Salinity Effects on Guard Cell Proteome in Chenopodium quinoa

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
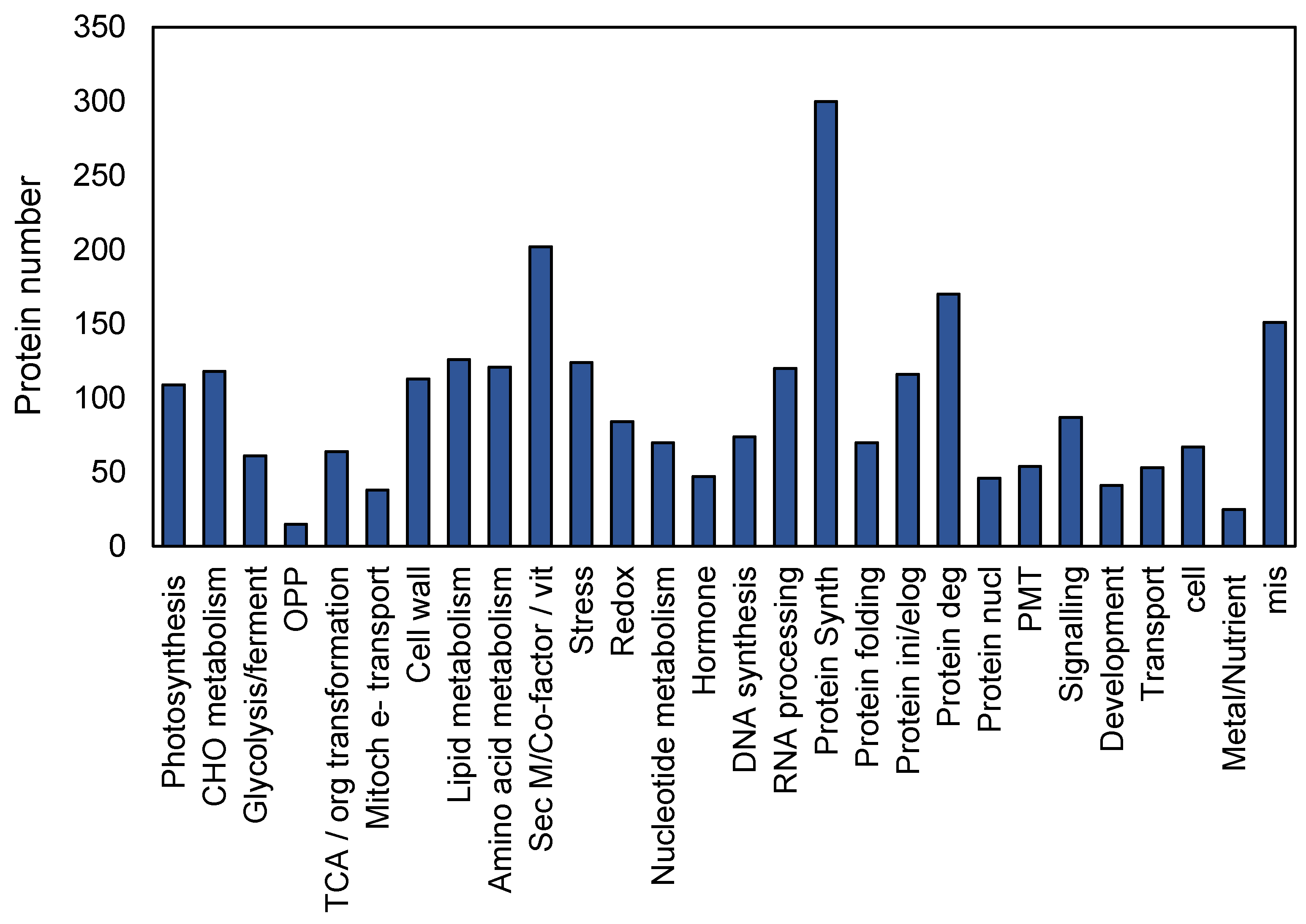
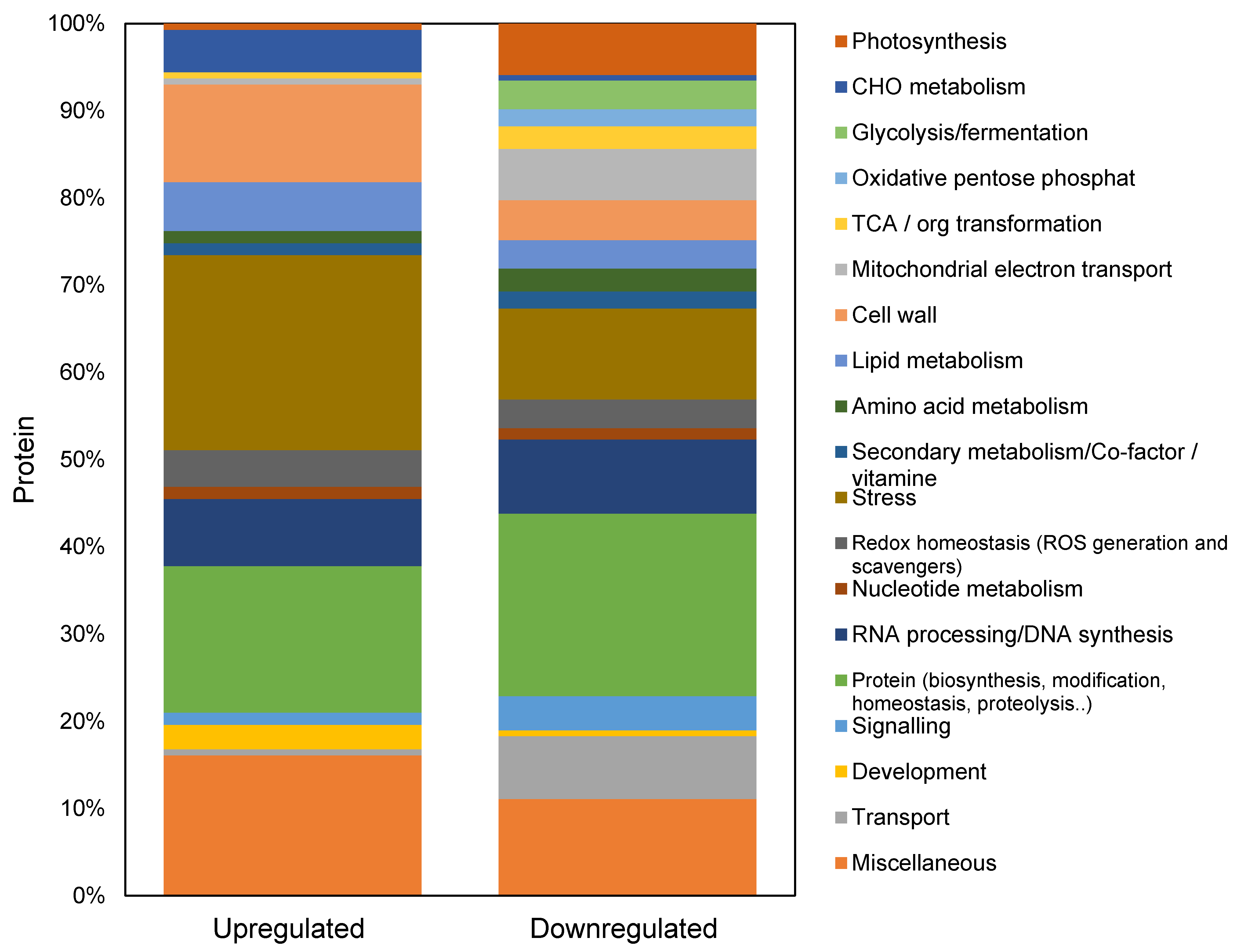
2.1. Protein Classification in the GC
2.2. Signaling Proteins in Quinoa GC Proteome
2.3. Transporters Proteins in Quinoa GC Proteome
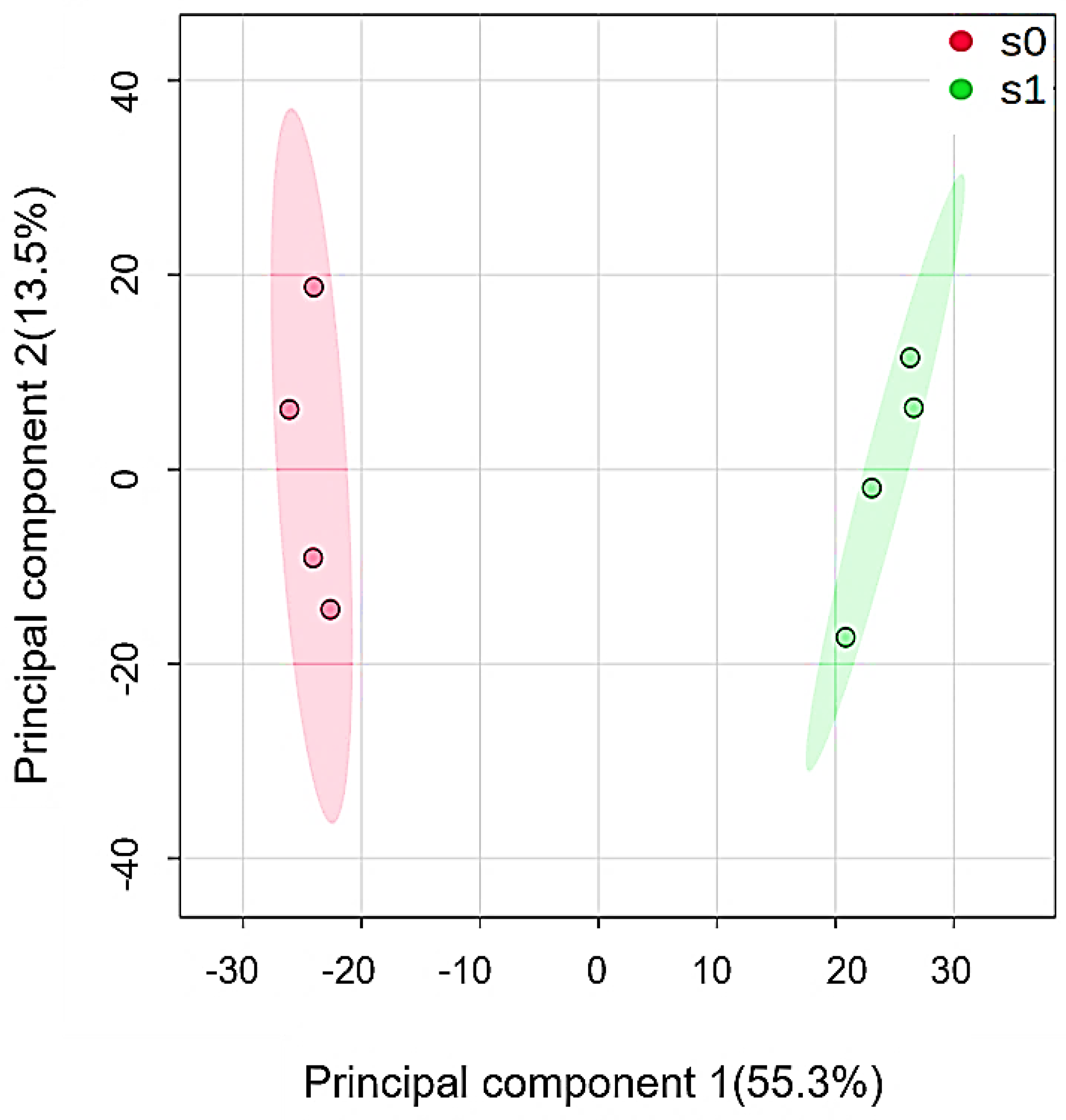
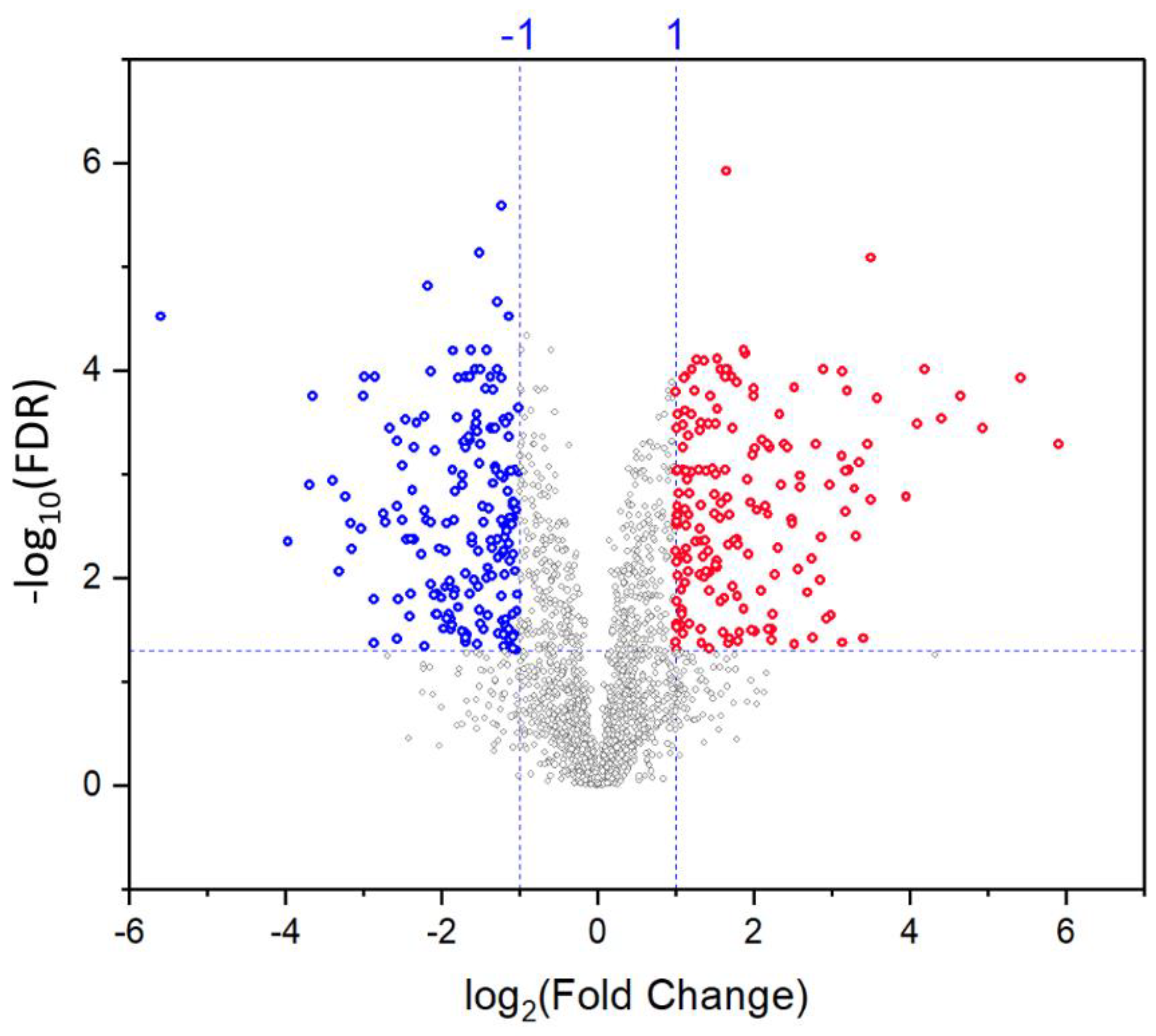
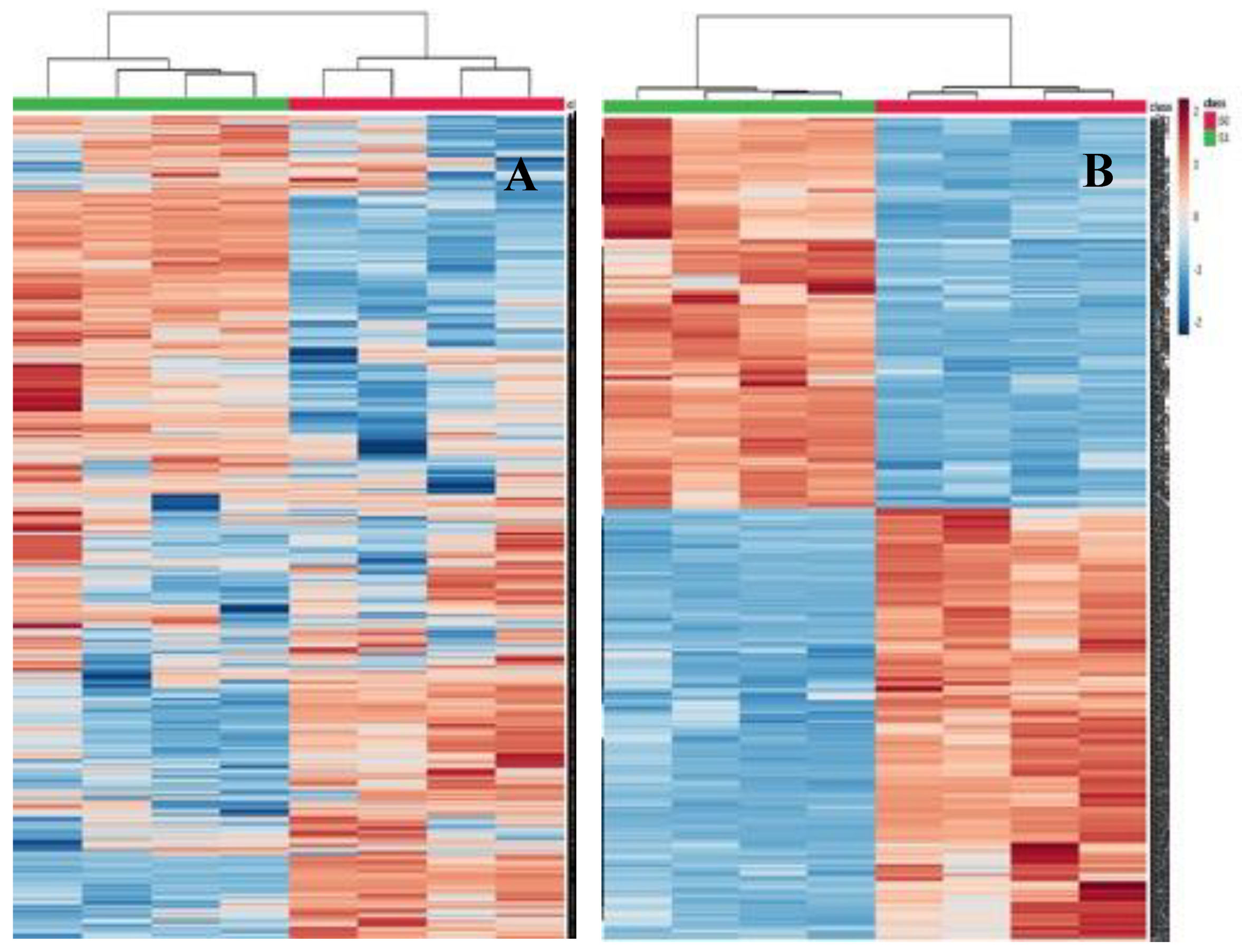
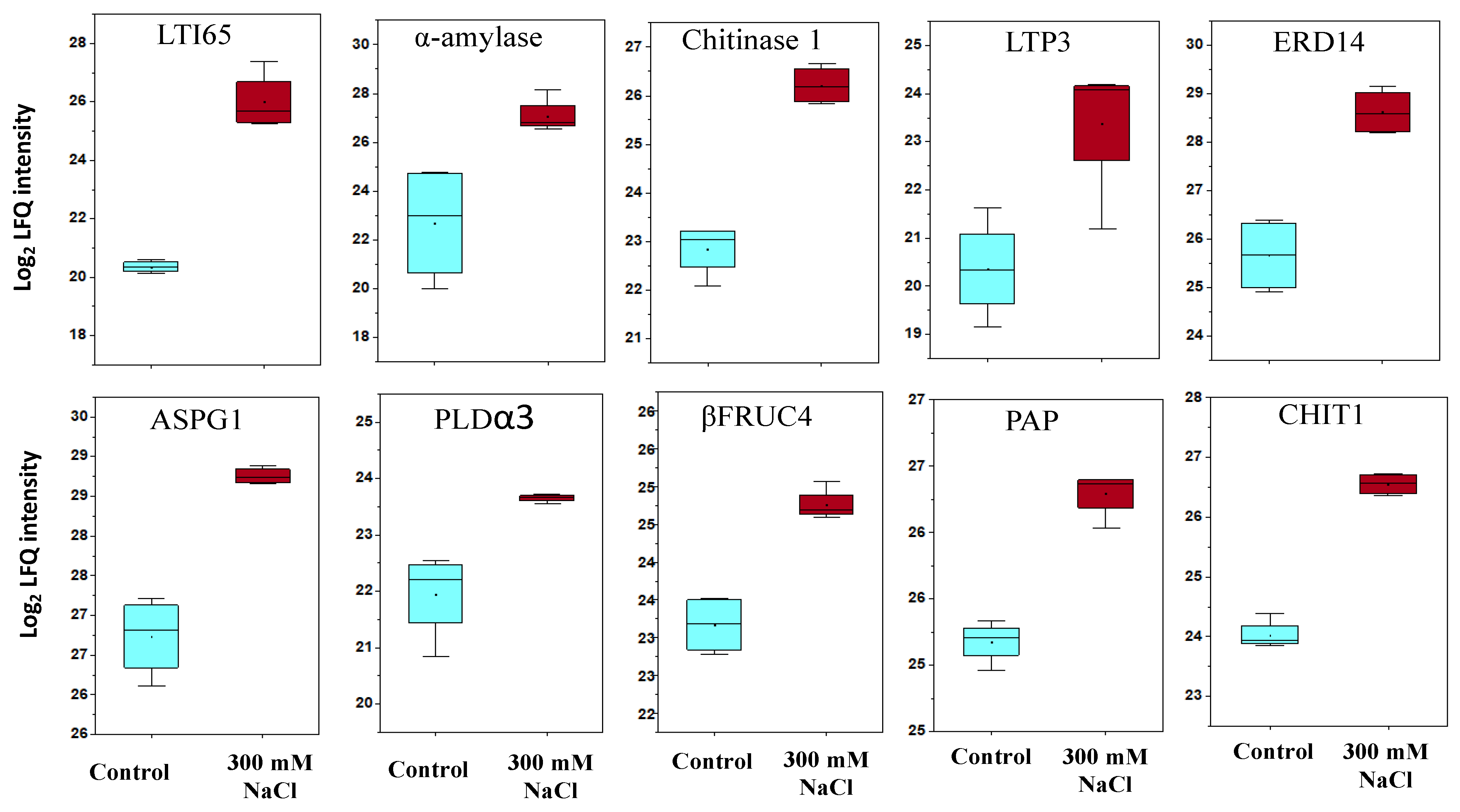
2.4. Differentially Abundant Proteins in Response to Salt Stress
2.5. Proteins with a Role in Stomatal Movement
2.6. Protein Involved in Response to ABA in GC Proteome
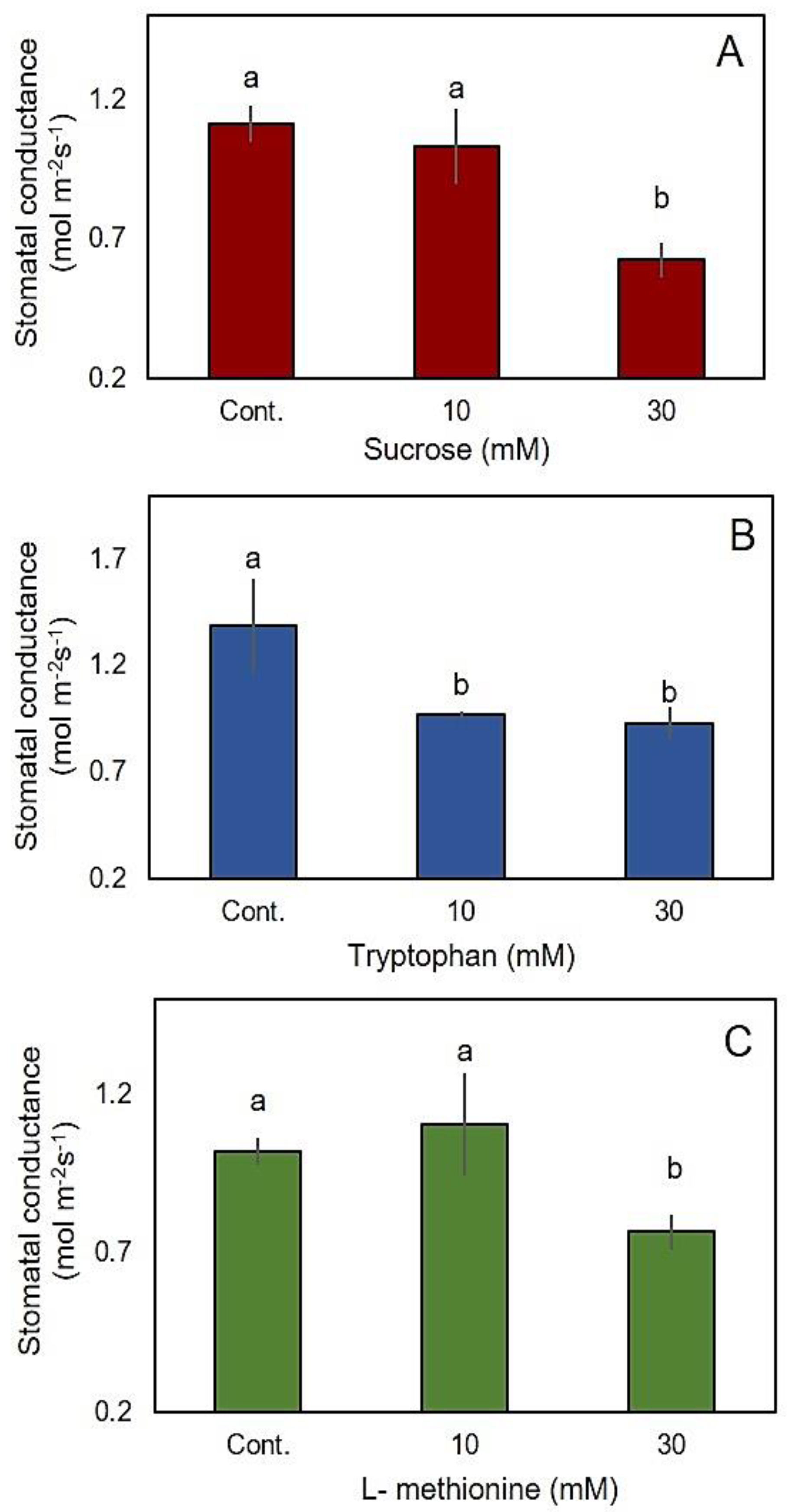
2.7. The Effect of Sucrose, Tryptophan and L-Methionine on Stomatal Conductance
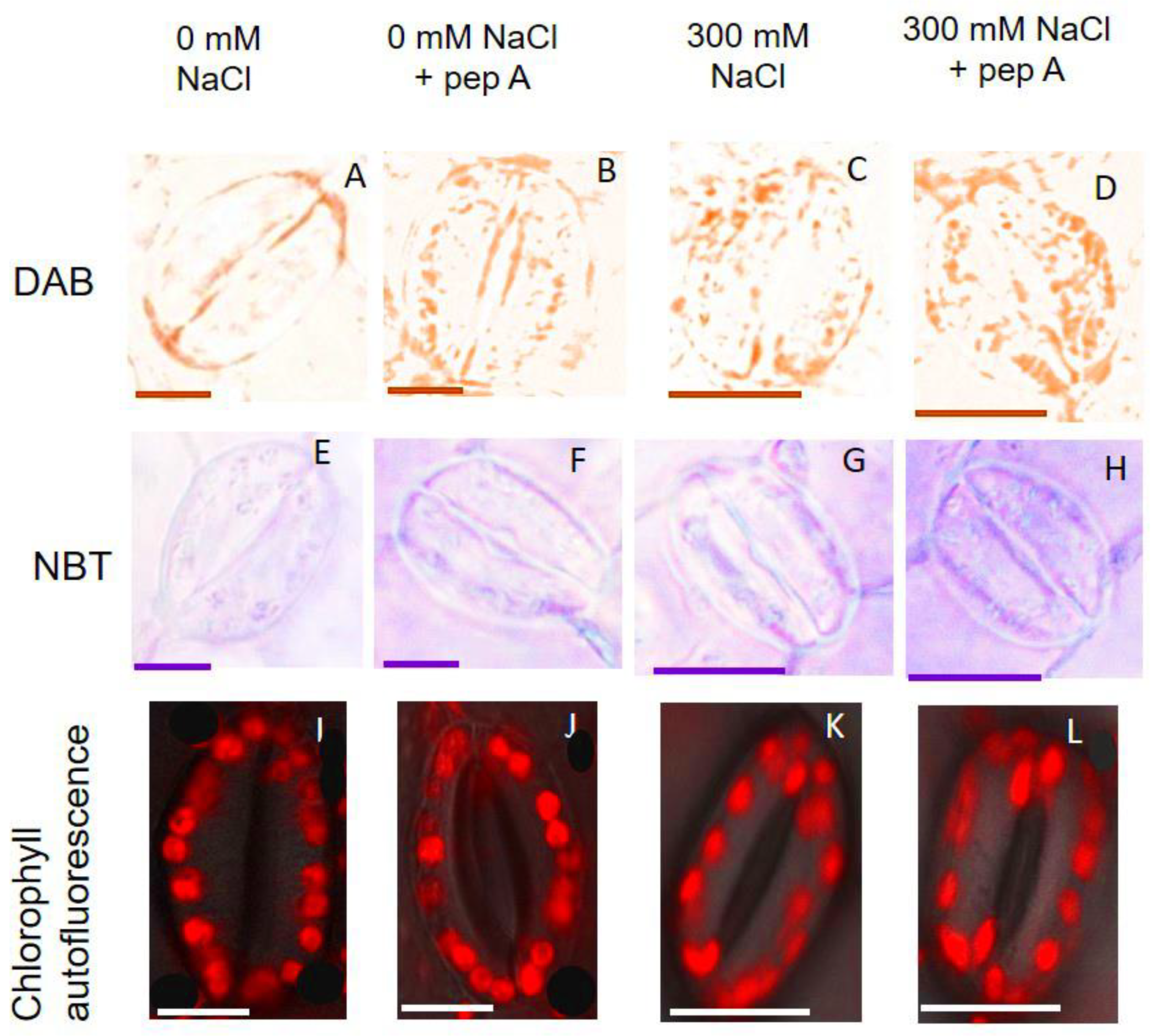
2.8. The Effect of Salinity and Pepstatin A on Quinoa Growth and Stomatal Traits
3. Discussion
3.1. Most Abundant Proteins in the GCs Isolated from Quinoa
3.2. Photosynthesis in the GCs
3.3. Stress and Defence-Related Proteins
3.4. Sucrose, Tryptophan and L-Methionine Induced Stomatal Closure
3.5. Aspartic Proteinases Are Important in Mitigating Oxidative Stress of GCs
4. Materials and Methods
4.1. Growth Conditions and GC Preparation
4.2. Label-Free Quantitative (LFQ) Proteomic Analysis of Quinoa GCs:
4.3. Determination of Relative Protein Abundance and Statistical Analysis
4.4. Stomatal Conductance in Plants Treated with Sucrose, Tryptophan, and L-Methionine
4.5. Exogenous Application of Pepstatin A
4.6. In Situ Detection of O2− and H2O2 in GCs and Imaging
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| DAB | 3,3′-diaminobenzidine |
| FDR | False Discovery Rate |
| GC | guard cells |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| NBT | nitro blue tetrazolium |
| NCBI | National Center for Biotechnology Information |
| ROS | Reactive oxygen species |
References
- Ache, P.; Bauer, H.; Kollist, H.; Al-Rasheid, K.A.; Lautner, S.; Hartung, W.; Hedrich, R. Stomatal action directly feeds back on leaf turgor: New insights into the regulation of the plant water status from non-invasive pressure probe measurements. Plant J. 2010, 62, 1072–1082. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.M.; Woodward, F.I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef]
- Lawson, T.; Blatt, M.R. Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency. Plant Physiol. 2014, 164, 1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kong, Z.; Liu, J.; Zhang, P.; Wang, Q.; Huan, X.; Li, L.; Qin, P. Non-targeted metabolomics of quinoa seed filling period based on liquid chromatography-mass spectrometry. Food Res. Int. 2020, 137, 109743. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Costa, J.M.; Zarrouk, O.; Pinheiro, C.; Lopes, C.M.; Pereira, J.S. Controlling stomatal aperture in semi-arid regions-The dilemma of saving water or being cool? Plant Sci. 2016, 251, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Li, D.H.; Zhou, R.; Wang, X.; Dossa, K.; Wang, L.H.; Zhang, Y.X.; Yu, J.Y.; Gong, H.H.; Zhang, X.R.; et al. Transcriptome and metabolome analyses of two contrasting sesame genotypes reveal the crucial biological pathways involved in rapid adaptive response to salt stress. BMC Plant Biol. 2019, 19, 66. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, R.; Shabala, S. Stomata in a saline world. Curr. Opin. Plant Biol. 2018, 46, 87–95. [Google Scholar] [CrossRef]
