Tissue Engineering of Cartilage Using a Random Positioning Machine

 , ,
, ,
Abstract
:1. Introduction
2. Results
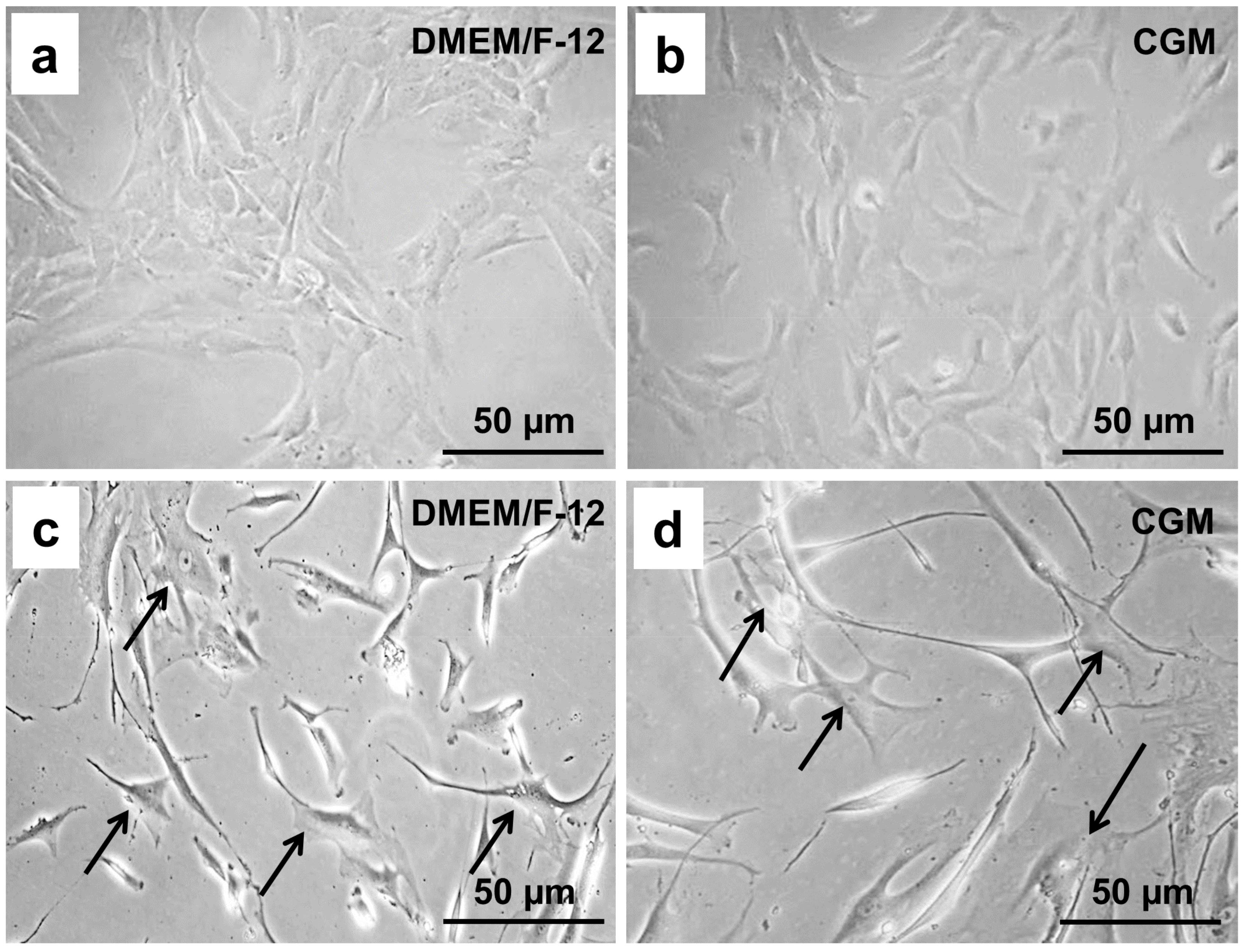
2.1. Chondrocyte Morphology Depends on the Cultivation Medium
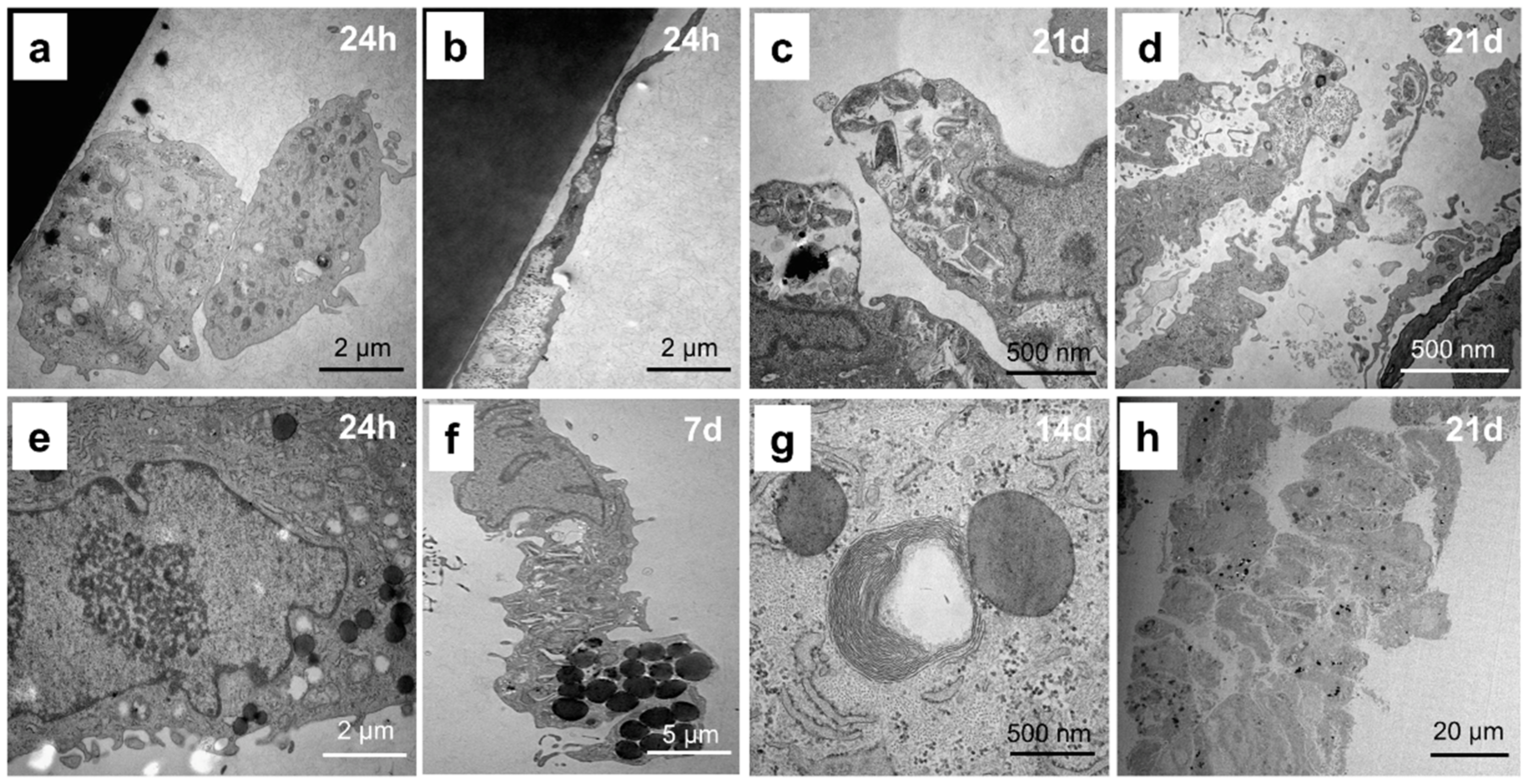
2.2. Chondrocytes Change Their Cellular Structure during Cultivation
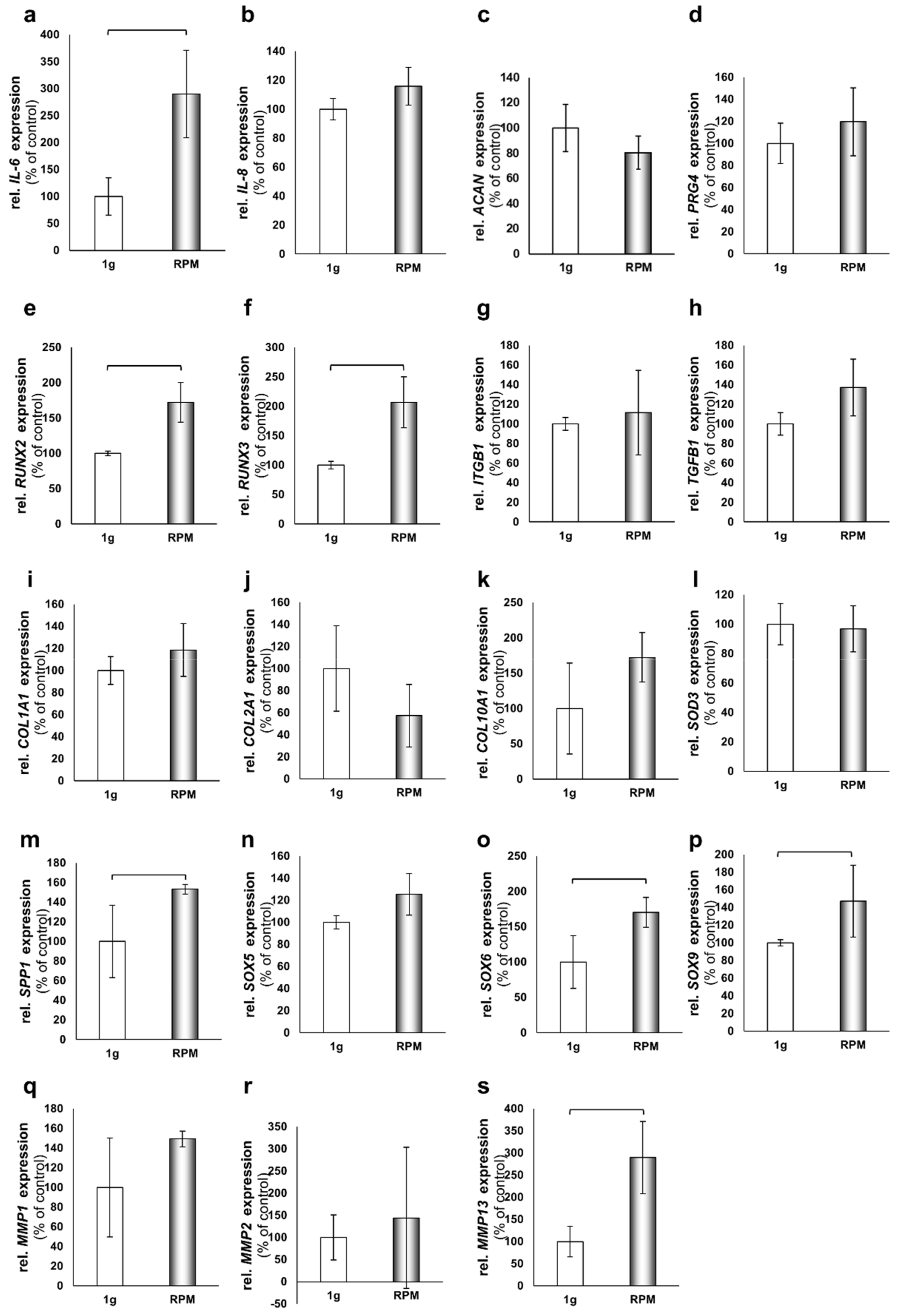
2.3. 24-h Short-Term Gene Expression Changes on the RPM
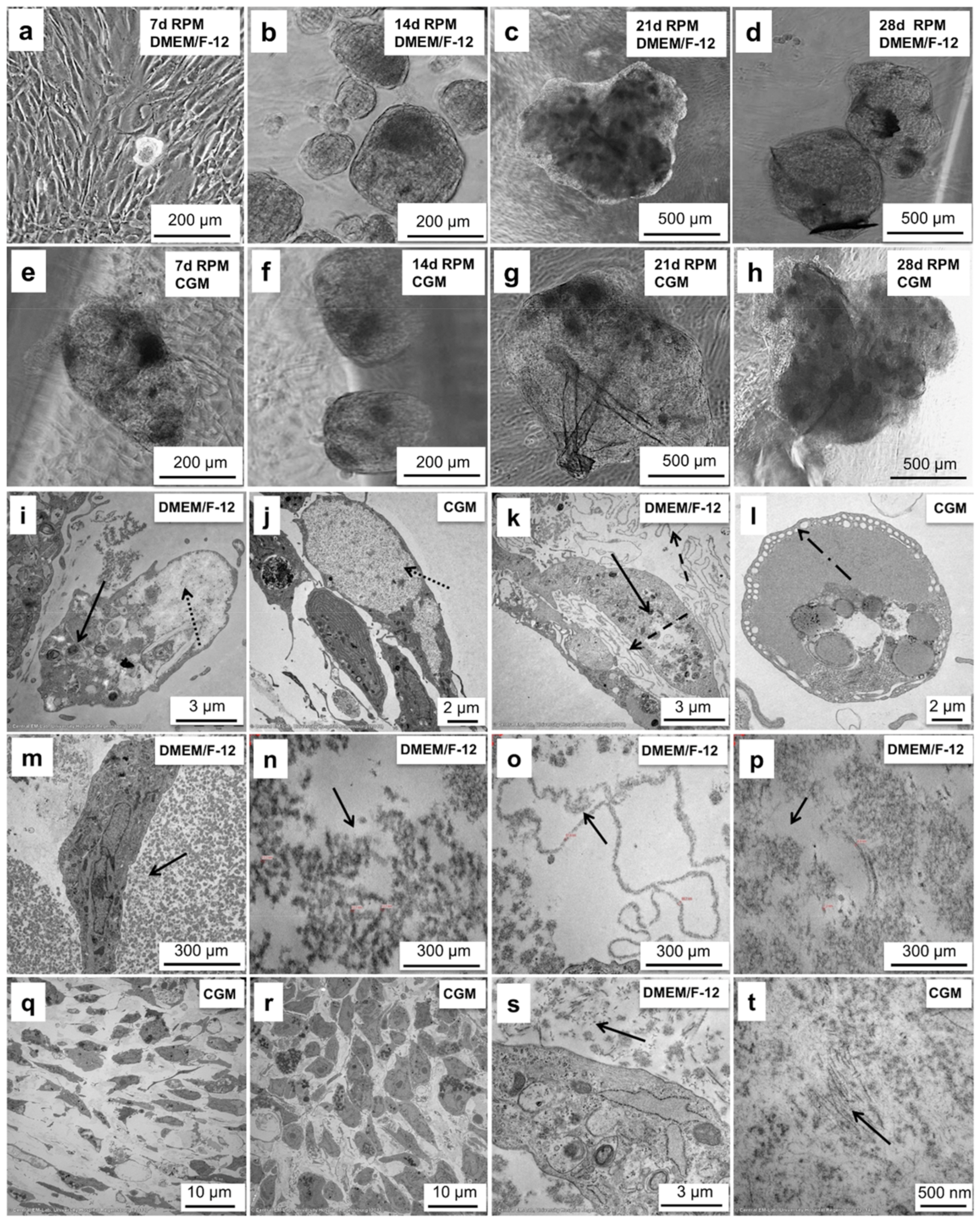
2.4. Chondrocytes Produce Spheroids on the RPM up to 2 mm in Size
2.5. Chondrocyte Spheroids Have a Rich Morphology and Form Collagen Fibrils
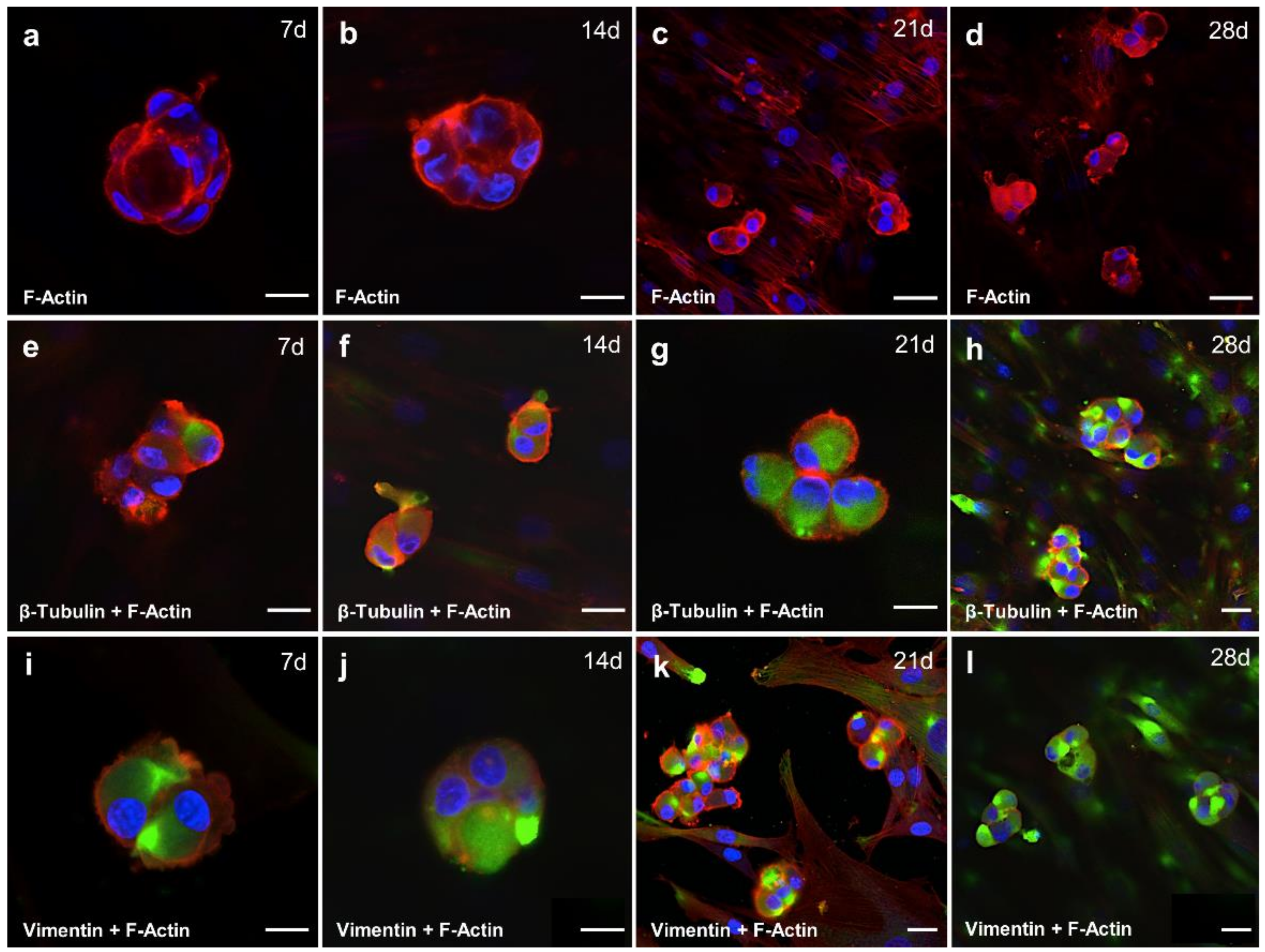
2.6. Cytoskeletal Composition of Spheroid Changes with the Duration of the Cultivation Time
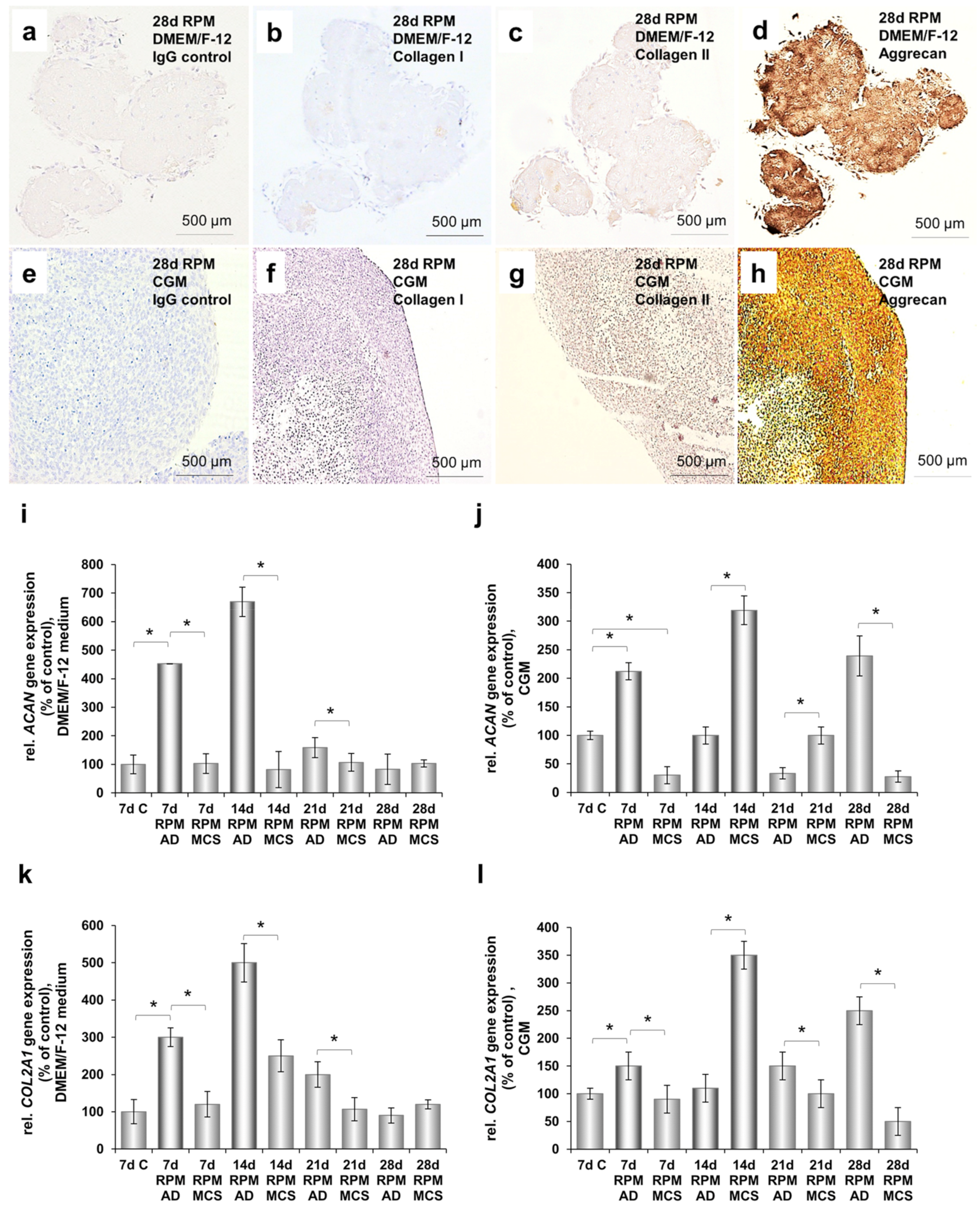
2.7. Cartilage-Like Tissue Reacts Positive Against Cartilage Markers and Produces Cartilage-Specific Proteins
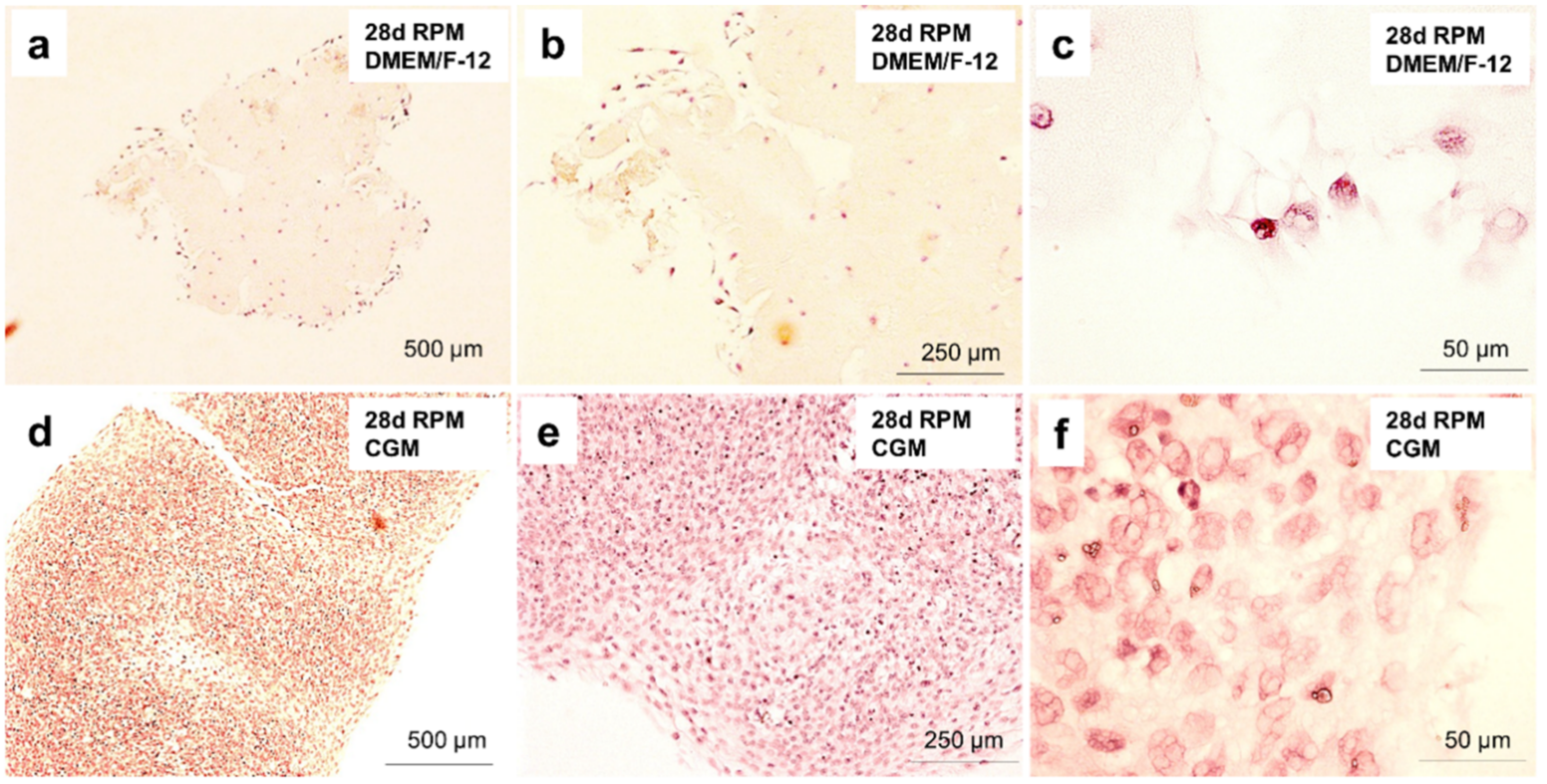
2.8. Cartilage Tissue Production is Medium-Dependent
2.9. Spheroids Produced on the RPM Form Monolayers under 1 g Culture Conditions
3. Discussion
4. Materials and Methods
4.1. Cells and Culture Medium
4.2. Random Positioning Machine
4.3. Histological Analysis
4.4. Immunohistochemical Analyses
4.5. Immunofluorescence Staining
4.6. F-Actin Staining
4.7. Microscopy
4.8. Transmission Electron Microscopy (TEM)
4.9. RNA Isolation
4.10. Quantitative Real-Time PCR
4.11. STRING-Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| µg | Microgravity |
| ACAN | Aggrecan |
| bFGF | Basic fibroblast growth factor |
| CGM | Chondrocyte growth medium |
| COL10A1 | Collagen alpha-1(X) chain |
| COL1A1 | Collagen, type I, alpha 1, |
| COL2A1 | Collagen, type II, alpha 1 |
| DMEM | Dulbecco’s Modified Eagle Medium |
| ECM | Extracellular matrix |
| EGF | Epidermal growth factor |
| ERK | Extracellular signal-regulated kinase |
| FCS | Fetal calf serum |
| IL6 | Interleukin 6 |
| IL8 | Interleukin 8 |
| ITGB1 | Integrin beta-1 |
| JAK | Janus kinase |
| MAPK | Mitogen-activated protein kinase |
| MMP1 | Matrix metalloproteinase-1 |
| MMP2 | Matrix metalloproteinase-2 |
| MMP13 | Matrix metalloproteinase-13 |
| PFA | Paraformaldehyde |
| OA | Ostheoarthritis |
| PRG4 | Proteoglycan 4 |
| qPCR | Quantitative real-time PCR |
| R3 IGF-1 | R3 insulin-like growth factor-1 |
| RPM | Random Positioning Machine |
| RUNX2 | Runt-related transcription factor 2 |
| RUNX3 | Runt-related transcription factor 3 |
| RWV | Rotating Wall Vessel |
| SOD3 | Extracellular superoxide dismutase |
| SOX5 | SRY-related HMG-box transcription factor 5 |
| SOX6 | SRY-related HMG-box transcription factor 6 |
| SOX9 | SRY-related HMG-box transcription factor 9 |
| SPP1 | Osteopontin |
| STAT | Signal transducer and activator of transcription |
| TEM | Transmission electron microscopy |
| TGFB1 | Transforming growth factor beta 1 |
| VEGF | Vascular endothelial growth factor |
References
- Chang, C.-H.; Lin, F.-H.; Kuo, T.-F.; Liu, H.-C. Cartilage Tissue Engineering. Biomed. Eng. Appl. Basis Commun. 2005, 17, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Mehrotra, C.; Remington, P.L.; Naimi, T.S.; Washington, W.; Miller, R. Trends in total knee replacement surgeries and implications for public health, 1990–2000. Public Health Rep. 2005, 120, 278–282. [Google Scholar] [CrossRef]
