Anti-Inflammatory Activity of Extracts and Pure Compounds Derived from Plants via Modulation of Signaling Pathways, Especially PI3K/AKT in Macrophages


Abstract
:1. Introduction
2. Medicinal Plants with Economic Value
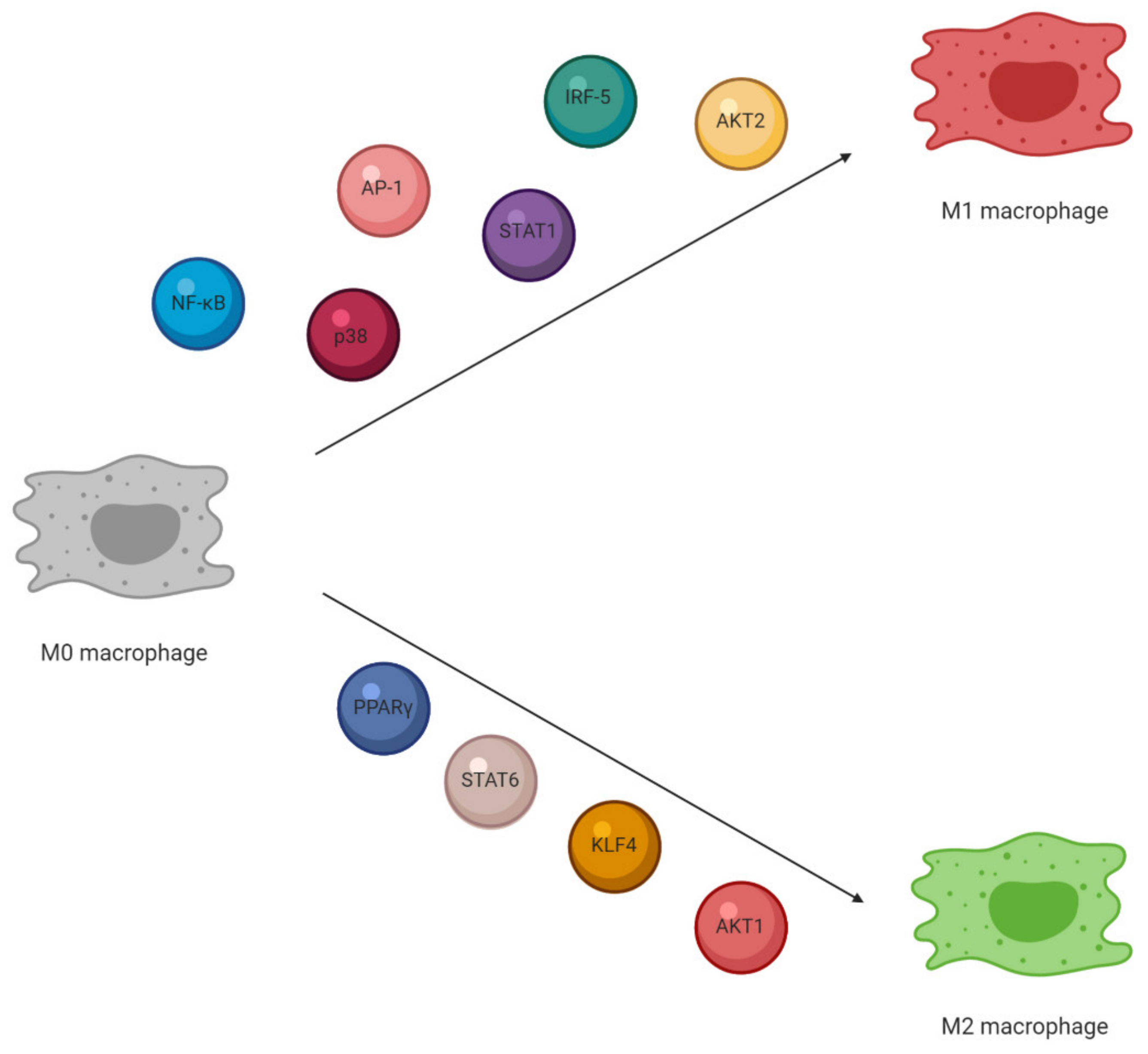
3. Macrophage Characterization and Activation
4. Plants and Their Active Compounds as Modulators of Macrophage Polarization
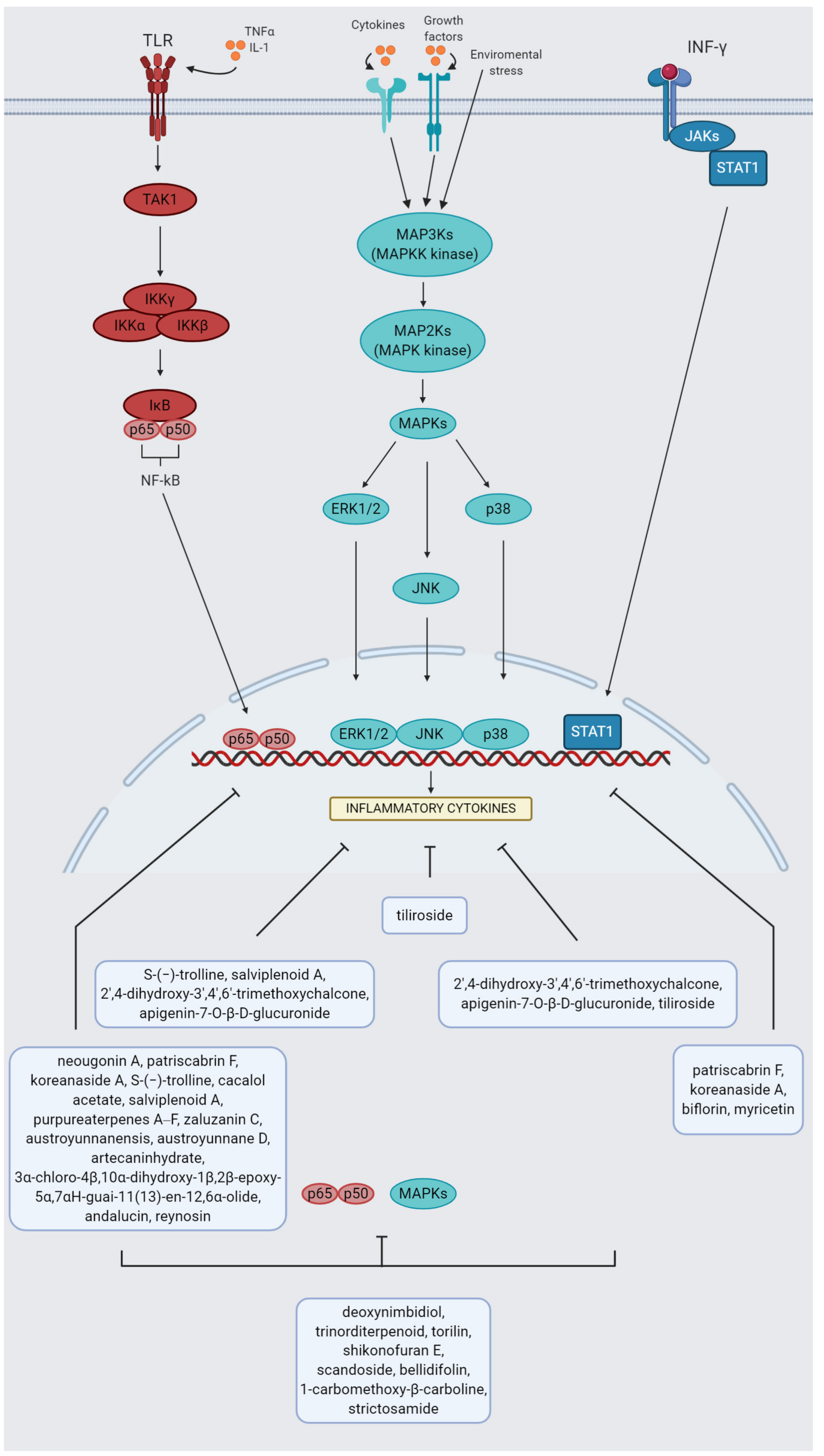
4.1. M1 Macrophages Stimuli and Plant-Based Modulators
4.2. M2 Macrophages Stimuli and Plant-Based Modulators
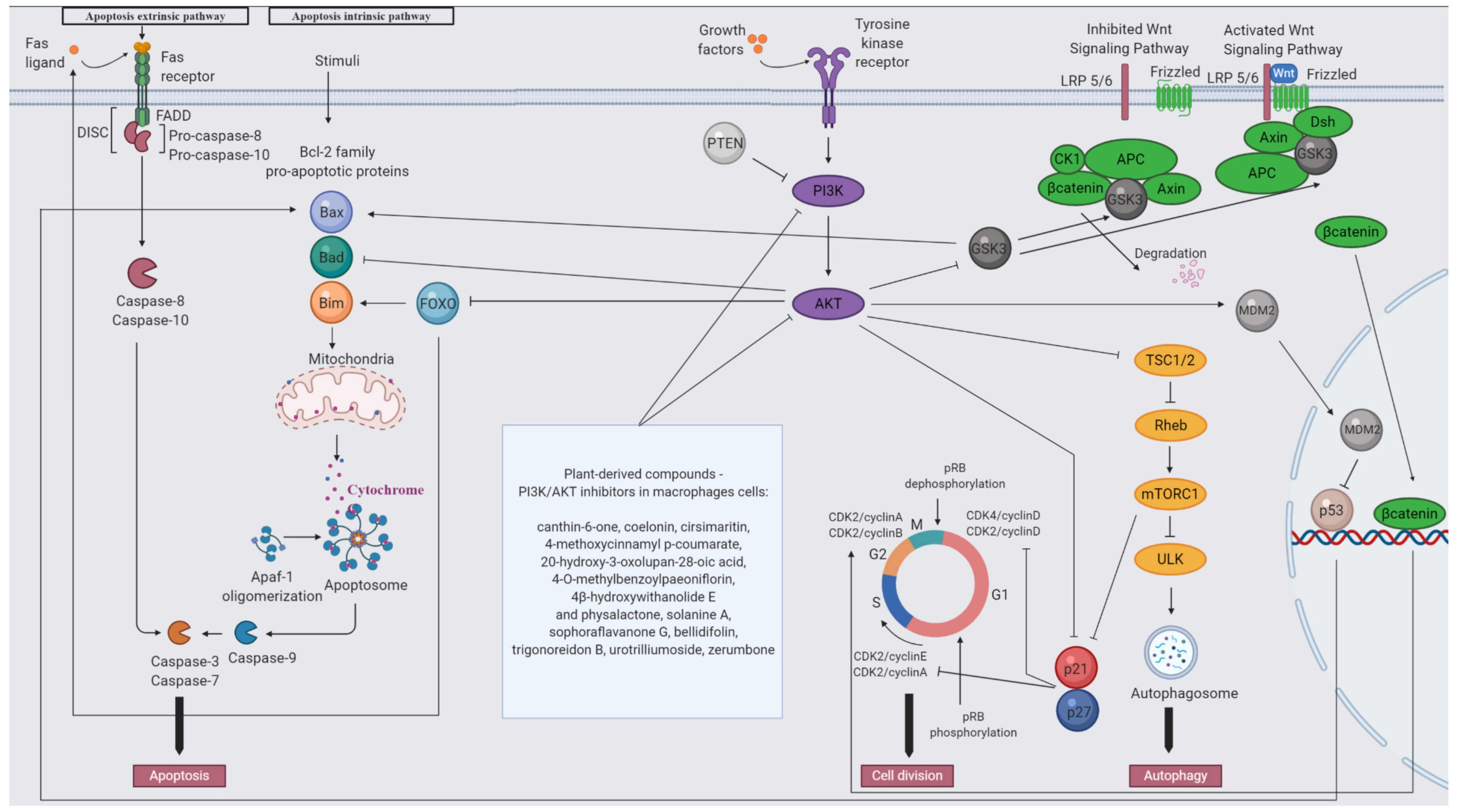
5. The PI3K/AKT Signaling Pathway
5.1. PI3K/AKT Signaling Modulates Macrophage Activation
5.2. PI3K/AKT Pathway Regulate Macrophage Polarization
5.3. The Role of the Upstream Regulators of AKT in Macrophages Polarization—PTEN, SHIP, PDK-1
5.4. The Role of AKT Downstream Signal in Macrophage Polarization—mTOR
5.5. The Contribution of AKT Signaling in Phagocytosis and Autophagy
5.6. The Contribution of Macrophage AKT in Inflammatory Diseases
6. Plants as Natural Modulators of PI3K/AKT Pathway in Macrophages
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kaufman, J.; Kalaitzandonakes, N. The economic potential of plant-made pharmaceuticals in the manufacture of biologic pharmaceuticals. J. Commer. Biotechnol. 2011, 17, 173–182. [Google Scholar] [CrossRef]
- Hefferon, K. Plant-derived pharmaceuticals for the developing world. Biotechnol. J. 2013, 8. [Google Scholar] [CrossRef]
- Pagare, S.; Bhatia, M.; Tripathi, N.; Pagare, S.; Bansal, Y.K. Secondary metabolites of plants and their role: Overview. Curr. Trends Biotechnol. Pharm. 2015, 9, 293–304. [Google Scholar]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Smith, W.; Hao, D.; He, B.; Kong, L. M1 and M2 macrophage polarization and potentially therapeutic naturally occurring compounds. Int. Immunopharmacol. 2019, 70, 459–466. [Google Scholar] [CrossRef]
- Varol, C.; Mildner, A.; Jung, S. Macrophages: Development and tissue specialization. Annu. Rev. Immunol. 2015, 33, 643–675. [Google Scholar] [CrossRef]
- Kumar, V. Macrophages: The Potent Immunoregulatory Innate Immune Cells. In Macrophage Activation—Biology and Disease; IntechOpen Limited: London, UK, 2020. [Google Scholar]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef]
- Liu, Y.C.; Zou, X.B.; Chai, Y.F.; Yao, Y.M. Macrophage polarization in inflammatory diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef]
- Gordon, S.; Martinez-Pomares, L. Physiological roles of macrophages. Pflug. Arch. Eur. J. Physiol. 2017, 469, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Troutman, T.D.; Bazan, J.F.; Pasare, C. Toll-like receptors, signalingadapters and regulation of the pro-inflammatory response by PI3K. Cell Cycle 2012, 11, 3559–3567. [Google Scholar] [CrossRef] [Green Version]
- Covarrubias, A.J.; Aksoylar, H.I.; Horng, T. Control of macrophage metabolism and activation by mTOR and Akt signaling. Semin. Immunol. 2015, 27, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Rauh, M.J.; Ho, V.; Pereira, C.; Sham, A.; Sly, L.M.; Lam, V.; Huxham, L.; Minchinton, A.I.; Mui, A.; Krystal, G. SHIP represses the generation of alternatively activated macrophages. Immunity 2005, 23, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Arranz, A.; Doxaki, C.; Vergadi, E.; De La Torre, Y.M.; Vaporidi, K.; Lagoudaki, E.D.; Ieronymaki, E.; Androulidaki, A.; Venihaki, M.; Margioris, A.N.; et al. Akt1 and Akt2 protein kinases differentially contribute to macrophage polarization. Proc. Natl. Acad. Sci. USA 2012, 109, 9517–9522. [Google Scholar] [CrossRef] [Green Version]
- Mercalli, A.; Calavita, I.; Dugnani, E.; Citro, A.; Cantarelli, E.; Nano, R.; Melzi, R.; Maffi, P.; Secchi, A.; Sordi, V.; et al. Rapamycin unbalances the polarization of human macrophages to M1. Immunology 2013, 140, 179–190. [Google Scholar] [CrossRef]
- Qian, M.; Fang, X.; Wang, X. Autophagy and inflammation. Clin. Transl. Med. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Van Kleunen, M.; Xu, X.; Yang, Q.; Maurel, N.; Zhang, Z.; Dawson, W.; Essl, F.; Kreft, H.; Pergl, J.; Pyšek, P.; et al. Economic use of plants is key to their naturalization success. Nat. Commun. 2020, 11, 3201. [Google Scholar] [CrossRef]
- Firenzuoli, F.; Gori, L. Herbal medicine today: Clinical and research issues. Evid. Based Complement. Alternat. Med. 2007, 4, 37–40. [Google Scholar] [CrossRef]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The traditional medicine and modern medicine from natural products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [Green Version]
- Böttger, A.; Vothknecht, U.; Bolle, C.; Wolf, A. Lessons on Caffeine, Cannabis & Co.; Springer International Publishing: New York, NY, USA, 2018. [Google Scholar]
- Hussein, A.R.; El-Anssary, A. Plants. In Herbal Medicine; IntechOpen Limited: London, UK, 2019. [Google Scholar]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [Green Version]
- Anand, U.; Jacobo-Herrera, N.; Altemimi, A.; Lakhssassi, N. A comprehensive review on medicinal plants as antimicrobial therapeutics: Potential avenues of biocompatible drug discovery. Metabolites 2019, 9, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitarek, P.; Merecz-Sadowska, A.; Kowalczyk, T.; Wieczfinska, J.; Zajdel, R.; Śliwiński, T. Potential synergistic action of bioactive compounds from plant extracts against skin infecting microorganisms. Int. J. Mol. Sci. 2020, 21, 5105. [Google Scholar] [CrossRef] [PubMed]
- Zielinska-Blizniewska, H.; Sitarek, P.; Merecz-Sadowska, A.; Malinowska, K.; Zajdel, K.; Jablonska, M.; Sliwinski, T.; Zajdel, R. Plant extracts and reactive oxygen species as two counteracting agents with anti- and pro-obesity properties. Int. J. Mol. Sci. 2019, 20, 4556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- Ghasemian, M.; Owlia, S.; Owlia, M.B. Review of Anti-Inflammatory Herbal Medicines. Adv. Pharmacol. Sci. 2016, 2016, 9130979. [Google Scholar] [CrossRef] [Green Version]
- Khan, T.; Ali, M.; Khan, A.; Nisar, P.; Jan, S.A.; Afridi, S.; Shinwari, Z.K. Anticancer plants: A review of the active phytochemicals, applications in animal models, and regulatory aspects. Biomolecules 2020, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Khan, R.A. Natural products chemistry: The emerging trends and prospective goals. Saudi Pharm. J. 2018, 26, 739–753. [Google Scholar] [CrossRef]
- Abdin, M.Z.; Alam, P. Genetic engineering of artemisinin biosynthesis: Prospects to improve its production. Acta Physiol. Plant. 2015, 37, 33. [Google Scholar] [CrossRef]
- Jansen, D.J.; Shenvi, R.A. Synthesis of medicinally relevant terpenes: Reducing the cost and time of drug discovery. Future Med. Chem. 2014, 6, 1127–1148. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K. The worldwide trend of using botanical drugs and strategies for developing global drugs. BMB Rep. 2017, 50, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Sofowora, A.; Ogunbodede, E.; Onayade, A. The role and place of medicinal plants in the strategies for disease prevention. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 210–229. [Google Scholar] [CrossRef]
- Tasneem, S.; Liu, B.; Li, B.; Choudhary, M.I.; Wang, W. Molecular pharmacology of inflammation: Medicinal plants as anti-inflammatory agents. Pharmacol. Res. 2019, 139, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Richardson, E.T.; Shukla, S.; Nagy, N.; Boom, W.H.; Beck, R.C.; Zhou, L.; Landreth, G.E.; Harding, C.V. ERK signaling is essential for macrophage development. PLoS ONE 2015, 10, e0140064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The metabolic signature of macrophage responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edholm, E.S.; Rhoo, K.H.; Robert, J. Evolutionary aspects of macrophages polarization. Results Probl. Cell Differ. 2017, 62, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Rath, M.; Müller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via arginase or nitric oxide synthase: Two competing arginine pathways in macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, P.; Tripathi, P.; Kashyap, L.; Singh, V. The role of nitric oxide in inflammatory reactions. FEMS Immunol. Med. Microbiol. 2007, 51, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Jiang, T.; Li, M.Q.; Zheng, X.L.; Zhao, G.J. Transcriptional regulation of macrophages polarization by microRNAs. Front. Immunol. 2018, 9, 1175. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Jantan, I.; Ahmad, W.; Bukhari, S.N.A. Plant-derived immunomodulators: An insight on their preclinical evaluation and clinical trials. Front. Plant Sci. 2015, 6, 655. [Google Scholar] [CrossRef]