- Kiani-Pouya, A.; Rasouli, F.; Bazihizina, N.; Zhang, H.; Hedrich, R.; Shabala, S. A large-scale screening of quinoa accessions reveals an important role of epidermal bladder cells and stomatal patterning in salinity tolerance. Environ. Exp. Bot. 2019, 168, 103885. [Google Scholar] [CrossRef]
- Takemori, A.; Nakashima, T.; Omura, H.; Tanaka, Y.; Nakata, K.; Nonami, H.; Takemori, N. Quantitative assay of targeted proteome in tomato trichome glandular cells using a large-scale selected reaction monitoring strategy. Plant Methods 2019, 15, 40. [Google Scholar] [CrossRef] [PubMed]
- Barkla, B.J.; Vera-Estrella, R.; Raymond, C. Single-cell-type quantitative proteomic and ionomic analysis of epidermal bladder cells from the halophyte model plant Mesembryanthemum crystallinum to identify salt-responsive proteins. BMC Plant Biol. 2016, 16, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geilfus, C.M.; Lan, J.; Carpentier, S. Dawn regulates guard cell proteins in Arabidopsis thaliana that function in ATP production from fatty acid beta-oxidation. Plant Mol. Biol. 2018, 98, 525–543. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.M.; Dai, S.J.; McClung, S.; Yan, X.F.; Chen, S.X. Functional differentiation of Brassica napus guard cells and mesophyll cells revealed by comparative proteomics. Mol. Cell Proteomics 2009, 8, 752–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adolf, V.I.; Jacobsen, S.-E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Lopez-Marques, R.L.; Norrevang, A.F.; Ache, P.; Moog, M.; Visintainer, D.; Wendt, T.; Osterberg, J.T.; Dockter, C.; Jorgensen, M.E.; Salvador, A.T.; et al. Prospects for the accelerated improvement of the resilient crop quinoa. J. Exp. Bot. 2020, 71, 5333–5347. [Google Scholar] [CrossRef]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef]
- Zhao, Z.X.; Zhang, W.; Stanley, B.A.; Assmann, S.M. Functional proteomics of Arabidopsis thaliana guard cells uncovers new stomatal signaling pathways. Plant Cell 2008, 20, 3210–3226. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhu, M.M.; Zhu, N.; Strul, J.M.; Dufresne, C.P.; Schneider, J.D.; Harmon, A.C.; Chen, S.X. Identification of thioredoxin targets in guard cell enriched epidermal peels using cysTMT proteomics. J. Proteomics 2016, 133, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Geng, S.; Yu, B.; Zhu, N.; Dufresne, C.; Chen, S. Metabolomics and proteomics of Brassica napus guard cells in response to low CO2. Front. Mol. Biosci. 2017, 4, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.X.; Stanley, B.A.; Zhang, W.; Assmann, S.M. ABA-Regulated G Protein Signaling in Arabidopsis Guard Cells: A Proteomic Perspective. J. Proteome Res. 2010, 9, 1637–1647. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, F.; Kiani-Pouya, A.; Li, L.; Zhang, H.; Chen, Z.; Hedrich, R.; Wilson, R.; Shabala, S. Sugar beet (Beta vulgaris) guard cells responses to salinity stress: A proteomic analysis. Int. J. Mol. Sci. 2020, 21, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, X.M.; Zhou, L.; He, Y.; Wang, D.; Qi, Y.H.; Jiang, D.A. Rubisco activase is also a multiple responder to abiotic stresses in rice. PLoS ONE 2015, 10, e0140934. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Kim, B.K.; Kwon, S.J.; Jin, H.C.; Park, O.K. Arabidopsis GDSL lipase 2 plays a role in pathogen defense via negative regulation of auxin signaling. Biochem. Bioph. Res. Co. 2009, 379, 1038–1042. [Google Scholar] [CrossRef]
