Human Adipose-Derived Stem Cells’ Paracrine Factors in Conditioned Medium Can Enhance Porcine Oocyte Maturation and Subsequent Embryo Development


Abstract
:1. Introduction
2. Results
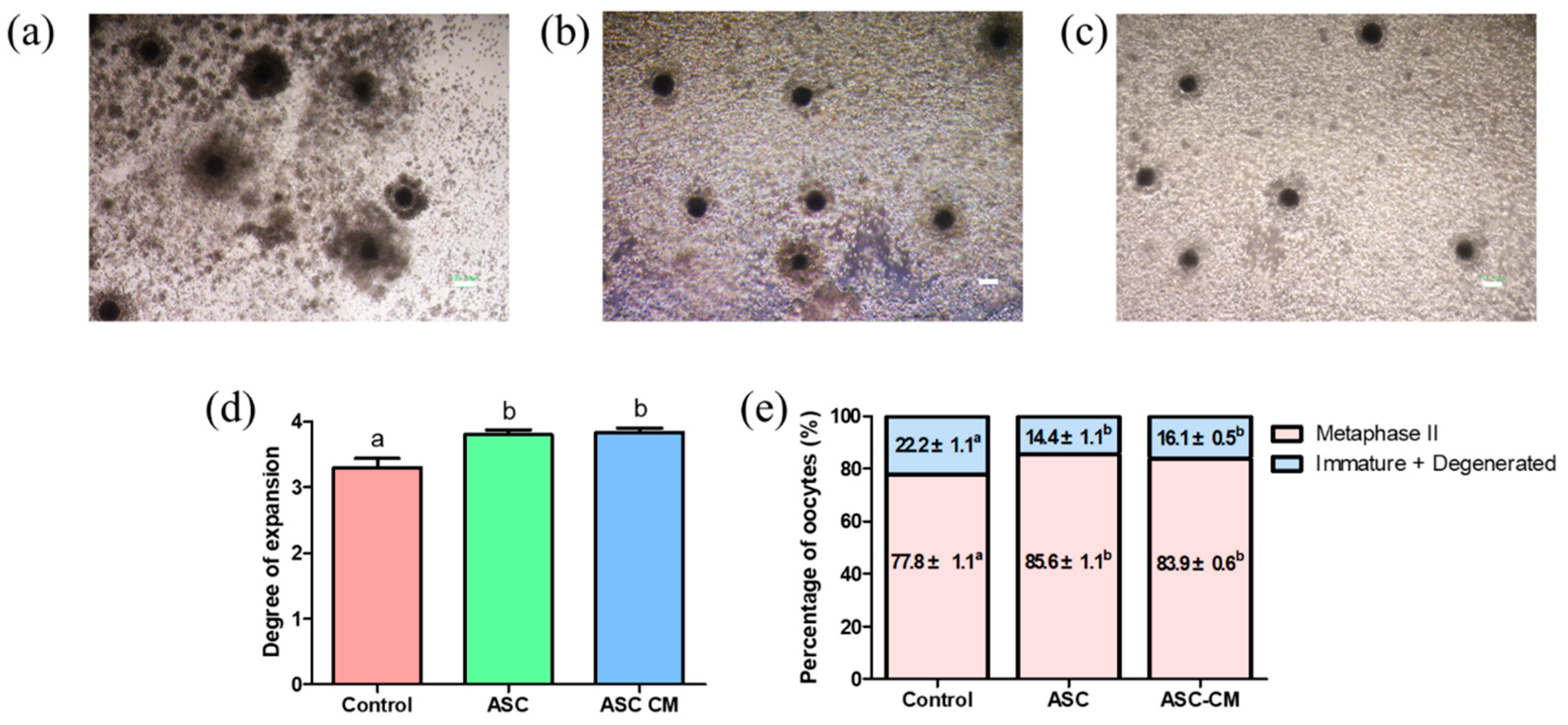
2.1. Effect of ASC and ASC-CM on Porcine Oocyte Maturation and Cumulus Expansion
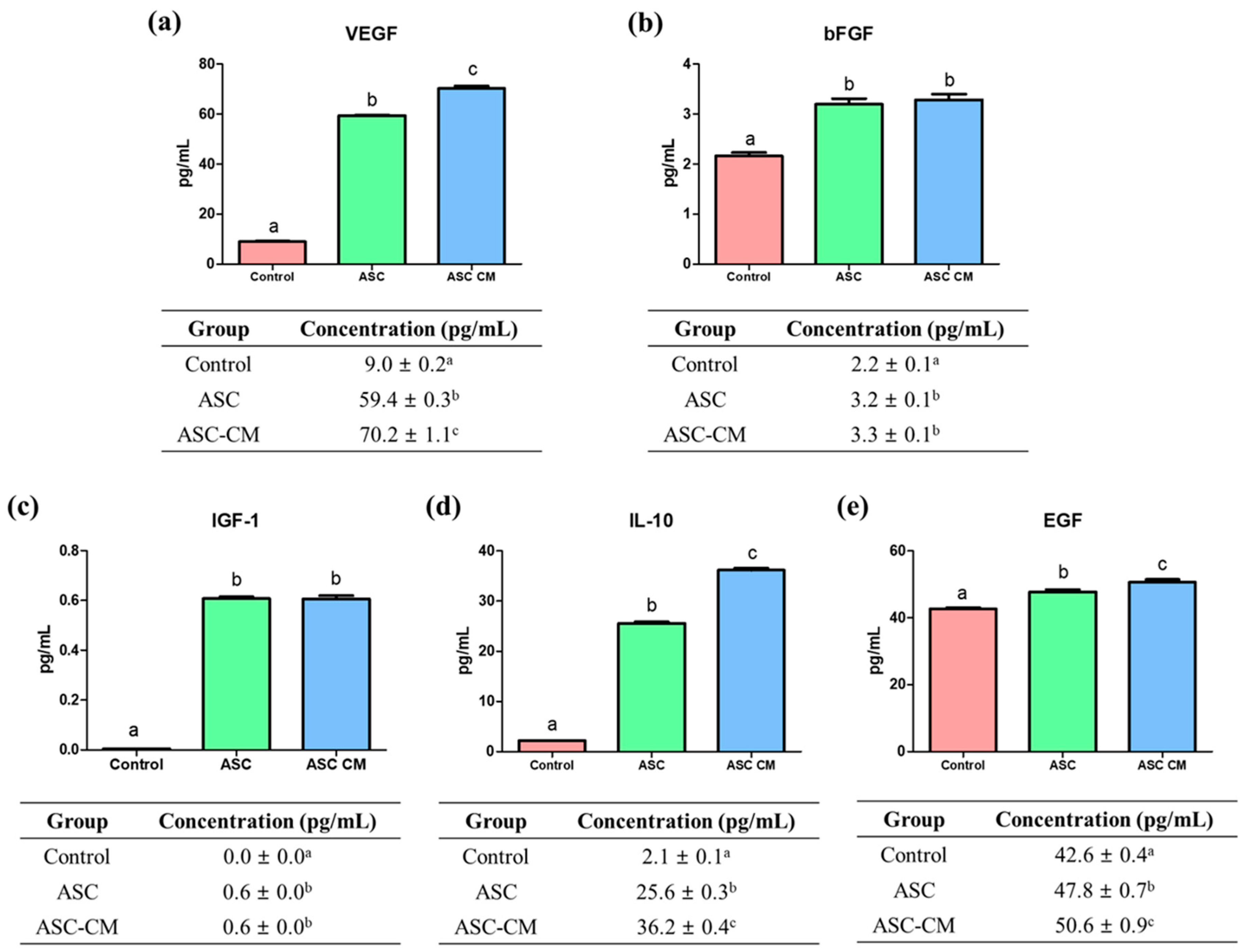
2.2. Quantification of Growth/Cytokine Factors from Culture Media
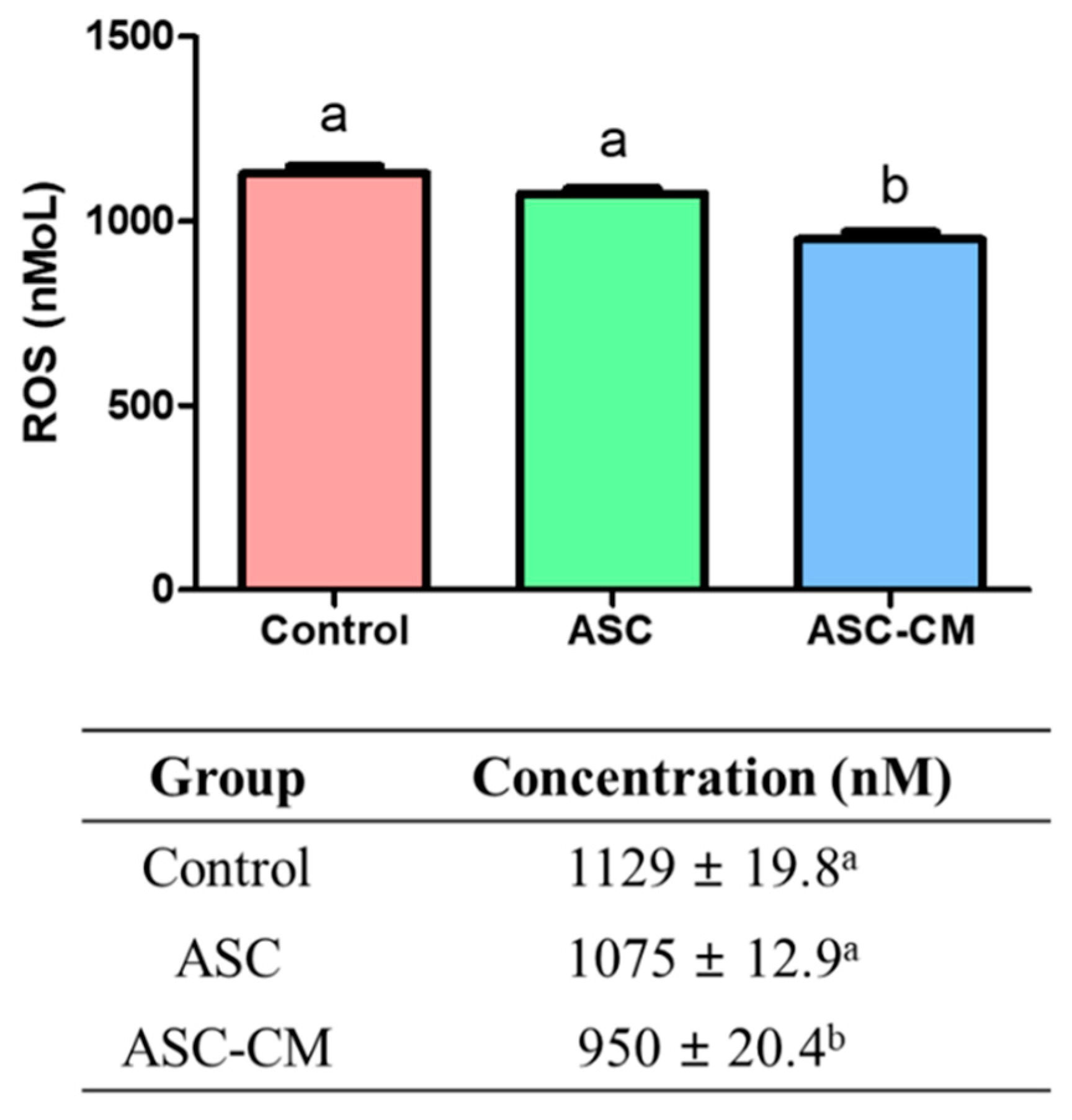
2.3. Quantification of ROS Concentration Derived from Culture Media
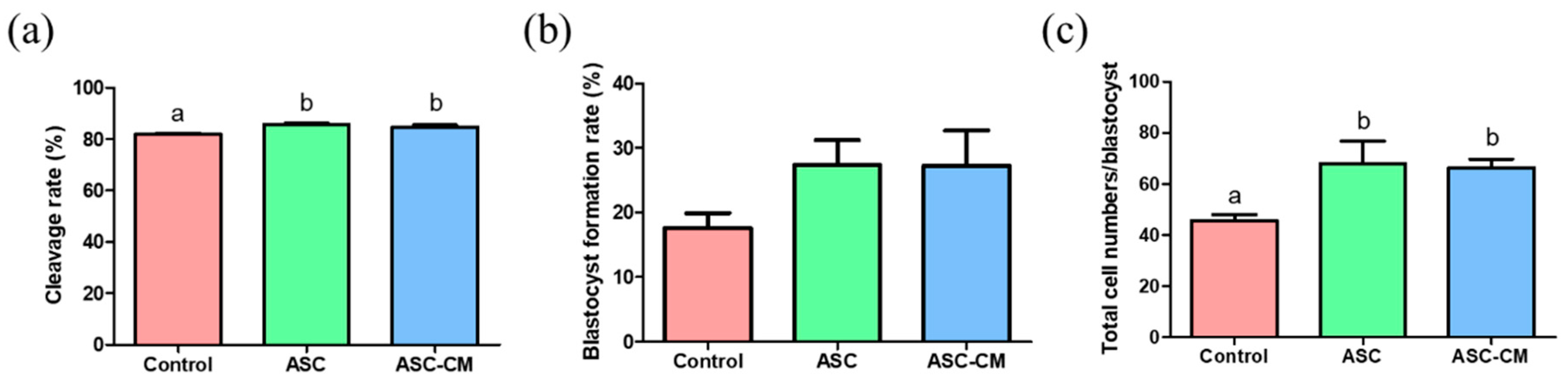
2.4. Effects of ASC and ASC-CM during IVM on In Vitro Development of Parthenotes
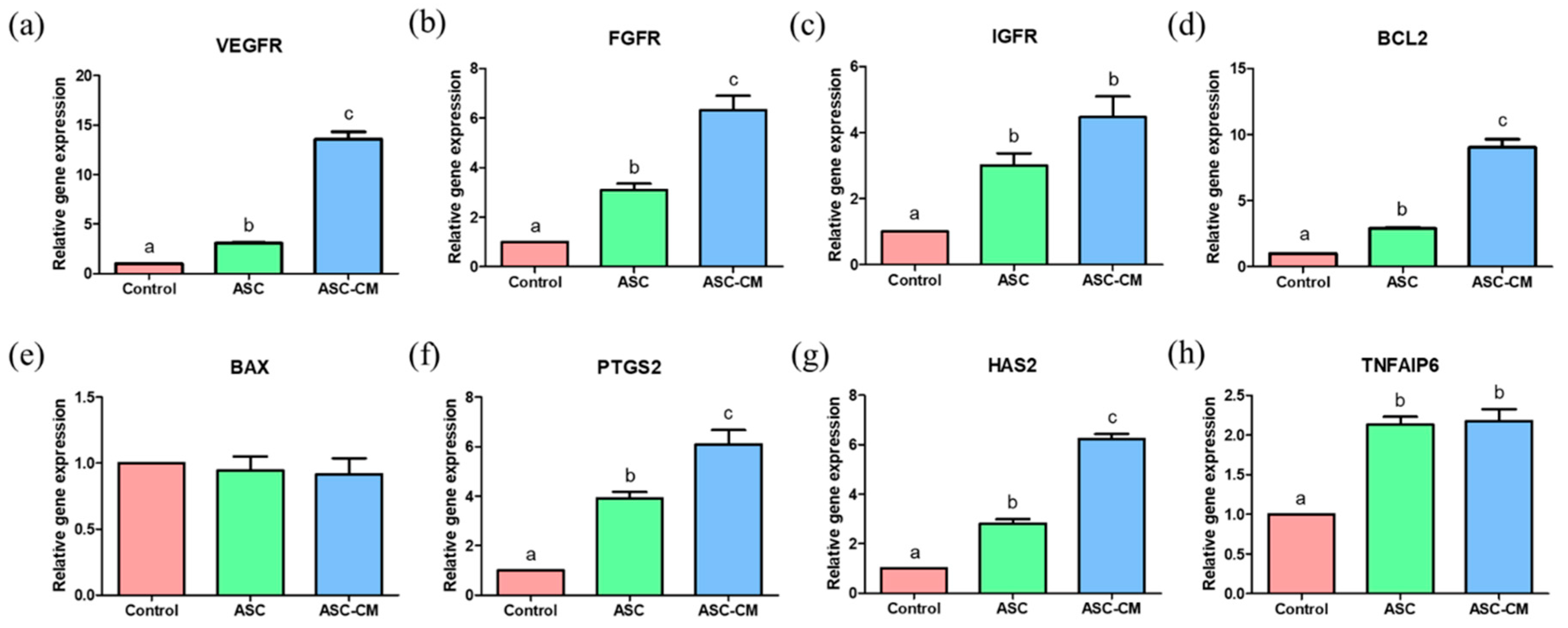
2.5. Effects of ASC and ASC-CM on the Relative Gene Expression in Cumulus Cells
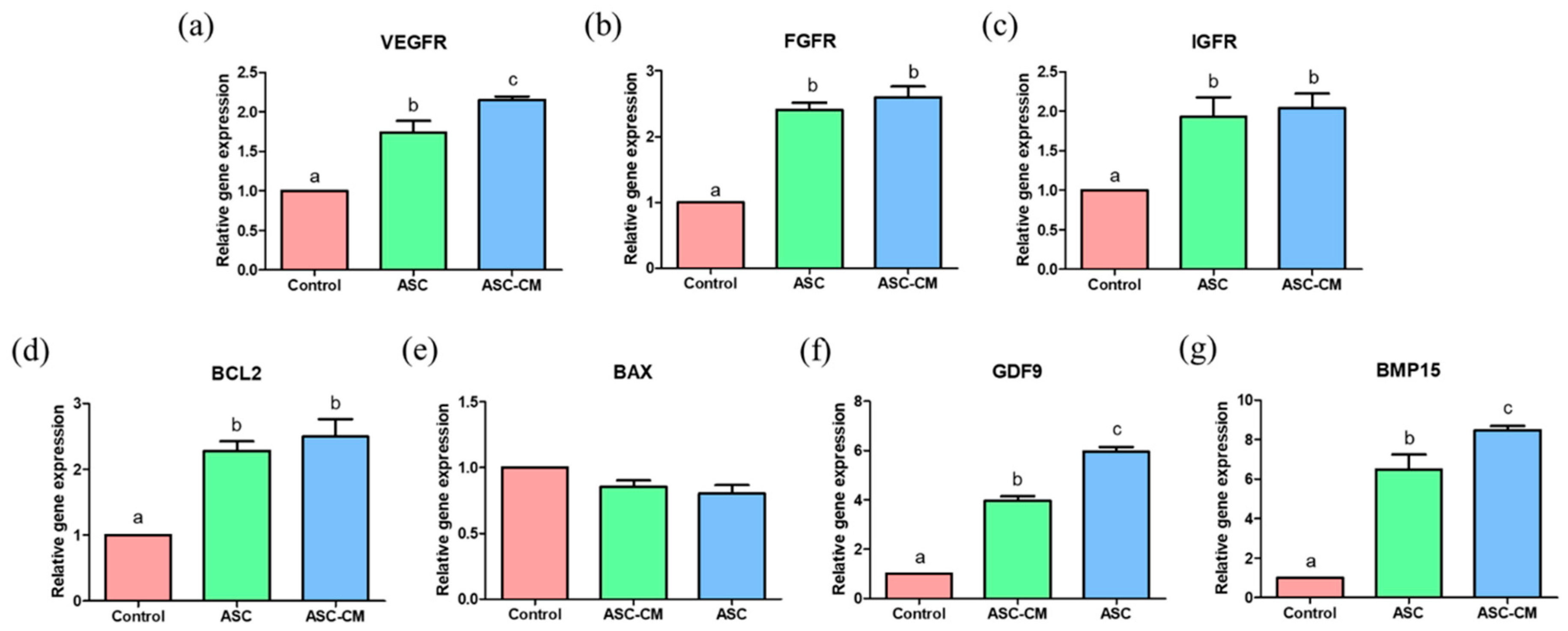
2.6. Effects of ASC and ASC-CM on the Relative Gene Expression in Oocytes
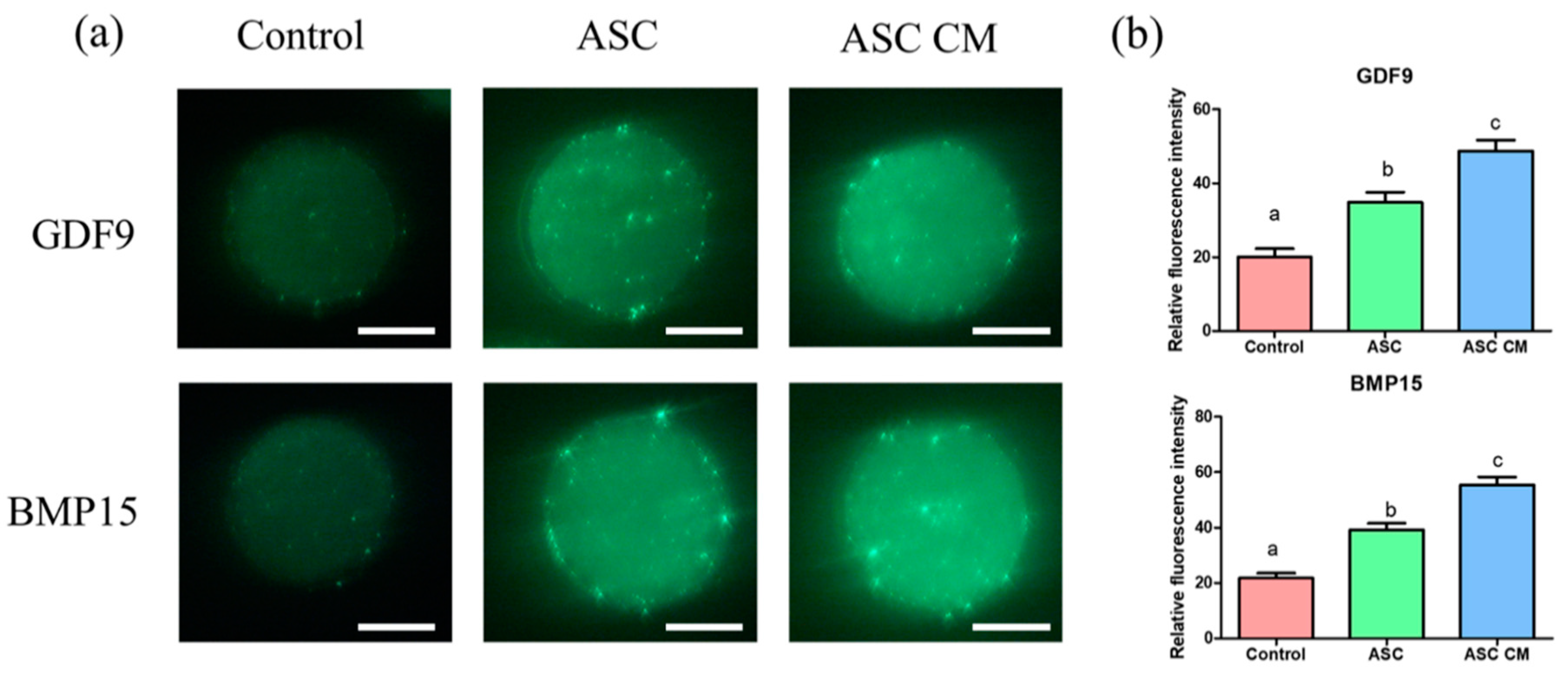
2.7. Effects of ASC and ASC-CM on the Fluorescence Intensity of GDF9 and BMP15 in Oocytes
3. Discussion
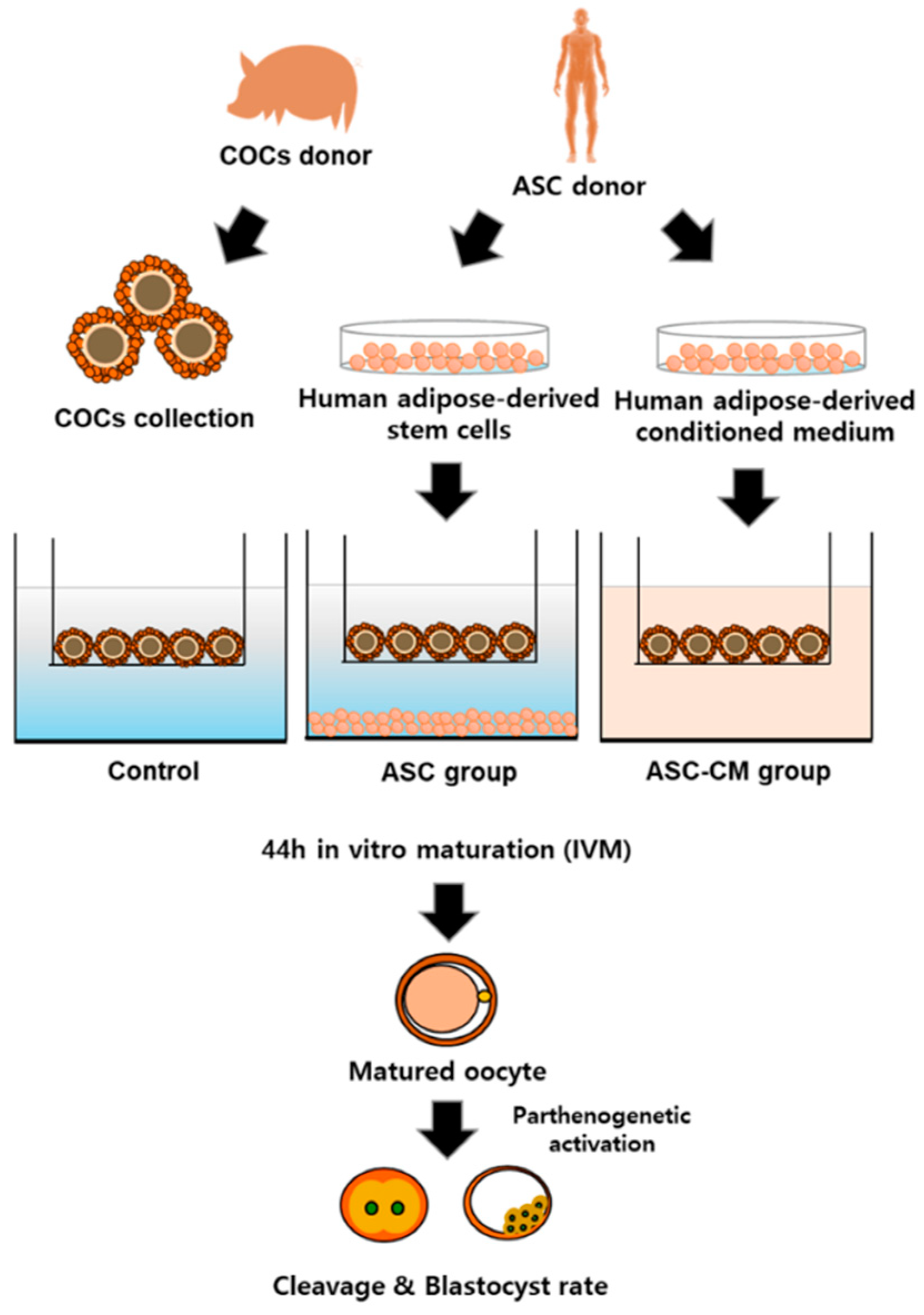
4. Materials and Methods
4.1. Chemical
4.2. Isolation and Culture of Adipose-Derived Stem Cells (ASCs)
4.3. Preparation of Human Adipose-Derived Stem Cells Conditioned Medium (ASC-CM)
4.4. In Vitro Maturation of Oocytes by Coculture with Human Adipose-Derived Stem Cells (ASC) and ASC Conditioned Medium (ASC-CM)
4.5. Evaluation of Cumulus Expansion Degree
4.6. ELISA Analysis
4.7. Assessment of In Vitro ROS Levels in Media
4.8. Parthenogenetic Activation and In Vitro Culture of Parthenotes
4.9. Total RNA Extraction and cDNA Synthesis
4.10. Real-Time PCR
4.11. Immunocytochemistry
4.12. Statistical Analysis
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ARTs | Assisted reproductive techniques |
| FBS | Fetal bovine serum |
| PBS | Phosphate buffer saline |
| TALP | Tyrode’s albumin lactate pyruvate |
| TNF | Tumor necrosis factor |
| CM bFGF | Conditioned medium Basic fibroblast growth factor |
| VEGF | Vascular endothelial growth factor |
| IGF-1 | Insulin growth factor 1 |
| IL-10 | Interleukin 10 |
| EGF | Epidermal growth factor |
| IVM | In vitro maturation |
| IVC | In vitro culture |
| ROS | Reactive oxygen species |
| FGFR | Fibroblast growth factor receptor |
| IGFR | Insulin growth factor 1 receptor |
| PTGS2 | Prostaglandin-endoperoxide synthase 2 |
| TNFAIP6 | Tumor necrosis factor α-induced protein 6 |
| HAS2 | Hyaluronan synthase 2 |
| GDF9 | Growth differentiation factor 9 |
| BMP15 | Bone morphogenetic protein 15 |
| ELISA | Enzyme-linked immunosorbent assay |
| COCs | Cumulus-oocyte complexes |
| PA | Parthenogenetic activation |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| TCM-199 | Tissue culture medium 199 |
| eCG | Equine chorionic gonadotropin |
| hCG | Human chorionic gonadotropin |
| PZM-5 | Porcine zygote medium-5 |
References
- Li, A.; Wang, F.; Li, L.; Fan, L.H.; Meng, T.G.; Li, Q.N.; Wang, Y.; Yue, W.; Wang, H.X.; Shi, Y.P.; et al. Mechanistic insights into the reduced developmental capacity of in vitro matured oocytes and importance of cumulus cells in oocyte quality determination. J. Cell Physiol. 2020, 235, 9743–9751. [Google Scholar] [CrossRef] [PubMed]
