Obese Mice with Dyslipidemia Exhibit Meibomian Gland Hypertrophy and Alterations in Meibum Composition and Aqueous Tear Production

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
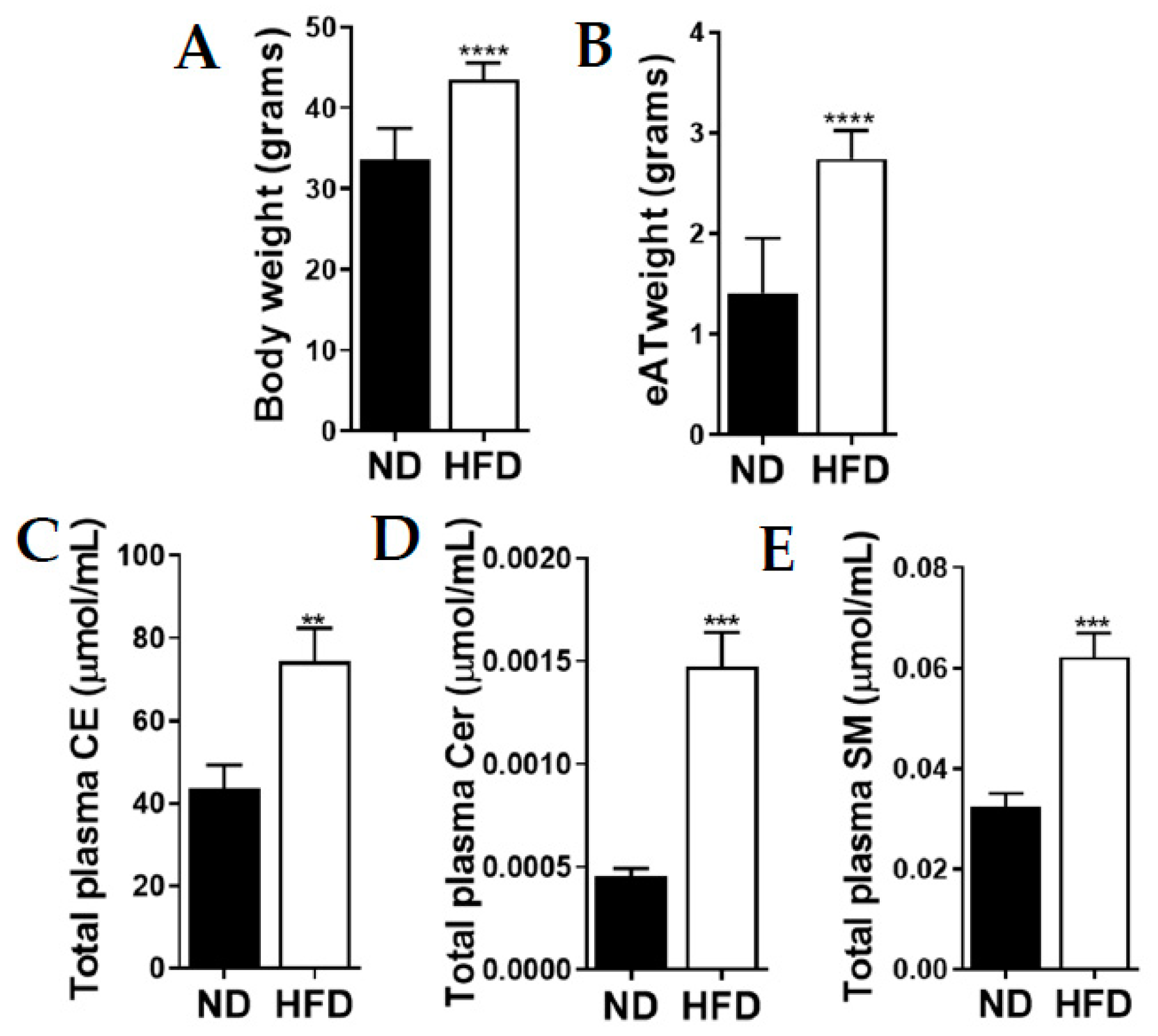
2.1. Mice Fed an Obesogenic Diet Gained More Weight with Elevated Total Plasma Lipid Levels