- Schwacke, R.; Ponce-Soto, G.Y.; Krause, K.; Bolger, A.M.; Arsova, B.; Hallab, A.; Gruden, K.; Stitt, M.; Bolger, M.E.; Usadel, B. MapMan4: A refined protein classification and annotation framework applicable to multi-omics data analysis. Mol. Plant 2019, 12, 879–892. [Google Scholar] [CrossRef]
- Cotelle, V.; Leonhardt, N. 14-3-3 proteins in guard cell signaling. Front. Plant Sci. 2015, 6, 1210. [Google Scholar] [CrossRef] [Green Version]
- Kretzschmar, T.; Burla, B.; Lee, Y.; Martinoia, E.; Nagy, R. Functions of ABC transporters in plants. Essays Biochem. 2011, 50, 145–160. [Google Scholar]
- Hubbard, K.E.; Webb, A.A. Circadian rhythms in stomata: Physiological and molecular aspects. In Rhythms in Plants, 2nd ed.; Mancuso, S., Shabala, S., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 231–255. [Google Scholar]
- Willmer, C.M.; Grammatikopoulos, G.; LascÈve, G.; Vavasseur, A. Characterization of the vacuolar-type H+-ATPase from guard cell protoplasts of Commelina. J. Exp. Bot. 1995, 46, 383–389. [Google Scholar] [CrossRef]
- Taji, T.; Seki, M.; Satou, M.; Sakurai, T.; Kobayashi, M.; Ishiyama, K.; Narusaka, Y.; Narusaka, M.; Zhu, J.K.; Shinozaki, K. Comparative genomics in salt tolerance between Arabidopsis and Arabidopsis-related halophyte salt cress using Arabidopsis microarray. Plant Physiol. 2004, 135, 1697–1709. [Google Scholar] [CrossRef] [Green Version]
- Laloum, T.; Martín, G.; Duque, P. Alternative splicing control of abiotic stress responses. Trends Plant Sci. 2018, 23, 140–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Jiang, H.; Xu, Y.; Li, H.; Wu, X.; Xie, Q.; Li, C. The CCCH-type zinc finger proteins AtSZF1 and AtSZF2 regulate salt stress responses in Arabidopsis. Plant Cell Physiol. 2007, 48, 1148–1158. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhong, Z.; Zhang, Y.; Xu, L.; Feng, S.; Rayatpisheh, S.; Wohlschlegel, J.A.; Wang, Z.; Jacobsen, S.E.; Ausin, I. NAP1-RELATED PROTEIN1 and 2 negatively regulate H2A.Z abundance in chromatin in Arabidopsis. Nat. Commun. 2020, 11, 2887. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Bulgakov, V.P.; Jinn, T.L. Pectin Methylesterases: Cell wall remodeling proteins are required for plant response to heat stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.C.; Wu, H.C.; Wang, Y.D.; Liu, C.H.; Lin, C.C.; Luo, D.L.; Jinn, T.L. PECTIN METHYLESTERASE34 Contributes to heat tolerance through its role in promoting stomatal movement. Plant Physiol. 2017, 174, 748–763. [Google Scholar] [CrossRef]
- Ringli, C.; Keller, B.; Ryser, U. Glycine-rich proteins as structural components of plant cell walls. Cell Mol. Life Sci. 2001, 58, 1430–1441. [Google Scholar] [CrossRef]