- Coticchio, G.; Dal Canto, M.; Mignini Renzini, M.; Guglielmo, M.C.; Brambillasca, F.; Turchi, D.; Novara, P.V.; Fadini, R. Oocyte maturation: Gamete-somatic cells interactions, meiotic resumption, cytoskeletal dynamics and cytoplasmic reorganization. Hum. Reprod. Update 2015, 21, 427–454. [Google Scholar] [CrossRef] [Green Version]
- Woods, D.C.; Tilly, J.L. The next (re)generation of ovarian biology and fertility in women: Is current science tomorrow’s practice? Fertil. Steril. 2012, 98, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, M.; Morselli, M.G.; Apparicio, M.; Luvoni, G.C. Granulosa cells in three-dimensional culture: A follicle-like structure for domestic cat vitrified oocytes. Reprod. Domest. Anim. 2020, 55 (Suppl. 2), 74–80. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Du, Z.; Zhou, Z.; Long, H.; Ni, Q. Effects of serum and follicular fluid on the in vitro maturation of canine oocytes. Theriogenology 2020, 143, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, S.; Gadea, J.; Coy, P.; Romar, R. Addition of exogenous proteins detected in oviductal secretions to in vitro culture medium does not improve the efficiency of in vitro fertilization in pigs. Theriogenology 2020, 157, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Jiang, Y.; Lin, T.; Kang, J.W.; Lee, J.E.; Jin, D.I. Lysophosphatidic acid improves porcine oocyte maturation and embryo development in vitro. Mol. Reprod. Dev. 2015, 82, 66–77. [Google Scholar] [CrossRef]
- Wani, N.A.; Hong, S.B. Effect of roscovitine pretreatment on in vitro maturation of oocytes and their subsequent developmental after chemical activation in dromedary camel (Camelus dromedarius). Theriogenology 2020, 157, 176–180. [Google Scholar] [CrossRef]
- Metcalf, E.S.; Masterson, K.R.; Battaglia, D.; Thompson, J.G.; Foss, R.; Beck, R.; Cook, N.L.; Thomas, O.L. Conditions to optimise the developmental competence of immature equine oocytes. Reprod. Fertil. Dev. 2020, 32, 1012–1021. [Google Scholar] [CrossRef]
- Fernandez-Hernandez, P.; Sanchez-Calabuig, M.J.; Garcia-Marin, L.J.; Bragado, M.J.; Gutierrez-Adan, A.; Millet, O.; Bruzzone, C.; Gonzalez-Fernandez, L.; Macias-Garcia, B. Study of the Metabolomics of Equine Preovulatory Follicular Fluid: A Way to Improve Current In Vitro Maturation Media. Animals 2020, 10, 883. [Google Scholar] [CrossRef]
- Yang, Y.; Kanno, C.; Sakaguchi, K.; Katagiri, S.; Yanagawa, Y.; Nagano, M. Theca cells can support bovine oocyte growth in vitro without the addition of steroid hormones. Theriogenology 2020, 142, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Kim, G.A.; Choi, Y.B.; Jo, Y.K.; Setyawan, E.M.N.; Lee, B.C. Effect of co-culture canine cumulus and oviduct cells with porcine oocytes during maturation and subsequent embryo development of parthenotes in vitro. Theriogenology 2018, 106, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, R.; Ansari, M.M.; Parmar, M.S.; Chandra, V.; Sharma, G.T. Stem Cell Conditioned Media Contains Important Growth Factors and Improves In Vitro Buffalo Embryo Production. Anim. Biotechnol. 2016, 27, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Umeki, H.; Shimura, H.; Kugumiya, K.; Shiga, K. Effect of group culture and embryo-culture conditioned medium on development of bovine embryos. J. Reprod. Dev. 2006, 52, 137–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Setyawan, E.M.N.; Choi, Y.B.; Lee, B.C. Effect of co-culture human endothelial progenitor cells with porcine oocytes during maturation and subsequent embryo development of parthenotes in vitro. Mol. Reprod. Dev. 2018, 85, 336–347. [Google Scholar] [CrossRef]
