
Extensive Histopathological Characterization of Inflamed Bowel in the Dextran Sulfate Sodium Mouse Model with Emphasis on Clinically Relevant Biomarkers and Targets for Drug Development

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results
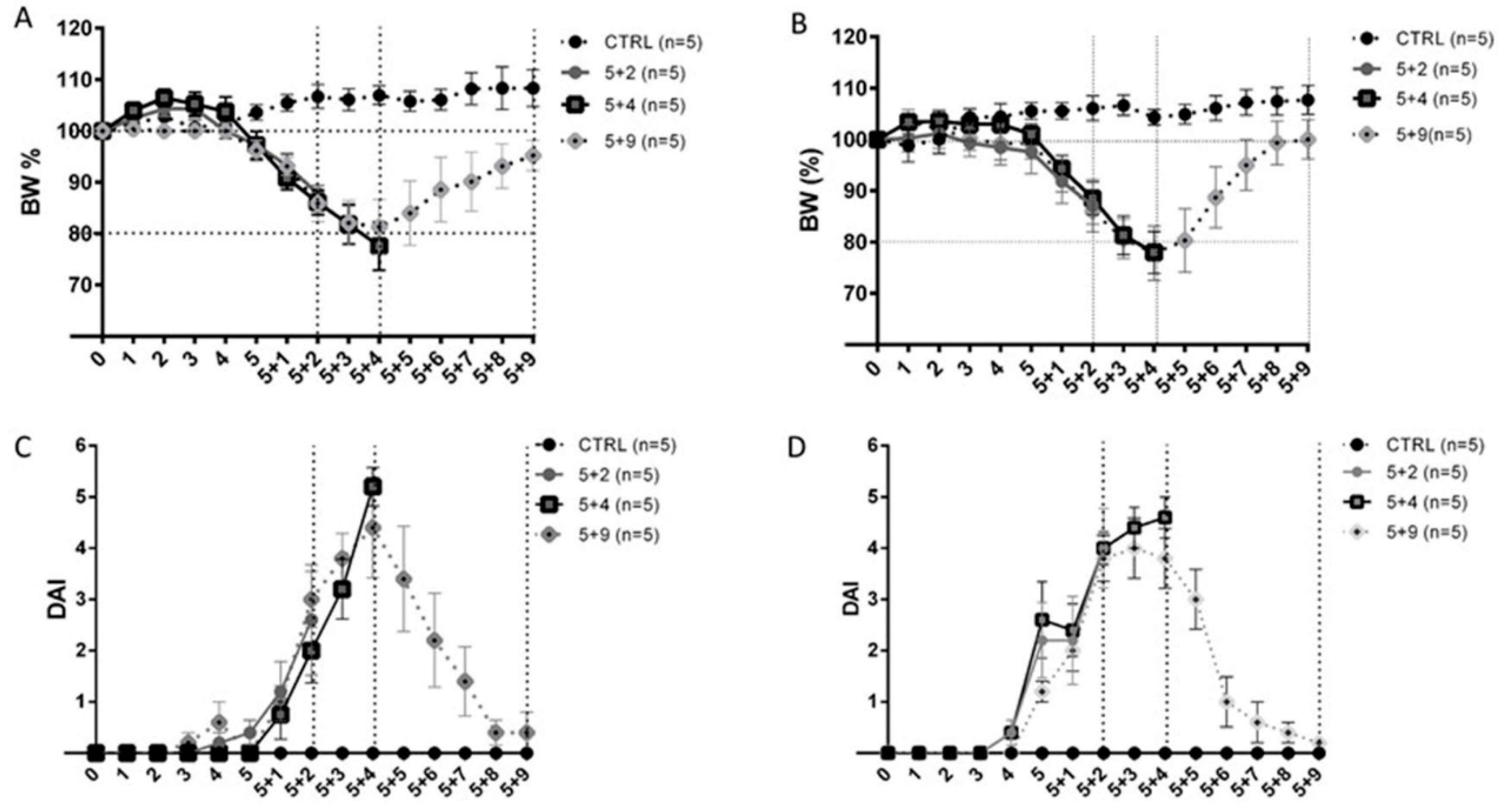
2.1. Clinical Assessment of Dextran Sulfate Sodium-Induced Colitis
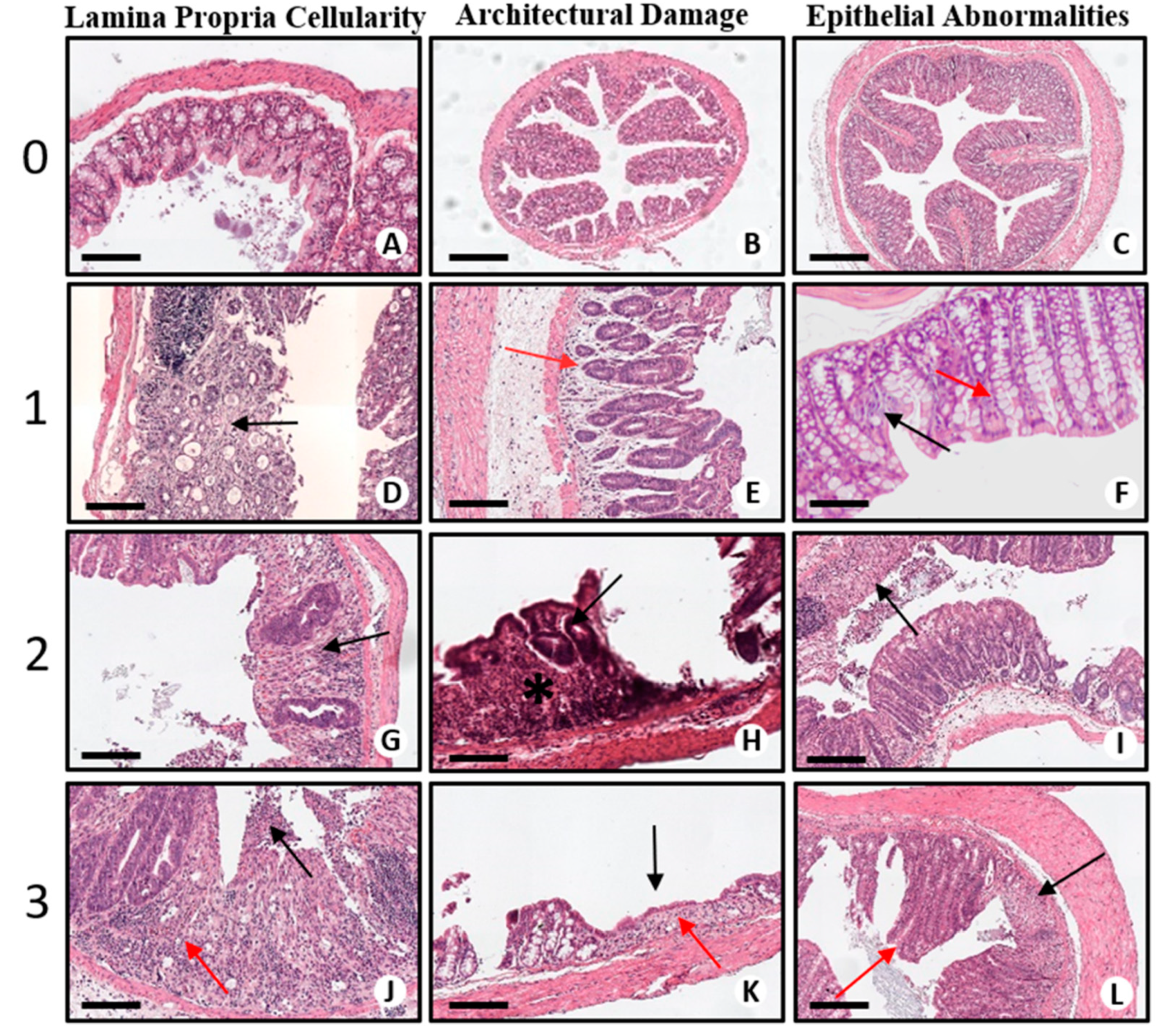
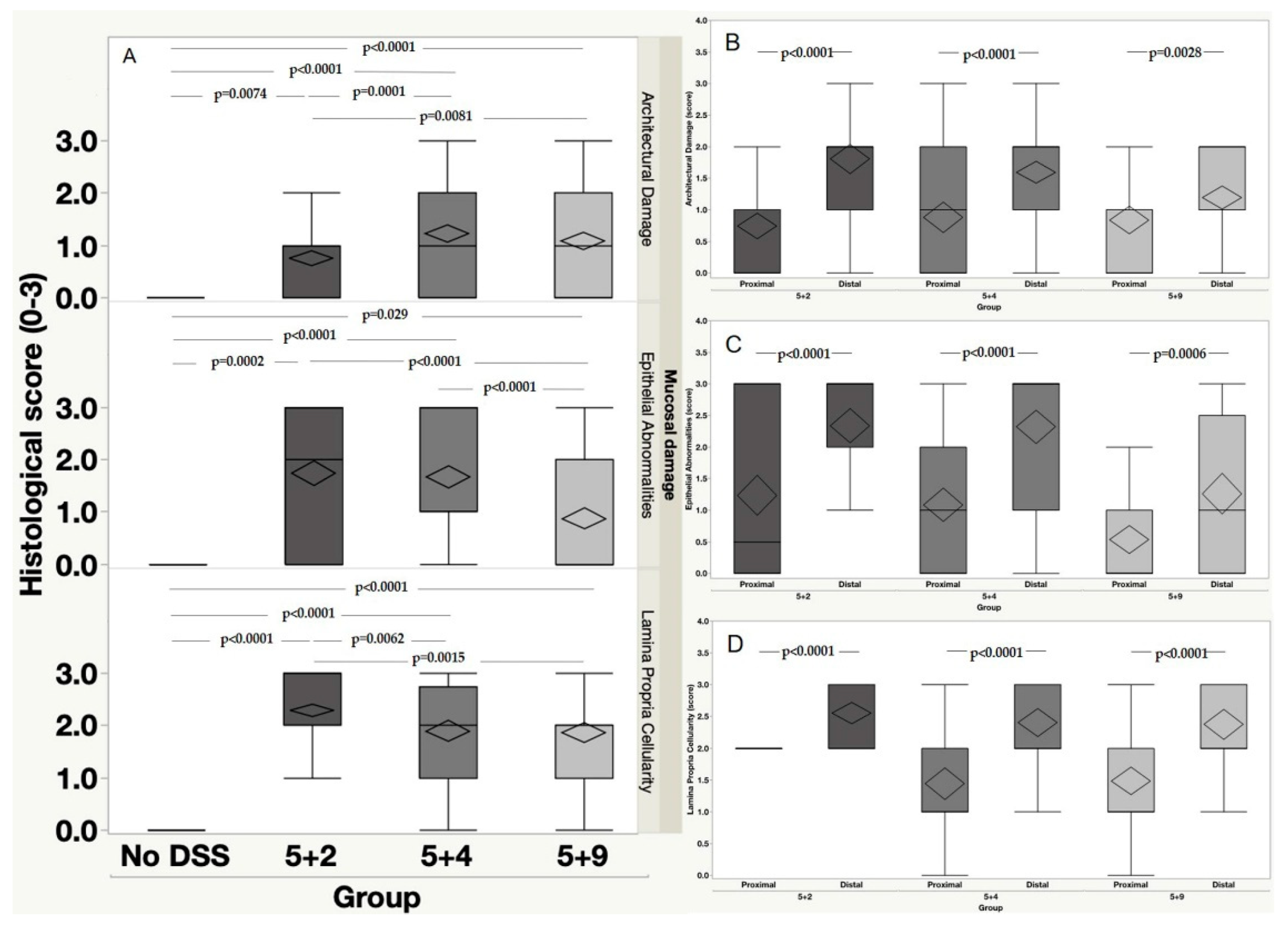
2.2. Histopathological Assessment of DSS-Induced Colitis
Mucosal Damage
2.3. Anatomical Distribution of the Mucosal Damage
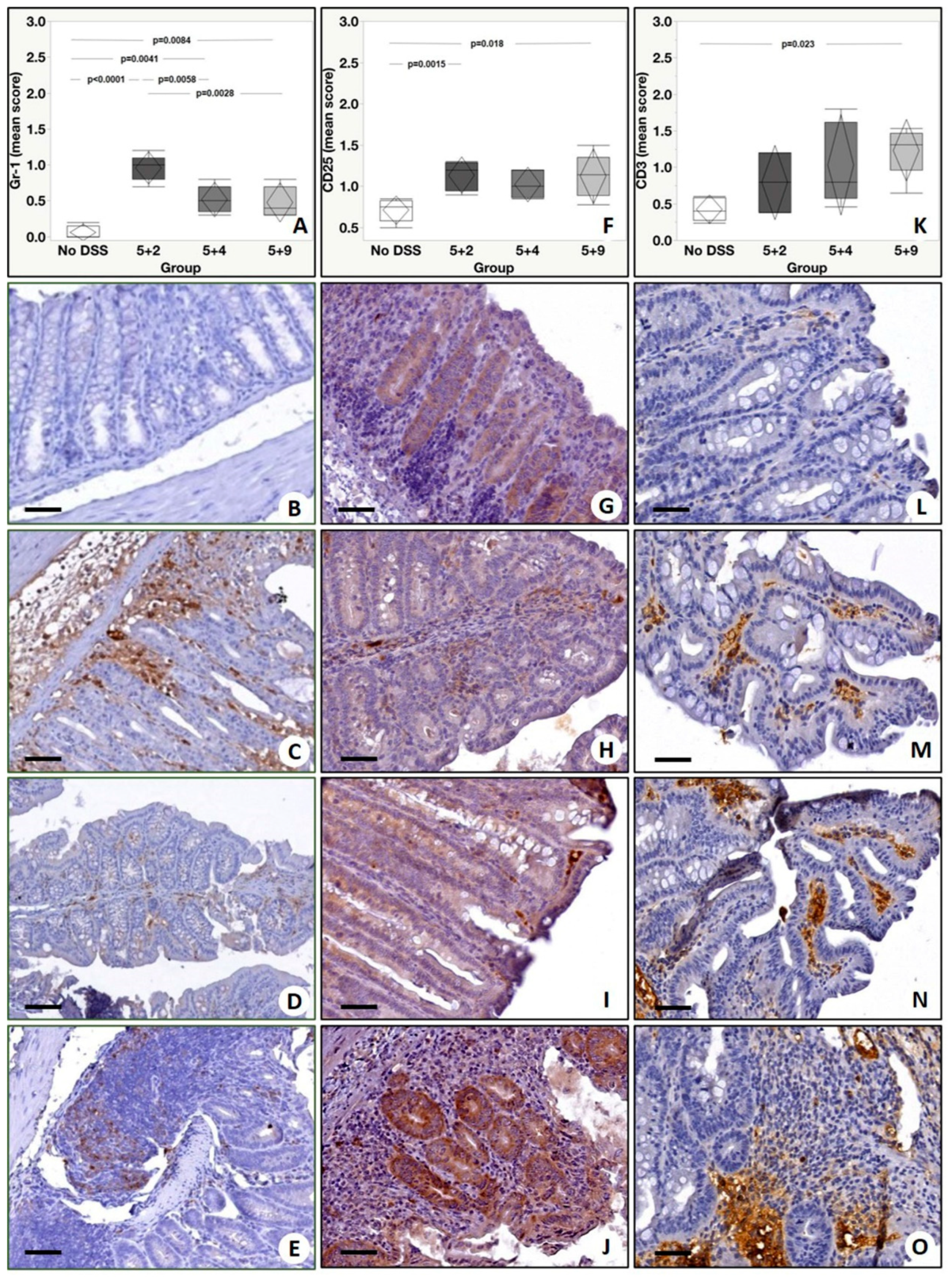
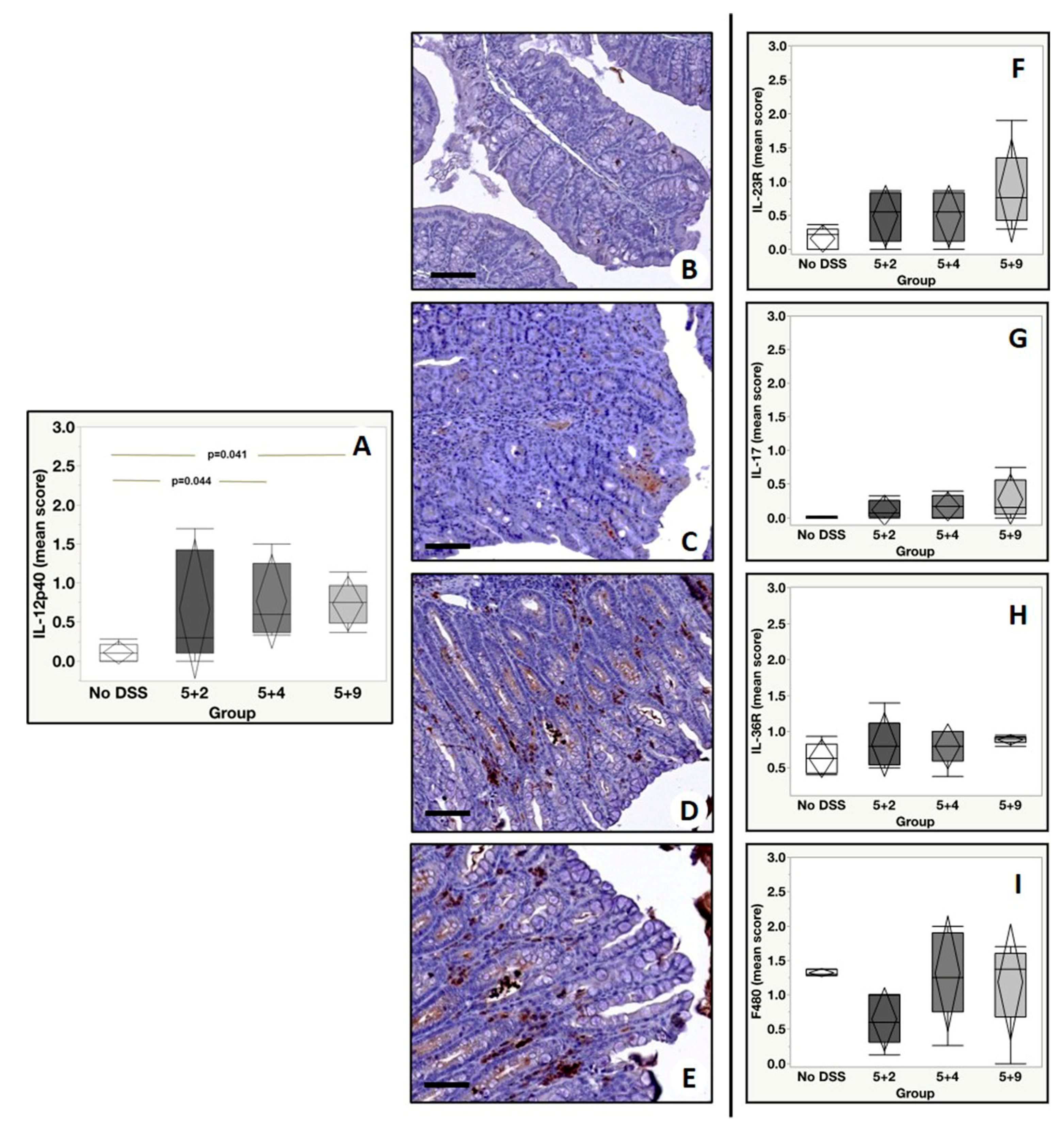
2.4. Immunohistochemistry
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Mice and Maintenance
4.3. Tissue Processing
4.4. Tissue Staining
4.5. Scoring System
4.6. Immunohistochemistry
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CD | Crohn’s disease |
| CS | Clinical Score |
| DAB | 3,3′-Diaminobenzidine |
| DAI | Diseases Activity Index |
| DSS | Dextran Sulfate Sodium |
| ECCO | European Crohn’s and Colitis Organisation |
| H&E | Hematoxylin and Eosin |
| HRP | Horseradish Peroxidase |
| IACUC | Institutional Animal Care and Use Committees |
| IBD | Inflammatory Bowel Disease |
| IL | Interleukin |
| PET | Positron Emission Tomography |
| SPECT | Single-Photon Emission Computed Tomography |
| TNF | Tumor Necrosis Factor |
| UC | Ulcerative Colitis |
| VEGFR | Vascular Endothelial Growth Factor |
References
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2018, 390, 2769–2778. [Google Scholar] [CrossRef]
- Kolios, G. Animal models of inflammatory bowel disease: How useful are they really? Curr. Opin. Gastroenterol. 2016, 32, 251–257. [Google Scholar] [CrossRef]