- Samec, M.; Liskova, A.; Koklesova, L.; Mathews, S.; Radovan, S.; Pavol, M. The role of plant—Derived natural substances as immunomodulatory agents in carcinogenesis. J. Cancer Res. Clin. Oncol. 2020, 146, 3137–3154. [Google Scholar] [CrossRef] [PubMed]
- Mendes, L.F.; Gaspar, V.M.; Conde, T.A.; Mano, J.F.; Duarte, I.F. Flavonoid-mediated immunomodulation of human macrophages involves key metabolites and metabolic pathways. Sci. Rep. 2019, 9, 14906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.; Song, B.; Cho, K.S.; Lee, I.S. Therapeutic potential of volatile terpenes and terpenoids from forests for inflammatory diseases. Int. J. Mol. Sci. 2020, 21, 2187. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Zhang, Y.; Li, H. Advances in research on immunoregulation of macrophages by plant polysaccharides. Front. Immunol. 2019, 10, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saqib, U.; Sarkar, S.; Suk, K.; Mohammad, O.; Baig, M.S.; Savai, R. Phytochemicals as modulators of M1-M2 macrophages in inflammation. Oncotarget 2018, 9, 17937–17950. [Google Scholar] [CrossRef] [Green Version]
- Davoodvandi, A.; Sahebnasagh, R.; Mardanshah, O.; Asemi, Z.; Nejati, M.; Shahrzad, M.K.; Mirzaei, H.R.; Mirzaei, H. Medicinal Plants As Natural Polarizers of Macrophages: Phytochemicals and Pharmacological Effects. Curr. Pharm. Des. 2019, 25, 3225–3238. [Google Scholar] [CrossRef]
- Dorrington, M.G.; Fraser, I.D.C. NF-κB signaling in macrophages: Dynamics, crosstalk, and signal integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Pomari, E.; Stefanon, B.; Colitti, M. Effect of plant extracts on H2O2-induced inflammatory gene expression in macrophages. J. Inflamm. Res. 2014, 7, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Hiben, M.G.; de Haan, L.; Spenkelink, B.; Wesseling, S.; Vervoort, J.; Rietjens, I.M.C.M. Induction of peroxisome proliferator activated receptor γ (PPARγ) mediated gene expression and inhibition of induced nitric oxide production by Maerua subcordata (Gilg) DeWolf. BMC Complement. Med. Ther. 2020, 20, 80. [Google Scholar] [CrossRef] [Green Version]
- Jo, M.; Yi, Y.S.; Cho, J.Y. Archidendron lucidum Exerts Anti-Inflammatory Effects by Targeting PDK1 in the NF-κB Pathway. Am. J. Chin. Med. 2020, 48, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Sethumadhavan, K. Regulation of Cell Viability and Anti-inflammatory Tristetraprolin Family Gene Expression in Mouse Macrophages by Cottonseed Extracts. Sci. Rep. 2020, 10, 775. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Ji, P.; Song, M.; Che, T.; Bravo, D.; Pettigrew, J.; Liu, Y. Dietary plant extracts modulate gene expression profiles in alveolar macrophages of pigs experimentally infected with porcine reproductive and respiratory syndrome virus. J. Anim. Sci. Biotechnol. 2020, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei, N.; Hou, S.; Kim, K.W.; Tahmasebi, S. Signaling pathways that control mRNA translation initiation in macrophages. Cell. Signal. 2020, 73, 109700. [Google Scholar] [CrossRef]
- Sun, L.R.; Zhou, W.; Zhang, H.M.; Guo, Q.S.; Yang, W.; Li, B.J.; Sun, Z.H.; Gao, S.H.; Cui, R.J. Modulation of Multiple Signaling Pathways of the Plant-Derived Natural Products in Cancer. Front. Oncol. 2019, 9, 1153. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Fofaria, N.M.; Kim, S.H.; Srivastava, S.K. Modulation of signal transduction pathways by natural compounds in cancer. Chin. J. Nat. Med. 2015, 13, 730–742. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. Signaling to NF-κB by Toll-like receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Platanitis, E.; Decker, T. Regulatory networks involving STATs, IRFs, and NFκB in inflammation. Front. Immunol. 2018, 9, 2542. [Google Scholar] [CrossRef] [Green Version]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-κB signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Hagemann, T.; Lawrence, T.; McNeish, I.; Charles, K.A.; Kulbe, H.; Thompson, R.G.; Robinson, S.C.; Balkwill, F.R. “Re-educating” tumor-associated macrophages by targeting NF-κB. J. Exp. Med. 2008, 205, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Fard, M.; Arulselvan, P.; Karthivashan, G.; Adam, S.; Fakurazi, S. Bioactive extract from moringa oleifera inhibits the pro-inflammatory mediators in lipopolysaccharide stimulated macrophages. Pharmacogn. Mag. 2015, 1, S556–S563. [Google Scholar] [CrossRef]
- Qomaladewi, N.P.; Aziz, N.; Kim, M.Y.; Cho, J.Y. Piper cubeba L. Methanol Extract Has Anti-Inflammatory Activity Targeting Src/Syk via NF-κB Inhibition. Evid. Based Complement. Altern. Med. 2019, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Araújo Moreira, M.D.R.; Sales-Campos, H.; Fontanari, C.; Galvão Meireles, A.F.; Borges Prado, M.K.; Zoccal, K.F.; Sorgi, C.A.; Tefé da Silva, C.; Groppo, M.; Faccioli, L.H. The ethanolic extract of Terminalia argentea Mart. & Zucc. bark reduces the inflammation through the modulation of cytokines and nitric oxide mediated by the downregulation of NF-κB. J. Ethnopharmacol. 2020, 261, 113150. [Google Scholar] [CrossRef]
- Harbeoui, H.; Hichami, A.; Wannes, W.A.; Lemput, J.; Tounsi, M.S.; Khan, N.A. Anti-inflammatory effect of grape (Vitis vinifera L.) seed extract through the downregulation of NF-κB and MAPK pathways in LPS-induced RAW264.7 macrophages. S. Afr. J. Bot. 2019, 125, 1–8. [Google Scholar] [CrossRef]
- Gutierrez, R.M.P.; Hoyo-Vadillo, C. Anti-inflammatory potential of Petiveria alliacea on activated RAW264.7 murine macrophages. Pharmacogn. Mag. 2017, 13, S174–S178. [Google Scholar] [CrossRef] [Green Version]
- Adam, G.O.; Kim, G.B.; Lee, S.J.; Lee, H.; Kang, H.S.; Kim, S.J. Red Ginseng Reduces Inflammatory Response via Suppression MAPK/P38 Signaling and p65 Nuclear Proteins Translocation in Rats and Raw 264.7 Macrophage. Am. J. Chin. Med. 2019, 47, 1589–1609. [Google Scholar] [CrossRef]
- Park, J.Y.; Park, S.D.; Koh, Y.J.; Kim, D.I.; Lee, J.H. Aqueous extract of Dipsacus asperoides suppresses lipopolysaccharide-stimulated inflammatory responses by inhibiting the ERK1/2 signaling pathway in RAW 264.7 macrophages. J. Ethnopharmacol. 2019, 231, 253–261. [Google Scholar] [CrossRef]
- Caban, M.; Chojnacka, K.; Owczarek, K.; Laskowska, J.; Fichna, J.; Podsedek, A.; Sosnowska, D.; Lewandowska, U. Spent hops (Humulus lupulus L.) extract as modulator of the inflammatory response in lipopolysaccharide stimulated raw 264.7 macrophages. J. Physiol. Pharmacol. 2020, 71. [Google Scholar] [CrossRef]
- Gil, T.Y.; Jin, B.R.; Hong, C.H.; Park, J.H.; An, H.J. Astilbe Chinensis ethanol extract suppresses inflammation in macrophages via NF-κB pathway. BMC Complement. Med. Ther. 2020, 20, 302. [Google Scholar] [CrossRef]
- Nam, N.-H. Naturally occurring NF-kappaB inhibitors. Mini Rev. Med. Chem. 2006, 6, 945–951. [Google Scholar] [CrossRef]
- Shin, J.S.; Kang, S.Y.; Lee, H.H.; Kim, S.Y.; Lee, D.H.; Jang, D.S.; Lee, K.T. Patriscabrin F from the roots of Patrinia scabra attenuates LPS-induced inflammation by downregulating NF-κB, AP-1, IRF3, and STAT1/3 activation in RAW 264.7 macrophages. Phytomedicine 2020, 68, 153167. [Google Scholar] [CrossRef]
