Admixture Analysis Using Genotyping-by-Sequencing Reveals Genetic Relatedness and Parental Lineage Distribution in Highbush Blueberry Genotypes and Cross Derivatives

 , , and
, , and
Abstract
:1. Introduction
2. Results
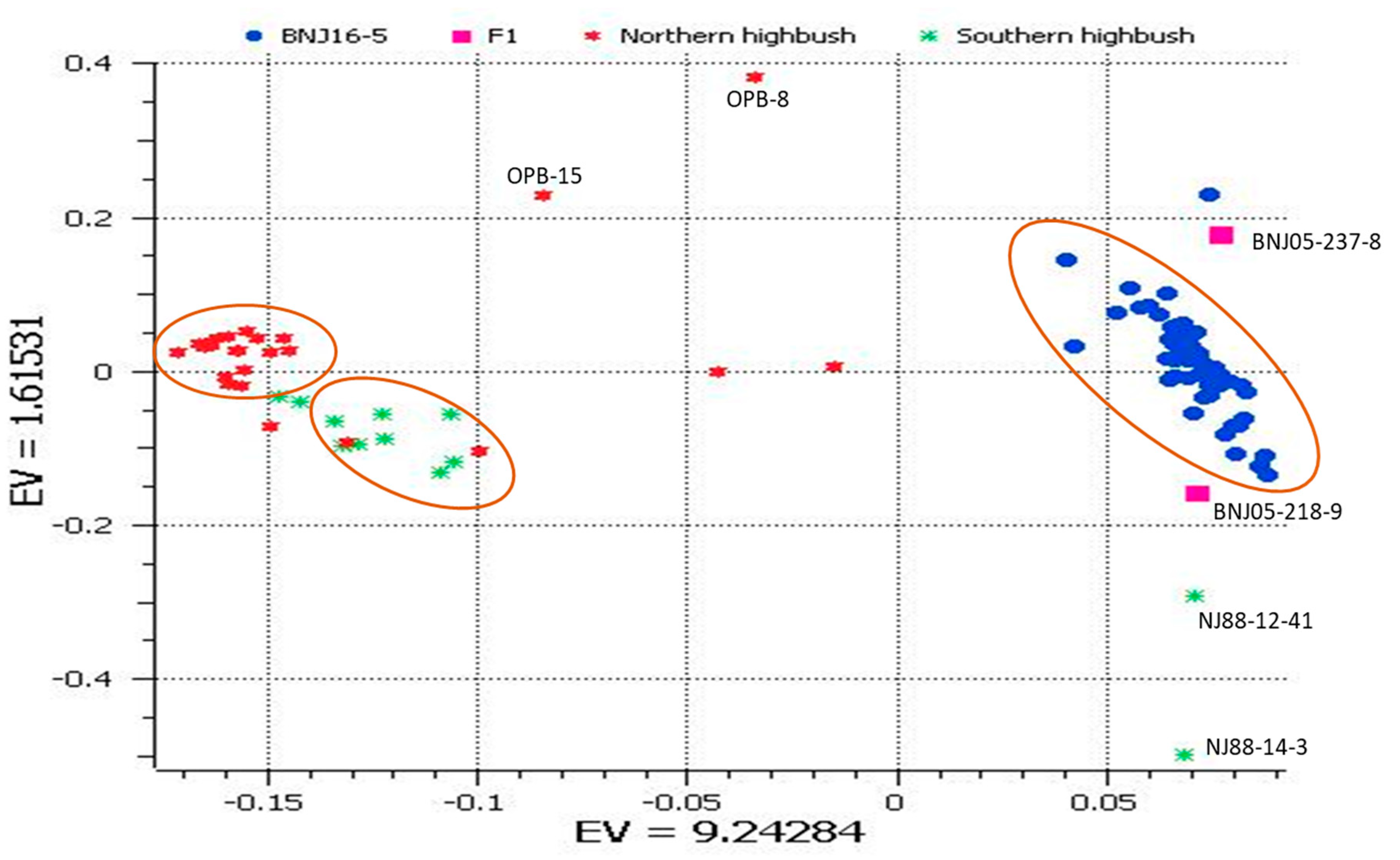
2.1. Principal Component Analysis (PCA)
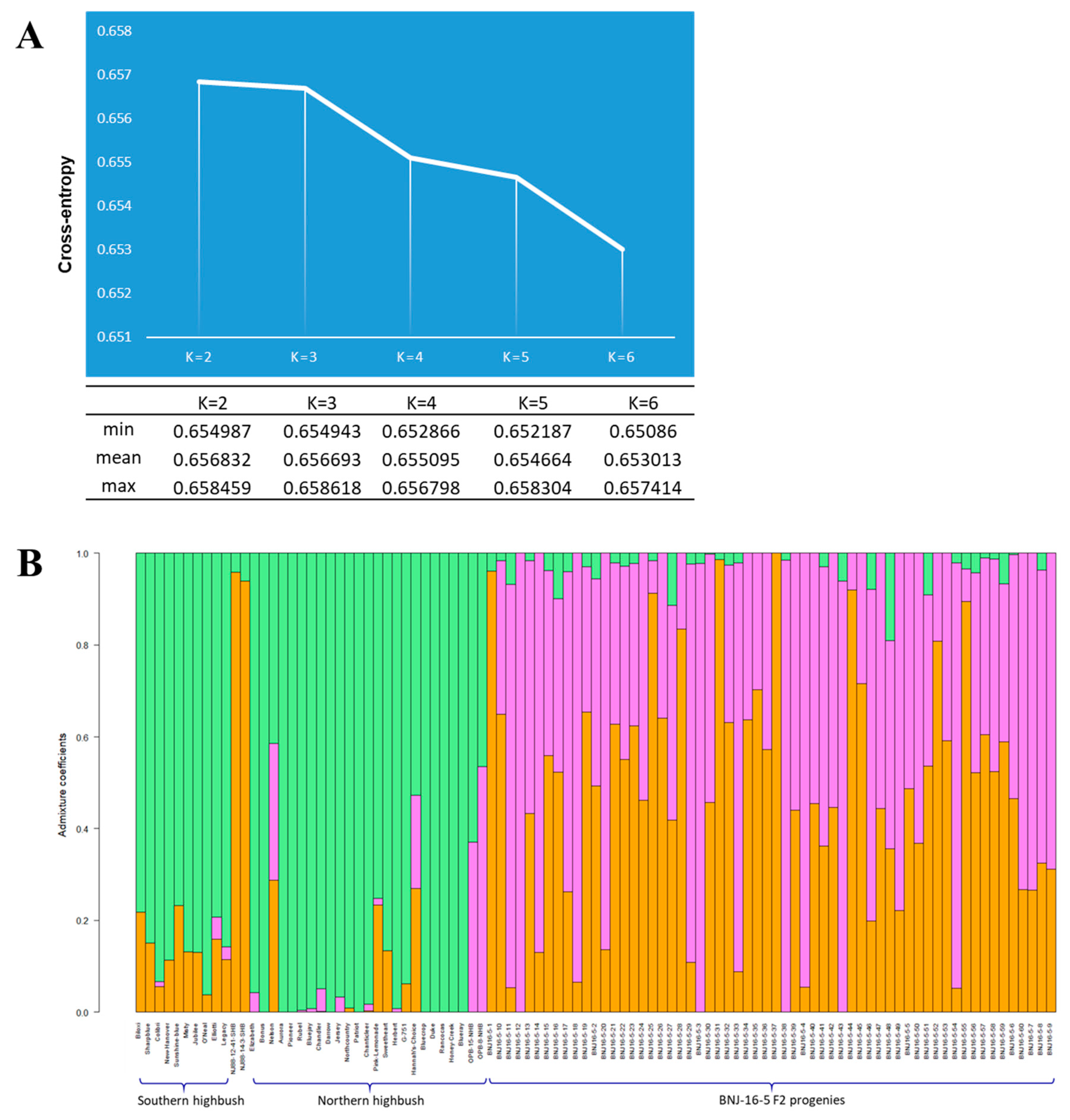
2.2. Admixture Analysis
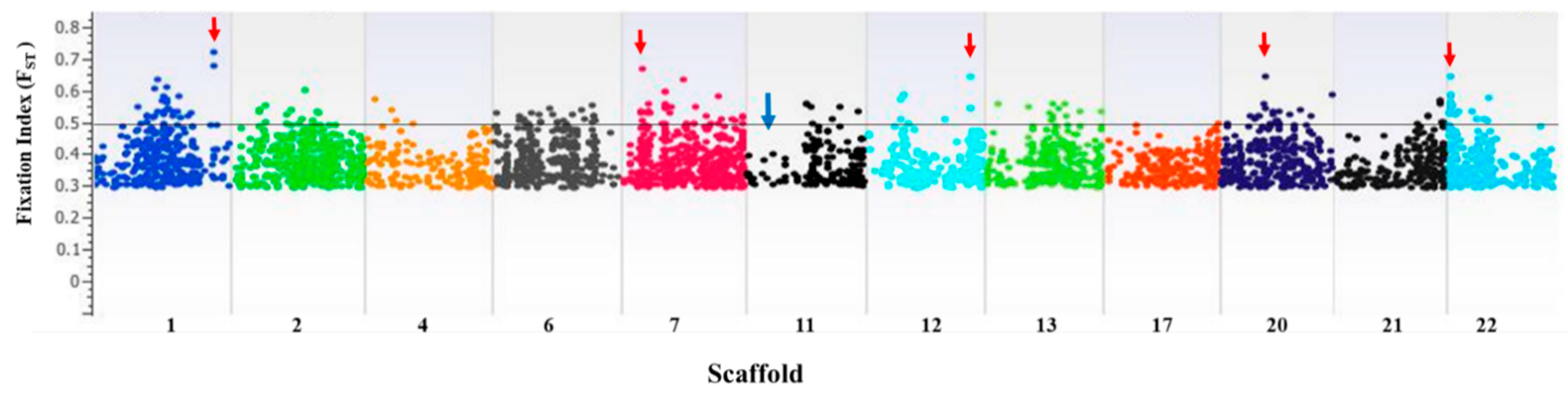
2.3. FST for Characterizing Selection Footprints
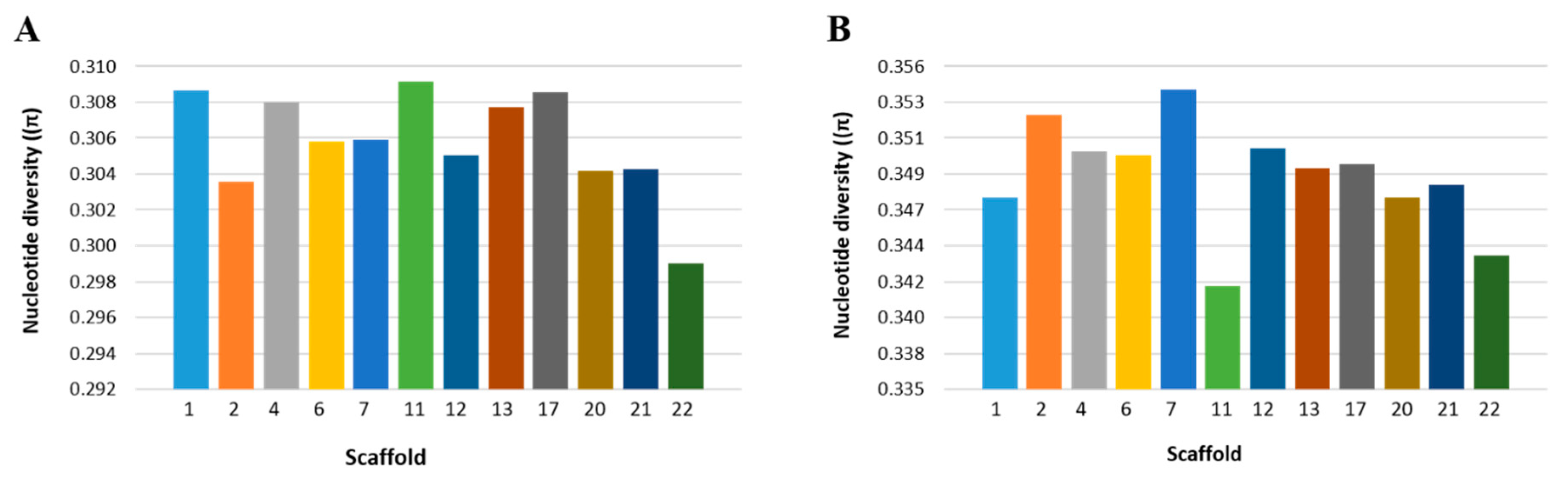
2.4. Nucleotide Diversity
2.5. LD and Haplotype Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. DNA Isolation
4.3. GBS Analysis
4.4. Sequence Alignment and SNP Identification
4.5. Principal Component Analysis
