar] [CrossRef] [PubMed]
- Soltanian-Zadeh, S.; Sahingur, K.; Blau, S.; Gong, Y.; Farsiu, S. Fast and Robust Active Neuron Segmentation in Two-Photon Calcium Imaging Using Spatiotemporal Deep Learning. Proc. Natl. Acad. Sci. USA 2019, 116, 8554–8563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, K.; Lyu, Y.; Pan, H.; Zhang, J.; Stambolian, D.; Susztak, K.; Reilly, M.P.; Hu, G.; Li, M. Deep Learning Enables Accurate Clustering with Batch Effect Removal in Single-Cell RNA-Seq Analysis. Nat. Commun. 2020, 11, 2338. [Google Scholar] [CrossRef]
- Tian, T.; Wan, J.; Song, Q.; Wei, Z. Clustering Single-Cell RNA-Seq Data with a Model-Based Deep Learning Approach. Nat. Mach. Intell. 2019, 1, 191–198. [Google Scholar] [CrossRef]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep Learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef]
- Lopez, R.; Regier, J.; Cole, M.B.; Jordan, M.I.; Yosef, N. Deep Generative Modeling for Single-Cell Transcriptomics. Nat. Methods 2018, 15, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Fan, J.; Emanuel, G.; Hao, J.; Zhuang, X. Spatial Transcriptome Profiling by MERFISH Reveals Subcellular RNA Compartmentalization and Cell Cycle-Dependent Gene Expression. Proc. Natl. Acad. Sci. USA 2019, 116, 19490–19499. [Google Scholar] [CrossRef] [Green Version]
- Eraslan, G.; Simon, L.M.; Mircea, M.; Mueller, N.S.; Theis, F.J. Single-Cell RNA-Seq Denoising Using a Deep Count Autoencoder. Nat. Commun. 2019, 10, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.E.; Georgescu, A.; Huh, D. Organoids-on-a-Chip. Science 2019, 364, 960–965. [Google Scholar] [CrossRef]