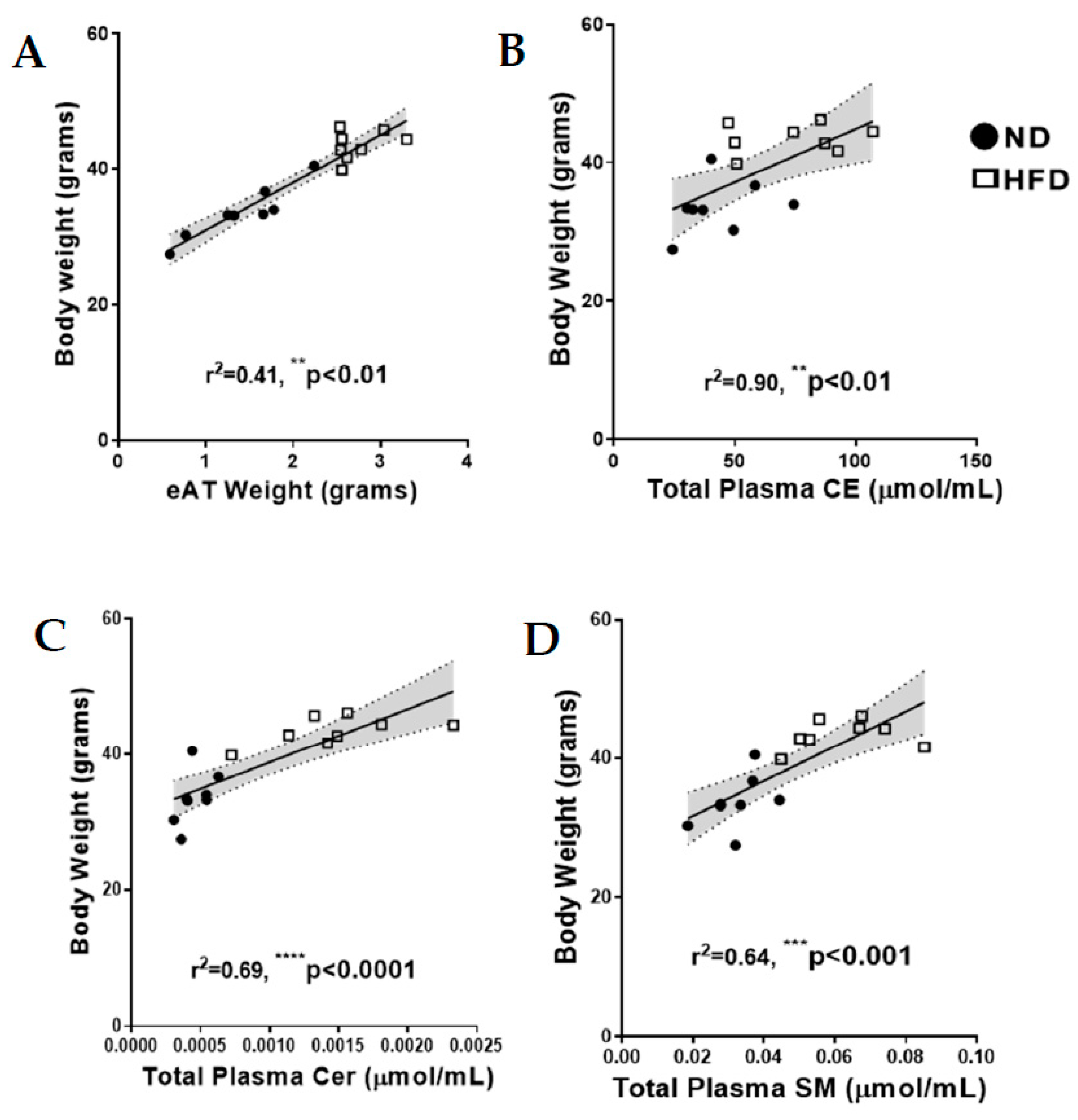
2.2. Relationship between Mouse Body Weight, eAT and Total Fasting Plasma Lipid Levels
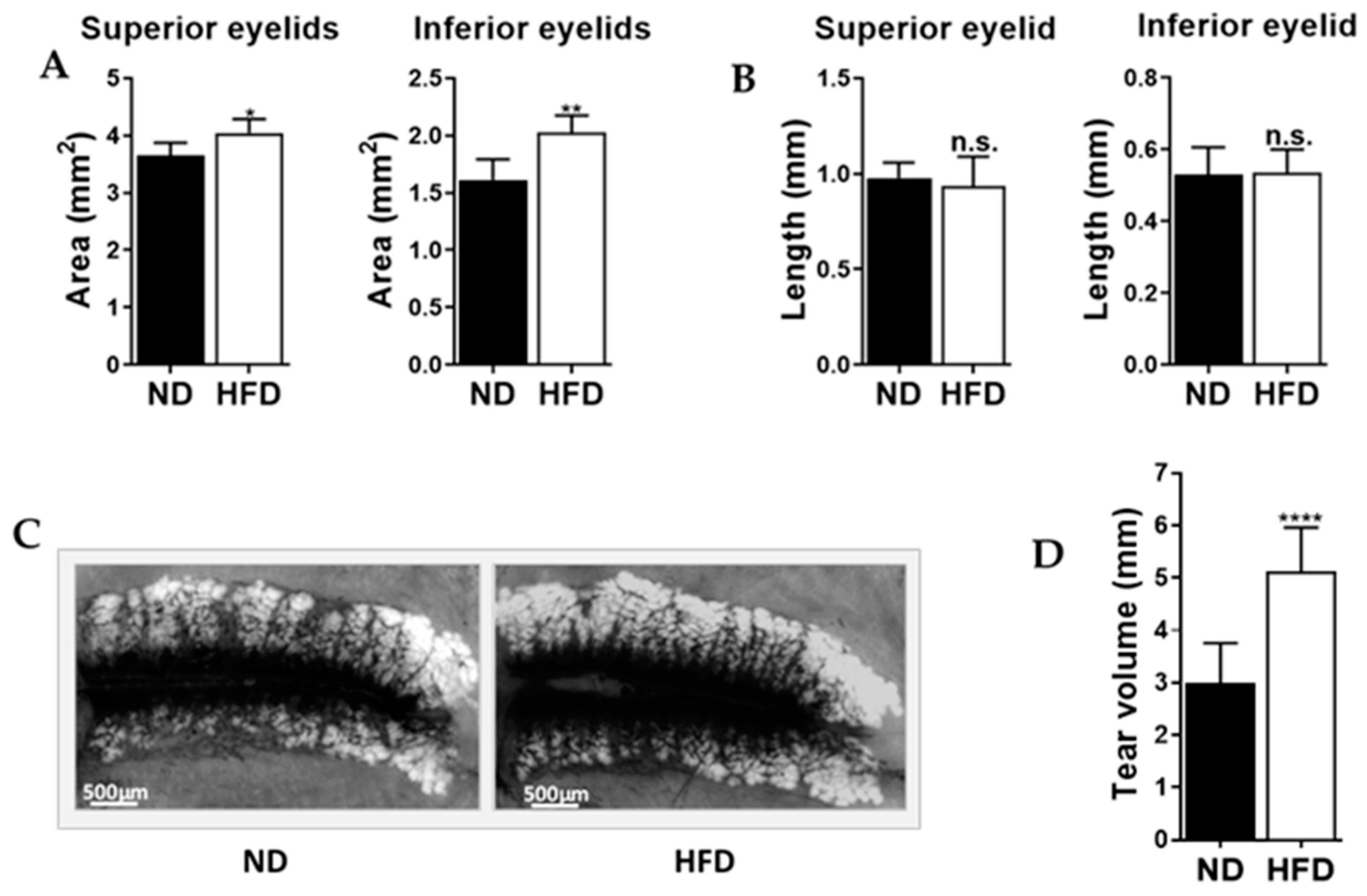
2.3. Mice Fed an Obesogenic Diet Showed Meibomian Gland Hypertrophy and Excessive Tear Production