- Kock, L.; van Donkelaar, C.C.; Ito, K. Tissue engineering of functional articular cartilage: The current status. Cell Tissue Res. 2012, 347, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Uemura, T.; Nishi, M.; Aoki, K.; Tsumura, T. Cartilage Regeneration from Bone Marrow Cells Using RWV Bioreactor and Its Automation System for Clinical Application. In Tissue Engineering for Tissue and Organ Regeneration; Eberli, D., Ed.; IntechOpen: Rijeka, Croatia, 2011. [Google Scholar]
- Wenger, R.; Hans, M.G.; Welter, J.F.; Solchaga, L.A.; Sheu, Y.R.; Malemud, C.J. Hydrostatic pressure increases apoptosis in cartilage-constructs produced from human osteoarthritic chondrocytes. Front. Biosci. 2006, 11, 1690–1695. [Google Scholar] [CrossRef] [Green Version]
- Marlovits, S.; Tichy, B.; Truppe, M.; Gruber, D.; Vecsei, V. Chondrogenesis of aged human articular cartilage in a scaffold-free bioreactor. Tissue Eng. 2003, 9, 1215–1226. [Google Scholar] [CrossRef]
- Giannoni, P.; Pagano, A.; Maggi, E.; Arbico, R.; Randazzo, N.; Grandizio, M.; Cancedda, R.; Dozin, B. Autologous chondrocyte implantation (ACI) for aged patients: Development of the proper cell expansion conditions for possible therapeutic applications. Osteoarthr. Cartil. 2005, 13, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Foldager, C.B.; Nielsen, A.B.; Munir, S.; Ulrich-Vinther, M.; Soballe, K.; Bunger, C.; Lind, M. Combined 3D and hypoxic culture improves cartilage-specific gene expression in human chondrocytes. Acta Orthopeadica 2011, 82, 234–240. [Google Scholar] [CrossRef]
- Barbero, A.; Grogan, S.; Schafer, D.; Heberer, M.; Mainil-Varlet, P.; Martin, I. Age related changes in human articular chondrocyte yield, proliferation and post-expansion chondrogenic capacity. Osteoarthr. Cartil. 2004, 12, 476–484. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.L.; Mauck, R.L.; Burdick, J.A. Hydrogel design for cartilage tissue engineering: A case study with hyaluronic acid. Biomaterials 2011, 32, 8771–8782. [Google Scholar] [CrossRef] [Green Version]
- Darling, E.M.; Athanasiou, K.A. Rapid phenotypic changes in passaged articular chondrocyte subpopulations. J. Orthop. Res. 2005, 23, 425–432. [Google Scholar] [CrossRef]
- Saltzman, W.M. Weaving cartilage at zero g: The reality of tissue engineering in space. Proc. Natl. Acad. Sci. USA 1997, 94, 13380–13382. [Google Scholar] [CrossRef] [Green Version]
- Stamenkovic, V.; Keller, G.; Nesic, D.; Cogoli, A.; Grogan, S.P. Neocartilage formation in 1 g, simulated, and microgravity environments: Implications for tissue engineering. Tissue Eng. Part A 2010, 16, 1729–1736. [Google Scholar] [CrossRef]
- Borst, A.G.; van Loon, J.J.W.A. Technology and Developments for the Random Positioning Machine, RPM. Microgravity Sci. Technol. 2008, 21, 287. [Google Scholar] [CrossRef]
- Kruger, M.; Melnik, D.; Kopp, S.; Buken, C.; Sahana, J.; Bauer, J.; Wehland, M.; Hemmersbach, R.; Corydon, T.J.; Infanger, M.; et al. Fighting Thyroid Cancer with Microgravity Research. Int. J. Mol. Sci. 2019, 20, 2553. [Google Scholar] [CrossRef] [Green Version]
- Kopp, S.; Warnke, E.; Wehland, M.; Aleshcheva, G.; Magnusson, N.E.; Hemmersbach, R.; Corydon, T.J.; Bauer, J.; Infanger, M.; Grimm, D. Mechanisms of three-dimensional growth of thyroid cells during long-term simulated microgravity. Sci. Rep. 2015, 5, 16691. [Google Scholar] [CrossRef] [Green Version]
