Brain Site-Specific Inhibitory Effects of the GLP-1 Analogue Exendin-4 on Alcohol Intake and Operant Responding for Palatable Food

 ,
, 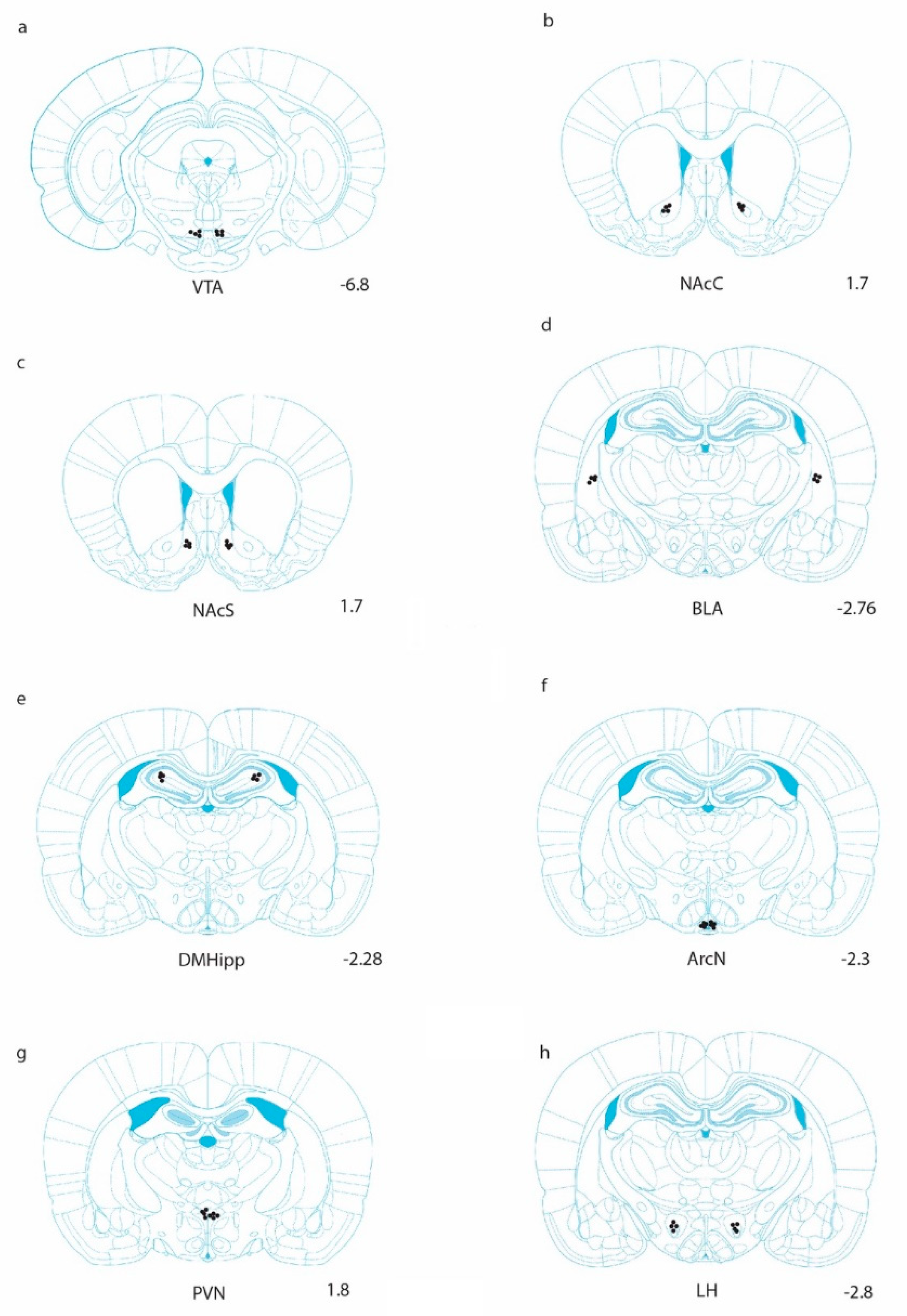{kind=link}
{kind=link}
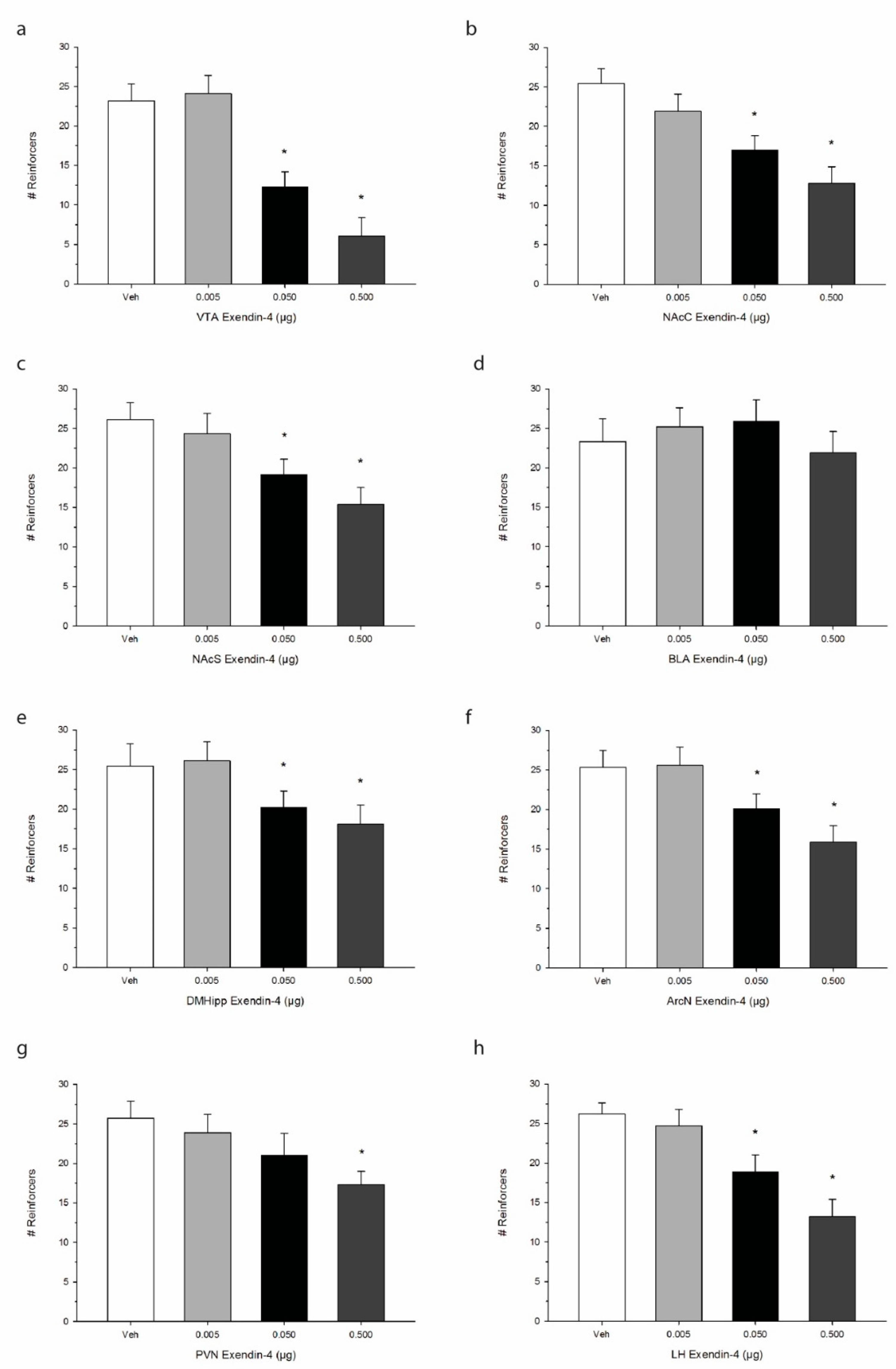{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Alcohol Consummatory Studies
2.2. Operant Conditioning Paradigms
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drug
4.3. Stereotaxic Surgery
4.4. Design and Procedure
4.4.1. Alcohol Consummatory Investigations
4.4.2. Operant Conditioning Paradigm
4.4.3. Histological and Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
References
- Lipari, R.N.; Park-Lee, E. Key Substance Use and Mental Health Indicators in the United States: Results from the 2018 National Survey on Drug Use and Health; Substance Abuse and Mental Health Services Administration: Rockville, MD, USA, 2019. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar] [CrossRef]
- Nguyen, L.-C.; Durazzo, T.C.; Dwyer, C.L.; Rauch, A.A.; Humphreys, K.; Williams, L.M.; Padula, C.B. Predicting Relapse after Alcohol Use Disorder Treatment in a High-Risk Cohort: The Roles of Anhedonia and Smoking. J. Psychiatr. Res. 2020, 126, 1–7. [Google Scholar] [CrossRef]
- Cannella, N.; Ubaldi, M.; Masi, A.; Bramucci, M.; Roberto, M.; Bifone, A.; Ciccocioppo, R. Building Better Strategies to Develop New Medications in Alcohol Use Disorder: Learning from Past Success and Failure to Shape a Brighter Future. Neurosci. Biobehav. Rev. 2019, 103, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Heilig, M.; Egli, M. Pharmacological Treatment of Alcohol Dependence: Target Symptoms and Target Mechanisms. Pharmacol. Ther. 2006, 111, 855–876. [Google Scholar] [CrossRef] [PubMed]
- Jerlhag, E. GLP-1 Signaling and Alcohol-Mediated Behaviors; Preclinical and Clinical Evidence. Neuropharmacology 2018, 136, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Bornebusch, A.B.; Fink-Jensen, A.; Wörtwein, G.; Seeley, R.J.; Thomsen, M. Glucagon-Like Peptide-1 Receptor Agonist Treatment Does Not Reduce Abuse-Related Effects of Opioid Drugs. eNeuro 2019, 6. [Google Scholar] [CrossRef]
- Górska, A.; Arciszewski, M.B. Distribution and Function of Glucagon-like Peptide 1 (GLP-1) in the Digestive Tract of Mammals and the Clinical Use of Its Analogues. Med. Weter. 2020, 76, 382–388. [Google Scholar] [CrossRef]