2.4. Similarities and Differences between Plasma and Meibum Lipid Species
2.5. Relative Abudance of the Lipid Species within the Various Lipid Classes in Plasma and Meibum
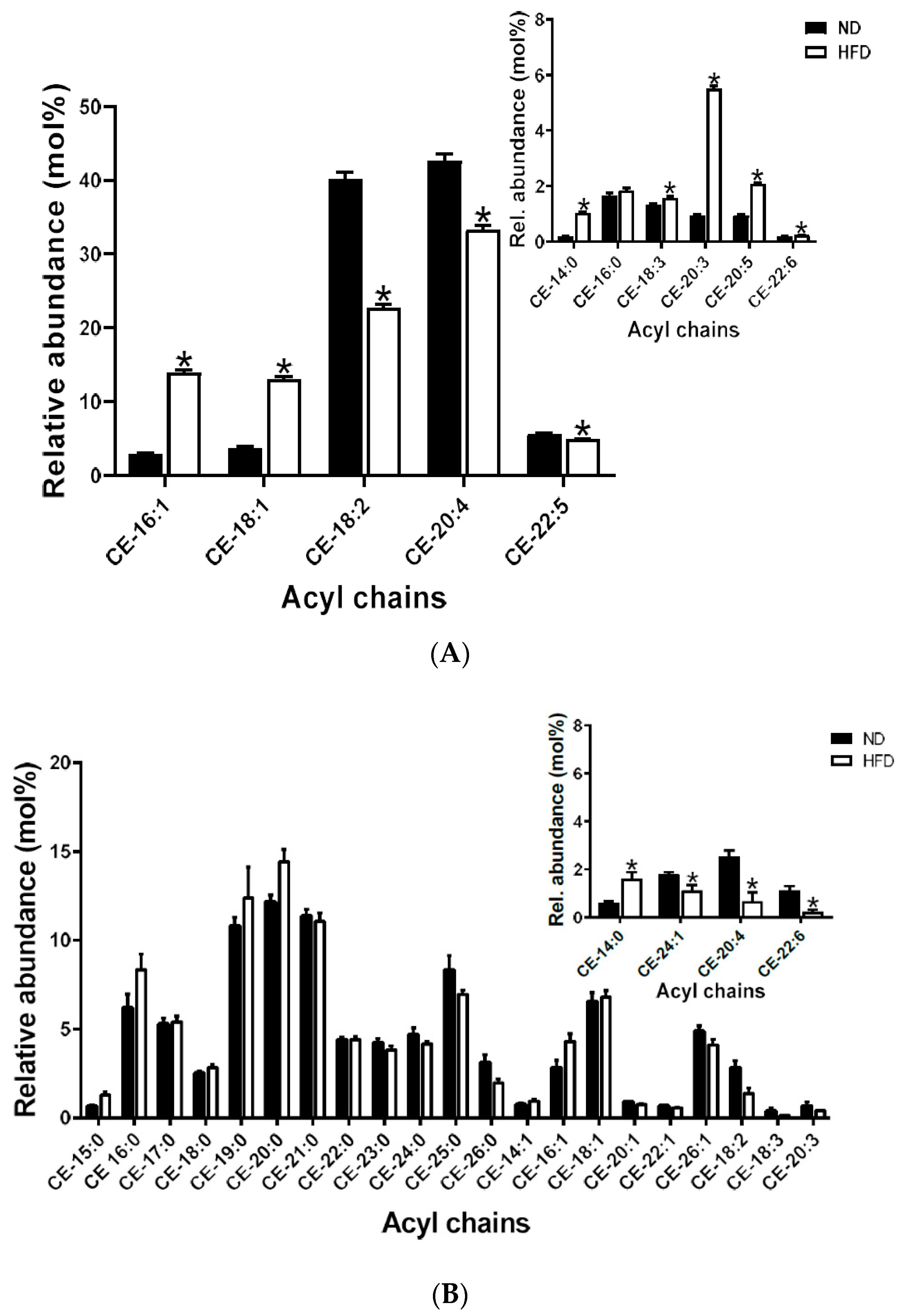
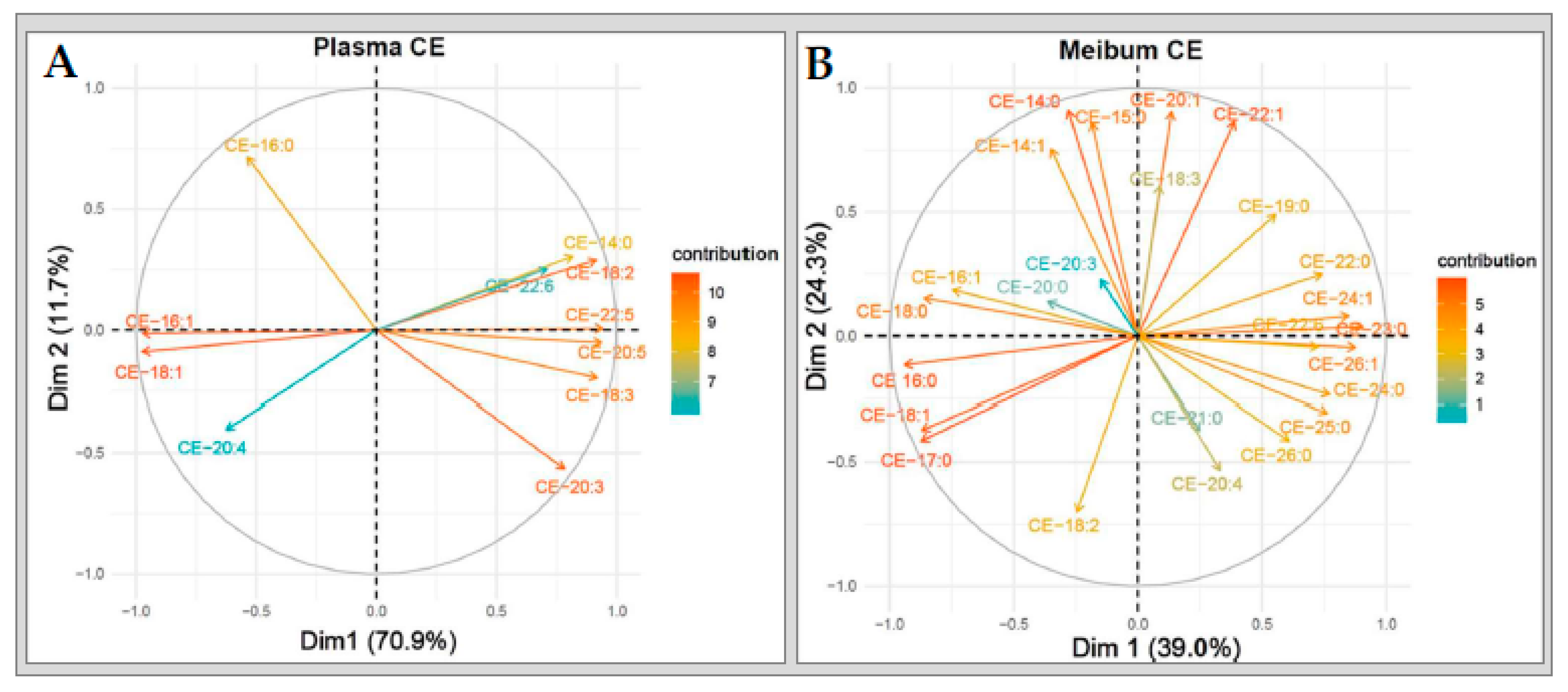
2.5.1. Cholesteryl Esters