Figure 1.
The conventional and iPSC-based drug discovery pipeline.

Figure 2.
Genome editing of organoids using Clustered Regularly Interspaced Short Palindromic RepeatsCRISPR-Cas9technology.
Figure 2.
Genome editing of organoids using Clustered Regularly Interspaced Short Palindromic RepeatsCRISPR-Cas9technology.

Figure 3.
Single-cell RNA sequencing approaches to investigate cell heterogeneity in human organoids.
Figure 3.
Single-cell RNA sequencing approaches to investigate cell heterogeneity in human organoids.


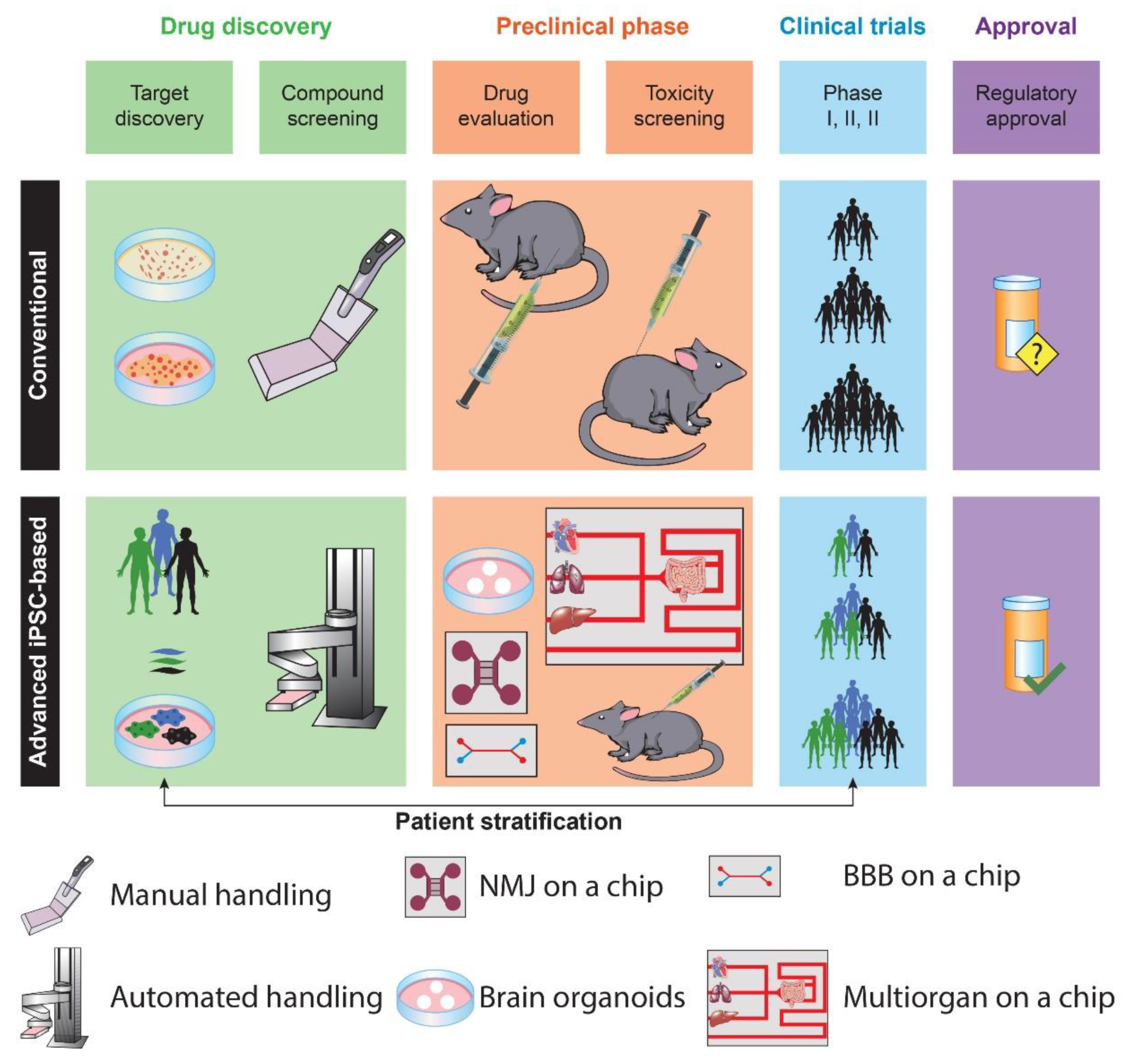{kind=link}
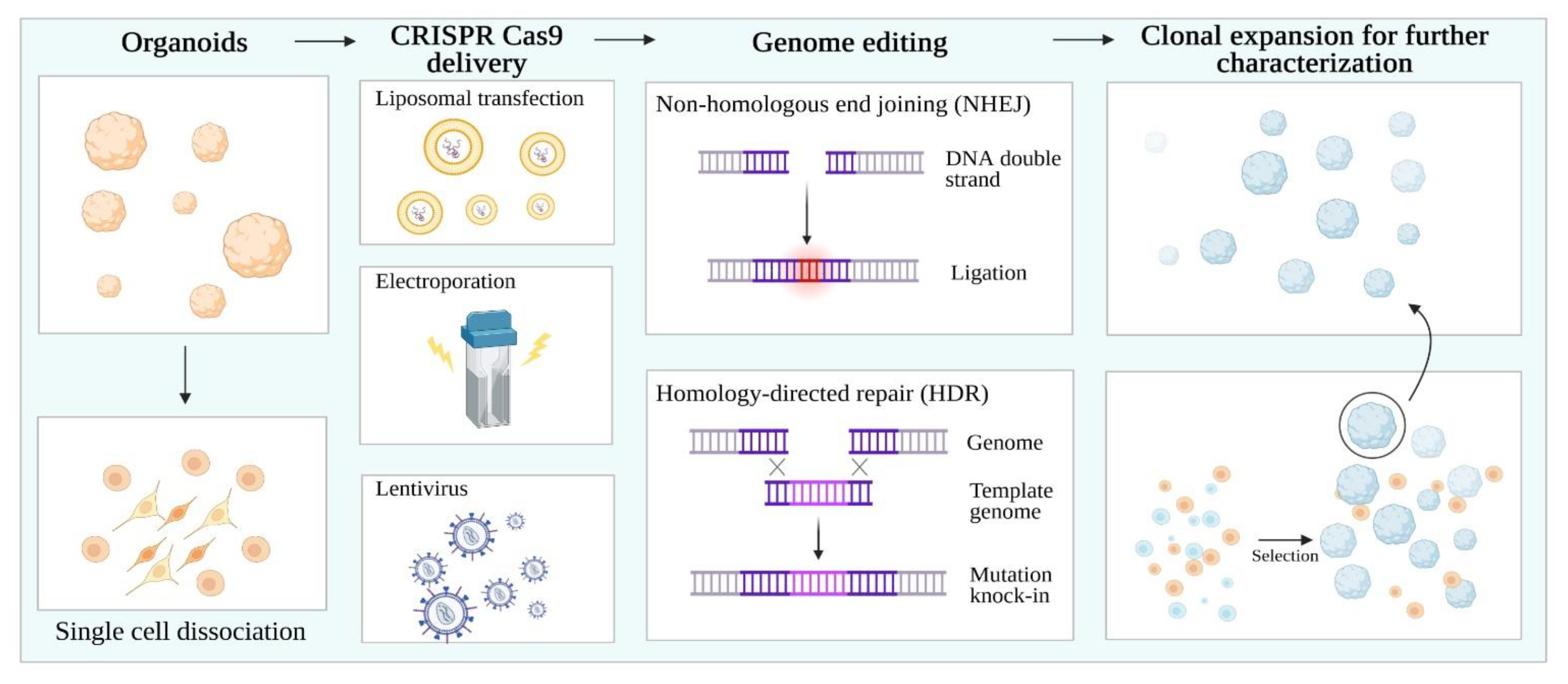{kind=link}
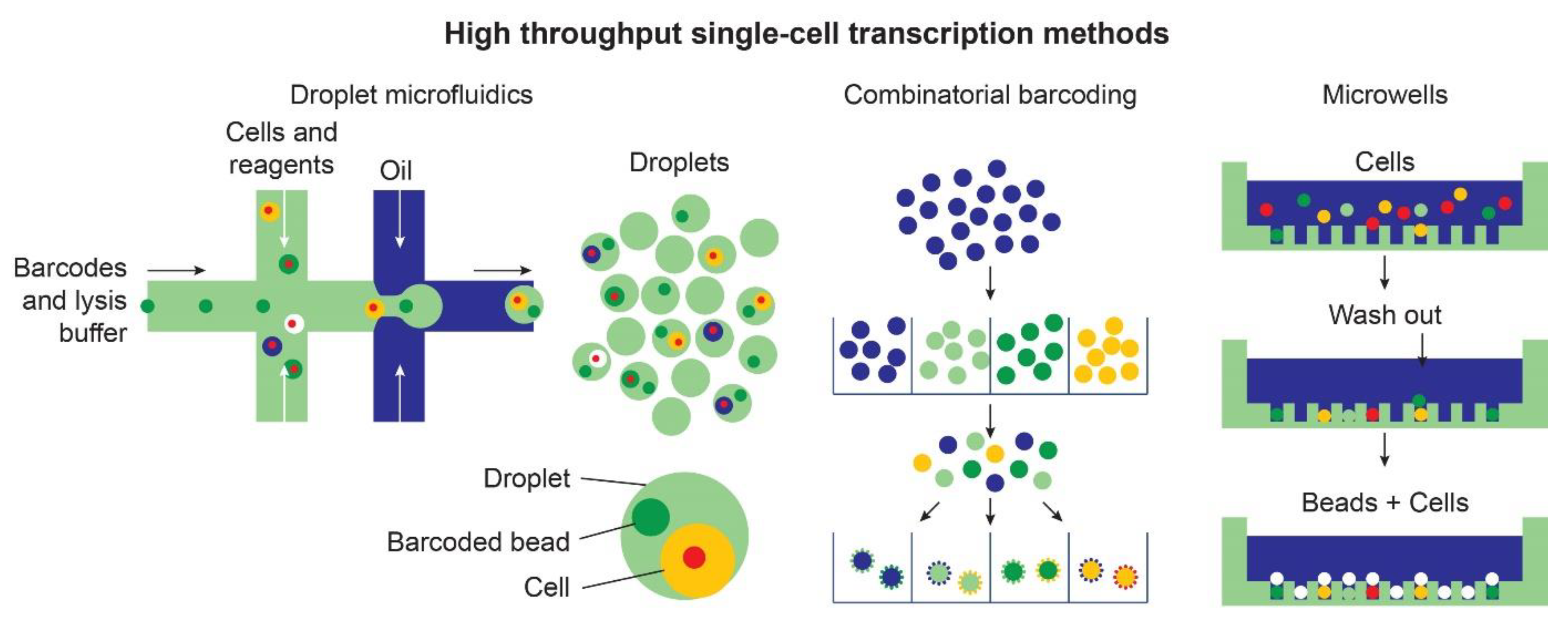{kind=link}
Table 1.
Selected studies using neural organoids to model neurological diseases.