- DeVoss, J.; Diehl, L. Murine Models of Inflammatory Bowel Disease (IBD): Challenges of Modeling Human Disease. Toxicol. Pathol. 2014, 42, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Valatas, V.; Vakas, M.; Kolios, G. The value of experimental models of colitis in predicting efficacy of biological therapies for inflammatory bowel diseases. Am. J. Physiol. Gastrointest Liver Physiol. 2013, 305, 763–785. [Google Scholar] [CrossRef]
- Ward, J.M.; Schofield, P.N.; Sundberg, J.P. Reproducibility of histopathological findings in experimental pathology of the mouse: A sorry tail. Lab Anim. 2017, 46, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Auletta, S.; Bonfiglio, R.; Wunder, A.; Varani, M.; Galli, F.; Borri, F.; Scimeca, M.; Niessen, H.G.; Schönberger, T.; Bonanno, E. Animal models for the study of inflammatory bowel diseases: A meta-analysis on modalities for imaging inflammatory lesions. Q. J. Nucl. Med. Mol. Imaging 2018, 621, 78–100. [Google Scholar]
- Magro, F.; Langner, C.; Driessen, A.; Ensari, A.; Geboes, K.; Mantzaris, G.J.; Villanacci, V.; Becheanu, G.; Borralho Nunes, P.; Cathomas, G.; et al. European consensus on the histopathology of inflammatory bowel disease. J. Crohns Colitis. 2013, 7, 827–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonovas, S.; Pantavou, K.; Evripidou, D.; Bastiampillai, A.J.; Nikolopoulos, G.K.; Peyrin-Biroulet, L.; Danese, S. Safety of biological therapies in ulcerative colitis: An umbrella review of meta-analyses. Best Pract. Res. Clin. Gastroenterol. 2018, 32–33, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Juillerat, P.; Manz, M.; Sauter, B.; Zeitz, J.; Vavricka, S.R.; Swiss IBDnet, an Official Working Group of the Swiss Society of Gastroenterology. Therapies in Inflammatory Bowel Disease Patients with Extraintestinal Manifestations. Digestion 2020, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Digby-Bell, J.L.; Atreya, R.; Monteleone, G.; Powell, N. Interrogating host immunity to predict treatment response in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 9–20. [Google Scholar] [CrossRef]