- Bose, J.; Munns, R.; Shabala, S.; Gilliham, M.; Pogson, B.; Tyerman, S.D. Chloroplast function and ion regulation in plants growing on saline soils: Lessons from halophytes. J. Exp. Bot. 2017, 68, 3129–3143. [Google Scholar] [CrossRef]
- Naranjo, M.A.; Forment, J.; Roldan, M.; Serrano, R.; Vicente, O. Overexpression of Arabidopsis thaliana LTL1, a salt-induced gene encoding a GDSL-motif lipase, increases salt tolerance in yeast and transgenic plants. Plant Cell Environ. 2006, 29, 1890–1900. [Google Scholar] [CrossRef]
- Le, J.; Liu, X.G.; Yang, K.Z.; Chen, X.L.; Zou, J.J.; Wang, H.Z.; Wang, M.; Vanneste, S.; Morita, M.; Tasaka, M.; et al. Auxin transport and activity regulate stomatal patterning and development. Nat. Commun. 2014, 5, 3090. [Google Scholar] [CrossRef] [Green Version]
- Thiel, G.; Blatt, M.R.; Fricker, M.D.; White, I.R.; Millner, P. Modulation of K+ channels in vicia stomatal guard-cells by peptide homologs to the auxin-binding protein-C terminus. Proc. Natl. Acad. Sci. USA 1993, 90, 11493–11497. [Google Scholar] [CrossRef] [Green Version]
- Gehring, C.A.; McConchie, R.M.; Venis, M.A.; Parish, R.W. Auxin-binding-protein antibodies and peptides influence stomatal opening and alter cytoplasmic pH. Planta 1998, 205, 581–586. [Google Scholar] [CrossRef]
- Azoulay-Shemer, T.; Palomares, A.; Bagheri, A.; Israelsson-Nordstrom, M.; Engineer, C.B.; Bargmann, B.O.; Stephan, A.B.; Schroeder, J.I. Guard cell photosynthesis is critical for stomatal turgor production, yet does not directly mediate CO2− and ABA-induced stomatal closing. Plant J. 2015, 83, 567–581. [Google Scholar] [CrossRef] [Green Version]
- Iwai, S.; Ogata, S.; Yamada, N.; Onjo, M.; Sonoike, K.; Shimazaki, K.I. Guard cell photosynthesis is crucial in abscisic acid-induced stomatal closure. Plant Direct 2019, 3, e00137. [Google Scholar] [CrossRef] [Green Version]
- Lüttge, U.; Beck, E.; Bartels, D. Plant dessication tolerance. In Ecological Studies Book 215; Springer: Berlin, Germany, 2011. [Google Scholar]
- Nylander, M.; Svensson, J.; Tapio Palva, E.; Welin, B.V. Stress-induced accumulation and tissue-specific localization of dehydrins in Arabidopsis thaliana. Plant Mol. Biol. 2001, 45, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Hyun, T.K.; van der Graaff, E.; Albacete, A.; Eom, S.H.; Grosskinsky, D.K.; Bohm, H.; Janschek, U.; Rim, Y.; Ali, W.W.; Kim, S.Y.; et al. The Arabidopsis PLAT domain protein1 is critically involved in abiotic stress tolerance. PLoS ONE 2014, 9, e112946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaashyap, M.; Ford, R.; Kudapa, H.; Jain, M.; Edwards, D.; Varshney, R.; Mantri, N. Differential regulation of genes involved in root morphogenesis and cell wall modification is associated with salinity tolerance in chickpea. Sci. Rep. 2018, 8, 4855. [Google Scholar] [CrossRef]
- Saucedo-Garcia, M.; Gavilanes-Ruiz, M.; Arce-Cervantes, O. Long-chain bases, phosphatidic acid, MAPKs, and reactive oxygen species as nodal signal transducers in stress responses in Arabidopsis. Front. Plant Sci. 2015, 6, 55. [Google Scholar]
- Yao, X.; Xiong, W.; Ye, T.T.; Wu, Y. Overexpression of the aspartic protease ASPG1 gene confers drought avoidance in Arabidopsis. J. Exp. Bot. 2012, 63, 2579–2593. [Google Scholar] [CrossRef] [Green Version]