- Manandhar, B.; Ahn, J.-M. Glucagon-like Peptide-1 (GLP-1) Analogs: Recent Advances, New Possibilities, and Therapeutic Implications. J. Med. Chem. 2015, 58, 1020–1037. [Google Scholar] [CrossRef]
- Müller, T.D.; Finan, B.; Bloom, S.R.; D’Alessio, D.; Drucker, D.J.; Flatt, P.R.; Fritsche, A.; Gribble, F.; Grill, H.J.; Habener, J.F.; et al. Glucagon-like Peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef]
- Kalra, S.; Das, A.K.; Sahay, R.K.; Baruah, M.P.; Tiwaskar, M.; Das, S.; Chatterjee, S.; Saboo, B.; Bantwal, G.; Bhattacharya, S.; et al. Consensus Recommendations on GLP-1 RA Use in the Management of Type 2 Diabetes Mellitus: South Asian Task Force. Diabetes Ther. 2019, 10, 1645–1717. [Google Scholar] [CrossRef] [Green Version]
- Bae, C.; Song, J. The Role of Glucagon-Like Peptide 1 (GLP1) in Type 3 Diabetes: GLP-1 Controls Insulin Resistance, Neuroinflammation and Neurogenesis in the Brain. IJMS 2017, 18, 2493. [Google Scholar] [CrossRef] [Green Version]
- Merchenthaler, I.; Lane, M.; Shughrue, P. Distribution of Pre-pro-Glucagon and Glucagon-like Peptide-1 Receptor Messenger RNAs in the Rat Central Nervous System. J. Comp. Neurol. 1999, 403, 261–280. [Google Scholar] [CrossRef]
- Alvarez, E.; Roncero, I.; Chowen, J.A.; Thorens, B.; Blázquez, E. Expression of the Glucagon-Like Peptide-1 Receptor Gene in Rat Brain. J. Neurochem. 2002, 66, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Cork, S.C.; Richards, J.E.; Holt, M.K.; Gribble, F.M.; Reimann, F.; Trapp, S. Distribution and Characterisation of Glucagon-like Peptide-1 Receptor Expressing Cells in the Mouse Brain. Mol. Metab. 2015, 4, 718–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, G.; Roland, B.; Tomaselli, K.; Dolman, C.S.; Lowe, C.; Heilig, J.S. Glucagon-like Peptide-1 in the Rat Brain: Distribution of Expression and Functional Implication. J. Comp. Neurol. 2013, 521, 2235–2261. [Google Scholar] [CrossRef] [PubMed]
- Heppner, K.M.; Kirigiti, M.; Secher, A.; Paulsen, S.J.; Buckingham, R.; Pyke, C.; Knudsen, L.B.; Vrang, N.; Grove, K.L. Expression and Distribution of Glucagon-Like Peptide-1 Receptor MRNA, Protein and Binding in the Male Nonhuman Primate (Macaca Mulatta) Brain. Endocrinology 2015, 156, 255–267. [Google Scholar] [CrossRef] [Green Version]