- Johansson, M.E.; Gustafsson, J.K.; Sjöberg, K.E.; Petersson, J.; Holm, L.; Sjövall, H.; Hansson, G.C. Bacteria penetrate the inner mucus layer before inflammation in the dextran sulfate colitis model. PLoS ONE 2010, 5, e12238. [Google Scholar] [CrossRef] [Green Version]
- Cooper, H.S.; Murthy, S.N.; Shah, R.S.; Sedergran, D.J. Clinicopathologic study of dextran sulfate sodium experimental murine colitis. Lab Invest. 1993, 69, 238–249. [Google Scholar] [PubMed]
- Yan, Y.; Kolachala, V.; Dalmasso, G.; Nguyen, H.; Laroui, H.; Sitaraman, S.V.; Merlin, D. Temporal and spatial analysis of clinical and molecular parameters in dextran sodium sulfate induced colitis. PLoS ONE 2009, 4, e6073. [Google Scholar] [CrossRef]
- Moolenbeek, C.; Ruitenberg, E.J. The “Swiss roll”: A simple technique for histological studies of the rodent intestine. Lab Anim. 1981, 15, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104. [Google Scholar] [CrossRef] [PubMed]
- Kühl, A.A.; Kakirman, H.; Janotta, M.; Dreher, S.; Cremer, P.; Pawlowski, N.N.; Loddenkemper, C.; Heimesaat, M.M.; Grollich, K.; Zeitz, M.; et al. Aggravation of different types of experimental colitis by depletion or adhesion blockade of neutrophils. Gastroenterology 2007, 1336, 1882–1892. [Google Scholar] [CrossRef] [PubMed]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef]
- Braegger, C.P.; Nicholls, S.; Murch, S.H.; Stephens, S.; MacDonald, T.T. Tumour necrosis factor alpha in stool as a marker of intestinal inflammation. Lancet 1992, 339, 89–91. [Google Scholar] [CrossRef]