| Disease Group | Diseases Modeled | Organoid Type | Intervention | Disease Phenotype | References |
|---|---|---|---|---|---|
| Neurodevelopmental | Primary microcephaly | Undirected | Electroporation-mediated CDK5RAP2 overexpression and knockdown (with shRNAs) | Smaller organoids; reduced number of radial glial stem cells; premature neural differentiation; rescue by shRNA for CDK5RAP2 | [12] |
| ASPM-related microcephaly | Forebrain | Electroporation-mediated knockdown of Aspm | Poorly organized neuroepithelium, reduced number of radial glial cells, and defective cortical layer lamination | [26] | |
| Seckel syndrome | Undirected | Small molecule-mediated and siRNA-mediated inhibition of the cilium disassembly complex | Smaller size and premature neuronal differentiation; increased number and length of cilium; retarded cilium disassembly. | [27] | |
| Periventricular heterotopia | Undirected | Induced pluripotent stem cell (iPSC)-derived organoids from patients carrying cadherin receptor-ligand DCHS1 and FAT4 gene mutations | Morphological alterations, defective migration, axonal guidance, and patterning of neural progenitors. | [28] | |
| Neonatal hypoxic injury | Forebrain | Low oxygen exposure controlled with a gas chamber and fiber-optic microsensors | Impairment of intermediate progenitors in the subventricular zone; altered activation of cellular stress response pathways and early neuronal differentiation; phenotype rescue by stress response inhibitor (ISRIB) | [29] | |
| Down syndrome | Ventral forebrain/chimeric mice | iPSC-derived organoids from patients | Increased expression of GABAergic interneurons; shRNA-knockdown of OLIG2 rescues phenotype in vitro and ameliorates behavioral deficits in mice | [30] | |
| Neurodegenerative | Alzheimer disease | Forebrain | Herpes Simplex virus-1 (HSV-1) -infected organoids derived from human induced neural stem cells seeded into a biomaterial-based scaffold. | Aβ plaque formation, neuronal loss, reactive gliosis, neuroinflammation, and diminished neural network functionality | [31] |
| Parkinson disease (PD) | Midbrain | iPSC-derived organoids from patients carrying the LRRK2-GS019S mutation data | Recapitulation of PD-specific pathological signatures, including increased α-synuclein. Upregulation of TXNIP favored disease phenotype. | [32] | |
| Creutzfeld-jakob disease (CJD) | Undirected | Cellular inoculation within the organoids with human brain homogenates from sporadic CJD subtypes | Reduced neuronal metabolism, deposition of insoluble protease-resistant (PrP) aggregates with seeding activity, over activation of astrocytes | [33] | |
| Brain Tumors | Glioblastoma | Forebrain | Electroporation-mediated plasmid infection leading to overexpression of MYC and inhibition of tumor suppressor genes | Glioma-like cells with poor differentiation, increased cell proliferation and disrupted cytoarchitecture, downregulation of PI3K-AK and RAS pathways, tumor invasion upon animal transplantation, partial rescue in glioma-like organoids using EGFR-inhibitors | [34] |
| Infectious | Herpes Simplex virus | Forebrain | iPSC-derived organoids infected with HSV-1 | HSV-1 transportation from the periphery to the central layers of the organoids, spontaneous reactivation of viral latent infection | [35] |
| Cytomegalovirus (CMV) | Undirected | CMV-infected hiPSCs differentiated into neural organoids | Impaired cell proliferation, tissue degeneration with necrosis, vacuoles, and cysts, disrupted cortical lamination | [36] | |
| Zikavirus (ZIKV) | Forebrain | Treatment with RNA interfering (RNAi) enhancer (enoxacin) | Microcephaly-like phenotype, production of small interfering RNAs in neural progenitors upon infection, viral clearance using enoxacin | [37] | |
| Psychiatric | Schizophrenia | Forebrain | iPSC and embryonic stem cell (ESC)-derived organoids from patients. Chemically mediated neuronal (n)FGFR modulation. | Abnormal neuronal migration, particularly towards the cortex, decreased intraneuronal intracortical connectivity. Both loss of function and hyperactivity of neuronal (n)FGFR1 affects cortical neurodevelopment. | [38] |
| Prenatal toxin exposure | Nicotine | Undirected | Organoids cultured with microfluidic devices. 3D cultures exposed to a physiologically relevant concentration of nicotine. | Premature neuronal differentiation, disrupted brain regional organization, abnormal cortical development, and neuronal outgrowth, | [39] |
| Methamphetamine | Undirected | 10-month-old ESC-derived organoids treated with methamphetamine for a week | Increased expression of astrocyte-specific gene networks related to inflammations. Upregulation of genes involved in complement activation, apoptosis, and immune response. | [40] | |
| Cannabis | Undirected | ESC-derived organoids grown in a microfluidic platform and perfused for 27 days with Δ-9-tetrahydrocannabinol (THC) | Reduced neuronal maturation and spontaneous firing, downregulation of cannabinoid receptor type 1 (CB1) receptors, and impaired neurite outgrowth. | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Costamagna, G.; Comi, G.P.; Corti, S. Advancing Drug Discovery for Neurological Disorders Using iPSC-Derived Neural Organoids. Int. J. Mol. Sci. 2021, 22, 2659. https://doi.org/10.3390/ijms22052659
AMA Style
Costamagna G, Comi GP, Corti S. Advancing Drug Discovery for Neurological Disorders Using iPSC-Derived Neural Organoids. International Journal of Molecular Sciences. 2021; 22(5):2659. https://doi.org/10.3390/ijms22052659
Chicago/Turabian StyleCostamagna, Gianluca, Giacomo Pietro Comi, and Stefania Corti. 2021. "Advancing Drug Discovery for Neurological Disorders Using iPSC-Derived Neural Organoids" International Journal of Molecular Sciences 22, no. 5: 2659. https://doi.org/10.3390/ijms22052659
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.