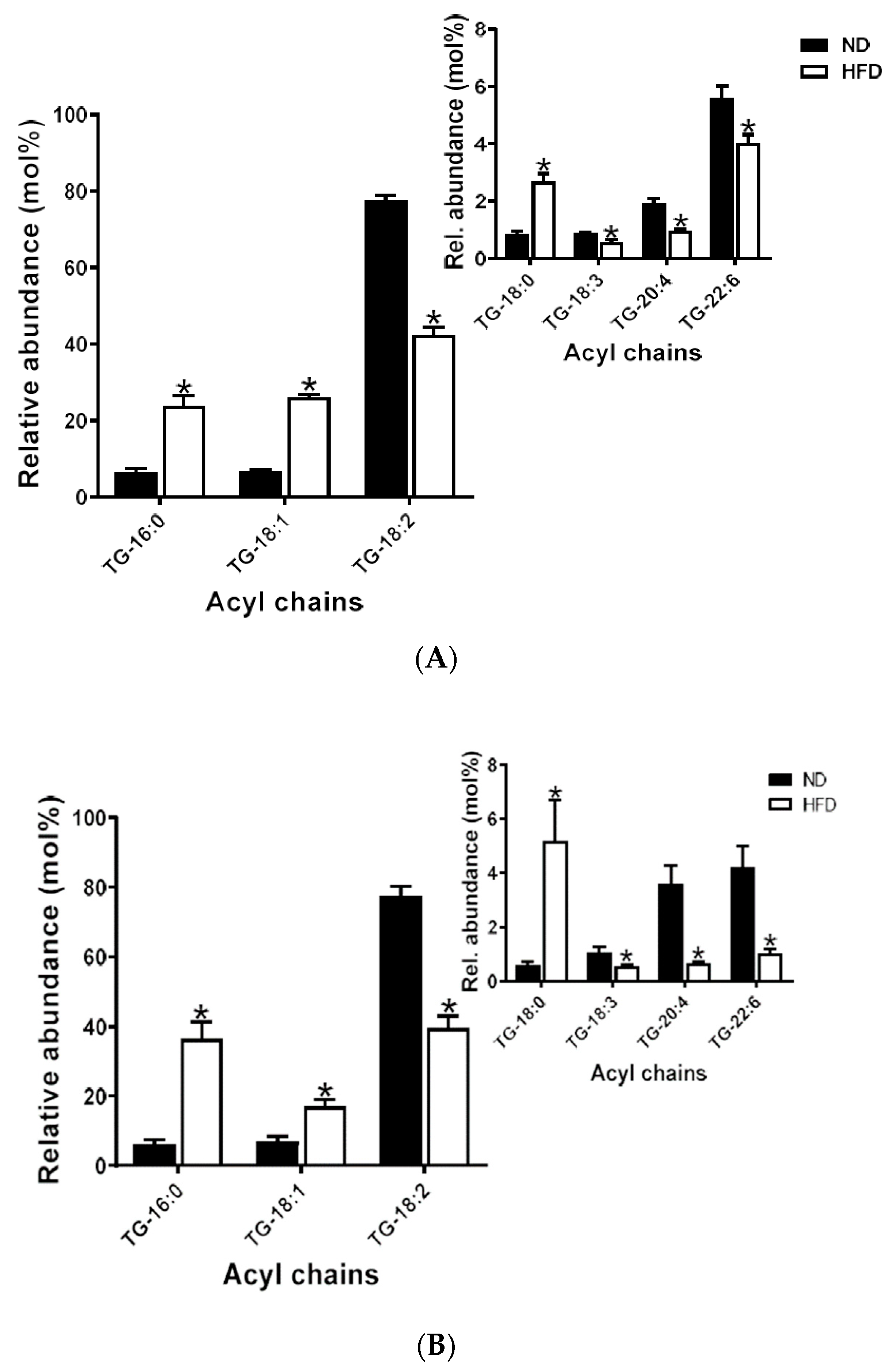
2.5.2. Triglycerides
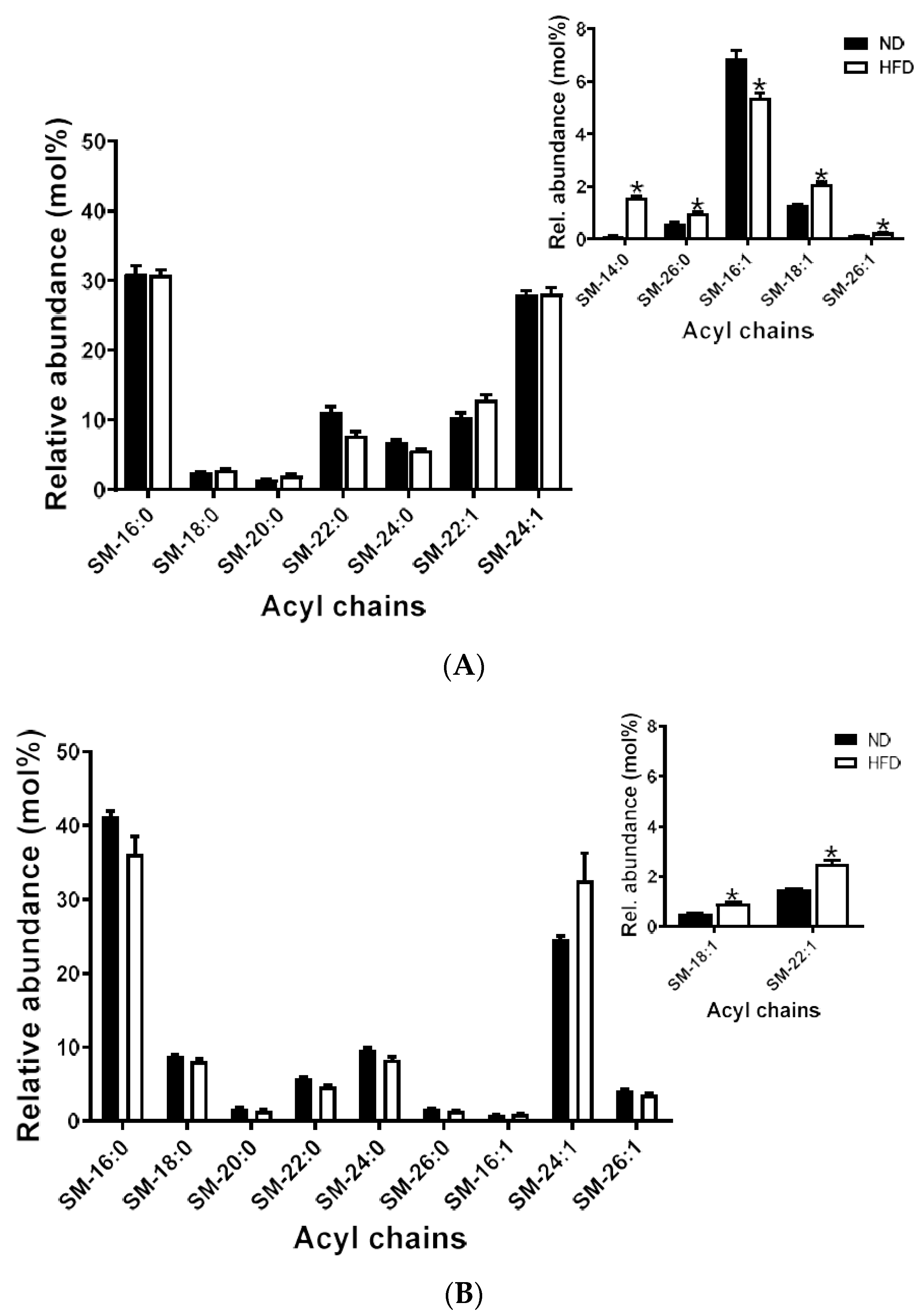
2.5.3. Sphingomyelins
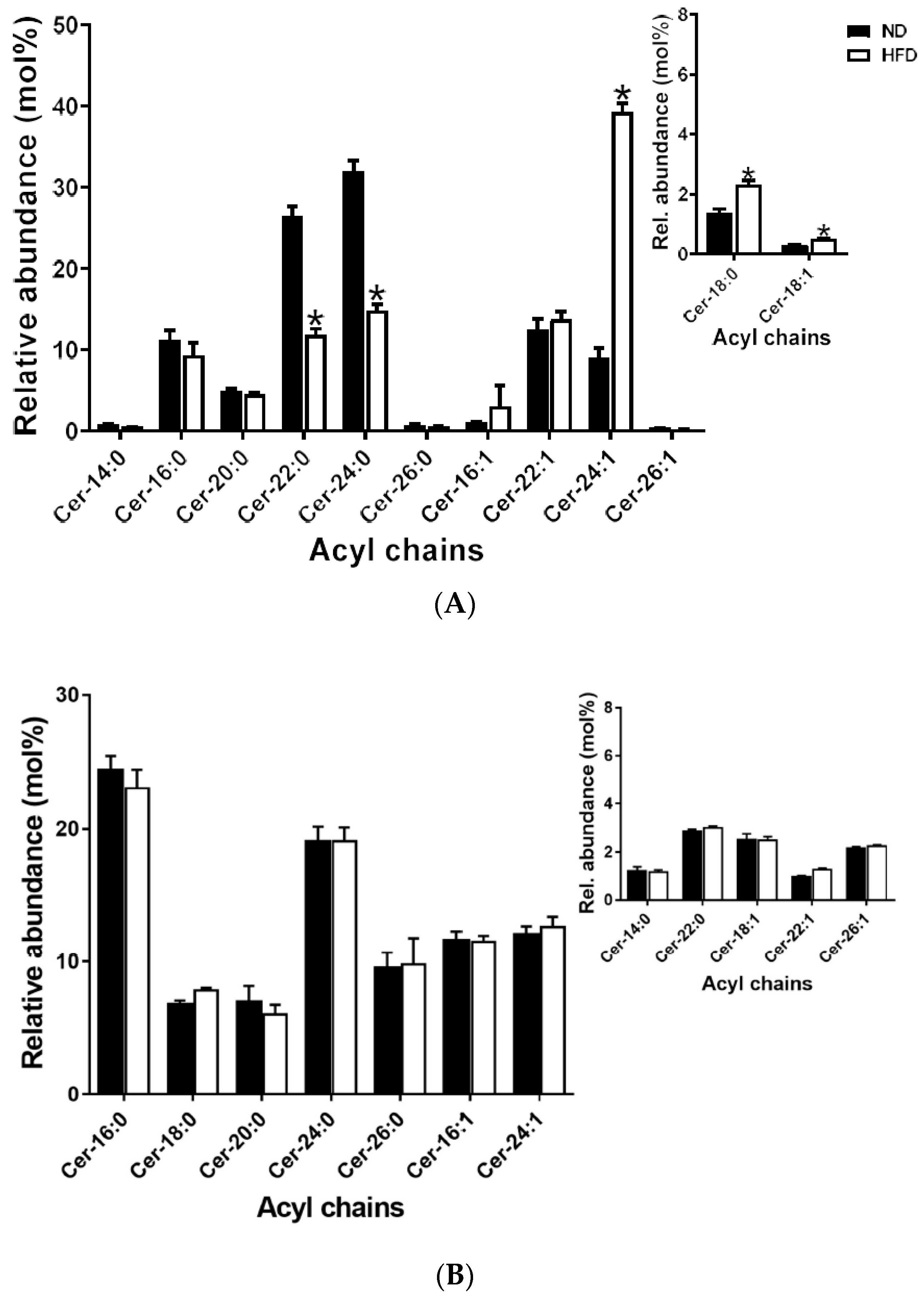
2.5.4. Ceramides
2.6. Patterns of Variation in the Different the Lipid Species within the Vairous Lipid Classes in Plasma and Meibum