- Park, H.Y.; Kim, E.Y.; Lee, S.E.; Choi, H.Y.; Moon, J.J.; Park, M.J.; Son, Y.J.; Lee, J.B.; Jeong, C.J.; Lee, D.S.; et al. Effect of human adipose tissue-derived mesenchymal-stem-cell bioactive materials on porcine embryo development. Mol. Reprod. Dev. 2013, 80, 1035–1047. [Google Scholar] [CrossRef]
- Kim, E.Y.; Lee, J.B.; Park, H.Y.; Jeong, C.J.; Riu, K.Z.; Park, S.P. The use of embryonic stem cell derived bioactive material as a new protein supplement for the in vitro culture of bovine embryos. J. Reprod. Dev. 2011, 57, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Maumus, M.; Rozier, P.; Boulestreau, J.; Jorgensen, C.; Noel, D. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Opportunities and Challenges for Clinical Translation. Front. Bioeng. Biotechnol. 2020, 8, 997. [Google Scholar] [CrossRef]
- Diomede, F.; Gugliandolo, A.; Scionti, D.; Merciaro, I.; Cavalcanti, M.F.; Mazzon, E.; Trubiani, O. Biotherapeutic Effect of Gingival Stem Cells Conditioned Medium in Bone Tissue Restoration. Int. J. Mol. Sci. 2018, 19, 329. [Google Scholar] [CrossRef] [Green Version]
- Salgado, A.J.; Reis, R.L.; Sousa, N.J.; Gimble, J.M. Adipose tissue derived stem cells secretome: Soluble factors and their roles in regenerative medicine. Curr. Stem. Cell Res. Ther. 2010, 5, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Bakhshi, T.; Zabriskie, R.C.; Bodie, S.; Kidd, S.; Ramin, S.; Paganessi, L.A.; Gregory, S.A.; Fung, H.C.; Christopherson, K.W., 2nd. Mesenchymal stem cells from the Wharton’s jelly of umbilical cord segments provide stromal support for the maintenance of cord blood hematopoietic stem cells during long-term ex vivo culture. Transfusion 2008, 48, 2638–2644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, R.; Betancur, M.; Boissel, L.; Tuncer, H.; Cetrulo, C.; Klingemann, H. Umbilical cord mesenchymal stem cells: Adjuvants for human cell transplantation. Biol. Blood Marrow Transplant. 2007, 13, 1477–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamari, M.; Nishino, Y.; Yamamoto, N.; Ueda, M. Acceleration of wound healing with stem cell-derived growth factors. Int. J. Oral. Maxillofac. Implants 2013, 28, e369–e375. [Google Scholar] [CrossRef] [Green Version]
- Jayaraman, P.; Nathan, P.; Vasanthan, P.; Musa, S.; Govindasamy, V. Stem cells conditioned medium: A new approach to skin wound healing management. Cell Biol. Int. 2013, 37, 1122–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocking, A.M.; Gibran, N.S. Mesenchymal stem cells: Paracrine signaling and differentiation during cutaneous wound repair. Exp. Cell Res. 2010, 316, 2213–2219. [Google Scholar] [CrossRef] [Green Version]
- Jafarzadeh, H.; Nazarian, H.; Ghaffari Novin, M.; Shams Mofarahe, Z.; Eini, F.; Piryaei, A. Improvement of oocyte in vitro maturation from mice with polycystic ovary syndrome by human mesenchymal stromal cell-conditioned media. J. Cell Biochem. 2018, 119, 10365–10375. [Google Scholar] [CrossRef]