- Kim, T.W.; Shin, J.S.; Chung, K.S.; Lee, Y.G.; Baek, N.I.; Lee, K.T. Anti-Inflammatory Mechanisms of Koreanaside A, a Lignan Isolated from the Flower of Forsythia koreana, against LPS-Induced Macrophage Activation and DSS-Induced Colitis Mice: The Crucial Role of AP-1, NF-κB, and JAK/STAT Signaling. Cells 2019, 8, 1163. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Jang, H.J.; Han, Y.K.; Su, X.D.; Lee, S.W.; Rho, M.C.; Wang, H.S.; Yang, S.Y.; Kim, Y.H. Alkaloids from Tetrastigma hemsleyanum and their anti-inflammatory effects on LPS-induced RAW264.7 cells. Molecules 2018, 23, 1445. [Google Scholar] [CrossRef] [Green Version]
- Mora-Ramiro, B.; Jiménez-Estrada, M.; Zentella-Dehesa, A.; Ventura-Gallegos, J.L.; Gomez-Quiroz, L.E.; Rosiles-Alanis, W.; Alarcón-Aguilar, F.J.; Almanza-Pérez, J.C. Cacalol Acetate, a Sesquiterpene from Psacalium decompositum, Exerts an Anti-inflammatory Effect through LPS/NF-KB Signaling in Raw 264.7 Macrophages. J. Nat. Prod. 2020, 8, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.H.; Zhao, L.; Xu, Y.K.; Bao, J.M.; Liu, X.; Zhang, J.S.; Li, W.; Ahmed, A.; Yin, S.; Tang, G.H. Anti-inflammatory sesquiterpenoids from the Traditional Chinese Medicine Salvia plebeia: Regulates pro-inflammatory mediators through inhibition of NF-κB and Erk1/2 signaling pathways in LPS-induced Raw264.7 cells. J. Ethnopharmacol. 2018, 210, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.Y.; Sun, X.; Liu, P.; Lin, B.; Li, L.Z.; Yao, G.D.; Huang, X.X.; Song, S.J. Sesquiterpenes from Echinacea purpurea and their anti-inflammatory activities. Phytochemistry 2020, 179, 112503. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.R.; Lee, H.W.; Kim, Y.H. Anti-inflammatory effect of zaluzanin C on lipopolysaccharide-stimulated murine macrophages. Korean J. Food Sci. Technol. 2016, 48, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Liu, D.; Xiang, C.; Dai, W.; Li, B.; Zhang, M. Sesquiterpene lactones from Artemisia austroyunnanensis suppresses ROS production and reduces cytokines, iNOS and COX-2 levels via NF-KB pathway in vitro. Nat. Prod. Res. 2020, 34, 1563–1566. [Google Scholar] [CrossRef]
- Kondoh, K.; Nishida, E. Regulation of MAP kinases by MAP kinase phosphatases. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 1227–1237. [Google Scholar] [CrossRef] [Green Version]
- Dhar, R.; Kimseng, R.; Chokchaisiri, R.; Hiransai, P.; Utaipan, T.; Suksamrarn, A.; Chunglok, W. 2′,4-Dihydroxy-3′,4′,6′-trimethoxychalcone from Chromolaena odorata possesses anti-inflammatory effects via inhibition of NF-κB and p38 MAPK in lipopolysaccharide-activated RAW 264.7 macrophages. Immunopharmacol. Immunotoxicol. 2018, 40, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, X.; Wu, L.; Shen, T.; Ji, L.; Zhao, X.; Si, C.L.; Jiang, Y.; Wang, G. Apigenin-7-O-β-d-glucuronide inhibits LPS-induced inflammation through the inactivation of AP-1 and MAPK signaling pathways in RAW 264.7 macrophages and protects mice against endotoxin shock. Food Funct. 2016, 7, 1002–1013. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Song, S.; Wang, J.; Zhang, Q.; Qiu, F.; Zhao, F. Tiliroside, the major component of Agrimonia pilosa Ledeb ethanol extract, inhibits MAPK/JNK/p38-mediated inflammation in lipopolysaccharide-activated RAW 264.7 macrophages. Exp. Ther. Med. 2016, 12, 499–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarubin, T.; Han, J. Activation and signaling of the p38 MAP kinase pathway. Cell Res. 2005, 15, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Kim, S.C.; Yu, T.; Yi, Y.S.; Rhee, M.H.; Sung, G.H.; Yoo, B.C.; Cho, J.Y. Functional roles of p38 mitogen-activated protein kinase in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2014, 2014, 352371. [Google Scholar] [CrossRef] [Green Version]
- Neamatallah, T. Mitogen-activated protein kinase pathway: A critical regulator in tumor-associated macrophage polarization. J. Microsc. Ultrastruct. 2019, 7, 53–56. [Google Scholar] [CrossRef]
- Lee, Y.M.; Son, E.; Kim, D.S. Treatment with peanut sprout root extract alleviates inflammation in a lipopolysaccharide-stimulated mouse macrophage cell line by inhibiting the MAPK signaling pathway. Int. J. Mol. Sci. 2019, 20, 5907. [Google Scholar] [CrossRef] [Green Version]
- Whitmarsh, A.J.; Davis, R.J. Transcription factor AP-1 regulation by mitogen-activated protein kinase signal transduction pathways. J. Mol. Med. 1996, 74, 589–607. [Google Scholar] [CrossRef]
- Srivastava, M.; Baig, M.S. NOS1 mediates AP1 nuclear translocation and inflammatory response. Biomed. Pharmacother. 2018, 102, 839–847. [Google Scholar] [CrossRef]
- Juhas, U.; Ryba-Stanisławowska, M.; Szargiej, P.; Myśliwska, J. Different pathways of macrophage activation and polarization. Postepy Hig. Med. Dosw. 2015, 69, 496–502. [Google Scholar] [CrossRef]
- Hossen, M.J.; Kim, S.C.; Son, Y.J.; Baek, K.S.; Kim, E.; Yang, W.S.; Jeong, D.; Park, J.G.; Kim, H.G.; Chung, W.J.; et al. AP-1-targeting anti-inflammatory activity of the methanolic extract of Persicaria Chinensis. Evid. Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.H.; Oh, Y.C.; Cho, W.K.; Yim, N.H.; Ma, J.Y. Hoveniae semen seu fructus ethanol extract exhibits anti-inflammatory activity via MAPK, AP-1, and STAT signaling pathways in LPS-stimulated RAW 264.7 and mouse peritoneal macrophages. Mediat. Inflamm. 2019, 2019, 9184769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endale, M.; Kim, T.H.; Kwak, Y.S.; Kim, N.M.; Kim, S.H.; Cho, J.Y.; Yun, B.S.; Rhee, M.H. Torilin Inhibits Inflammation by Limiting TAK1-Mediated MAP Kinase and NF-κB Activation. Mediat. Inflamm. 2017, 2017, 7250968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.W.; Kwon, O.K.; Ryu, H.W.; Paik, J.H.; Paryanto, I.; Yuniato, P.; Choi, S.H.; Oh, S.R.; Ahn, K.S. Anti-inflammatory effects of Passiflora foetida L. In LPS-stimulated RAW264.7 macrophages. Int. J. Mol. Med. 2018, 41, 3709–3716. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.N.; Kim, J.D.; Yeo, J.H.; Son, H.J.; Park, S.B.; Park, G.H.; Eo, H.J.; Jeong, J.B. Heracleum moellendorffii roots inhibit the production of pro-inflammatory mediators through the inhibition of NF-κB and MAPK signaling, and activation of ROS/Nrf2/HO-1 signaling in LPS-stimulated RAW264.7 cells. BMC Complement. Altern. Med. 2019, 19, 310. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.N.; Baek, J.K.; Park, S.B.; Kim, J.D.; Son, H.J.; Park, G.H.; Eo, H.J.; Park, J.H.; Jung, H.S.; Jeong, J.B. Anti-inflammatory effect of Vaccinium oldhamii stems through inhibition of NF-κB and MAPK/ATF2 signaling activation in LPS-stimulated RAW264.7 cells. BMC Complement. Altern. Med. 2019, 19, 291. [Google Scholar] [CrossRef]
- Choo, G.S.; Lim, D.P.; Kim, S.M.; Yoo, E.S.; Kim, S.H.; Kim, C.H.; Woo, J.S.; Kim, H.J.; Jung, J.Y. Anti-inflammatory effects of Dendropanax morbifera in lipopolysaccharide-stimulated RAW264.7 macrophages and in an animal model of atopic dermatitis. Mol. Med. Rep. 2019, 19, 2087–2096. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.A.; Jantan, I.; Harikrishnan, H.; Ghazalee, S. Standardized extract of Zingiber zerumbet suppresses LPS-induced pro-inflammatory responses through NF-κB, MAPK and PI3K-Akt signaling pathways in U937 macrophages. Phytomedicine 2019, 54, 195–205. [Google Scholar] [CrossRef]
- Chu, Q.; Yu, X.; Jia, R.; Wang, Y.; Zhang, Y.; Zhang, S.; Liu, Y.; Li, Y.; Chen, W.; Ye, X.; et al. Flavonoids from Apios americana Medikus Leaves Protect RAW264.7 Cells against Inflammation via Inhibition of MAPKs, Akt-mTOR Pathways, and Nfr2 Activation. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Tursun, X.; Zhao, Y.; Alat, Z.; Xin, X.; Tursun, A.; Abdulla, R.; Akber Aisa, H. Anti- inflammatory effect of Rosa rugosa flower extract in lipopolysaccharide-stimulated RAW264.7 macrophages. Biomol. Ther. 2016, 24, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Harikrishnan, H.; Jantan, I.; Haque, M.A.; Kumolosasi, E. Anti-inflammatory effects of Phyllanthus amarus Schum. & Thonn. Through inhibition of NF-ΚB, MAPK, and PI3K-Akt signaling pathways in LPS-induced human macrophages. BMC Complement. Altern. Med. 2018, 18, 224. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, Y.; Aisa, H.A. Anti-inflammatory effect of pomegranate flower in lipopolysaccharide (Lps)-stimulated raw264.7 macrophages. Pharm. Biol. 2017, 55, 2095–2101. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.; Park, J.W.; Kwon, O.K.; Lee, J.W.; Lee, H.S.; Lee, S.; Choi, S.; Li, W.; Jin, H.; Han, S.B.; et al. Anti-inflammatory effects of a methanolic extract of Castanea seguinii Dode in LPS-induced RAW264.7 macrophage cells. Int. J. Mol. Med. 2018, 41, 391–398. [Google Scholar] [CrossRef]