- Schick, R.R.; Zimmermann, J.P.; vorm Walde, T.; Schusdziarra, V. Glucagon-like Peptide 1-(7–36) Amide Acts at Lateral and Medial Hypothalamic Sites to Suppress Feeding in Rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R1427–R1435. [Google Scholar] [CrossRef] [Green Version]
- McMahon, L.R.; Wellman, P.J. PVN Infusion of GLP-1-(7—36) Amide Suppresses Feeding but Does Not Induce Aversion or Alter Locomotion in Rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 274, R23–R29. [Google Scholar] [CrossRef]
- Abtahi, S.; VanderJagt, H.L.; Currie, P.J. The Glucagon-like Peptide-1 Analog Exendin-4 Antagonizes the Effect of Acyl Ghrelin on the Respiratory Exchange Ratio. NeuroReport 2016, 27, 992–996. [Google Scholar] [CrossRef]
- Abtahi, S.; Howell, E.; Salvucci, J.T.; Bastacky, J.M.R.; Dunn, D.P.; Currie, P.J. Exendin-4 Antagonizes the Metabolic Action of Acylated Ghrelinergic Signaling in the Hypothalamic Paraventricular Nucleus. Gen. Comp. Endocrinol. 2019, 270, 75–81. [Google Scholar] [CrossRef]
- Crovetti, R.; Lucchetti, F.; Mancia, M.; Mariotti, M.; Porrini, M.; Spinnler, P.; Testolin, G. Food-Intake After Amygdaloid Lesion in Rats. Nutr. Res. 1995, 15, 565–570. [Google Scholar] [CrossRef]
- Ganaraj, B.; Jeganathan, P.S. Involvement of Basolateral Nucleus & Central Nucleus of Amygdala in the Regulation of Ingestive Behaviour in Rat. Indian J. Med Res. 1998, 108, 98–103. [Google Scholar] [PubMed]
- Anderberg, R.H.; Anefors, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. Dopamine Signaling in the Amygdala, Increased by Food Ingestion and GLP-1, Regulates Feeding Behavior. Physiol. Behav. 2014, 136, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Ren, W.; Li, H.; Hou, Y. Inhibitory Effects of Peripheral Administration of Exendin-4 on Food Intake Are Attenuated by Lesions of the Central Nucleus of Amygdala. Brain Res. Bull. 2019, 148, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Oka, J.-I.; Goto, N.; Kameyama, T. Glucagon-like Peptide-1 Modulates Neuronal Activity in the Rat’s Hippocampus. NeuroReport 1999, 10, 1643–1646. [Google Scholar] [CrossRef]
- Hsu, T.M.; Hahn, J.D.; Konanur, V.R.; Lam, A.; Kanoski, S.E. Hippocampal GLP-1 Receptors Influence Food Intake, Meal Size, and Effort-Based Responding for Food through Volume Transmission. Neuropsychopharmacology 2015, 40, 327–337. [Google Scholar] [CrossRef]