- Akbari, H.; Eftekhar Vaghefi, S.H.; Shahedi, A.; Habibzadeh, V.; Mirshekari, T.R.; Ganjizadegan, A.; Mollaei, H.; Ahmadi, M.; Nematollahi-Mahani, S.N. Mesenchymal Stem Cell-Conditioned Medium Modulates Apoptotic and Stress-Related Gene Expression, Ameliorates Maturation and Allows for the Development of Immature Human Oocytes after Artificial Activation. Genes 2017, 8, 371. [Google Scholar] [CrossRef] [Green Version]
- Ra, J.C.; Shin, I.S.; Kim, S.H.; Kang, S.K.; Kang, B.C.; Lee, H.Y.; Kim, Y.J.; Jo, J.Y.; Yoon, E.J.; Choi, H.J.; et al. Safety of intravenous infusion of human adipose tissue-derived mesenchymal stem cells in animals and humans. Stem. Cells Dev. 2011, 20, 1297–1308. [Google Scholar] [CrossRef]
- Borgese, M.; Barone, L.; Rossi, F.; Raspanti, M.; Papait, R.; Valdatta, L.; Bernardini, G.; Gornati, R. Effect of Nanostructured Scaffold on Human Adipose-Derived Stem Cells: Outcome of In Vitro Experiments. Nanomaterials 2020, 10, 1822. [Google Scholar] [CrossRef]
- Baek, S.J.; Kang, S.K.; Ra, J.C. In vitro migration capacity of human adipose tissue-derived mesenchymal stem cells reflects their expression of receptors for chemokines and growth factors. Exp. Mol. Med. 2011, 43, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Capella-Monsonis, H.; De Pieri, A.; Peixoto, R.; Korntner, S.; Zeugolis, D.I. Extracellular matrix-based biomaterials as adipose-derived stem cell delivery vehicles in wound healing: A comparative study between a collagen scaffold and two xenografts. Stem. Cell Res. Ther. 2020, 11, 510. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.G.; He, Z.Y.; Liang, S.; Yang, Q.; Cheng, P.; Chen, A.M. Comprehensive proteomic analysis of exosomes derived from human bone marrow, adipose tissue, and umbilical cord mesenchymal stem cells. Stem. Cell Res. Ther. 2020, 11, 511. [Google Scholar] [CrossRef] [PubMed]
- Maqsood, M.; Kang, M.; Wu, X.; Chen, J.; Teng, L.; Qiu, L. Adult mesenchymal stem cells and their exosomes: Sources, characteristics, and application in regenerative medicine. Life Sci. 2020, 256, 118002. [Google Scholar] [CrossRef] [PubMed]
- Raty, M.; Ketoja, E.; Pitkanen, T.; Ahola, V.; Kananen, K.; Peippo, J. In vitro maturation supplements affect developmental competence of bovine cumulus-oocyte complexes and embryo quality after vitrification. Cryobiology 2011, 63, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Lapa, M.; Marques, C.C.; Alves, S.P.; Vasques, M.I.; Baptista, M.C.; Carvalhais, I.; Silva Pereira, M.; Horta, A.E.; Bessa, R.J.; Pereira, R.M. Effect of trans-10 cis-12 conjugated linoleic acid on bovine oocyte competence and fatty acid composition. Reprod. Domest. Anim. 2011, 46, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Javvaji, P.K.; Dhali, A.; Francis, J.R.; Kolte, A.P.; Roy, S.C.; Selvaraju, S.; Mech, A.; Sejian, V. IGF-1 treatment during in vitro maturation improves developmental potential of ovine oocytes through the regulation of PI3K/Akt and apoptosis signaling. Anim. Biotechnol. 2020, 1–8. [Google Scholar] [CrossRef]
- Son, Y.J.; Lee, S.E.; Hyun, H.; Shin, M.Y.; Park, Y.G.; Jeong, S.G.; Kim, E.Y.; Park, S.P. Fibroblast growth factor 10 markedly improves in vitro maturation of porcine cumulus-oocyte complexes. Mol. Reprod. Dev. 2017, 84, 67–75. [Google Scholar] [CrossRef]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Lee, B.C. Canine oviductal exosomes improve oocyte development via EGFR/MAPK signaling pathway. Reproduction 2020, 160, 613–625. [Google Scholar] [CrossRef]