- Kopp, S.; Slumstrup, L.; Corydon, T.J.; Sahana, J.; Aleshcheva, G.; Islam, T.; Magnusson, N.E.; Wehland, M.; Bauer, J.; Infanger, M.; et al. Identifications of novel mechanisms in breast cancer cells involving duct-like multicellular spheroid formation after exposure to the Random Positioning Machine. Sci. Rep. 2016, 6, 26887. [Google Scholar] [CrossRef]
- Ulbrich, C.; Westphal, K.; Pietsch, J.; Winkler, H.D.; Leder, A.; Bauer, J.; Kossmehl, P.; Grosse, J.; Schoenberger, J.; Infanger, M.; et al. Characterization of human chondrocytes exposed to simulated microgravity. Cell. Physiol. Biochem. 2010, 25, 551–560. [Google Scholar] [CrossRef]
- Grimm, D.; Infanger, M.; Westphal, K.; Ulbrich, C.; Pietsch, J.; Kossmehl, P.; Vadrucci, S.; Baatout, S.; Flick, B.; Paul, M.; et al. A delayed type of three-dimensional growth of human endothelial cells under simulated weightlessness. Tissue Eng. Part A 2009, 15, 2267–2275. [Google Scholar] [CrossRef]
- Stockwell, R.A.; Meachim, G. The chondrocytes. In Adult Articular Cartilage; Freeman, M.A.R., Ed.; Pitman Medical: London, UK, 1973; pp. 51–99. [Google Scholar]
- Dingle, J.T. The secretion of enzymes into the pericellular environment. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1975, 271, 315–324. [Google Scholar]
- Stockwell, R.A. Chondrocytes. Suppl. (R. Coll. Pathol.) 1978, 12, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Meachim, G.; Roy, S. Intracytoplasmic filaments in the cells of adult human articular cartilage. Ann. Rheum. Dis. 1967, 26, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, H.; Bischoff, R.; Holtzer, H. Formation of arrowhead complexes with heavy meromyosin in a variety of cell types. J. Cell Biol. 1969, 43, 312–328. [Google Scholar] [CrossRef]
- Bush, P.G.; Hoemann, C.D.; Hall, A.C. A Similarity between Morphology of Chondrocytes within Degenerate Cartilage and Those Cultured in a Weak 3D Matrix. In Proceedings of the 47th Annual Meeting of the Orthopaedic Research Society, San Francisco, CA, USA, 25–28 February 2001; Volume 26, p. 0661. [Google Scholar]
- Canty, E.G.; Kadler, K.E. Procollagen trafficking, processing and fibrillogenesis. J. Cell Sci. 2005, 118 Pt 7, 1341–1353. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.E.; Burridge, K. Transmembrane molecular assemblies in cell-extracellular matrix interactions. Curr. Opin. Cell Biol. 1991, 3, 849–853. [Google Scholar] [CrossRef]
- Sims, J.R.; Karp, S.; Ingber, D.E. Altering the cellular mechanical force balance results in integrated changes in cell, cytoskeletal and nuclear shape. J. Cell Sci. 1992, 103 Pt 4, 1215–1222. [Google Scholar]
- Juliano, R.L.; Haskill, S. Signal transduction from the extracellular matrix. J. Cell Biol. 1993, 120, 577–585. [Google Scholar] [CrossRef]
- Simon, V.R.; Pon, L.A. Actin-based organelle movement. Experientia 1996, 52, 1117–1122. [Google Scholar] [CrossRef]
- Riezman, H.; Munn, A.; Geli, M.I.; Hicke, L. Actin-, myosin- and ubiquitin-dependent endocytosis. Experientia 1996, 52, 1033–1041. [Google Scholar] [CrossRef]
- Vorselen, D.; Roos, W.H.; MacKintosh, F.C.; Wuite, G.J.; van Loon, J.J. The role of the cytoskeleton in sensing changes in gravity by nonspecialized cells. FASEB J. 2014, 28, 536–547. [Google Scholar] [CrossRef] [Green Version]
- Vale, R.D. Intracellular transport using microtubule-based motors. Annu. Rev. Cell Biol. 1987, 3, 347–378. [Google Scholar] [CrossRef]
- Mitchison, T.; Evans, L.; Schulze, E.; Kirschner, M. Sites of microtubule assembly and disassembly in the mitotic spindle. Cell 1986, 45, 515–527. [Google Scholar] [CrossRef]