- Hsu, T.M.; Noble, E.E.; Liu, C.M.; Cortella, A.M.; Konanur, V.R.; Suarez, A.N.; Reiner, D.J.; Hahn, J.D.; Hayes, M.R.; Kanoski, S.E. A Hippocampus to Prefrontal Cortex Neural Pathway Inhibits Food Motivation through Glucagon-like Peptide-1 Signaling. Mol. Psychiatry 2018, 23, 1555–1565. [Google Scholar] [CrossRef] [Green Version]
- Alhadeff, A.L.; Rupprecht, L.E.; Hayes, M.R. GLP-1 Neurons in the Nucleus of the Solitary Tract Project Directly to the Ventral Tegmental Area and Nucleus Accumbens to Control for Food Intake. Endocrinology 2012, 153, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Dickson, S.L.; Shirazi, R.H.; Hansson, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. The Glucagon-Like Peptide 1 (GLP-1) Analogue, Exendin-4, Decreases the Rewarding Value of Food: A New Role for Mesolimbic GLP-1 Receptors. J. Neurosci. 2012, 32, 4812–4820. [Google Scholar] [CrossRef]
- Dossat, A.M.; Lilly, N.; Kay, K.; Williams, D.L. Glucagon-Like Peptide 1 Receptors in Nucleus Accumbens Affect Food Intake. J. Neurosci. 2011, 31, 14453–14457. [Google Scholar] [CrossRef] [Green Version]
- Dossat, A.M.; Diaz, R.; Gallo, L.; Panagos, A.; Kay, K.; Williams, D.L. Nucleus Accumbens GLP-1 Receptors Influence Meal Size and Palatability. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1314–E1320. [Google Scholar] [CrossRef] [Green Version]
- Howell, E.; Baumgartner, H.; Zallar, L.; Selva, J.; Engel, L.; Currie, P. Glucagon-Like Peptide-1 (GLP-1) and 5-Hydroxytryptamine 2c (5-HT2c) Receptor Agonists in the Ventral Tegmental Area (VTA) Inhibit Ghrelin-Stimulated Appetitive Reward. IJMS 2019, 20, 889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egecioglu, E.; Steensland, P.; Fredriksson, I.; Feltmann, K.; Engel, J.A.; Jerlhag, E. The Glucagon-like Peptide 1 Analogue Exendin-4 Attenuates Alcohol Mediated Behaviors in Rodents. Psychoneuroendocrinology 2013, 38, 1259–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi, R.H.; Dickson, S.L.; Skibicka, K.P. Gut Peptide GLP-1 and Its Analogue, Exendin-4, Decrease Alcohol Intake and Reward. PLoS ONE 2013, 8, e61965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørensen, G.; Caine, S.B.; Thomsen, M. Effects of the GLP-1 Agonist Exendin-4 on Intravenous Ethanol Self-Administration in Mice. Alcohol Clin. Exp. Res. 2016, 40, 2247–2252. [Google Scholar] [CrossRef]