4.6. Admixture Analysis
4.7. Haplotype Block Analysis
4.8. Nucleotide Diversity Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galletta, G.; Ballington, J. Blueberries, cranberries and lingonberries. Fruit Breed. 1996, 2, 107. [Google Scholar]
- Ratnaparkhe, M.B. Blueberry. In Fruits and Nuts; Springer: Berlin/Heidelberg, Germany, 2007; pp. 217–227. [Google Scholar]
- Die, J.V.; Rowland, L.J. Advent of genomics in blueberry. Mol. Breed. 2013, 32, 493–504. [Google Scholar]
- Yang, B.; Kortesniemi, M. Clinical evidence on potential health benefits of berries. Curr. Opin. Food Sci. 2015, 2, 36–42. [Google Scholar] [CrossRef]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent research on the health benefits of blueberries and their anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [PubMed]
- Levi, A.; Rowland, L. Identifying blueberry cultivars and evaluating their genetic relationships using randomly amplified polymorphic DNA (RAPD) and simple sequence repeat-(SSR-) anchored primers. J. Am. Soc. Hortic. Sci. 1997, 122, 74–78. [Google Scholar]
- Coville, F.V. Improving the wild blueberry. In Yearbook of Agriculture; United States Department of Agriculture: Washington, DC, USA, 1937; pp. 559–574. [Google Scholar]
- Song, G.-Q.; Hancock, J.F. Vaccinium. In Wild Crop Relatives: Genomic and Breeding Resources; Springer: Berlin/Heidelberg, Germany, 2011; pp. 197–221. [Google Scholar]
- Ballington, J.R. Collection, utilization, and preservation of genetic resources in Vaccinium. HortScience 2001, 36, 206–213. [Google Scholar]
- Camp, W.H. The North American blueberries with notes on other groups of Vacciniaceae. Brittonia 1945, 5, 203–275. [Google Scholar]
- Camp, W.H. A survey of the American species of Vaccinium, subgenus Euvaccinium. Brittonia 1942, 4, 205–247. [Google Scholar]
- Megalos, B.S.; Ballington, J.R. Unreduced pollen frequencies versus hybrid production in diploid-tetraploid Vaccinium crosses. Euphytica 1988, 39, 271–278. [Google Scholar]
- Vander Kloet, S.P. The Genus Vaccinium in North America; Research Branch, Agriculture Canada, Canadian Government Publication Centre: Ottawa, ON, Canada, 1988.