- Jeong, Y.H.; Oh, Y.C.; Cho, W.K.; Lee, B.; Ma, J.Y. Anti-Inflammatory Effects of Melandrii Herba Ethanol Extract via Inhibition of NF-κB and MAPK Signaling Pathways and Induction of HO-1 in RAW 264.7 Cells and Mouse Primary Macrophages. Molecules 2016, 21, 818. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.C.; Bach, T.T.; Kim, B.R.; Vuong, H.L.; Cho, S. Spilanthes acmella inhibits inflammatory responses via inhibition of NF-κB and MAPK signaling pathways in RAW 264.7 macrophages. Mol. Med. Rep. 2017, 16, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngabire, D.; Seong, Y.A.; Patil, M.P.; Niyonizigiye, I.; Seo, Y.B.; Kim, G. Do Anti-inflammatory effects of aster incisus through the inhibition of NF-κB, MAPK, and akt pathways in LPS-stimulated RAW 264.7 macrophages. Mediat. Inflamm. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H.; Kang, H.J. Fructus sophorae attenuates secretion of proinflammatory mediators and cytokines through the modulation of NF-κB and MAPK signaling pathways in LPS-stimulated RAW 264.7 macrophages. Gen. Physiol. Biophys. 2016, 35, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Eo, H.J.; Kwon, H.Y.; Da Kim, S.; Kang, Y.; Park, Y.; Park, G.H. GC/MS analysis and anti-inflammatory effect of leaf extracts from Hibiscus syriacus through inhibition of NF-κB and MAPKs signaling in LPS-stimulated RAW264.7 macrophages. Plant Biotechnol. Rep. 2020, 14, 539–546. [Google Scholar] [CrossRef]
- Kim, H.N.; Kim, J.D.; Park, S.B.; Son, H.J.; Park, G.H.; Eo, H.J.; Kim, H.S.; Jeong, J.B. Anti-inflammatory activity of the extracts from Rodgersia podophylla leaves through activation of Nrf2/HO-1 pathway, and inhibition of NF-κB and MAPKs pathway in mouse macrophage cells. Inflamm. Res. 2020, 69, 233–244. [Google Scholar] [CrossRef]
- Jeong, Y.E.; Lee, M.Y. Anti-inflammatory activity of populus deltoides leaf extract via modulating NF-κB and p38/JNK pathways. Int. J. Mol. Sci. 2018, 19, 3746. [Google Scholar] [CrossRef] [Green Version]
- Yeom, M.; Kim, J.H.; Min, J.H.; Hwang, M.K.; Jung, H.S.; Sohn, Y. Xanthii fructus inhibits inflammatory responses in LPS-stimulated RAW 264.7 macrophages through suppressing NF-κB and JNK/p38 MAPK. J. Ethnopharmacol. 2015, 176, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Kim, K.H.; Park, E.J.; Kang, J.A.; Yun, B.S.; Lee, S.J.; Park, C.S.; Lee, S.; Lee, S.W.; Rho, M.C. Anti-Inflammatory Activity of Diterpenoids from Celastrus orbiculatus in Lipopolysaccharide-Stimulated RAW264.7 Cells. J. Immunol. Res. 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Xue, Y.; Yang, Z.; Li, Y.; Li, H.; Chen, X.; Li, R.; Liu, D. Shikonofuran E plays an anti-inflammatory role by down-regulating MAPK and NF-κB signaling pathways in lipopolysaccharide-stimulated RAW264.7 macrophages. J. Nat. Med. 2019, 73, 244–251. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Li, J.; Liu, H.; Yang, Z.; Zhou, F.; Wei, T.; Dong, Y.; Xue, H.; Tang, L.; Liu, M. Scandoside exerts anti-inflammatory effect via suppressing NF-κb and MAPK signaling pathways in lps-induced raw 264.7 macrophages. Int. J. Mol. Sci. 2018, 19, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.Y.; Ju, J.M.; Mo, L.H.; Ma, L.; Hu, W.H.; You, R.R.; Chen, X.Q.; Chen, Y.Y.; Liu, Z.Q.; Qiu, S.Q.; et al. Anti-inflammation action of xanthones from Swertia chirayita by regulating COX-2/NF-κB/MAPKs/Akt signaling pathways in RAW 264.7 macrophage cells. Phytomedicine 2019, 55, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Park, E.J.; Jang, H.J.; Lee, S.J.; Park, C.S.; Yun, B.S.; Lee, S.W.; Rho, M.C. 1-carbomethoxy-β-carboline, derived from portulaca oleracea l., ameliorates lps-mediated inflammatory response associated with mapk signaling and nuclear translocation of NF-κB. Molecules 2019, 24, 4042. [Google Scholar] [CrossRef] [Green Version]
- Li, D.Y.; Chen, J.Q.; Ye, J.Q.; Zhai, X.T.; Song, J.; Jiang, C.H.; Wang, J.; Zhang, H.; Jia, X.B.; Zhu, F.X. Anti-inflammatory effect of the six compounds isolated from Nauclea officinalis Pierrc ex Pitard, and molecular mechanism of strictosamide via suppressing the NF-κB and MAPK signaling pathway in LPS-induced RAW 264.7 macrophages. J. Ethnopharmacol. 2017, 196, 66–74. [Google Scholar] [CrossRef]
- Yan, Z.; Gibson, S.A.; Buckley, J.A.; Qin, H.; Benveniste, E.N. Role of the JAK/STAT signaling pathway in regulation of innate immunity in neuroinflammatory diseases. Clin. Immunol. 2018, 189, 4–13. [Google Scholar] [CrossRef]
- Arora, S.; Dev, K.; Agarwal, B.; Das, P.; Syed, M.A. Macrophages: Their role, activation and polarization in pulmonary diseases. Immunobiology 2018, 223, 383–396. [Google Scholar] [CrossRef]
- Eom, S.H.; Jin, S.J.; Jeong, H.Y.; Song, Y.; Lim, Y.J.; Kim, J.I.; Lee, Y.H.; Kang, H. Kudzu leaf extract suppresses the production of inducible nitric oxide synthase, cyclooxygenase-2, tumor necrosis factor-alpha, and interleukin-6 via inhibition of JNK, TBK1 and STAT1 in inflammatory macrophages. Int. J. Mol. Sci. 2018, 19, 1536. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.K.; Kim, Y.S.; Natarajan, S.B.; Kim, W.S.; Hwang, J.W.; Jeon, N.J.; Jeong, J.H.; Moon, S.H.; Jeon, B.T.; Park, P.J. Antioxidant and Anti-Inflammatory Effects of Chaenomeles sinensis Leaf Extracts on LPS-Stimulated RAW 264.7 Cells. Molecules 2016, 21, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.H.; Shin, J.S.; Lee, W.S.; Ryu, B.; Jang, D.S.; Lee, K.T. Biflorin, Isolated from the Flower Buds of Syzygium aromaticum L., Suppresses LPS-Induced Inflammatory Mediators via STAT1 Inactivation in Macrophages and Protects Mice from Endotoxin Shock. J. Nat. Prod. 2016, 79, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.O.; Yin, H.H.; Park, S.H.; Byun, E.B.; Ha, H.Y.; Jang, S.I. Anti-inflammatory activity of myricetin from diospyros lotus through suppression of nf-κb and stat1 activation and nrf2-mediated ho-1 induction in lipopolysaccharide-stimulated raw264.7 macrophages. Biosci. Biotechnol. Biochem. 2016, 80, 1520–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, K.; Takaoka, A.; Taniguchi, T. Type I Inteferon Gene Induction by the Interferon Regulatory Factor Family of Transcription Factors. Immunity 2006, 25, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chistiakov, D.A.; Myasoedova, V.A.; Revin, V.V.; Orekhov, A.N.; Bobryshev, Y.V. The impact of interferon-regulatory factors to macrophage differentiation and polarization into M1 and M2. Immunobiology 2018, 223, 101–111. [Google Scholar] [CrossRef]
- Wei, Z.; Yan, L.; Chen, Y.; Bao, C.; Dang, J.; Deng, J. Mangiferin inhibits macrophage classical activation via downregulating interferon regulatory factor 5 expression. Mol. Med. Rep. 2016, 14, 1091–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Jin, Z.; Yu, J.; Liang, J.; Yang, Q.; Li, F.; Shi, X.; Zhu, X.; Zhang, X. Baicalin ameliorates experimental inflammatory bowel disease through polarization of macrophages to an M2 phenotype. Int. Immunopharmacol. 2016, 35, 119–126. [Google Scholar] [CrossRef]
- Jiménez-Garcia, L.; Herránz, S.; Luque, A.; Hortelano, S. Critical role of p38 MAPK in IL-4-induced alternative activation of peritoneal macrophages. Eur. J. Immunol. 2015, 45. [Google Scholar] [CrossRef]
- Czimmerer, Z.; Daniel, B.; Horvath, A.; Rückerl, D.; Nagy, G.; Kiss, M.; Peloquin, M.; Budai, M.M.; Cuaranta-Monroy, I.; Simandi, Z.; et al. The Transcription Factor STAT6 Mediates Direct Repression of Inflammatory Enhancers and Limits Activation of Alternatively Polarized Macrophages. Immunity 2018, 48, 75–90.e6. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Hong, J.H.; Lee, J.E.; Lee, Y.C. 18β-Glycyrrhetinic acid, the major bioactive component of Glycyrrhizae Radix, attenuates airway inflammation by modulating Th2 cytokines, GATA-3, STAT6, and Foxp3 transcription factors in an asthmatic mouse model. Environ. Toxicol. Pharmacol. 2017, 52, 99–113. [Google Scholar] [CrossRef]