- Vallöf, D.; Maccioni, P.; Colombo, G.; Mandrapa, M.; Jörnulf, J.W.; Egecioglu, E.; Engel, J.A.; Jerlhag, E. The Glucagon-like Peptide 1 Receptor Agonist Liraglutide Attenuates the Reinforcing Properties of Alcohol in Rodents. Addict. Biol. 2016, 21, 422–437. [Google Scholar] [CrossRef]
- Vallöf, D.; Kalafateli, A.L.; Jerlhag, E. Long-Term Treatment with a Glucagon-like Peptide-1 Receptor Agonist Reduces Ethanol Intake in Male and Female Rats. Transl. Psychiatry 2020, 10, 238. [Google Scholar] [CrossRef]
- Thomsen, M.; Holst, J.J.; Molander, A.; Linnet, K.; Ptito, M.; Fink-Jensen, A. Effects of Glucagon-like Peptide 1 Analogs on Alcohol Intake in Alcohol-Preferring Vervet Monkeys. Psychopharmacology 2019, 236, 603–611. [Google Scholar] [CrossRef]
- Vallöf, D.; Vestlund, J.; Jerlhag, E. Glucagon-like Peptide-1 Receptors within the Nucleus of the Solitary Tract Regulate Alcohol-Mediated Behaviors in Rodents. Neuropharmacology 2019, 149, 124–132. [Google Scholar] [CrossRef]
- Dixon, T.N.; McNally, G.P.; Ong, Z.Y. Glucagon-Like Peptide-1 Receptor Signaling in the Ventral Tegmental Area Reduces Alcohol Self-Administration in Male Rats. Alcohol. Clin. Exp. Res. 2020, 44, 2118–2129. [Google Scholar] [CrossRef]
- Vallöf, D.; Kalafateli, A.L.; Jerlhag, E. Brain Region Specific Glucagon-like Peptide-1 Receptors Regulate Alcohol-Induced Behaviors in Rodents. Psychoneuroendocrinology 2019, 103, 284–295. [Google Scholar] [CrossRef]
- Abtahi, S.; Howell, E.; Currie, P.J. Accumbal Ghrelin and Glucagon-like Peptide 1 Signaling in Alcohol Reward in Female Rats. Neuroreport 2018, 29, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. Paxino’s and Watson’s The Rat Brain in Stereotaxic Coordinates, 7th ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Boston, MA, USA, 2014. [Google Scholar]
- Egecioglu, E.; Engel, J.A.; Jerlhag, E. The Glucagon-Like Peptide 1 Analogue, Exendin-4, Attenuates the Rewarding Properties of Psychostimulant Drugs in Mice. PLoS ONE 2013, 8, e69010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Highfield, D.; Clements, A.; Shalev, U.; McDonald, R.; Featherstone, R.; Stewart, J.; Shaham, Y. Involvement of the Medial Septum in Stress-Induced Relapse to Heroin Seeking in Rats: Medial Septum and Stress-Induced Relapse. Eur. J. Neurosci. 2000, 12, 1705–1713. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, G.; Reddy, I.A.; Weikop, P.; Graham, D.L.; Stanwood, G.D.; Wortwein, G.; Galli, A.; Fink-Jensen, A. The Glucagon-like Peptide 1 (GLP-1) Receptor Agonist Exendin-4 Reduces Cocaine Self-Administration in Mice. Physiol. Behav. 2015, 149, 262–268. [Google Scholar] [CrossRef]