- Reimund, J.M.; Wittersheim, C.; Dumont, S.; Muller, C.D.; Kenney, J.S.; Baumann, R.; Poindron, P.; Duclos, B. Increased production of tumour necrosis factor-alpha interleukin-1 beta, and interleukin-6 by morphologically normal intestinal biopsies from patients with Crohn’s disease. Gut 1996, 39, 684–689. [Google Scholar] [CrossRef] [Green Version]
- Sabino, J.; Verstockt, B.; Vermeire, S.; Ferrante, M. New biologics and small molecules in inflammatory bowel disease: An update. Therap. Adv. Gastroenterol. 2019, 12, 1756284819853208. [Google Scholar] [CrossRef] [Green Version]
- Jovani, M.; Danese, S. Vedolizumab for the treatment of IBD: A selective therapeutic approach targeting pathogenic a4b7 cells. Curr. Drug Targets. 2013, 14, 1433–1443. [Google Scholar] [CrossRef]
- Cherry, L.N.; Yunker, N.S.; Lambert, E.R.; Vaughan, D.; Lowe, D.K. Vedolizumab: An α4β7 integrin antagonist for ulcerative colitis and Crohn’s disease. Ther. Adv. Chronic. Dis. 2015, 6, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Benelli, R.; Lorusso, G.; Albini, A.; Noonan, D.M. Cytokines and chemokines as regulators of angiogenesis in health and disease. Curr. Pharm. Des. 2006, 12, 3101–3115. [Google Scholar] [CrossRef] [PubMed]
- Pousa, I.D.; Maté, J.; Gisbert, J.P. Angiogenesis in inflammatory bowel disease. Eur. J. Clin. Invest. 2008, 38, 73–81. [Google Scholar] [CrossRef]
- Scaldaferri, F.; Vetrano, S.; Sans, M.; Arena, V.; Straface, G.; Stigliano, E.; Repici, A.; Sturm, A.; Malesci, A.; Panes, J.; et al. VEGF-A links angiogenesis and inflammation in inflammatory bowel disease pathogenesis. Gastroenterology 2009, 136, 585–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taurone, S.; Galli, F.; Signore, A.; Agostinelli, E.; Dierckx, R.A.; Minni, A.; Pucci, M.; Artico, M. VEGF in nuclear medicine: Clinical application in cancer and future perspectives (Review). Int. J. Oncol. 2016, 49, 437–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buell, M.G.; Berin, M.C. Neutrophil-independence of the initiation of colonic injury. Comparison of results from three models of experimental colitis in the rat. Dig. Dis. Sci. 1994, 39, 2575–2588. [Google Scholar] [CrossRef] [PubMed]