- Bruederle, L.P.; Vorsa, N. Genetic differentiation of diploid blueberry, Vaccinium sect. Cyanococcus (Ericaceae). Syst. Bot. 1994, 19, 337–349. [Google Scholar]
- Bruederle, L.P.; Vorsa, N.; Ballington, J.R. Population genetic structure in diploid blueberry Vaccinium section Cyanococcus (Ericaceae). Am. J. Bot. 1991, 78, 230–237. [Google Scholar]
- Ballington, J.R.; Galletta, G.J. Comparative crossability of 4 diploid Vaccinium species. J. Am. Soc. Hortic. Sci. 1978, 103, 554–560. [Google Scholar]
- Chavez, D.J.; Lyrene, P.M. Effects of self-pollination and cross-pollination of Vaccinium darrowii (Ericaceae) and other low-chill blueberries. HortScience 2009, 44, 1538. [Google Scholar]
- Beers, L.; Rowland, L.J.; Drummond, F. Genetic diversity of lowbush blueberry throughout the United States in managed and non-managed populations. Agriculture 2019, 9, 113. [Google Scholar]
- Boches, P.; Bassil, N.V.; Rowland, L. Genetic diversity in the highbush blueberry evaluated with microsatellite markers. J. Am. Soc. Hortic. Sci. 2006, 131, 674–686. [Google Scholar]
- Retamales, J.B.; Hancock, J.F. Blueberries; CABI Publishing: Wallingford, UK, 2012. [Google Scholar]
- Vander Kloet, S.P. The taxonomy of Vaccinium Cyanococcus: A summation. Can. J. Bot. 1983, 61, 256–266. [Google Scholar]
- Skotte, L.; Korneliussen, T.S.; Albrechtsen, A. Estimating individual admixture proportions from next generation sequencing data. Genetics 2013, 195, 693–702. [Google Scholar]
- Bian, Y.; Ballington, J.; Raja, A.; Brouwer, C.; Reid, R.; Burke, M.; Wang, X.; Rowland, L.J.; Bassil, N.; Brown, A. Patterns of simple sequence repeats in cultivated blueberries (Vaccinium section Cyanococcus spp.) and their use in revealing genetic diversity and population structure. Mol. Breed. 2014, 34, 675–689. [Google Scholar]
- Bassil, N.; Bidani, A.; Hummer, K.; Rowland, L.J.; Olmstead, J.; Lyrene, P.; Richards, C. Assessing genetic diversity of wild southeastern North American Vaccinium species using microsatellite markers. Genet. Resour. Crop Evol. 2018, 65, 939–950. [Google Scholar]
- Rowland, L.J.; Mehra, S.; Dhanaraj, A.L.; Ogden, E.L.; Slovin, J.P.; Ehlenfeldt, M.K. Development of EST-PCR markers for DNA fingerprinting and genetic relationship studies in blueberry (Vaccinium, section Cyanococcus). J. Am. Soc. Hortic. Sci. 2003, 128, 682–690. [Google Scholar]
- Bell, D.J.; Rowland, L.J.; Polashock, J.J.; Drummond, F.A. Suitability of EST-PCR markers developed in highbush blueberry for genetic fingerprinting and relationship studies in lowbush blueberry and related species. J. Am. Soc. Hortic. Sci. 2008, 133, 701–707. [Google Scholar] [CrossRef] [Green Version]
- Zong, Y.; Kang, H.; Fang, Q.; Chen, X.; Zhou, M.; Ni, J.; Zhang, Y.; Wang, L.; Zhu, Y.; Guo, W. Phylogenetic relationship and genetic background of blueberry (Vaccinium spp.) based on retrotransposon-based SSAP molecular markers. Sci. Hortic. 2019, 247, 116–122. [Google Scholar] [CrossRef]