- Granot, D.; Kelly, G. Evolution of guard-cell theories: The story of sugars. Trends Plant Sci. 2019, 24, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Outlaw, W.H.; Manchester, J. Guard cell starch concentration quantitatively related to stomatal aperture. Plant Physiol. 1979, 64, 79–82. [Google Scholar] [CrossRef] [Green Version]
- Kelly, G.; Moshelion, M.; David-Schwartz, R.; Halperin, O.; Wallach, R.; Attia, Z.; Belausov, E.; Granot, D. Hexokinase mediates stomatal closure. Plant J. 2013, 75, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Kottapalli, J.; David-Schwartz, R.; Khamaisi, B.; Brandsma, D.; Lugassi, N.; Egbaria, A.; Kelly, G.; Granot, D. Sucrose-induced stomatal closure is conserved across evolution. PLoS ONE 2018, 13, e0205359. [Google Scholar] [CrossRef] [PubMed]
- Rasouli, F.; Kiani-Pouya, A.; Tahir, A.; Shabala, L.; Chen, Z.; Shabala, S. A comparative analysis of stomatal traits and photosynthetic responses in closely related halophytic and glycophytic species under saline conditions. Environ. Exp. Bot. 2021, 181, 104300. [Google Scholar] [CrossRef]
- Jaarsma, R.; de Vries, R.S.; de Boer, A.H. Effect of salt stress on growth, Na+ accumulation and proline metabolism in potato (Solanum tuberosum) cultivars. PLoS ONE 2013, 8, e60183. [Google Scholar] [CrossRef]
- Cuin, T.A.; Shabala, S. Amino acids regulate salinity-induced potassium efflux in barley root epidermis. Planta 2007, 225, 753–761. [Google Scholar] [CrossRef]
- Sharma, U.D.; Rai, V.K. Modulation of osmotic closure of stomata, stomatal resistance and K+ fluxes by exogenous amino acids in Vicia faba L. Leaves. Biochem. Physiol. Pfl. 1989, 185, 369–376. [Google Scholar] [CrossRef]
- Kong, D.; Hu, H.C.; Okuma, E.; Lee, Y.; Lee, H.S.; Munemasa, S.; Cho, D.; Ju, C.; Pedoeim, L.; Rodriguez, B.; et al. L-Met activates Arabidopsis GLR Ca2+ channels upstream of ROS production and regulates stomatal movement. Cell Rep. 2016, 17, 2553–2561. [Google Scholar] [CrossRef]
- Simões, I.; Faro, C. Structure and function of plant aspartic proteinases. Eur. J. Biochem. 2004, 271, 2067–2075. [Google Scholar] [CrossRef] [Green Version]
- Soares, A.; Ribeiro Carlton, S.M.; Simoes, I. Atypical and nucellin-like aspartic proteases: Emerging players in plant developmental processes and stress responses. J. Exp. Bot. 2019, 70, 2059–2076. [Google Scholar] [CrossRef]
- Guo, R.; Zhao, J.; Wang, X.; Guo, C.; Li, Z.; Wang, Y.; Wang, X. Constitutive expression of a grape aspartic protease gene in transgenic Arabidopsis confers osmotic stress tolerance. Plant Cell Tissue Organ Cult. 2015, 121, 275–287. [Google Scholar] [CrossRef]
- Williams, M.; Woodward, F.I.; Baldocchi, D.D.; Ellsworth, D.S. Leaf to landscape. In Photosynthetic Adaptation; Smith, W.K., Vogelmann, T.C., Critchley, C., Eds.; Springer: New York, NY, USA, 2004; pp. 133–168. [Google Scholar]
- Rawlings, N.D.; Barrett, A.J. Families of aspartic peptidases, and those of unknown catalytic mechanism. Method Enzymol. 1995, 28, 105–120. [Google Scholar]