- Natsui, M.; Kawasaki, K.; Takizawa, H.; Hayashi, S.I.; Matsuda, Y.; Sugimura, K.; Seki, K.; Narisawa, R.; Sendo, F.; Asakura, H. Selective depletion of neutrophils by a monoclonal antibody, RP-3, suppresses dextran sulphate sodium-induced colitis in rats. J. Gastroenterol. Hepatol. 1997, 12, 801–808. [Google Scholar] [CrossRef]
- Zhang, R.; Ito, S.; Nishio, N.; Cheng, Z.; Suzuki, H.; Isobe, K. Up-regulation of Gr1 + CD11b+ population in spleen of dextran sulfate sodium administered mice works to repair colitis. Inflamm. Allergy Drug Targets. 2011, 10, 39–46. [Google Scholar] [CrossRef]
- Locke, L.W.; Chordia, M.D.; Zhang, Y.; Kundu, B.; Kennedy, D.; Landseadel, J.; Xiao, L.; Fairchild, K.D.; Berr, S.S.; Linden, J.; et al. A novel neutrophil-specific PET imaging agent: cFLFLFK-PEG-64Cu. J. Nucl. Med. 2009, 50, 790–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Xiao, L.; Chordia, M.D.; Locke, L.W.; Williams, M.B.; Berr, S.S.; Pan, D. Neutrophil targeting heterobivalent SPECT imaging probe: cFLFLF-PEG-TKPPR-99mTc. Bioconjug. Chem. 2010, 21, 1788–1793. [Google Scholar] [CrossRef]
- Spier, B.J.; Perlman, S.B.; Jaskowiak, C.J.; Reichelderfer, M. PET/CT in the evaluation of inflammatory bowel disease: Studies in patients before and after treatment. Mol. Imaging Biol. 2010, 12, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Hartimath, S.V.; Draghiciu, O.; van de Wall, S.; Manuelli, V.; Dierckx, R.A.; Nijman, H.W.; Daemen, T.; de Vries, E.F. Noninvasive monitoring of cancer therapy induced activated T cells using [18F] FB-IL-2 PET imaging. Oncoimmunology 2016, 6, e1248014. [Google Scholar] [CrossRef] [Green Version]