- Gibbons, I.R. Cilia and flagella of eukaryotes. J. Cell Biol. 1981, 91 Pt 2, 107s–124s. [Google Scholar] [CrossRef]
- Farquharson, C.; Lester, D.; Seawright, E.; Jefferies, D.; Houston, B. Microtubules are potential regulators of growth-plate chondrocyte differentiation and hypertrophy. Bone 1999, 25, 405–412. [Google Scholar] [CrossRef]
- Lazarides, E. Intermediate filaments as mechanical integrators of cellular space. Nature 1980, 283, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Traub, P. Intermediate filaments and gene regulation. Physiol. Chem. Phys. Med. NMR 1995, 27, 377–400. [Google Scholar] [PubMed]
- Aubin, J.E.; Liu, F.; Malaval, L.; Gupta, A.K. Osteoblast and chondroblast differentiation. Bone 1995, 17 (Suppl. S2), 77s–83s. [Google Scholar] [CrossRef]
- Johnstone, B.; Hering, T.M.; Caplan, A.I.; Goldberg, V.M.; Yoo, J.U. In vitro chondrogenesis of bone marrow-derived mesenchymal progenitor cells. Exp. Cell Res. 1998, 238, 265–272. [Google Scholar] [CrossRef]
- Temenoff, J.S.; Mikos, A.G. Review: Tissue engineering for regeneration of articular cartilage. Biomaterials 2000, 21, 431–440. [Google Scholar] [CrossRef]
- Stockwell, R.A. The cell density of human articular and costal cartilage. J. Anat. 1967, 101 Pt 4, 753–763. [Google Scholar]
- Buckwalter, J.A.; Mankin, H.J. Articular cartilage: Tissue design and chondrocyte-matrix interactions. Instr. Course Lect. 1998, 47, 477–486. [Google Scholar]
- Nakamura, S.; Arai, Y.; Takahashi, K.A.; Terauchi, R.; Ohashi, S.; Mazda, O.; Imanishi, J.; Inoue, A.; Tonomura, H.; Kubo, T. Hydrostatic pressure induces apoptosis of chondrocytes cultured in alginate beads. J. Orthop. Res. 2006, 24, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Akmal, M.; Anand, A.; Anand, B.; Wiseman, M.; Goodship, A.E.; Bentley, G. The culture of articular chondrocytes in hydrogel constructs within a bioreactor enhances cell proliferation and matrix synthesis. J. Bone Jt. Surg. Br. 2006, 88, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Buravkova, L.; Romanov, Y.; Rykova, M.; Grigorieva, O.; Merzlikina, N. Cell-to-cell interactions in changed gravity: Ground-based and flight experiments. Acta Astronaut. 2005, 57, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Cerwinka, W.H.; Sharp, S.M.; Boyan, B.D.; Zhau, H.E.; Chung, L.W.; Yates, C. Differentiation of human mesenchymal stem cell spheroids under microgravity conditions. Cell Regen. 2012, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Freed, L.E.; Langer, R.; Martin, I.; Pellis, N.R.; Vunjak-Novakovic, G. Tissue engineering of cartilage in space. Proc. Natl. Acad. Sci. USA 1997, 94, 13885–13890. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, S.; Satoh, T.; Chiba, J.; Ju, C.; Inoue, K.; Kagawa, J. Interleukin-6 and interleukin-8 levels in serum and synovial fluid of patients with osteoarthritis. Cytokines Cell. Mol. Ther. 2000, 6, 71–79. [Google Scholar] [CrossRef]
- Kamiya, N.; Kuroyanagi, G.; Aruwajoye, O.; Kim, H.K.W. IL6 receptor blockade preserves articular cartilage and increases bone volume following ischemic osteonecrosis in immature mice. Osteoarthr. Cartil. 2019, 27, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Aida, Y.; Honda, K.; Tanigawa, S.; Nakayama, G.; Matsumura, H.; Suzuki, N.; Shimizu, O.; Takeichi, O.; Makimura, M.; Maeno, M. IL-6 and soluble IL-6 receptor stimulate the production of MMPs and their inhibitors via JAK-STAT and ERK-MAPK signalling in human chondrocytes. Cell Biol. Int. 2012, 36, 367–376. [Google Scholar] [CrossRef]