- Jia, B.Y.; Xiang, D.C.; Zhang, B.; Quan, G.B.; Shao, Q.Y.; Hong, Q.H.; Wu, G.Q. Quality of vitrified porcine immature oocytes is improved by coculture with fresh oocytes during in vitro maturation. Mol. Reprod. Dev. 2019, 86, 1615–1627. [Google Scholar] [CrossRef]
- Appeltant, R.; Somfai, T.; Kikuchi, K.; Maes, D.; Van Soom, A. Influence of co-culture with denuded oocytes during in vitro maturation on fertilization and developmental competence of cumulus-enclosed porcine oocytes in a defined system. Anim. Sci. J. 2016, 87, 503–510. [Google Scholar] [CrossRef]
- Dos Santos-Neto, P.C.; Vilarino, M.; Cuadro, F.; Barrera, N.; Crispo, M.; Menchaca, A. Cumulus cells during in vitro fertilization and oocyte vitrification in sheep: Remove, maintain or add? Cryobiology 2020, 92, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Setyawan, E.M.N.; Lee, B.C. Interaction of the EGFR signaling pathway with porcine cumulus oocyte complexes and oviduct cells in a coculture system. J. Cell Physiol. 2019, 234, 4030–4043. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Oh, H.J.; Kim, M.J.; Kim, G.A.; Choi, Y.B.; Jo, Y.K.; Setyawan, E.M.N.; Lee, B.C. Oocyte maturation-related gene expression in the canine oviduct, cumulus cells, and oocytes and effect of co-culture with oviduct cells on in vitro maturation of oocytes. J. Assist. Reprod. Genet. 2017, 34, 929–938. [Google Scholar] [CrossRef] [PubMed]
- LaFramboise, W.A.; Petrosko, P.; Krill-Burger, J.M.; Morris, D.R.; McCoy, A.R.; Scalise, D.; Malehorn, D.E.; Guthrie, R.D.; Becich, M.J.; Dhir, R. Proteins secreted by embryonic stem cells activate cardiomyocytes through ligand binding pathways. J. Proteom. 2010, 73, 992–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Graham-Evans, B.; Broxmeyer, H.E. Murine embryonic stem cells secrete cytokines/growth modulators that enhance cell survival/anti-apoptosis and stimulate colony formation of murine hematopoietic progenitor cells. Stem. Cells 2006, 24, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Park, B.S.; Sung, J.H. The wound-healing and antioxidant effects of adipose-derived stem cells. Expert Opin. Biol. Ther. 2009, 9, 879–887. [Google Scholar] [CrossRef]
- Richter, K.S. The importance of growth factors for preimplantation embryo development and in-vitro culture. Curr. Opin. Obstet. Gynecol. 2008, 20, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Fraser, H.M.; Lunn, S.F. Angiogenesis and its control in the female reproductive system. Br. Med. Bull. 2000, 56, 787–797. [Google Scholar] [CrossRef] [Green Version]
- Anasti, J.N.; Kalantaridou, S.N.; Kimzey, L.M.; George, M.; Nelson, L.M. Human follicle fluid vascular endothelial growth factor concentrations are correlated with luteinization in spontaneously developing follicles. Hum. Reprod. 1998, 13, 1144–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shweiki, D.; Itin, A.; Neufeld, G.; Gitay-Goren, H.; Keshet, E. Patterns of expression of vascular endothelial growth factor (VEGF) and VEGF receptors in mice suggest a role in hormonally regulated angiogenesis. J. Clin. Investig. 1993, 91, 2235–2243. [Google Scholar] [CrossRef] [Green Version]