- Loose, D.; Signore, A.; Bonanno, E.; Vermeersch, H.; Dierckx, R.; Deron, P.; Van de Wiele, C. Prognostic value of CD25 expression on lymphocytes and tumor cells in squamous-cell carcinoma of the head and neck. Cancer Biother. Radiopharm. 2008, 23, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marafini, I.; Angelucci, E.; Pallone, F.; Monteleone, G. The IL-12/23/STAT Axis as a Therapeutic Target in Inflammatory Bowel Disease: Mechanisms and Evidence in Man. Dig. Dis. 2015, 33 (Suppl. 1), 113–119. [Google Scholar] [CrossRef] [Green Version]
- Breynaert, C.; Dresselaers, T.; Perrier, C.; Arijs, I.; Cremer, J.; Van Lommel, L.; Van Steen, K.; Ferrante, M.; Schuit, F.; Vermeire, S.; et al. Unique gene expression and MR T2 relaxometry patterns define chronic murine dextran sodium sulphate colitis as a model for connective tissue changes in human Crohn’s disease. PLoS ONE 2013, 8, e68876. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, J.R.; Brown, W.A.; Smith, C.L.; Byrne, F.R.; Viney, J.L. Methods of inducing inflammatory bowel disease in mice. Curr. Protoc. Pharmacol. 2009. [Google Scholar] [CrossRef]
- Bonfiglio, R.; Milano, F.; Cranga, A.; De Caro, M.T.; Lamsira, H.K.; Trivigno, D.; Urso, S.; Scimeca, M.; Bonanno, E. Negative prognostic value of intra-ductal fat infiltrate in breast cancer. Pathol. Res. Pract. 2019, 215, 11152634. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Lamina Propria Cellularity | Architectural Damage | Epithelial Abnormalities |
|---|---|---|---|
| 0 | Normal presence of mononuclear inflammatory cells (lymphocytes, plasma cells, eosinophils) in lamina propria. | Normal crypts (straight, parallel and extend from immediately above the muscularis mucosae) | No signs of epithelial damage. |