- Müller, T.D.; Nogueiras, R.; Andermann, M.L.; Andrews, Z.B.; Anker, S.D.; Argente, J.; Batterham, R.L.; Benoit, S.C.; Bowers, C.Y.; Broglio, F.; et al. Ghrelin. Mol. Metab. 2015, 4, 437–460. [Google Scholar] [CrossRef]
- Skibicka, K.P.; Hansson, C.; Alvarez-Crespo, M.; Friberg, P.A.; Dickson, S.L. Ghrelin Directly Targets the Ventral Tegmental Area to Increase Food Motivation. Neuroscience 2011, 180, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Skibicka, K.P.; Hansson, C.; Egecioglu, E.; Dickson, S.L. Role of Ghrelin in Food Reward: Impact of Ghrelin on Sucrose Self-Administration and Mesolimbic Dopamine and Acetylcholine Receptor Gene Expression. Addict Biol 2012, 17, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, H.D.; Mietlicki-Baase, E.G.; Ige, K.Y.; Maurer, J.J.; Reiner, D.J.; Zimmer, D.J.; Van Nest, D.S.; Guercio, L.A.; Wimmer, M.E.; Olivos, D.R.; et al. Glucagon-Like Peptide-1 Receptor Activation in the Ventral Tegmental Area Decreases the Reinforcing Efficacy of Cocaine. Neuropsychopharmacol 2016, 41, 1917–1928. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, M.; Dencker, D.; Wörtwein, G.; Weikop, P.; Egecioglu, E.; Jerlhag, E.; Fink-Jensen, A.; Molander, A. The Glucagon-like Peptide 1 Receptor Agonist Exendin-4 Decreases Relapse-like Drinking in Socially Housed Mice. Pharmacol. Biochem. Behav. 2017, 160, 14–20. [Google Scholar] [CrossRef]
- Fuchs, R.A.; Evans, K.A.; Ledford, C.C.; Parker, M.P.; Case, J.M.; Mehta, R.H.; See, R.E. The Role of the Dorsomedial Prefrontal Cortex, Basolateral Amygdala, and Dorsal Hippocampus in Contextual Reinstatement of Cocaine Seeking in Rats. Neuropsychopharmacol 2005, 30, 296–309. [Google Scholar] [CrossRef]
- McGlinchey, E.M.; Aston-Jones, G. Dorsal Hippocampus Drives Context-Induced Cocaine Seeking via Inputs to Lateral Septum. Neuropsychopharmacol. 2018, 43, 987–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, C.T.; Mitra, S.; Auerbach, B.D.; Wang, Z.-J.; Martin, J.A.; Stewart, A.F.; Gobira, P.H.; Iida, M.; An, C.; Cobb, M.M.; et al. Neuroadaptations in the Dorsal Hippocampus Underlie Cocaine Seeking during Prolonged Abstinence. Proc. Natl. Acad. Sci. USA 2020, 117, 26460–26469. [Google Scholar] [CrossRef] [PubMed]