- Tyagi, S.; Gupta, P.; Saini, A.; Kaushal, C.; Sharma, S. The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.W.; Kang, B.Y.; Kim, S.H.; Pak, Y.K.; Cho, D.; Trinchieri, G.; Kim, T.S. Oxidized low density lipoprotein inhibits interleukin-12 production in lipopolysaccharide-activated mouse macrophages via direct interactions between peroxisome proliferator-activated receptor-gamma and nuclear factor-kappa B. J. Biol. Chem. 2000, 275, 32681–32687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korbecki, J.; Bobiński, R.; Dutka, M. Self-regulation of the inflammatory response by peroxisome proliferator-activated receptors. Inflamm. Res. 2019, 68, 443–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelombitko, M.A. Role of Reactive Oxygen Species in Inflammation: A Minireview. Mosc. Univ. Biol. Sci. Bull. 2018, 73, 199–202. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, Y.; Zhang, P.; Ruan, W.; Zhang, L.; Yuan, S.; Pang, T.; Jia, A.Q. Smiglaside A ameliorates LPS-induced acute lung injury by modulating macrophage polarization via AMPK-PPARγ pathway. Biochem. Pharmacol. 2018, 156, 385–395. [Google Scholar] [CrossRef]
- Wen, Q.; Mei, L.; Ye, S.; Liu, X.; Xu, Q.; Miao, J.; Du, S.; Chen, D.; Li, C.; Li, H. Chrysophanol demonstrates anti-inflammatory properties in LPS-primed RAW 264.7 macrophages through activating PPAR-γ. Int. Immunopharmacol. 2018, 56, 90–97. [Google Scholar] [CrossRef]
- McConnell, B.B.; Yang, V.W. Mammalian Krüppel-Like factors in health and diseases. Physiol. Rev. 2010, 90, 1337–1381. [Google Scholar] [CrossRef]
- Sweet, D.R.; Fan, L.; Hsieh, P.N.; Jain, M.K. Krüppel-Like Factors in Vascular Inflammation: Mechanistic Insights and Therapeutic Potential. Front. Cardiovasc. Med. 2018, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Liao, X.; Sharma, N.; Kapadia, F.; Zhou, G.; Lu, Y.; Hong, H.; Paruchuri, K.; Mahabeleshwar, G.H.; Dalmas, E.; Venteclef, N.; et al. Krüppel-like factor 4 regulates macrophage polarization. J. Clin. Investig. 2011, 121, 2736–2749. [Google Scholar] [CrossRef] [Green Version]
- Mao, Q.; Quan, T.; Luo, B.; Guo, X.; Liu, L.; Zheng, Q. MiR-375 targets KLF4 and impacts the proliferation of colorectal carcinoma. Tumor Biol. 2016, 37, 463–471. [Google Scholar] [CrossRef]
- Chen, W.; Li, X.; Guo, S.; Song, N.; Wang, J.; Jia, L.; Zhu, A. Tanshinone IIA harmonizes the crosstalk of autophagy and polarization in macrophages via miR-375/KLF4 pathway to attenuate atherosclerosis. Int. Immunopharmacol. 2019, 70, 486–497. [Google Scholar] [CrossRef]
- Sharif, O.; Brunner, J.S.; Vogel, A.; Schabbauer, G. Macrophage rewiring by nutrient associated PI3K dependent pathways. Front. Immunol. 2019, 10, 2002. [Google Scholar] [CrossRef]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Backer, J.M. The regulation of class IA PI 3-kinases by inter-subunit interactions. Curr. Top. Microbiol. Immunol. 2010, 346, 87–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carracedo, A.; Pandolfi, P.P. The PTEN-PI3K pathway: Of feedbacks and cross-talks. Oncogene 2008, 27, 5527–5541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prestwich, G.D. Phosphoinositide signaling: From affinity probes to pharmaceutical targets. Chem. Biol. 2004, 11, 619–637. [Google Scholar] [CrossRef]
- Madhunapantula, S.R.V.; Mosca, P.J.; Robertson, G.P. The Akt signaling pathway: An emerging therapeutic target in malignant melanoma. Cancer Biol. Ther. 2011, 12, 1032–1049. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Cantley, L.C. AKT/PKB Signaling: Navigating Downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [Green Version]
- Manning, B.D.; Toker, A. AKT/PKB Signaling: Navigating the Network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Tang, N.; Hadden, T.J.; Rishi, A.K. Akt, FoxO and regulation of apoptosis. Biochim. Biophys. Acta 2011, 1813, 1978–1986. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Tindall, D.J. FOXOs, cancer and regulation of apoptosis. Oncogene 2008, 27, 2312–2319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, U.; Preiss, F.; Brauns-Schubert, P.; Schlicher, L.; Charvet, C. GSK-3 -at the crossroads of cell death and survival. J. Cell Sci. 2014, 127, 1369–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nag, S.; Qin, J.; Srivenugopal, K.S.; Wang, M.; Zhang, R. The MDM2-p53 pathway revisited. J. Biomed. Res. 2013, 27, 254–271. [Google Scholar] [CrossRef] [PubMed]
- Tee, A.R. The target of rapamycin and mechanisms of cell growth. Int. J. Mol. Sci. 2018, 19, 880. [Google Scholar] [CrossRef] [Green Version]
- Lv, D.; Guo, L.; Zhang, T.; Huang, L. PRAS40 signaling in tumor. Oncotarget 2017, 8, 69076–69085. [Google Scholar] [CrossRef] [Green Version]
- Alers, S.; Loffler, A.S.; Wesselborg, S.; Stork, B. Role of AMPK-mTOR-Ulk1/2 in the Regulation of Autophagy: Cross Talk, Shortcuts, and Feedbacks. Mol. Cell. Biol. 2012, 32, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Abukhdeir, A.M.; Park, B.H. p21 and p27. Expert Rev. Mol. Med. 2008, 10, e19. [Google Scholar] [CrossRef] [Green Version]
- Komiya, Y.; Habas, R. Wnt signal transduction pathways. Organogenesis 2008, 4, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Pan, W. GSK3: A multifaceted kinase in Wnt signaling. Trends Biochem. Sci. 2010, 35, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Shtutman, M.; Zhurinsky, J.; Simcha, I.; Albanese, C.; D’Amico, M.; Pestell, R.; Ben-Ze’ev, A. The cyclin D1 gene is a target of the β-catenin/LEF-1 pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 5522–5527. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.S.; Mazur, T.; Ji, W.; Liu, S.T.; Taylor, W.R. Analysis of the role of GSK3 in the mitotic checkpoint. Sci. Rep. 2018, 8, 14259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proud, C.G. mTORC1 and cell cycle control. Enzymes 2010, 27, 129–146. [Google Scholar] [CrossRef]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Xie, L.; Liu, C.; Zhang, Q.; Sun, S. PTEN/PI3k/AKT Regulates Macrophage Polarization in Emphysematous mice. Scand. J. Immunol. 2017, 85, 395–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Guerra, M.J.; Castrillo, A.; Martín-Sanz, P.; Boscá, L. Negative regulation by phosphatidylinositol 3-kinase of inducible nitric oxide synthase expression in macrophages. J. Immunol. 1999, 162, 6184–6190. [Google Scholar] [PubMed]
- Joshi, S.; Liu, K.X.; Zulcic, M.; Singh, A.R.; Skola, D.; Glass, C.K.; Dominick Sanders, P.; Sharabi, A.B.; Pham, T.V.; Tamayo, P.; et al. Macrophage Syk–PI3Kg inhibits antitumor immunity: SRX3207, a novel dual Syk–PI3K inhibitory chemotype relieves tumor immunosuppression. Mol. Cancer Ther. 2020, 19, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Solinas, G.; Becattini, B. The role of PI3Kγ in metabolism and macrophage activation. Oncotarget 2017, 8, 106145–106146. [Google Scholar] [CrossRef]
- Sakai, K.; Suzuki, H.; Oda, H.; Akaike, T.; Azuma, Y.; Murakami, T.; Sugi, K.; Ito, T.; Ichinose, H.; Koyasu, S.; et al. Phosphoinositide 3-kinase in nitric oxide synthesis in macrophage: Critical dimerization of inducible nitric-oxide synthase. J. Biol. Chem. 2006, 281, 17736–17742. [Google Scholar] [CrossRef] [Green Version]
- Fukao, T.; Koyasu, S. PI3K and negative regulation of TLR signaling. Trends Immunol. 2003, 24, 358–363. [Google Scholar] [CrossRef]
- Kuijl, C.; Savage, N.D.L.; Marsman, M.; Tuin, A.W.; Janssen, L.; Egan, D.A.; Ketema, M.; Van Den Nieuwendijk, R.; Van Den Eeden, S.J.F.; Geluk, A.; et al. Intracellular bacterial growth is controlled by a kinase network around PKB/AKT1. Nature 2007, 450, 725–730. [Google Scholar] [CrossRef]