- Mackay, I.; Powell, W. Methods for linkage disequilibrium mapping in crops. Trends Plant Sci. 2007, 12, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Ranger, C.; Singh, A.; Johnson-Cicalese, J.; Polavarapu, S.; Vorsa, N. Intraspecific variation in aphid resistance and constitutive phenolics exhibited by the wild blueberry Vaccinium darrowii. J. Chem. Ecol. 2007, 33, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baloga, D.W.; Vorsa, N.; Lawter, L. Dynamic headspace gas chromatography—Mass spectrometry analysis of volatile flavor compounds from wild diploid blueberry species. In Fruit Flavors: Biogenesis, Characterization and Authentication, 1st ed.; Rousseff, R.L., Leahy, M.M., Eds.; ACS Publications: Washington, DC, USA, 1995. [Google Scholar]
- Wang, Y.; Fong, S.K.; Singh, A.P.; Vorsa, N.; Johnson-Cicalese, J. Variation of anthocyanins, proanthocyanidins, flavonols, and organic acids in cultivated and wild diploid blueberry species. HortScience 2019, 54, 576–585. [Google Scholar] [CrossRef] [Green Version]
- Colle, M.; Leisner, C.P.; Wai, C.M.; Ou, S.; Bird, K.A.; Wang, J.; Wisecaver, J.H.; Yocca, A.E.; Alger, E.I.; Tang, H.; et al. Haplotype-phased genome and evolution of phytonutrient pathways of tetraploid blueberry. GigaScience 2019, 8, giz012. [Google Scholar] [CrossRef] [Green Version]
- Frichot, E.; François, O. LEA: An R package for landscape and ecological association studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Michalska, A.; Łysiak, G. Bioactive compounds of blueberries: Post-harvest factors influencing the nutritional value of products. Int. J. Mol. Sci. 2015, 16, 18642–18663. [Google Scholar] [CrossRef]
- Eck, P.; Childers, N. Blueberry Culture; Rutgers University Press: New Brunswick, NJ, USA, 1966. [Google Scholar]
- Rodriguez-Saona, C.; Vincent, C.; Isaacs, R. Blueberry IPM: Past successes and future challenges. Annu. Rev. Entomol. 2019, 64, 95–114. [Google Scholar] [CrossRef]
- Lyrene, P.; Vorsa, N.; Ballington, J. Polyploidy and sexual polyploidization in the genus Vaccinium. Euphytica 2003, 133, 27–36. [Google Scholar] [CrossRef]
- Moore, J. Improving highbush blueberries by breeding and selection. Euphytica 1965, 14, 39–48. [Google Scholar]
- Ehlenfeldt, M. Domestication of the highbush blueberry at Whitesbog, New Jersey, 1911–1916. In Proceedings of the IX International Vaccinium Symposium 810, Corvallis, OR, USA, 14–18 July 2008; pp. 147–152. [Google Scholar]
- Rodriguez-Saona, C.; Cloonan, K.R.; Sanchez-Pedraza, F.; Zhou, Y.; Giusti, M.M.; Benrey, B. Differential susceptibility of wild and cultivated blueberries to an invasive frugivorous pest. J. Chem. Ecol. 2019, 45, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Mengist, M.F.; Grace, M.H.; Xiong, J.; Kay, C.D.; Bassil, N.; Hummer, K.; Ferruzzi, M.G.; Lila, M.A.; Iorizzo, M. Diversity in metabolites and fruit quality traits in blueberry enables ploidy and species differentiation and establishes a strategy for future genetic studies. Front. Plant Sci. 2020, 11, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debnath, S.C. Structured diversity using EST-PCR and EST-SSR markers in a set of wild blueberry clones and cultivars. Biochem. Syst. Ecol. 2014, 54, 337–347. [Google Scholar] [CrossRef]
- Brevis, P.A.; Bassil, N.V.; Ballington, J.R.; Hancock, J.F. Impact of wide hybridization on highbush blueberry breeding. J. Am. Soc. Hortic. Sci. 2008, 133, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Draper, A.D. Tetraploid hybrids from crosses of diploid, tetraploid and hexaploid Vaccinium species. In Proceedings of the Symposium on Vaccinium Culture in Europe 61, Hannover, Germany, 22 July 1976; pp. 33–36. [Google Scholar]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PloS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Wickland, D.P.; Battu, G.; Hudson, K.A.; Diers, B.W.; Hudson, M.E. A comparison of genotyping-by-sequencing analysis methods on low-coverage crop datasets shows advantages of a new workflow, GB-eaSy. BMC Bioinform. 2017, 18, 586. [Google Scholar] [CrossRef]