- Puaud, M.; Higuera-Matas, A.; Brunault, P.; Everitt, B.J.; Belin, D. The Basolateral Amygdala to Nucleus Accumbens Core Circuit Mediates the Conditioned Reinforcing Effects of Cocaine-Paired Cues on Cocaine Seeking. Biol. Psychiatry 2020, S0006322320318084. [Google Scholar] [CrossRef] [PubMed]
- Schönfeld, L.-M.; Schäble, S.; Zech, M.-P.; Kalenscher, T. 5-HT1A Receptor Agonism in the Basolateral Amygdala Increases Mutual-Reward Choices in Rats. Sci. Rep. 2020, 10, 16622. [Google Scholar] [CrossRef]
- Falco, A.M.; Bergstrom, H.C.; Bachus, S.E.; Smith, R.F. Persisting Changes in Basolateral Amygdala MRNAs after Chronic Ethanol Consumption. Physiol. Behav. 2009, 96, 169–173. [Google Scholar] [CrossRef]
- Sharp, B.M. Basolateral Amygdala and Stress-Induced Hyperexcitability Affect Motivated Behaviors and Addiction. Transl. Psychiatry 2017, 7, e1194. [Google Scholar] [CrossRef]
- López-Ferreras, L.; Richard, J.E.; Noble, E.E.; Eerola, K.; Anderberg, R.H.; Olandersson, K.; Taing, L.; Kanoski, S.E.; Hayes, M.R.; Skibicka, K.P. Lateral Hypothalamic GLP-1 Receptors Are Critical for the Control of Food Reinforcement, Ingestive Behavior and Body Weight. Mol. Psychiatry 2018, 23, 1157–1168. [Google Scholar] [CrossRef] [Green Version]
- Secher, A.; Jelsing, J.; Baquero, A.F.; Hecksher-Sørensen, J.; Cowley, M.A.; Dalbøge, L.S.; Hansen, G.; Grove, K.L.; Pyke, C.; Raun, K.; et al. The Arcuate Nucleus Mediates GLP-1 Receptor Agonist Liraglutide-Dependent Weight Loss. J. Clin. Invest. 2014, 124, 4473–4488. [Google Scholar] [CrossRef] [Green Version]
- Barbano, M.F.; Wang, H.-L.; Morales, M.; Wise, R.A. Feeding and Reward Are Differentially Induced by Activating GABAergic Lateral Hypothalamic Projections to VTA. J. Neurosci. 2016, 36, 2975–2985. [Google Scholar] [CrossRef] [Green Version]
- Gigante, E.D.; Benaliouad, F.; Zamora-Olivencia, V.; Wise, R.A. Optogenetic Activation of a Lateral Hypothalamic-Ventral Tegmental Drive-Reward Pathway. PLoS ONE 2016, 11, e0158885. [Google Scholar] [CrossRef]