- Babaev, V.R.; Hebron, K.E.; Wiese, C.B.; Toth, C.L.; Ding, L.; Zhang, Y.; May, J.M.; Fazio, S.; Vickers, K.C.; Linton, M.F. Macrophage deficiency of Akt2 reduces atherosclerosis in Ldlr null mice. J. Lipid Res. 2014, 55, 2296–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Androulidaki, A.; Iliopoulos, D.; Arranz, A.; Doxaki, C.; Zacharioudaki, V.; Margioris, A.N.; Tsichlis, P.N.; Tsatsanis, C. Akt1 controls macrophage response to LPS by regulating microRNAs. Immunity 2009, 31, 220–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawano, Y.; Nakae, J.; Watanabe, N.; Fujisaka, S.; Iskandar, K.; Sekioka, R.; Hayashi, Y.; Tobe, K.; Kasuga, M.; Noda, T.; et al. Loss of Pdk1-Foxo1 signaling in myeloid cells predisposes to adipose tissue inflammation and insulin resistance. Diabetes 2012, 61, 1935–1948. [Google Scholar] [CrossRef] [Green Version]
- Linton, M.F.; Moslehi, J.J.; Babaev, V.R. Akt signaling in macrophage polarization, survival, and atherosclerosis. Int. J. Mol. Sci. 2019, 20, 2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luyendyk, J.P.; Schabbauer, G.A.; Tencati, M.; Holscher, T.; Pawlinski, R.; Mackman, N. Genetic Analysis of the Role of the PI3K-Akt Pathway in Lipopolysaccharide-Induced Cytokine and Tissue Factor Gene Expression in Monocytes/Macrophages. J. Immunol. 2008, 180, 4218–4226. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Haubenwallner, S.; Kuttke, M.; Kollmann, I.; Halfmann, A.; Dohnal, A.B.; Chen, L.; Cheng, P.; Hoesel, B.; Einwallner, E.; et al. Macrophage PTEN Regulates Expression and Secretion of Arginase I Modulating Innate and Adaptive Immune Responses. J. Immunol. 2014, 193, 1717–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, S.; Rao, J.; Zhu, J.; Busuttil, R.W.; Kupiec-Weglinski, J.W.; Lu, L.; Wang, X.; Zhai, Y. Myeloid PTEN Deficiency Protects Livers from Ischemia Reperfusion Injury by Facilitating M2 Macrophage Differentiation. J. Immunol. 2014, 192, 5343–5353. [Google Scholar] [CrossRef] [Green Version]
- Weisser, S.B.; Mclarren, K.W.; Voglmaier, N.; van Netten-Thomas, C.J.; Antov, A.; Flavell, R.A.; Sly, L.M. Alternative activation of macrophages by IL-4 requires SHIP degradation. Eur. J. Immunol. 2011, 41, 1742–1753. [Google Scholar] [CrossRef]
- Kuroda, E.; Ho, V.; Ruschmann, J.; Antignano, F.; Hamilton, M.; Rauh, M.J.; Antov, A.; Flavell, R.A.; Sly, L.M.; Krystal, G. SHIP Represses the Generation of IL-3-Induced M2 Macrophages by Inhibiting IL-4 Production from Basophils. J. Immunol. 2009, 183, 3652–3660. [Google Scholar] [CrossRef]
- Tan, Z.; Xie, N.; Cui, H.; Moellering, D.R.; Abraham, E.; Thannickal, V.J.; Liu, G. Pyruvate Dehydrogenase Kinase 1 Participates in Macrophage Polarization via Regulating Glucose Metabolism. J. Immunol. 2015, 194, 6082–6089. [Google Scholar] [CrossRef]
- Fang, C.; Yu, J.; Luo, Y.; Chen, S.; Wang, W.; Zhao, C.; Sun, Z.; Wu, W.; Guo, W.; Han, Z.; et al. Tsc1 is a Critical Regulator of Macrophage Survival and Function. Cell. Physiol. Biochem. 2015, 36, 1406–1418. [Google Scholar] [CrossRef] [PubMed]
- Weichhart, T.; Costantino, G.; Poglitsch, M.; Rosner, M.; Zeyda, M.; Stuhlmeier, K.M.; Kolbe, T.; Stulnig, T.M.; Hörl, W.H.; Hengstschläger, M.; et al. The TSC-mTOR Signaling Pathway Regulates the Innate Inflammatory Response. Immunity 2008, 29, 565–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covarrubias, A.J.; Aksoylar, H.I.; Yu, J.; Snyder, N.W.; Worth, A.J.; Iyer, S.S.; Wang, J.; Ben-Sahra, I.; Byles, V.; Polynne-Stapornkul, T.; et al. Akt-mTORC1 signaling regulates Acly to integrate metabolic input to control of macrophage activation. eLife 2016, 5, e11612. [Google Scholar] [CrossRef] [PubMed]
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. Principles of innate and adaptive immunity. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Hirayama, D.; Iida, T.; Nakase, H. The phagocytic function of macrophage-enforcing innate immunity and tissue homeostasis. Int. J. Mol. Sci. 2018, 19, 92. [Google Scholar] [CrossRef] [Green Version]
- Bao, H.R.; Chen, J.L.; Li, F.; Zeng, X.L.; Liu, X.J. Relationship between pi3k/mtor/rhoa pathway-regulated cytoskeletal rearrangements and phagocytic capacity of macrophages. Braz. J. Med. Biol. Res. 2020, 53, e9207. [Google Scholar] [CrossRef]
- Zhang, B.; Ma, Y.; Guo, H.; Sun, B.; Niu, R.; Ying, G.; Zhang, N. Akt2 is required for macrophage chemotaxis. Eur. J. Immunol. 2009, 39, 894–901. [Google Scholar] [CrossRef]
- Vural, A.; Kehrl, J.H. Autophagy in Macrophages: Impacting Inflammation and Bacterial Infection. Scientifica 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Cescon, M.; Bonaldo, P. Autophagy-mediated regulation of macrophages and its applications for cancer. Autophagy 2014, 10, 192–200. [Google Scholar] [CrossRef]
- Rocher, C.; Singla, D.K. SMAD-PI3K-Akt-mTOR pathway mediates BMP-7 polarization of monocytes into M2 macrophages. PLoS ONE 2013, 8, e84009. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Ma, T.; Shen, X.N.; Xia, X.F.; Xu, G.D.; Bai, X.L.; Liang, T.B. Macrophage-induced tumor angiogenesis is regulated by the TSC2-mTOR pathway. Cancer Res. 2012, 72, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freire, M.O.; Van Dyke, T.E. Natural resolution of inflammation. Periodontol. 2000 2013, 63, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Alexander, M.; Misharin, A.V.; Budinger, G.R.S. The role of macrophages in the resolution of inflammation. J. Clin. Investig. 2019, 129, 2619–2628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pahwa, R.; Goyal, A.; Bansal, P.; Jialal, I. Chronic Inflammation; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Schabbauer, G.; Tencati, M.; Pedersen, B.; Pawlinski, R.; Mackman, N. PI3K-Akt pathway suppresses coagulation and inflammation in endotoxemic mice. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1963–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson-Casey, J.L.; Deshane, J.S.; Ryan, A.J.; Thannickal, V.J.; Carter, A.B. Macrophage Akt1 Kinase-Mediated Mitophagy Modulates Apoptosis Resistance and Pulmonary Fibrosis. Immunity 2016, 44, 582–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, F.; Lee, J.T.; Navolanic, P.M.; Steelman, L.S.; Shelton, J.G.; Blalock, W.L.; Franklin, R.A.; McCubrey, J.A. Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: A target for cancer chemotherapy. Leukemia 2003, 17, 590–603. [Google Scholar] [CrossRef] [Green Version]
- Deretic, V.; Saitoh, T.; Akira, S. Autophagy in infection, inflammation and immunity. Nat. Rev. Immunol. 2013, 13, 722–737. [Google Scholar] [CrossRef]
- Hankittichai, P.; Buacheen, P.; Pitchakarn, P.; Takuathung, M.N.; Wikan, N.; Smith, D.R.; Potikanond, S.; Nimlamool, W. Artocarpus lakoocha extract inhibits lps-induced inflammatory response in raw 264.7 macrophage cells. Int. J. Mol. Sci. 2020, 21, 1355. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.J.; Jeong, J.S.; Park, N.J.; Go, G.B.; Kim, S.O.; Park, C.; Kim, B.W.; Hong, S.H.; Choi, Y.H. An ethanol extract of Aster yomena (Kitam.) Honda inhibits lipopolysaccharide-induced inflammatory responses in murine RAW 264.7 macrophages. Biosci. Trends 2017, 11, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liang, T.; Yang, W.; Zhang, L.; Wu, S.; Yan, C.; Li, Q. Astragalus membranaceus Injection Suppresses Production of Interleukin-6 by Activating Autophagy through the AMPK-mTOR Pathway in Lipopolysaccharide-Stimulated Macrophages. Oxid. Med. Cell. Longev. 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.S.; Kim, D.; Yi, Y.S.; Kim, J.H.; Jeong, H.Y.; Hwang, K.; Kim, J.H.; Park, J.; Cho, J.Y. AKT-targeted anti-inflammatory activity of the methanol extract of Chrysanthemum indicum var. albescens. J. Ethnopharmacol. 2017, 201, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Kunworarath, N.; Rangkadilok, N.; Suriyo, T.; Thiantanawat, A.; Satayavivad, J. Longan (Dimocarpus longan Lour.) inhibits lipopolysaccharide-stimulated nitric oxide production in macrophages by suppressing NF-κB and AP-1 signaling pathways. J. Ethnopharmacol. 2016, 179, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Schmiech, M.; El Gaafary, M.; Zhang, X.; Syrovets, T.; Simmet, T. A comparative study on root and bark extracts of Eleutherococcus senticosus and their effects on human macrophages. Phytomedicine 2020, 68, 153181. [Google Scholar] [CrossRef]