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigenanalysis. PloS Genet. 2006, 2, e190. [Google Scholar] [CrossRef]
- Frichot, E.; Mathieu, F.; Trouillon, T.; Bouchard, G.; François, O. Fast and efficient estimation of individual ancestry coefficients. Genetics 2014, 196, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Korneliussen, T.S.; Moltke, I.; Albrechtsen, A.; Nielsen, R. Calculation of Tajima’s D and other neutrality test statistics from low depth next-generation sequencing data. BMC Bioinform. 2013, 14, 289. [Google Scholar] [CrossRef] [Green Version]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 1358–1370. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosome | Chromosome Length (bp) | Raw SNPs | Filtered SNPs (MAF < 0.05; Call Rate < 0.9; DP > 3) | Average Number of Filtered SNPs per kb |
|---|---|---|---|---|
| VACCDSCAFF1 | 46,295,995 | 223,567 | 8719 | 5 |
| VACCDSCAFF2 | 44,818,276 | 188,522 | 8994 | 5 |
| VACCDSCAFF4 | 42,981,373 | 162,229 | 7758 | 6 |
| VACCDSCAFF6 | 42,795,824 | 190,595 | 7090 | 6 |
| VACCDSCAFF7 | 41,705,179 | 175,039 | 7783 | 5 |
| VACCDSCAFF11 | 40,122,599 | 211,211 | 8194 | 5 |
| VACCDSCAFF12 | 39,741,682 | 170,459 | 6191 | 6 |
| VACCDSCAFF13 | 39,652,356 | 177,072 | 7654 | 5 |
| VACCDSCAFF17 | 38,874,919 | 173,797 | 7901 | 5 |
| VACCDSCAFF20 | 37,996,905 | 187,631 | 7116 | 5 |
| VACCDSCAFF21 | 37,975,728 | 184,224 | 7395 | 5 |
| VACCDSCAFF22 | 37,315,645 | 199,693 | 7253 | 5 |
| Total Number of SNPs | 2,244,039 | 92,048 | ||
| Chromosome | Largest LD Block (kb) | |
|---|---|---|
| Northern Highbush Blueberry | Southern Highbush Blueberry | |
| VACCDSCAFF1 | 285 | 285 |
| VACCDSCAFF2 | 425 | 425 |
| VACCDSCAFF4 | 425 | 425 |
| VACCDSCAFF6 | 518 | 418 |
| VACCDSCAFF7 | 154 | 251 |
| VACCDSCAFF11 | 255 | 329 |
| VACCDSCAFF12 | 454 | 382 |
| VACCDSCAFF13 | 672 | 261 |
| VACCDSCAFF17 | 545 | 243 |
| VACCDSCAFF20 | 247 | 255 |
| VACCDSCAFF21 | 231 | 366 |
| VACCDSCAFF22 | 189 | 194 |
| Category | Genotype | Accession a | Ploidy | Taxon | Pedigree | Improvement Status |
|---|---|---|---|---|---|---|
| Tetraploid northern highbush (23) b | Elizabeth | 4× | Vc | (Katharine × Jersey) × Scammel | ||
| Bonus | PI 666839 | 4× | Vc | |||
| Nelson | PI 618100 | 4× | Vc | Bluecrop × G-107 | Cultivar | |
| Aurora | 4× | Vc | Brigitta Blue × Elliott | |||
| Pioneer | PI 554815 | 4× | Vc | Brooks × Sooy | Cultivar | |
| Rubel | PI 554817 | 4× | Vc | Selection from wild V. corymbosum in NJ selected from the pine barrens of NJ | Cultivar | |
| Bluejay | PI 554846 | 4× | Vc | Berkeley × Michigan Highbush Sel. 241 (Pioneer × Taylor) | Cultivar | |
| Chandler | PI 657260 | 4× | Vc | Darrow × M-23 | Cultivar | |
| Darrow | PI 618035 | 4× | Vc | F 72 × Bluecrop | Cultivar | |
| Jersey | PI 554897 | 4× | Vc | Rubel × Grover | Cultivar | |
| Northcountry | PI 554953 | 4× | Vc × Va | B6 (G65 × ‘Ashworth’ V. corymbosum) × R2P4 (open pollinated V. corymbosum × V. angustifolium hybrid) | Cultivar | |