| 1 | Mild increase in mononuclear inflammatory cells in the lamina propria. | Mild crypt distortion (includes loss of parallelism between crypts, variation in size and shape) | Mucin depletion (reduction in number of goblet cells and/or intracellular mucin) and/or repairing/reactive changes (may include nuclear enlargement, loss of nuclear polarity, prominent nucleoli, presence of mitotic figures). |
| 2 | Striking increase in mononuclear inflammatory cells and possible presence of a minor neutrophilic component in the lamina propria | Moderate crypt distortion (more evident crypt distortion) and shortening: ≤1/3 of crypt height from muscularis mucosae | Mucin depletion, repairing/reactive changes and erosions (epithelial necrosis involving epithelium mainly superficially). |
| 3 | Striking increase in mononuclear inflammatory cells and easily recognizable presence of neutrophils in the lamina propria. | Severe crypt distortion and shortening of ≥1/2 of crypt height from muscularis mucosae (atrophy). Additional evidence: wider spacing of crypts | Mucin depletion, repairing/reactive changes and ulcers (epithelial necrosis involving the total thickness of the mucosa). |
| Antibody | Characteristics | Dilution | Retrieval |
|---|---|---|---|
| Anti-TNF-α | Rat monoclonal, clone XT3.11; BioXCell, NH, USA | 1:1000 | Citrate pH 6.0 |
| Anti-α4β7 | Rat monoclonal, clone DATK32; BioXCell, NH, USA) | 1:700 | EDTA–Citrate Ph 7.8 |
| Anti-IL-12B p40 | Rat monoclonal, clone C17.8; Santa Cruz Biotechnology, Texas, USA | 1:100 | Citrate pH 6.0 |
| anti-CD3 | mouse monoclonal clone V9; Ventana, Tucson, AZ, USA | Pre-diluted | EDTA citrate pH 7.8 |
| anti-CD25 | Rabbit monoclonal, clone SP176; Spring Bioscience, Pleasanton, CA, USA | 1:50 | EDTA–Citrate pH 7.8 |
| anti-IL-17A | Rat monoclonal, clone TC11-18H10; Novus Biologicals, Littleton, CO, USA | 1:50 | EDTA–Citrate pH 7.8 |