- Yamauchi, S.; Mano, S.; Oikawa, K.; Hikino, K.; Teshima, K.M.; Kimori, Y.; Nishimura, M.; Shimazaki, K.I.; Takemiya, A. Autophagy controls reactive oxygen species homeostasis in guard cells that is essential for stomatal opening. Proc. Natl. Acad. Sci. USA 2019, 116, 19187–19192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, C.; Chen, A.; Xiao, L.; Muller, H.M.; Ache, P.; Haberer, G.; Zhang, M.; Jia, W.; Deng, P.; Huang, R.; et al. A high-quality genome assembly of quinoa provides insights into the molecular basis of salt bladder-based salinity tolerance and the exceptional nutritional value. Cell Res. 2017, 27, 1327–1340. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmockel, S.M.; Li, B.; Borm, T.J.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasouli, F.; Kiani-Pouya, A.; Zhang, H.; Shabala, S. Developing and validating protocols for mechanical isolation of guard-cell enriched epidermal peels for omics studies. Funct. Plant Biol. 2020, 47, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Moggridge, S.; Muller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 2019, 14, 68–85. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Term | Term | Query Item | FDR * |
|---|---|---|---|
| GO:0006412 | translation | 122 | 2.4 × 10−22 |
| GO:0008152 | metabolic process | 854 | 3.1 × 10−18 |
| GO:0006091 | generation of precursor metabolites and energy | 46 | 4.5 × 10−15 |
| GO:0015979 | photosynthesis | 39 | 3.9 × 10−12 |
| GO:0005975 | carbohydrate metabolic process | 115 | 4.4 × 10−12 |
| GO:0009056 | catabolic process | 59 | 4.4 × 10−10 |
| GO:0009058 | biosynthetic process | 253 | 9.6 × 10−7 |
| GO:0006950 | response to stress | 76 | 1.8 × 10−4 |
| Protein Code | Name | Signal Type |
|---|---|---|
| XP_021772273 | mitochondrial proton/calcium exchanger protein-like | Calcium |
| XP_021720398 | Calcium-binding EF-hand family protein | Calcium |
| XP_021730224 | probable calcium-binding protein CML13 | Calcium |
| XP_021743759 | calcium-binding allergen Ole e 8-like | Calcium |
| XP_021724195 | calcium-binding protein CML49 | Calcium |
| XP_021740220 | cryptochrome-1-like isoform X3 | Calcium |
| XP_021761282 | calnexin homolog | Calcium |
| XP_021742145 | serine/threonine protein phosphatase 2A | Calcium |
| XP_021751211 | 14-3-3-like protein | 14.3.3 |
| XP_021775698 | 14-3-3-like protein D | 14.3.3 |
| XP_021772761 | GTP-binding protein SAR1A | G-proteins |
| XP_021772956 | mitochondrial Rho GTPase 1 | G-proteins |
| XP_021760299 | dynamin-related protein 5A | G-proteins |
| XP_021768021 | ras-related protein RABD2c-like | G-proteins |
| XP_021763216 | guanylate-binding protein 2-like | G-proteins |
| XP_021738598 | ras-related protein Rab7-like | G-proteins |
| XP_021731959 | nuclear pore complex protein NUP50A-like | G-proteins |
| XP_021765995 | guanylate-binding protein 3-like | G-proteins |
| XP_021736812 | nuclear pore complex protein NUP50A-like | G-proteins |
| XP_021735409 | nucleolin 1-like | G-proteins |
| XP_021748152 | RAN GTPase-activating protein 2 | G-proteins |
| XP_021736812 | nuclear pore complex protein NUP50A | G-proteins |
| XP_021735409 | nucleolin 1-like | G-proteins |
| XP_021748152 | RAN GTPase-activating protein 2 | G-proteins |
| XP_021751681 | G- nucleotide diphosphate dissociation inhibitor | G-proteins |
| XP_021716254 | phototropin-1-like isoform X1 | Light |
| XP_021740244 | protein EXORDIUM-like | Light |