| Patriot | PI 554843 | 4× | Vc | US 3 (Dixi × Mich LB-1) × Earliblue | Cultivar | |
| Chanticleer | PI 638765 | 4× | Vc | G-180 × MEUS 6620 | Cultivar | |
| Pink Lemonade | PI 641330 | 4× | Vc | NJ89-158-1 × Delite (V. ashei) | Cultivar | |
| Sweetheart | 4× | Vc | Cultivar | |||
| Herbert | PI 554805 | 4× | Vc | Stanley (Katharine × Rubel) × GS-149 (Jersey × Pioneer) | Cultivar | |
| G-751 | 4× | Vc | Wild material | |||
| Hannah’s Choice | PI 657259 | 4× | Vc | G-136 × G-358 | Cultivar | |
| Bluecrop | PI 554885 | 4× | Vc | GM-37 (Jersey × Pioneer) × CU-5 (Stanley × June) | Cultivar | |
| Duke | PI 554872 | 4× | Vc | G-100 (Ivanhoe × Earliblue) × 192-8 (E-30 × E-11) | Cultivar | |
| Rancocas | PI 554816 | 4× | Vc | 394Y (Brooks × Russell) × Rubel | Cultivar | |
| Honey Creek | 4× | Vc | ||||
| Blueray | PI 554887 | 4× | Vc | (Jersey × Pioneer) × (Stanley × June) | ||
| Tetraploid southern highbush (9) | Biloxi | PI 618193 | 4× | Vc | Sharpblue × US 329 [US210 (US67 × US132) × FL 4-76 (Bluecrop × 13-236)] | Cultivar |
| Sharpblue | PI 554948 | 4× | Vc | V. corymbosum × V. ashei & V. darrowii (Fla 61-5 × Fla 62-4) tetraploid | Cultivar | |
| Colibri | 4× | Vc | ||||
| New Hanover | 4× | Vc | ||||
| Sunshine blue | PI 555316 | Vc × Va | Avonblue OP | |||
| Misty | PI 555317 | 4× | Vc | Florida 67-I × Avonblue | ||
| Jubilee | PI 618195 | 4× | Vc | Sharpblue × MS60 [(Ashworth × Earliblue] × Bluecrop) × US-75] | Cultivar | |
| O’Neal | PI 554944 | 4× | Vc | Wolcott x Fla. 4-15 mainly V. corymbosum, some V. angustifolium, V. ashei, V. darrowii | Cultivar | |
| Legacy | PI 618164 | 4× | Vc | Elizabeth × (Fla. 4B × Bluecrop) | Cultivar | |
| Vaccinium elliottii (1) | Elliottii | PI 657176 | 2× | Ve | Wild material | |
| Vaccinium darrowii (2) | NJ88-12-41 | 2× | Vd | Wild material | ||
| NJ88-14-3 | 2× | Vd | Wild material | |||
| Vaccinium corymbosum (2) | NJOPB-8 | 2× | Vc | Wild material | ||
| NJOPB-15 | 2× | Vc | Wild material | |||
| F1 (2) | BNJ05-237-8 | Vc × Vd | Cross derivative | |||
| BNJ05-218-9 | Vd × Vc | Cross derivative | ||||
| F2 (60) | BNJ16-5 population | Cross derivatives |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulkarni, K.P.; Vorsa, N.; Natarajan, P.; Elavarthi, S.; Iorizzo, M.; Reddy, U.K.; Melmaiee, K. Admixture Analysis Using Genotyping-by-Sequencing Reveals Genetic Relatedness and Parental Lineage Distribution in Highbush Blueberry Genotypes and Cross Derivatives. Int. J. Mol. Sci. 2021, 22, 163. https://doi.org/10.3390/ijms22010163
Kulkarni KP, Vorsa N, Natarajan P, Elavarthi S, Iorizzo M, Reddy UK, Melmaiee K. Admixture Analysis Using Genotyping-by-Sequencing Reveals Genetic Relatedness and Parental Lineage Distribution in Highbush Blueberry Genotypes and Cross Derivatives. International Journal of Molecular Sciences. 2021; 22(1):163. https://doi.org/10.3390/ijms22010163
Chicago/Turabian StyleKulkarni, Krishnanand P., Nicholi Vorsa, Purushothaman Natarajan, Sathya Elavarthi, Massimo Iorizzo, Umesh K. Reddy, and Kalpalatha Melmaiee. 2021. "Admixture Analysis Using Genotyping-by-Sequencing Reveals Genetic Relatedness and Parental Lineage Distribution in Highbush Blueberry Genotypes and Cross Derivatives" International Journal of Molecular Sciences 22, no. 1: 163. https://doi.org/10.3390/ijms22010163