| anti-Ly6G (Gr-1) | Mouse monoclonal, clone RB6-85C; Novus Biologicals, Littleton, CO, USA | 1:100 | EDTA–Citrate pH 7.8 |
| anti-IL-36R | IgG2a anti-mouse IL36 with mutated Fc portion Boehringer Ingelheim, Germany | 1:100 | Citrate pH 6.0 |
| anti-FLK1 (VEGFRII) | Rabbit polyclonal; Spring Bioscience, Pleasanton, CA, USA. | 1:100 | Citrate pH 6.0 |
| anti-F4/80 | Rat monoclonal, clone CI:A3-1; BioXCell, NH, USA | 1:500 | EDTA–Citrate pH 7.8 |
| Anti-IL-23R | Rabbit polyclonal, Novus Biologicals, Littleton, CO, USA | 1:100 | Citrate pH 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonfiglio, R.; Galli, F.; Varani, M.; Scimeca, M.; Borri, F.; Fazi, S.; Cicconi, R.; Mattei, M.; Campagna, G.; Schönberger, T.; et al. Extensive Histopathological Characterization of Inflamed Bowel in the Dextran Sulfate Sodium Mouse Model with Emphasis on Clinically Relevant Biomarkers and Targets for Drug Development. Int. J. Mol. Sci. 2021, 22, 2028. https://doi.org/10.3390/ijms22042028
Bonfiglio R, Galli F, Varani M, Scimeca M, Borri F, Fazi S, Cicconi R, Mattei M, Campagna G, Schönberger T, et al. Extensive Histopathological Characterization of Inflamed Bowel in the Dextran Sulfate Sodium Mouse Model with Emphasis on Clinically Relevant Biomarkers and Targets for Drug Development. International Journal of Molecular Sciences. 2021; 22(4):2028. https://doi.org/10.3390/ijms22042028
Chicago/Turabian StyleBonfiglio, Rita, Filippo Galli, Michela Varani, Manuel Scimeca, Filippo Borri, Sara Fazi, Rosella Cicconi, Maurizio Mattei, Giuseppe Campagna, Tanja Schönberger, and et al. 2021. "Extensive Histopathological Characterization of Inflamed Bowel in the Dextran Sulfate Sodium Mouse Model with Emphasis on Clinically Relevant Biomarkers and Targets for Drug Development" International Journal of Molecular Sciences 22, no. 4: 2028. https://doi.org/10.3390/ijms22042028