- Ravindranath, N.; Little-Ihrig, L.; Phillips, H.S.; Ferrara, N.; Zeleznik, A.J. Vascular endothelial growth factor messenger ribonucleic acid expression in the primate ovary. Endocrinology 1992, 131, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Christenson, L.K.; Stouffer, R.L. Follicle-stimulating hormone and luteinizing hormone/chorionic gonadotropin stimulation of vascular endothelial growth factor production by macaque granulosa cells from pre- and periovulatory follicles. J. Clin. Endocrinol. Metab. 1997, 82, 2135–2142. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N. The Fgf families in humans, mice, and zebrafish: Their evolutional processes and roles in development, metabolism, and disease. Biol. Pharm. Bull. 2007, 30, 1819–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ornitz, D.M.; Itoh, N. Fibroblast growth factors. Genome Biol. 2001, 2, REVIEWS3005. [Google Scholar] [CrossRef] [Green Version]
- Buratini, J., Jr.; Pinto, M.G.; Castilho, A.C.; Amorim, R.L.; Giometti, I.C.; Portela, V.M.; Nicola, E.S.; Price, C.A. Expression and function of fibroblast growth factor 10 and its receptor, fibroblast growth factor receptor 2B, in bovine follicles. Biol. Reprod. 2007, 77, 743–750. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, K.; Su, Y.Q.; Diaz, F.J.; Pangas, S.A.; Sharma, S.; Wigglesworth, K.; O’Brien, M.J.; Matzuk, M.M.; Shimasaki, S.; Eppig, J.J. Oocyte-derived BMP15 and FGFs cooperate to promote glycolysis in cumulus cells. Development 2007, 134, 2593–2603. [Google Scholar] [CrossRef] [Green Version]
- Ben-Haroush, A.; Abir, R.; Ao, A.; Jin, S.; Kessler-Icekson, G.; Feldberg, D.; Fisch, B. Expression of basic fibroblast growth factor and its receptors in human ovarian follicles from adults and fetuses. Fertil. Steril. 2005, 84 (Suppl. 2), 1257–1268. [Google Scholar] [CrossRef]
- Berisha, B.; Sinowatz, F.; Schams, D. Expression and localization of fibroblast growth factor (FGF) family members during the final growth of bovine ovarian follicles. Mol. Reprod. Dev. 2004, 67, 162–171. [Google Scholar] [CrossRef]
- Cardilli, D.J.; Sousa-Oliveira, K.; Franchi-Joao, C.; Azevedo-Voorwald, F.; Machado-Silva, M.A.; Oliveira, J.A.; Sanchez-Calabuig, M.J.; Toniollo, G.H.; Perez-Gutierrez, J.F. Immunolocalization and effect of low concentrations of Insulin like growth factor-1 (IGF-1) in the canine ovary. Vet. Med. Sci. 2020. [Google Scholar] [CrossRef]
- Kwintkiewicz, J.; Giudice, L.C. The interplay of insulin-like growth factors, gonadotropins, and endocrine disruptors in ovarian follicular development and function. Semin. Reprod. Med. 2009, 27, 43–51. [Google Scholar] [CrossRef]
- Oberlender, G.; Murgas, L.D.; Zangeronimo, M.G.; da Silva, A.C.; Menezes Tde, A.; Pontelo, T.P.; Vieira, L.A. Role of insulin-like growth factor-I and follicular fluid from ovarian follicles with different diameters on porcine oocyte maturation and fertilization in vitro. Theriogenology 2013, 80, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.V.; Dukesova, J.; Makarevich, A.V.; Kubek, A.; Bulla, J. Evidence that growth factors IGF-I, IGF-II and EGF can stimulate nuclear maturation of porcine oocytes via intracellular protein kinase A. Reprod. Nutr. Dev. 2000, 40, 559–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbs, S.A.; Webber, L.J.; Stark, J.; Rice, S.; Margara, R.; Lavery, S.; Trew, G.H.; Hardy, K.; Franks, S. Role of Insulin-like growth factors in initiation of follicle growth in normal and polycystic human ovaries. J. Clin. Endocrinol. Metab. 2013, 98, 3298–3305. [Google Scholar] [CrossRef] [PubMed]