- Koh, W.; Shin, J.S.; Lee, J.; Lee, I.H.; Lee, S.K.; Ha, I.H.; Chung, H.J. Anti-inflammatory effect of Cortex Eucommiae via modulation of the toll-like receptor 4 pathway in lipopolysaccharide-stimulated RAW 264.7 macrophages. J. Ethnopharmacol. 2017, 209, 255–263. [Google Scholar] [CrossRef]
- Dung, T.T.M.; Lee, J.; Kim, E.; Yoo, B.C.; Ha, V.T.; Kim, Y.; Yoon, D.H.; Hong, S.; Baek, K.S.; Sung, N.Y.; et al. Anti-inflammatory activities of gouania leptostachya methanol extract and its constituent resveratrol. Phyther. Res. 2015, 29, 381–392. [Google Scholar] [CrossRef]
- Hu, W.; Wu, L.; Qiang, Q.; Ji, L.; Wang, X.; Luo, H.; Wu, H.; Jiang, Y.; Wang, G.; Shen, T. The dichloromethane fraction from Mahonia bealei (Fort.) Carr. leaves exerts an anti-inflammatory effect both in vitro and in vivo. J. Ethnopharmacol. 2016, 188, 134–143. [Google Scholar] [CrossRef]
- Jeong, S.G.; Kim, S.; Kim, H.G.; Kim, E.; Jeong, D.; Kim, J.H.; Yang, W.S.; Oh, J.; Sung, G.H.; Hossain, M.A.; et al. Mycetia cauliflora methanol extract exerts anti-inflammatory activity by directly targeting PDK1 in the NF-κB pathway. J. Ethnopharmacol. 2019, 231, 1–9. [Google Scholar] [CrossRef]
- Nguyen, C.T.; Luong, T.T.; Lee, S.Y.; Kim, G.L.; Kwon, H.; Lee, H.G.; Park, C.K.; Rhee, D.K. Panax ginseng aqueous extract prevents pneumococcal sepsis in vivo by potentiating cell survival and diminishing inflammation. Phytomedicine 2015, 22, 1055–1061. [Google Scholar] [CrossRef]
- Han, S.Y.; Kim, J.; Kim, E.; Kim, S.H.; Seo, D.B.; Kim, J.H.; Shin, S.S.; Cho, J.Y. AKT-targeted anti-inflammatory activity of Panax ginseng calyx ethanolic extract. J. Ginseng Res. 2018, 42, 496–503. [Google Scholar] [CrossRef]
- Um, Y.; Eo, H.J.; Kim, H.J.; Kim, K.; Jeon, K.S.; Jeong, J.B. Wild simulated ginseng activates mouse macrophage, RAW264.7 cells through TRL2/4-dependent activation of MAPK, NF-κB and PI3K/AKT pathways. J. Ethnopharmacol. 2020, 263, 113218. [Google Scholar] [CrossRef] [PubMed]
- Hossen, M.J.; Baek, K.S.; Kim, E.; Yang, W.S.; Jeong, D.; Kim, J.H.; Kweon, D.H.; Yoon, D.H.; Kim, T.W.; Kim, J.H.; et al. In vivo and in vitro anti-inflammatory activities of Persicaria chinensis methanolic extract targeting Src/Syk/NF-κB. J. Ethnopharmacol. 2015, 159, 9–16. [Google Scholar] [CrossRef]
- Hossen, M.J.; Jeon, S.H.; Kim, S.C.; Kim, J.H.; Jeong, D.; Sung, N.Y.; Yang, S.; Baek, K.S.; Kim, J.H.; Yoon, D.H.; et al. In vitro and in vivo anti-inflammatory activity of Phyllanthus acidus methanolic extract. J. Ethnopharmacol. 2015, 168, 217–228. [Google Scholar] [CrossRef]
- Gong, G.; Xie, F.; Zheng, Y.; Hu, W.; Qi, B.; He, H.; Dong, T.T.; Tsim, K.W. The effect of methanol extract from Saussurea involucrata in the lipopolysaccharide-stimulated inflammation in cultured RAW 264.7 cells. J. Ethnopharmacol. 2020, 251, 112532. [Google Scholar] [CrossRef]
- Kuo, C.E.A.; Wu, S.Y.; Lee, C.H.; Lai, Y.R.; Lu, C.H.; Chen, P.C.; Cheng, J.H.; Tsai, L.Y.; Yen, K.T.; Tsao, Y.; et al. Toona sinensis modulates autophagy and cytokines in lipopolysaccharide-induced RAW 264.7 macrophages. Biomed. Pharmacother. 2020, 129, 110386. [Google Scholar] [CrossRef] [PubMed]
- Park, G.; Kim, E.; Son, Y.J.; Yoon, D.H.; Sung, G.H.; Aravinthan, A.; Park, Y.C.; Kim, J.H.; Cho, J.Y. Anti-inflammatory effect of torilidis fructus ethanol extract through inhibition of Src. Pharm. Biol. 2017, 55, 2074–2082. [Google Scholar] [CrossRef]
- Shin, J.Y.; Kang, J.S.; Byun, H.W.; Ahn, E.K. Regulatory effects and molecular mechanism of trigonostemon reidioides on lipopolysaccharide-induced inflammatory responses in RAW264.7 cells. Mol. Med. Rep. 2017, 16, 5137–5142. [Google Scholar] [CrossRef]
- Cho, S.K.; Jeong, M.; Jang, D.S.; Choi, J.H. Anti-inflammatory Effects of Canthin-6-one Alkaloids from Ailanthus altissima. Planta Med. 2018, 84, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Q.; Zhang, S.; Yu, L.; Li, Y.; Liu, Y.; Ye, X.; Zheng, X. Apios americana Medikus tuber polysaccharide exerts anti-inflammatory effects by activating autophagy. Int. J. Biol. Macromol. 2019, 130, 892–902. [Google Scholar] [CrossRef]
- Jiang, F.; Li, M.; Wang, H.; Ding, B.; Zhang, C.; Ding, Z.; Yu, X.; Lv, G. Coelonin, an anti-inflammation active component of Bletilla striata and its potential mechanism. Int. J. Mol. Sci. 2019, 20, 4422. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.S.; Park, J.Y.; Lee, J.; Yoo, H.H.; Hahm, D.H.; Lee, S.C.; Lee, S.; Hwang, G.S.; Jung, K.; Kang, K.S. Anti-inflammatory effects and corresponding mechanisms of cirsimaritin extracted from Cirsium japonicum var. maackii Maxim. Bioorganic Med. Chem. Lett. 2017, 27, 3076–3080. [Google Scholar] [CrossRef] [PubMed]
- Mankhong, S.; Iawsipo, P.; Srisook, E.; Srisook, K. 4-methoxycinnamyl p-coumarate isolated from Etlingera pavieana rhizomes inhibits inflammatory response via suppression of NF-κB, Akt and AP-1 signaling in LPS-stimulated RAW 264.7 macrophages. Phytomedicine 2019, 54, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, F.; Luo, Y.; Zhang, L.; Lu, S.; Xing, R.; Yan, B.; Zhang, H.; Hu, W. 20-Hydroxy-3-oxolupan-28-oic acid attenuates inflammatory responses by regulating PI3K–AKT and MAPKs signaling pathways in LPS-stimulated RAW264.7 macrophages. Molecules 2019, 24, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, X.; Han, L.; Qu, T.; Mu, Y.; Guan, P.; Qu, X.; Wang, Z.; Huang, X. Anti-inflammatory effects, SAR, and action mechanism of monoterpenoids from radix paeoniae alba on LPS-stimulated RAW264.7 cells. Molecules 2017, 22, 715. [Google Scholar] [CrossRef] [Green Version]
- Park, E.J.; Sang-Ngern, M.; Chang, L.C.; Pezzuto, J.M. Physalactone and 4β-Hydroxywithanolide e Isolated from Physalis peruviana Inhibit LPS-Induced Expression of COX-2 and iNOS Accompanied by Abatement of Akt and STAT1. J. Nat. Prod. 2019, 82, 492–499. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, L.; Di, S.n.; Xu, Q.; Ren, Q.c.; Chen, S.z.; Huang, N.; Jia, D.; Shen, X.-f. Steroidal alkaloid solanine A from Solanum nigrum Linn. exhibits anti-inflammatory activity in lipopolysaccharide/interferon γ-activated murine macrophages and animal models of inflammation. Biomed. Pharmacother. 2018, 105, 606–615. [Google Scholar] [CrossRef]
- Guo, C.; Yang, L.; Luo, J.; Zhang, C.; Xia, Y.; Ma, T.; Kong, L. Sophoraflavanone G from Sophora alopecuroides inhibits lipopolysaccharide-induced inflammation in RAW264.7 cells by targeting PI3K/Akt, JAK/STAT and Nrf2/HO-1 pathways. Int. Immunopharmacol. 2016, 38, 349–356. [Google Scholar] [CrossRef]
- Utaipan, T.; Suksamrarn, A.; Kaemchantuek, P.; Chokchaisiri, R.; Stremmel, W.; Chamulitrat, W.; Chunglok, W. Diterpenoid trigonoreidon B isolated from Trigonostemon reidioides alleviates inflammation in models of LPS-stimulated murine macrophages and inflammatory liver injury in mice. Biomed. Pharmacother. 2018, 101, 961–971. [Google Scholar] [CrossRef]
- Yan, T.; Yu, X.; Sun, X.; Meng, D.; Jia, J.-m. A new steroidal saponin, furotrilliumoside from Trillium tschonoskii inhibits lipopolysaccharide-induced inflammation in Raw264.7 cells by targeting PI3K/Akt, MARK and Nrf2/HO-1 pathways. Fitoterapia 2016, 115, 37–45. [Google Scholar] [CrossRef]
- Haque, M.A.; Jantan, I.; Harikrishnan, H. Zerumbone suppresses the activation of inflammatory mediators in LPS-stimulated U937 macrophages through MyD88-dependent NF-κB/MAPK/PI3K-Akt signaling pathways. Int. Immunopharmacol. 2018, 55, 312–322. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name of the Species | Name of the Family | Part of the Plant | Type of Extract | Determined Chemicals | Cell Line | Mechanism of Action | Ref. |
|---|---|---|---|---|---|---|---|
| Apios americana Medikus | Fabaceae | leaves | ethanolic | vicenin-2, schaftoside, baimaside, apigenin (6-C-α-l-arabinopyranosyl)-8-C-β-d-glucopyranoside, vitexin, isquercetin, 6-C-β-d-glucopyranosyl-7-O-methylluteolin | RAW 264.7 cells | Inhibition of AKT and mTORC1 activity | [102] |