| XP_021774295 | phytochrome B-like | Light |
| XP_021750479 | NAD(P)-binding Rossmann-fold superfamily | Light |
| XP_021738894 | COP9 signalosome complex subunit 5a-like | Light |
| XP_021714332 | PLC-like phosphodiesterases superfamily | MAP kinases |
| XP_021774801 | mitogen-activated protein kinase MMK1 | MAP kinases |
| XP_021754221 | mitogen-activated protein kinase MMK2 | MAP kinases |
| XP_021723308 | Leucine-rich repeat protein kinase family protein | RK. LRR III |
| XP_021746237 | LRR receptor-like ser/thre-protein kinase | RK. LRR VI |
| XP_021749023 | inactive LLR receptor-like protein kinase | RK. LRR VII |
| XP_021775620 | leucine-rich repeat receptor-like protein kinase | RK. LRR VII |
| XP_021718843 | leucine-rich repeat receptor-like protein kinase | RK. LRR VII |
| XP_021772032 | DNA damage-repair/toleration protein DRT100 | RK. LRR XI |
| Accession No. | Name | Transporter |
|---|---|---|
| XP_021715294 | plasma membrane-associated cation-binding protein | cation |
| XP_021762724 | ABC transporter F family member 4-like | ABC |
| XP_021743351 | ABC transporter C family member 2-like | ABC |
| XP_021776035 | ABC transporter G family member 22-like | ABC |
| XP_021766195 | ABC transporter I family member 19-like | ABC |
| XP_021720939 | calcium-transporting ATPase 4 | calcium |
| XP_021751211 | calcium-transporting ATPase 10 | calcium |
| XP_021744756 | bifunctional monothiol glutaredoxin-S16 | calcium |
| XP_021760697 | pyrophosphate-energized vacuolar membrane H+ pump | H+ pump |
| XP_021769151 | probable aquaporin PIP1-4 | PIP |
| XP_021765658 | mitochondrial carnitine/acylcarnitine carrier-like protein | metabolite |
| XP_021738293 | mitochondrial dicarboxylate/tricarboxylate transporter DTC | metabolite |
| XP_021756032 | mitochondrial phosphate carrier protein 3, mitochondrial | metabolite |
| XP_021757591 | cation/H+ antiporter 18-like | cation |
| XP_021736780 | plastidic ATP/ADP-transporter-like | Misc |
| XP_021738681 | V-type proton ATPase subunit a3-like | ATPases |
| XP_021730105 | V-type proton ATPase subunit C-like | ATPases |
| XP_021761683 | ATPase 11, plasma membrane-type-like | ATPases |
| XP_021765334 | V-type proton ATPase subunit G 1-like | ATPases |
| XP_021739675 | V-type proton ATPase catalytic subunit A | ATPases |
| XP_021732700 | V-type proton ATPase catalytic subunit A-like | ATPases |
| XP_021738896 | plasma membrane ATPase 4-like | ATPases |
| XP_021762284 | V-type proton ATPase subunit d2 | ATPases |
| XP_021765533 | V-type proton ATPase subunit B 2 | ATPases |
| XP_021754298 | V-type proton ATPase subunit E-like | ATPases |
| XP_021772280 | V-type proton ATPase subunit H-like isoform X2 | ATPases |
| XP_021714458 | mitochondrial outer membrane protein porin 2-like | Porin |
| XP_021761841 | mitochondrial import receptor subunit TOM40-1-like | Porin |
| XP_021758463 | mitochondrial outer membrane protein porin of 34 kDa | Porin |
| XP_021717525 | K+ efflux antiporter 2, chloroplastic-like | Potassium |
| XP_021762166 | probable voltage-gated potassium channel subunit beta | Potassium |
| XP_021753247 | monosaccharide-sensing protein 2-like | Sugar |
| XP_021726328 | sugar carrier protein C-like | Sugar |