- Baumgarten, S.C.; Convissar, S.M.; Fierro, M.A.; Winston, N.J.; Scoccia, B.; Stocco, C. IGF1R signaling is necessary for FSH-induced activation of AKT and differentiation of human Cumulus granulosa cells. J. Clin. Endocrinol. Metab. 2014, 99, 2995–3004. [Google Scholar] [CrossRef] [Green Version]
- Jatesada, J.; Elisabeth, P.; Anne-Marie, D. Seminal plasma did not influence the presence of transforming growth factor-beta1, interleukine-10 and interleukin-6 in porcine follicles shortly after insemination. Acta Vet. Scand. 2013, 55, 66. [Google Scholar] [CrossRef] [Green Version]
- Kollmann, Z.; Schneider, S.; Fux, M.; Bersinger, N.A.; von Wolff, M. Gonadotrophin stimulation in IVF alters the immune cell profile in follicular fluid and the cytokine concentrations in follicular fluid and serum. Hum. Reprod. 2017, 32, 820–831. [Google Scholar] [CrossRef]
- Pereira, L.M.C.; Bersano, P.R.O.; Rocha, D.D.; Lopes, M.D. Effect of EGF on expression and localization of maturation-promoting factor, mitogen-activated protein kinase, p34(cdc2) and cyclin B during different culture periods on in vitro maturation of canine oocytes. Reprod. Domest. Anim. 2019, 54, 325–341. [Google Scholar] [CrossRef]
- Bezerra, F.T.G.; Paulino, L.; Silva, B.R.; Silva, A.W.B.; Souza Batista, A.L.P.; Silva, J.R.V. Effects of epidermal growth factor and progesterone on oocyte meiotic resumption and the expression of maturation-related transcripts during prematuration of oocytes from small and medium-sized bovine antral follicles. Reprod. Fertil. Dev. 2020. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Falcone, T.; Mohamed, M.S.; Aleem, A.A.; Sharma, R.K.; Worley, S.E.; Thornton, J.; Agarwal, A. Differential growth of human embryos in vitro: Role of reactive oxygen species. Fertil. Steril. 2004, 82, 593–600. [Google Scholar] [CrossRef]
- Guerin, P.; El Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Goto, Y.; Noda, Y.; Mori, T.; Nakano, M. Increased generation of reactive oxygen species in embryos cultured in vitro. Free Radic. Biol. Med. 1993, 15, 69–75. [Google Scholar] [CrossRef]
- Nasr-Esfahani, M.H.; Aitken, J.R.; Johnson, M.H. Hydrogen peroxide levels in mouse oocytes and early cleavage stage embryos developed in vitro or in vivo. Development 1990, 109, 501–507. [Google Scholar] [PubMed]
- de Lamirande, E.; Jiang, H.; Zini, A.; Kodama, H.; Gagnon, C. Reactive oxygen species and sperm physiology. Rev. Reprod. 1997, 2, 48–54. [Google Scholar] [CrossRef]
- Iwata, H.; Ohota, M.; Hashimoto, S.; Nagai, Y. Free oxygen radicals are generated at the time of aspiration of oocytes from ovaries that have been stored for a long time. Zygote 2003, 11, 1–5. [Google Scholar] [CrossRef]