| Artocarpus lakoocha Roxb. | Moraceae | heartwood | ethanolic | oxyresveratrol (trans-2,30,4,50-tetrahydroxystilbene) | RAW 264.7 cells | Inhibition of AKT activity | [204] |
| Aster incisus Fisch. | Asteraceae | whole plant | methanolic | 4-((1E)-3-Hydroxy-1-propenyl)-2-methoxyphenol, liliolide, neophytadiene, triterpene lupeol, trans-phytol, palmitic acid beta-monoglyceride, chondrillasterol, olean-12-en-3-one, palmitic acid | RAW 264.7 cells | Inhibition of PI3K and AKT proteins expression | [109] |
| Aster yomena (Kitam.) Honda | Asteraceae | leaves | ethanolic | - | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [205] |
| Astragalus membranaceus (Fisch.) Bunge | Fabaceae | whole plant | - | - | ANA-1 cells | Inhibition of AKT activity | [206] |
| Chrysanthemum indicum L. | Asteraceae | methanolic | luteolin | RAW264.7 cells | Inhibition of AKT1 and AKT2 activity | [207] | |
| Dimocarpus longan Lour. | Sapindaceae | flowers, seeds, and pulp | aqueous | corilagin, ellagic acid, and gallic acid | RAW 264.7 cells | Inhibition of AKT activity | [208] |
| Eleutherococcus senticosus (Rupr. et Maxim.) Maxim | Araliaceae | roots and bark | root—ethanolic bark—methanolic | root extract containing more syringin, caffeic acid, and isofraxidin than the bark the bark extract contained more sesamin and oleanolic acid | human primary macrophages | Inhibition of AKT activity | [209] |
| Eucommia ulmoides Oliv. | Eucommiaceae | bark | aqueous | pinoresinol diglucoside, geniposide, aucubin | RAW 264.7 cells | Inhibition of PI3K, AKT, mTORC1 activity | [210] |
| Gouania leptostachya DC. var. tonkinensis Pitard. | Rhamnaceae | leaves | methanolic | resveratrol | RAW264.7 cells and mouse peritoneal macrophages | Inhibition of AKT activity | [211] |
| Mahonia bealei (Fort.) Carr. | Berberidaceae | leaves | aqueous extract and fractionated using a series of organic solvents, including n-hexane, dichloromethane, ethylacetate, n-butanol | - | RAW 264.7 cells | Inhibition of PI3K, AKT activity | [212] |
| Mycetia cauliflora Reinw. | Rubiaceae | aerial parts | methanolic | - | RAW 264.7 cells | Inhibition of AKT activity | [213] |
| Panax ginseng C.A. Meyer | Araliaceae | roots | aqueous | - | RAW 264.7 cells | Activation of PI3K and AKT activity | [214] |
| Panax ginseng Mayer | Araliaceae | berry calyx | ethanolic | ginsenosides | RAW264.7 cells | Inhibition of AKT1 and AKT2 activity | [215] |
| Panax ginseng Meyer | Araliaceae | roots | aqueous | ginsenosides | RAW264.7 cells | Induction of PI3K, AKT activity | [216] |
| Persicaria chinensis L. | Polygonaceae | aerial parts | methanolic | caffeic acid, kaempferol, and quercetin | peritoneal macrophages, and RAW264.7 cells | Inhibition of AKT activity | [217] |
| Phyllanthus acidus (L.) Skeels | Phyllanthaceae | leaves | methanolic | quercetin, kaempferol, caffeic acid | RAW 264.7 cells, U937 cells | Inhibition of AKT activity | [218] |
| Phyllanthus amarus Schum. & Thonn. | Phyllanthaceae | whole plant | ethanolic | lignans, phyllanthin, hypophyllahtin and niranthin, and polyphenols, gallic acid, geraniin, corilagin, ellagic acid | U937 cells | Inhibition of AKT protein expression | [104] |
| Piper cubeba L. | Piperaceae | whole plant | methanolic | quercetin, luteolin, and kaempferol | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [66] |
| Saussurea involucrate (Kar. et Kir.) Sch.-Bip. | Asteraceae | aerial parts | methanolic | rutin, chlorogenic acid | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [219] |
| Toona sinensis (Juss.) M. Roem | Meliaceae | leaves | aqueous | gallic acid, methyl gallate, ethyl gallate, kaempferol, kaempferol 3-O-b-d-glucoside, quercetin, quercitrin, quercetin 3-O-b-d-glucoside, rutin | RAW 264.7 cells | Inhibition of mTORC1 activity | [220] |
| Torilis japonica (Houtt.) D.C. | Umbelliferae | fructus | ethanolic | - | RAW 264.7 cells | Inhibition of AKT activity | [221] |
| Trigonostemon reidioides (Kurz). Craib | Euphorbiaceae | stems | ethanolic | - | RAW 264.7 cells | Inhibition of PI3K, AKT activity | [222] |
| Zingiber zerumbet (L.) Roscoe ex Sm. | Zingiberaceae | rhizomes | ethanolic | zerumbone, ethyl gallate, gallic acid, catechin, kaempferol rhamnoside, kaempferol, kaempferol-3-O-(2”, 4”-diacetyl) rhamnoside isomers, kaempferol methylether, kaempferol methylether isomer, kaempferol-3-O-(3”,4”-diacetyl) rhamnoside isomers, kaempferol glucoside conjugate, demethoxycurcumin, curcumin, bisdemethoxycurcumin | U937 cells | Inhibition of AKT activity | [101] |
| Name of the Species | Name of the Family | Part of the Plant | Name of the Chemical | Class of the Chemical | Cell Line | Mechanism of Action | Ref. |
|---|---|---|---|---|---|---|---|
| Ailanthus altissima (Mill.) Swingle | Simarubaceae | stem bark | canthin-6-one | alkaloid | RAW 264.7 cells | Inhibition of AKT activity | [223] |
| Apios americana Medikus | Fabaceae | tubers | polysaccharide (glucose, arabinose, galactose and galacturonic acid) | polysaccharide | RAW 264.7 cells | Inhibition of AKT and mTOR activity | [224] |
| Bletilla striata (Thunb.) Rchb.f. | Orchidaceae | whole plant | coelonin | dihydrophenanthrene | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [225] |
| Cirsium japonicum Fisch. ex DC. | Asteraceae | aerial parts | cirsimaritin | flavonoid | RAW 264.7 cells | Inhibition of AKT activity | [226] |
| Etlingera pavieana (Pierre ex Gagnep) R.M.Sm. | Zingiberaceae | rhizomes | 4-methoxycinnamyl p-coumarate | phenolic compound | RAW 264.7 cells | Inhibition of AKT activity | [227] |
| Mahonia bealei (Fortune) Carrière | Berberidaceae | leaves | 20-hydroxy-3-oxolupan-28-oic acid | triterpene | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [228] |
| Paeonia lactiflora Pall. | Paeoniaceae | rhizomes | 4-O-methylbenzoylpaeoniflorin | monoterpene | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [229] |
| Physalis peruviana L. | Solanaceae | aerial part | 4β-hydroxywithanolide E and physalactone | withanolides | RAW 264.7 cells | Inhibition of AKT activity | [230] |
| Solanum nigrum L. | Solanaceae | fruits | solanine A | alkaloid | RAW 264.7 cells | Inhibition of AKT activity | [231] |
| Sophora alopecuroides L. | Fabaceae | roots | sophoraflavanone G | flavonoid | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [232] |
| Swertia chirayita (Roxb. ex Fleming) H. Karst. | Gentianaceae | whole plant | bellidifolin | xanthone | RAW 264.7 cells | Inhibition of AKT activity | [118] |
| Trigonostemon reidioides (Kurz) Craib. | Euphorbiaceae | roots | trigonoreidon B | diterpene | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [233] |
| Trillium tschonoskii Maxim. | Liliaceae | roots and rhizomes | urotrilliumoside | saponin | RAW 264.7 cells | Inhibition of PI3K and AKT activity | [234] |
| Zingiber zerumbet (L.) Sm. | Zingiberaceae | rhizomes | zerumbone | sesquiterpene | U937 cells | Inhibition of AKT activity | [235] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merecz-Sadowska, A.; Sitarek, P.; Śliwiński, T.; Zajdel, R. Anti-Inflammatory Activity of Extracts and Pure Compounds Derived from Plants via Modulation of Signaling Pathways, Especially PI3K/AKT in Macrophages. Int. J. Mol. Sci. 2020, 21, 9605. https://doi.org/10.3390/ijms21249605
Merecz-Sadowska A, Sitarek P, Śliwiński T, Zajdel R. Anti-Inflammatory Activity of Extracts and Pure Compounds Derived from Plants via Modulation of Signaling Pathways, Especially PI3K/AKT in Macrophages. International Journal of Molecular Sciences. 2020; 21(24):9605. https://doi.org/10.3390/ijms21249605
Chicago/Turabian StyleMerecz-Sadowska, Anna, Przemysław Sitarek, Tomasz Śliwiński, and Radosław Zajdel. 2020. "Anti-Inflammatory Activity of Extracts and Pure Compounds Derived from Plants via Modulation of Signaling Pathways, Especially PI3K/AKT in Macrophages" International Journal of Molecular Sciences 21, no. 24: 9605. https://doi.org/10.3390/ijms21249605