| XP_021757156 | plastidic glucose transporter 4-like | Sugar |
| XP_021760460 | sucrose transport protein-like isoform X1 | Sucrose |
| XP_021752898 | chloride channel protein CLC-b-like | anions |
| XP_021739774 | ATPase ASNA1 homolog | anions |
| XP_021772050 | ADP, ATP carrier protein 1, mitochondrial-like | cation |
| Accession No | Name | Fold Change NaCl/Control |
|---|---|---|
| XP_02174988 | abscisic acid receptor PYL2 | 1.1 |
| XP_021736717 | phospholipase D alpha 1-like | 3.3 * |
| XP_021739586 | GDPDL3-like | 2.1 * |
| XP_021760770 | ricin B-like lectin EULS3 | 2.0 * |
| XP_021754221 | MMK2-like | 1.6 * |
| XP_021759066 | GDPDL3-like | 1.5 * |
| XP_021738483 | OST1 | 1.3 * |
| XP_021723531 | hexokinase-1-like | 0.9 * |
| XP_021715572 | clathrin heavy chain 1-like | 0.8 * |
| XP_021723433 | plasma membrane ATPase 4-like | 0.5 * |
| XP_021761237 | calcium sensing receptor, chloroplastic-like | 0.5 * |
| XP_021734330 | protein flowering locus t-like | 0.3 * |
| XP_021776446 | protein thylakoid formation1 | 0.3 * |
| XP_021775775 | uncharacterized protein LOC110739633 | 0.2 * |
| XP_021776385 | protein phosphatase 2C | 0.4 |
| XP_021714641 | DNA-directed RNA polymerases II, IV and V | 2.5 |
| XP_021721800 | BLUS1-like | 2.2 |
| XP_021757551 | MMK2-like | 1.9 |
| XP_021723484 | serine/threonine-protein kinase STY8-like | 1.8 |
| XP_021760806 | carbonic anhydrase, chloroplastic | 1.7 |
| XP_021740220 | cryptochrome-1-like isoform X3 | 1.4 |
| XP_021753299 | phototropin-2-like | 1.3 |
| XP_021731588 | glycine-rich RNA-binding, ABA-inducible protein | 1.1 |
| XP_021774295 | phytochrome B-like | 1.1 |
| XP_021742577 | translationally-controlled tumor protein homolog | 1.1 |
| XP_021716254 | phototropin-1-like isoform X1 | 1.0 |
| XP_021741723 | carbonic anhydrase, chloroplastic-like | 1.0 |
| XP_021774801 | MMK1-like | 0.9 |
| XP_021760799 | phospholipase D alpha 1-like | 0.9 |
| XP_021758723 | chlorophyll a-b binding protein 3, chloroplastic | 0.7 |
| XP_021745518 | plasma membrane-associated cation-binding protein 1 | 0.7 |
| XP_021731589 | glycine-rich RNA-binding protein-like | 0.7 |
| XP_021723344 | phosphoglycerate mutase-like | 0.6 |
| XP_021760744 | vesicle-associated membrane protein 711 | 0.6 |
| XP_021769207 | clathrin heavy chain 1-like | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasouli, F.; Kiani-Pouya, A.; Shabala, L.; Li, L.; Tahir, A.; Yu, M.; Hedrich, R.; Chen, Z.; Wilson, R.; Zhang, H.; et al. Salinity Effects on Guard Cell Proteome in Chenopodium quinoa. Int. J. Mol. Sci. 2021, 22, 428. https://doi.org/10.3390/ijms22010428
Rasouli F, Kiani-Pouya A, Shabala L, Li L, Tahir A, Yu M, Hedrich R, Chen Z, Wilson R, Zhang H, et al. Salinity Effects on Guard Cell Proteome in Chenopodium quinoa. International Journal of Molecular Sciences. 2021; 22(1):428. https://doi.org/10.3390/ijms22010428
Chicago/Turabian StyleRasouli, Fatemeh, Ali Kiani-Pouya, Lana Shabala, Leiting Li, Ayesha Tahir, Min Yu, Rainer Hedrich, Zhonghua Chen, Richard Wilson, Heng Zhang, and et al. 2021. "Salinity Effects on Guard Cell Proteome in Chenopodium quinoa" International Journal of Molecular Sciences 22, no. 1: 428. https://doi.org/10.3390/ijms22010428