- Dahlberg, L.; Billinghurst, R.C.; Manner, P.; Nelson, F.; Webb, G.; Ionescu, M.; Reiner, A.; Tanzer, M.; Zukor, D.; Chen, J.; et al. Selective enhancement of collagenase-mediated cleavage of resident type II collagen in cultured osteoarthritic cartilage and arrest with a synthetic inhibitor that spares collagenase 1 (matrix metalloproteinase 1). Arthritis Rheum. 2000, 43, 673–682. [Google Scholar] [CrossRef]
- Gao, S.G.; Li, K.H.; Zeng, K.B.; Tu, M.; Xu, M.; Lei, G.H. Elevated osteopontin level of synovial fluid and articular cartilage is associated with disease severity in knee osteoarthritis patients. Osteoarthr. Cartil. 2010, 18, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Honsawek, S.; Tanavalee, A.; Sakdinakiattikoon, M.; Chayanupatkul, M.; Yuktanandana, P. Correlation of plasma and synovial fluid osteopontin with disease severity in knee osteoarthritis. Clin. Biochem. 2009, 42, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Zhang, L.; Zhao, L.; Gao, S.; Han, R.; Su, D.; Lei, G. Phosphorylation of osteopontin in osteoarthritis degenerative cartilage and its effect on matrix metalloprotease 13. Rheumatol. Int. 2013, 33, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Pullig, O.; Weseloh, G.; Gauer, S.; Swoboda, B. Osteopontin is expressed by adult human osteoarthritic chondrocytes: Protein and mRNA analysis of normal and osteoarthritic cartilage. Matrix Biol. 2000, 19, 245–255. [Google Scholar] [CrossRef]
- Matsui, Y.; Iwasaki, N.; Kon, S.; Takahashi, D.; Morimoto, J.; Matsui, Y.; Denhardt, D.T.; Rittling, S.; Minami, A.; Uede, T. Accelerated development of aging-associated and instability-induced osteoarthritis in osteopontin-deficient mice. Arthritis Rheum. 2009, 60, 2362–2371. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Lefebvre, V. Unraveling the transcriptional regulatory machinery in chondrogenesis. J. Bone Miner. Metab. 2011, 29, 390–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozhemyakina, E.; Lassar, A.B.; Zelzer, E. A pathway to bone: Signaling molecules and transcription factors involved in chondrocyte development and maturation. Development 2015, 142, 817–831. [Google Scholar] [CrossRef] [Green Version]
- Oh, C.D.; Lu, Y.; Liang, S.; Mori-Akiyama, Y.; Chen, D.; de Crombrugghe, B.; Yasuda, H. SOX9 regulates multiple genes in chondrocytes, including genes encoding ECM proteins, ECM modification enzymes, receptors, and transporters. PLoS ONE 2014, 9, e107577. [Google Scholar] [CrossRef]
- Ohba, S.; He, X.; Hojo, H.; McMahon, A.P. Distinct Transcriptional Programs Underlie Sox9 Regulation of the Mammalian Chondrocyte. Cell Rep. 2015, 12, 229–243. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.F.; Lefebvre, V. The transcription factors SOX9 and SOX5/SOX6 cooperate genome-wide through super-enhancers to drive chondrogenesis. Nucleic Acids Res. 2015, 43, 8183–8203. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Wang, C.; Acton, A.J.; Eckert, G.J.; Trippel, S.B. Role of sox9 in growth factor regulation of articular chondrocytes. J. Cell. Biochem. 2015, 116, 1391–1400. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Zheng, Q.; Engin, F.; Munivez, E.; Chen, Y.; Sebald, E.; Krakow, D.; Lee, B. Dominance of SOX9 function over RUNX2 during skeletogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19004–19009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikegami, D.; Akiyama, H.; Suzuki, A.; Nakamura, T.; Nakano, T.; Yoshikawa, H.; Tsumaki, N. Sox9 sustains chondrocyte survival and hypertrophy in part through Pik3ca-Akt pathways. Development 2011, 138, 1507–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, H.; Enomoto-Iwamoto, M.; Iwamoto, M.; Nomura, S.; Himeno, M.; Kitamura, Y.; Kishimoto, T.; Komori, T. Cbfa1 is a positive regulatory factor in chondrocyte maturation. J. Biol. Chem. 2000, 275, 8695–8702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, S.; Bonnamy, J.P.; Owen, M.J.; Ducy, P.; Karsenty, G. Continuous expression of Cbfa1 in nonhypertrophic chondrocytes uncovers its ability to induce hypertrophic chondrocyte differentiation and partially rescues Cbfa1-deficient mice. Genes Dev. 2001, 15, 467–481. [Google Scholar] [CrossRef] [Green Version]
- Ueta, C.; Iwamoto, M.; Kanatani, N.; Yoshida, C.; Liu, Y.; Enomoto-Iwamoto, M.; Ohmori, T.; Enomoto, H.; Nakata, K.; Takada, K.; et al. Skeletal malformations caused by overexpression of Cbfa1 or its dominant negative form in chondrocytes. J. Cell Biol. 2001, 153, 87–100. [Google Scholar] [CrossRef]
- Jiménez, M.J.; Balbín, M.; López, J.M.; Alvarez, J.; Komori, T.; López-Otín, C. Collagenase 3 is a target of Cbfa1, a transcription factor of the runt gene family involved in bone formation. Mol. Cell. Biol. 1999, 19, 4431–4442. [Google Scholar] [CrossRef] [Green Version]
- Hess, J.; Porte, D.; Munz, C.; Angel, P. AP-1 and Cbfa/runt physically interact and regulate parathyroid hormone-dependent MMP13 expression in osteoblasts through a new osteoblast-specific element 2/AP-1 composite element. J. Biol. Chem. 2001, 276, 20029–20038. [Google Scholar] [CrossRef] [Green Version]
- Komori, T. Regulation of bone development and extracellular matrix protein genes by RUNX2. Cell Tissue Res. 2010, 339, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Morii, E.; Komori, T.; Kawahata, H.; Sugimoto, M.; Terai, K.; Shimizu, H.; Yasui, T.; Ogihara, H.; Yasui, N.; et al. Transcriptional regulation of osteopontin gene in vivo by PEBP2alphaA/CBFA1 and ETS1 in the skeletal tissues. Oncogene 1998, 17, 1517–1525. [Google Scholar] [CrossRef] [Green Version]
- Kamekura, S.; Kawasaki, Y.; Hoshi, K.; Shimoaka, T.; Chikuda, H.; Maruyama, Z.; Komori, T.; Sato, S.; Takeda, S.; Karsenty, G.; et al. Contribution of runt-related transcription factor 2 to the pathogenesis of osteoarthritis in mice after induction of knee joint instability. Arthritis Rheum. 2006, 54, 2462–2470. [Google Scholar] [CrossRef]
- Liao, L.; Zhang, S.; Gu, J.; Takarada, T.; Yoneda, Y.; Huang, J.; Zhao, L.; Oh, C.D.; Li, J.; Wang, B.; et al. Deletion of Runx2 in Articular Chondrocytes Decelerates the Progression of DMM-Induced Osteoarthritis in Adult Mice. Sci. Rep. 2017, 7, 2371. [Google Scholar] [CrossRef] [PubMed]
- Marlovits, S.; Tichy, B.; Truppe, M.; Gruber, D.; Schlegel, W. Collagen expression in tissue engineered cartilage of aged human articular chondrocytes in a rotating bioreactor. Int. J. Artif. Organs 2003, 26, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Aleshcheva, G.; Bauer, J.; Hemmersbach, R.; Egli, M.; Wehland, M.; Grimm, D. Tissue Engineering of Cartilage on Ground-Based Facilities. Microgravity Sci. Technol. 2016, 28, 237–245. [Google Scholar] [CrossRef]