- Li, H.K.; Kuo, T.Y.; Yang, H.S.; Chen, L.R.; Li, S.S.; Huang, H.W. Differential gene expression of bone morphogenetic protein 15 and growth differentiation factor 9 during in vitro maturation of porcine oocytes and early embryos. Anim. Reprod. Sci. 2008, 103, 312–322. [Google Scholar] [CrossRef] [PubMed]
- Hussein, T.S.; Thompson, J.G.; Gilchrist, R.B. Oocyte-secreted factors enhance oocyte developmental competence. Dev. Biol. 2006, 296, 514–521. [Google Scholar] [CrossRef]
- Yan, C.; Wang, P.; DeMayo, J.; DeMayo, F.J.; Elvin, J.A.; Carino, C.; Prasad, S.V.; Skinner, S.S.; Dunbar, B.S.; Dube, J.L.; et al. Synergistic roles of bone morphogenetic protein 15 and growth differentiation factor 9 in ovarian function. Mol. Endocrinol. 2001, 15, 854–866. [Google Scholar] [CrossRef]
- Su, Y.Q.; Wu, X.; O’Brien, M.J.; Pendola, F.L.; Denegre, J.N.; Matzuk, M.M.; Eppig, J.J. Synergistic roles of BMP15 and GDF9 in the development and function of the oocyte-cumulus cell complex in mice: Genetic evidence for an oocyte-granulosa cell regulatory loop. Dev. Biol. 2004, 276, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Dragovic, R.A.; Ritter, L.J.; Schulz, S.J.; Amato, F.; Thompson, J.G.; Armstrong, D.T.; Gilchrist, R.B. Oocyte-secreted factor activation of SMAD 2/3 signaling enables initiation of mouse cumulus cell expansion. Biol. Reprod. 2007, 76, 848–857. [Google Scholar] [CrossRef]
- Prochazka, R.; Nemcova, L.; Nagyova, E.; Kanka, J. Expression of growth differentiation factor 9 messenger RNA in porcine growing and preovulatory ovarian follicles. Biol. Reprod. 2004, 71, 1290–1295. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′ → 3′) | GenBank No. | Product Size (bp) |
|---|---|---|---|
| GAPDH | F-CTTCCACTTTTGATGCTGGGG R-TCCAGGGGCTCTTACTCCTT | NM_001206359.1 | 145 |
| VEGFR | F-AAGCCATTCCGATAACAACG R-GCTGTTTTCGATGCTTCACA | NM_021065525.1 | 173 |
| FGFR | F: TCATCTGCCTGGTTGTGGTC R: CGCAGCCACGTAAACTTCTG | NM_001099924.2 | 140 |
| IGFR | F: CCCAATGGCAACCTGAGCTA R: TCCTCGACATCAATGGTGCC | NM_214172.1 | 137 |
| BCL2 | F-AGGGCATTCAGTGACCTGAC R-CGATCCGACTCACCAATACC | NM_214285 | 193 |
| BAX | F-TGCCTCAGGATGCATCTACC R-AAGTAGAAAAGCGCGACCAC | XM_003127290 | 199 |
| GDF9 | F-ACATGACTCTTCTGGCAGCC R-ACCCTCAGACAGCCCTCTTT | NM_001001909.1 | 140 |
| BMP15 | F-AGCTCTGGAATCACAAGGGG R-ACAAGAAGGCAGTGTCCAGG | NM_001005155.1 | 123 |
| PTGS2 | F-TGGGGAGACCATGGTAGAAG R-CTGAATCGAGGCAGTGTTGA | NM_214321.1 | 142 |
| HAS2 | F-AGTTTATGGGCAGCCAATGTAGTT R-GCACTTGGACCGAGCTGTGT | AB050389 | 101 |
| TNFAIP6 | F-AGAAGCGAAAGATGGGATGCT R-CATTTGGGAAGCCTGGAGATT | NM_001159607 | 106 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.H. Human Adipose-Derived Stem Cells’ Paracrine Factors in Conditioned Medium Can Enhance Porcine Oocyte Maturation and Subsequent Embryo Development. Int. J. Mol. Sci. 2021, 22, 579. https://doi.org/10.3390/ijms22020579
Lee SH. Human Adipose-Derived Stem Cells’ Paracrine Factors in Conditioned Medium Can Enhance Porcine Oocyte Maturation and Subsequent Embryo Development. International Journal of Molecular Sciences. 2021; 22(2):579. https://doi.org/10.3390/ijms22020579
Chicago/Turabian StyleLee, Seok Hee. 2021. "Human Adipose-Derived Stem Cells’ Paracrine Factors in Conditioned Medium Can Enhance Porcine Oocyte Maturation and Subsequent Embryo Development" International Journal of Molecular Sciences 22, no. 2: 579. https://doi.org/10.3390/ijms22020579