- Jennings, J.H.; Ung, R.L.; Resendez, S.L.; Stamatakis, A.M.; Taylor, J.G.; Huang, J.; Veleta, K.; Kantak, P.A.; Aita, M.; Shilling-Scrivo, K.; et al. Visualizing Hypothalamic Network Dynamics for Appetitive and Consummatory Behaviors. Cell 2015, 160, 516–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieh, E.H.; Matthews, G.A.; Allsop, S.A.; Presbrey, K.N.; Leppla, C.A.; Wichmann, R.; Neve, R.; Wildes, C.P.; Tye, K.M. Decoding Neural Circuits That Control Compulsive Sucrose Seeking. Cell 2015, 160, 528–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korol, S.V.; Jin, Z.; Babateen, O.; Birnir, B. GLP-1 and Exendin-4 Transiently Enhance GABA A Receptor–Mediated Synaptic and Tonic Currents in Rat Hippocampal CA3 Pyramidal Neurons. Diabetes 2015, 64, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.N.; Pise, A.; Sharma, J.N.; Shukla, P. Glucagon-like Peptide-1 (GLP-1) Receptor Agonist Prevents Development of Tolerance to Anti-Anxiety Effect of Ethanol and Withdrawal-Induced Anxiety in Rats. Metab. Brain Dis. 2015, 30, 719–730. [Google Scholar] [CrossRef]
- Antonsen, K.K.; Klausen, M.K.; Brunchmann, A.S.; le Dous, N.; Jensen, M.E.; Miskowiak, K.W.; Fisher, P.M.; Thomsen, G.K.; Rindom, H.; Fahmy, T.P.; et al. Does Glucagon-like Peptide-1 (GLP-1) Receptor Agonist Stimulation Reduce Alcohol Intake in Patients with Alcohol Dependence: Study Protocol of a Randomised, Double-Blinded, Placebo-Controlled Clinical Trial. BMJ Open 2018, 8, e019562. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, Z.Y.; Nicholson, M.L.; Currie, P.J. 6-Hydroxydopamine Lesions of the Ventral Tegmental Area Suppress Ghrelin’s Ability to Elicit Food-Reinforced Behavior. Neurosci. Lett. 2011, 499, 70–73. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colvin, K.J.; Killen, H.S.; Kanter, M.J.; Halperin, M.C.; Engel, L.; Currie, P.J. Brain Site-Specific Inhibitory Effects of the GLP-1 Analogue Exendin-4 on Alcohol Intake and Operant Responding for Palatable Food. Int. J. Mol. Sci. 2020, 21, 9710. https://doi.org/10.3390/ijms21249710
Colvin KJ, Killen HS, Kanter MJ, Halperin MC, Engel L, Currie PJ. Brain Site-Specific Inhibitory Effects of the GLP-1 Analogue Exendin-4 on Alcohol Intake and Operant Responding for Palatable Food. International Journal of Molecular Sciences. 2020; 21(24):9710. https://doi.org/10.3390/ijms21249710
Chicago/Turabian StyleColvin, Kayla J., Henry S. Killen, Maxwell J. Kanter, Maximilian C. Halperin, Liv Engel, and Paul J. Currie. 2020. "Brain Site-Specific Inhibitory Effects of the GLP-1 Analogue Exendin-4 on Alcohol Intake and Operant Responding for Palatable Food" International Journal of Molecular Sciences 21, no. 24: 9710. https://doi.org/10.3390/ijms21249710