- Grimm, D.; Wehland, M.; Corydon, T.J.; Richter, P.; Prasad, B.; Bauer, J.; Egli, M.; Kopp, S.; Lebert, M.; Krüger, M. The effects of microgravity on differentiation and cell growth in stem cells and cancer stem cells. Stem Cells Transl. Med. 2020, 9, 882–894. [Google Scholar] [CrossRef] [PubMed]
- Ohyabu, Y.; Kida, N.; Kojima, H.; Taguchi, T.; Tanaka, J.; Uemura, T. Cartilaginous tissue formation from bone marrow cells using rotating wall vessel (RWV) bioreactor. Biotechnol. Bioeng. 2006, 95, 1003–1008. [Google Scholar] [CrossRef]
- Luo, W.; Xiong, W.; Qiu, M.; Lv, Y.; Li, Y.; Li, F. Differentiation of mesenchymal stem cells towards a nucleus pulposus-like phenotype utilizing simulated microgravity In vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2011, 31, 199. [Google Scholar] [CrossRef]
- Yuge, L.; Kajiume, T.; Tahara, H.; Kawahara, Y.; Umeda, C.; Yoshimoto, R.; Wu, S.L.; Yamaoka, K.; Asashima, M.; Kataoka, K.; et al. Microgravity potentiates stem cell proliferation while sustaining the capability of differentiation. Stem Cells Dev. 2006, 15, 921–929. [Google Scholar] [CrossRef]
- Wu, X.; Li, S.H.; Lou, L.M.; Chen, Z.R. The effect of the microgravity rotating culture system on the chondrogenic differentiation of bone marrow mesenchymal stem cells. Mol. Biotechnol. 2013, 54, 331–336. [Google Scholar] [CrossRef]
- Mayer-Wagner, S.; Hammerschmid, F.; Redeker, J.I.; Schmitt, B.; Holzapfel, B.M.; Jansson, V.; Betz, O.B.; Müller, P.E. Simulated microgravity affects chondrogenesis and hypertrophy of human mesenchymal stem cells. Int. Orthop. 2014, 38, 2615–2621. [Google Scholar] [CrossRef]
- Grimm, D.; Wehland, M.; Pietsch, J.; Aleshcheva, G.; Wise, P.; van Loon, J.; Ulbrich, C.; Magnusson, N.E.; Infanger, M.; Bauer, J. Growing tissues in real and simulated microgravity: New methods for tissue engineering. Tissue Eng. Part B Rev. 2014, 20, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Krüger, M.; Pietsch, J.; Bauer, J.; Kopp, S.; Carvalho, D.T.O.; Baatout, S.; Moreels, M.; Melnik, D.; Wehland, M.; Egli, M.; et al. Growth of Endothelial Cells in Space and in Simulated Microgravity—A Comparison on the Secretory Level. Cell. Physiol. Biochem. 2019, 52, 1039–1060. [Google Scholar] [PubMed] [Green Version]
- Mann, V.; Grimm, D.; Corydon, T.J.; Krüger, M.; Wehland, M.; Riwaldt, S.; Sahana, J.; Kopp, S.; Bauer, J.; Reseland, J.E.; et al. Changes in Human Foetal Osteoblasts Exposed to the Random Positioning Machine and Bone Construct Tissue Engineering. Int. J. Mol. Sci. 2019, 20, 1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietsch, J.; Gass, S.; Nebuloni, S.; Echegoyen, D.; Riwaldt, S.; Baake, C.; Bauer, J.; Corydon, T.J.; Egli, M.; Infanger, M.; et al. Three-dimensional growth of human endothelial cells in an automated cell culture experiment container during the SpaceX CRS-8 ISS space mission—The SPHEROIDS project. Biomaterials 2017, 124, 126–156. [Google Scholar] [CrossRef] [PubMed]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-Based Facilities for Simulation of Microgravity: Organism-Specific Recommendations for Their Use, and Recommended Terminology. Astrobiology 2012, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated Microgravity: Critical Review on the Use of Random Positioning Machines for Mammalian Cell Culture. BioMed Res. Int. 2015, 2015, 971474. [Google Scholar] [CrossRef] [Green Version]
- Infanger, M.; Ulbrich, C.; Baatout, S.; Wehland, M.; Kreutz, R.; Bauer, J.; Grosse, J.; Vadrucci, S.; Cogoli, A.; Derradji, H.; et al. Modeled gravitational unloading induced downregulation of endothelin-1 in human endothelial cells. J. Cell. Biochem. 2007, 101, 1439–1455. [Google Scholar] [CrossRef]
- Riecke, K.; Grimm, D.; Shakibaei, M.; Kossmehl, P.; Schulze-Tanzil, G.; Paul, M.; Stahlmann, R. Low doses of 2,3,7,8-tetrachlorodibenzo- p-dioxin increase transforming growth factor beta and cause myocardial fibrosis in marmosets (Callithrix jacchus). Arch. Toxicol. 2002, 76, 360–366. [Google Scholar] [CrossRef]
- Rothermund, L.; Kreutz, R.; Kossmehl, P.; Fredersdorf, S.; Shakibaei, M.; Schulze-Tanzil, G.; Paul, M.; Grimm, D. Early onset of chondroitin sulfate and osteopontin expression in angiotensin II-dependent left ventricular hypertrophy. Am. J. Hypertens. 2002, 15 Pt 1, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Grimm, D.; Bauer, J.; Ulbrich, C.; Westphal, K.; Wehland, M.; Infanger, M.; Aleshcheva, G.; Pietsch, J.; Ghardi, M.; Beck, M.; et al. Different responsiveness of endothelial cells to vascular endothelial growth factor and basic fibroblast growth factor added to culture media under gravity and simulated microgravity. Tissue Eng. Part A 2010, 16, 1559–1573. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| 18S rRNA | GGAGCCTGCGGCTTAATTT | CAACTAAGAACGGCCATGCA |
| ACAN | AGTCCAACTCTTCAAGGTGAACTA | ACTCAGCGAGTTGTCATGGT |
| COL1A1 | ACGAAGACATCCCACCAATCAC | CGTTGTCGCAGACGCAGAT |
| COL2A1 | GGCAATAGCAGGTTCACGTACA | CGATAACAGTCTTGCCCCACTT |
| COL10A1 | GGGCAGAGGAAGCTTCAGAAA | TCTCAGATGGATTCTGCGTGC |
| IL6 | CGGGAACGAAAGAGAAGCTCTA | GAGCAGCCCCAGGGAGAA |
| CXCL8 | TGGCAGCCTTCCTGATTTCT | GGGTGGAAAGGTTTGGAGTATG |
| ITGB1 | GAAAACAGCGCATATCTGGAAATT | CAGCCAATCAGTGATCCACAA |
| MMP1 | GTCAGGGGAGATCATCGGG | GAGCATCCCCTCCAATACCTG |
| MMP2 | CCATGATGGAGAGGCAGACA | CCATGATGGAGAGGCAGACA |
| MMP13 | AGCCTTCAAAGTTTGGTCCGA | TCGCCATGCTCCTTAATTCCA |
| PRG4 | CCCCCAAACCACCAGTTGTA | ACGTGTCAGGAGTTGTGACC |
| RUNX2 | GAACCCAGAAGGCACAGACA | GGATGAGGAATGCGCCCTAA |
| RUNX3 | GTGGGCGAGGGAAGAGTTTC | CCTTGATGGCTCGGTGGTAG |
| SOD3 | CTGGAAAGGTGCCCGACTCC | ATGTCTCGGATCCACTCCGC |
| SOX5 | TCCTCCCTCCAGGCTTCAG | CTGCCATGGTAGTTGGGATCA |
| SOX6 | GCCACACATTAAGCG | TCCAGCGAGATCCTAAGATTTTG |
| SOX9 | AGGAAGTCGGTGAAGAACGG | CGCCTTGAAGATGGCGTTG |
| SPP1 | CGAGGTGATAGTGTGGTTTATGGA | CGTCTGTAGCATCAGGGTACTG |
| TGFB1 | CACCCGCGTGCTAATGGT | AGAGCAACACGGGTTCAGGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wehland, M.; Steinwerth, P.; Aleshcheva, G.; Sahana, J.; Hemmersbach, R.; Lützenberg, R.; Kopp, S.; Infanger, M.; Grimm, D. Tissue Engineering of Cartilage Using a Random Positioning Machine. Int. J. Mol. Sci. 2020, 21, 9596. https://doi.org/10.3390/ijms21249596
Wehland M, Steinwerth P, Aleshcheva G, Sahana J, Hemmersbach R, Lützenberg R, Kopp S, Infanger M, Grimm D. Tissue Engineering of Cartilage Using a Random Positioning Machine. International Journal of Molecular Sciences. 2020; 21(24):9596. https://doi.org/10.3390/ijms21249596
Chicago/Turabian StyleWehland, Markus, Paul Steinwerth, Ganna Aleshcheva, Jayashree Sahana, Ruth Hemmersbach, Ronald Lützenberg, Sascha Kopp, Manfred Infanger, and Daniela Grimm. 2020. "Tissue Engineering of Cartilage Using a Random Positioning Machine" International Journal of Molecular Sciences 21, no. 24: 9596. https://doi.org/10.3390/ijms21249596