2.6.1. Cholesteryl Esters
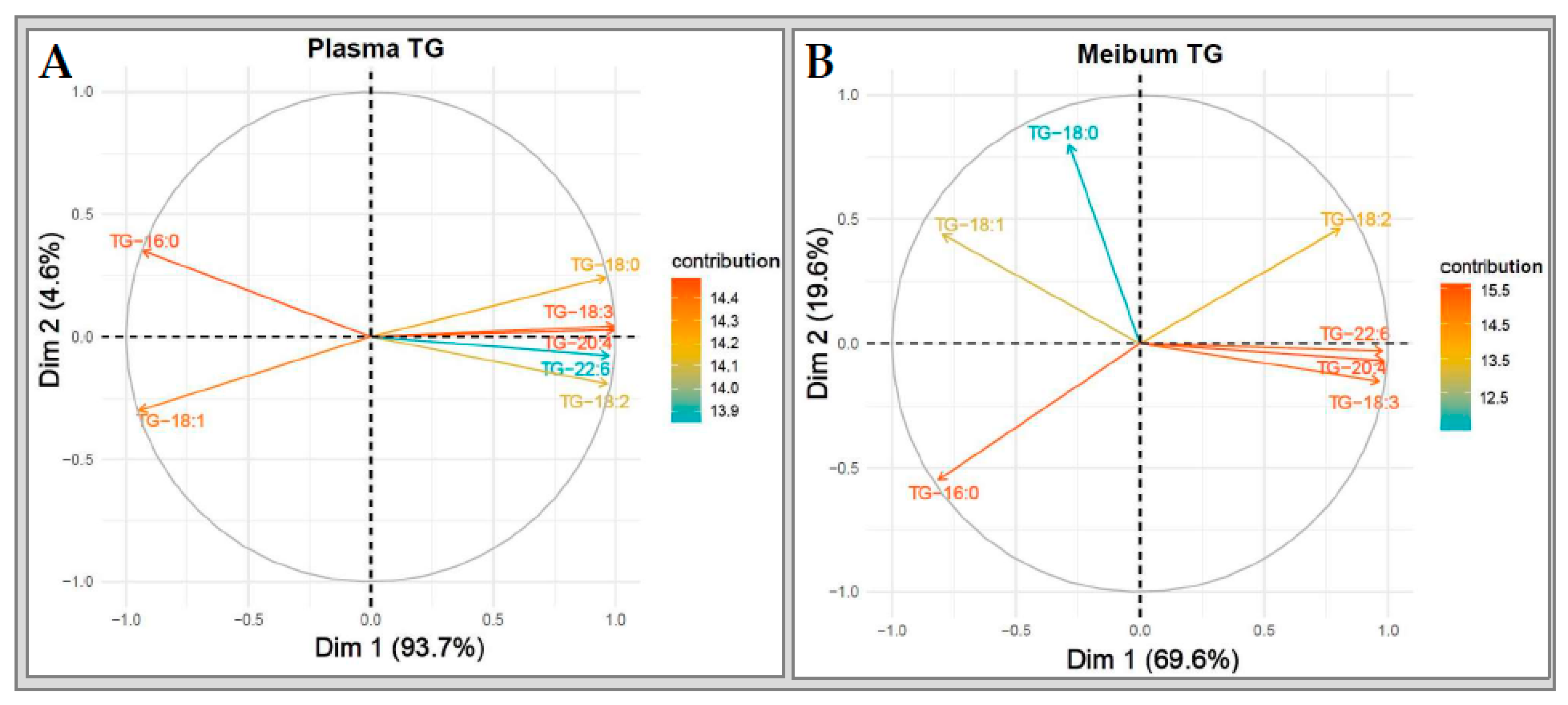
2.6.2. Triglycerides
2.6.3. Sphingomyelins and Ceramides
3. Discussion
4. Methods
4.1. Ethical Cosniderations
4.2. Mouse Model, Tear Production Assessments, Meibography and Sample Collection
4.3. Lipidomic Analyses of Plasma and Meibum Samples
4.3.1. Analysis of Cholesteryl Ester and Triglycerides Species
4.3.2. Analysis of Ceramides and Sphingomyelins
4.4. Nomenclature and Presentation of Lipid Quantities
4.5. Image and Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| eAT | Epididymal adipose tissue |
| CE | Cholesteryl ester |
| Cer | Ceramide |
| HFD | High-fat diet |
| MGD | Meibomian gland dysfunction |
| MUFAs | Monounsaturated fatty acids |
| ND | Normal diet |
| PUFAs | Polyunsaturated fatty acids |
| SM | Sphingomyelins |
| TG | Triglcyerides |
Appendix A. Analysis of Meibographs

- Meibomian gland tissue.
- Pigmentation—little darker spots sometimes on the glandular tissue and very dense along the lid margins.
- Background—includes areas without glands.

Appendix B. Principal Component Analysis Using Custom R Code
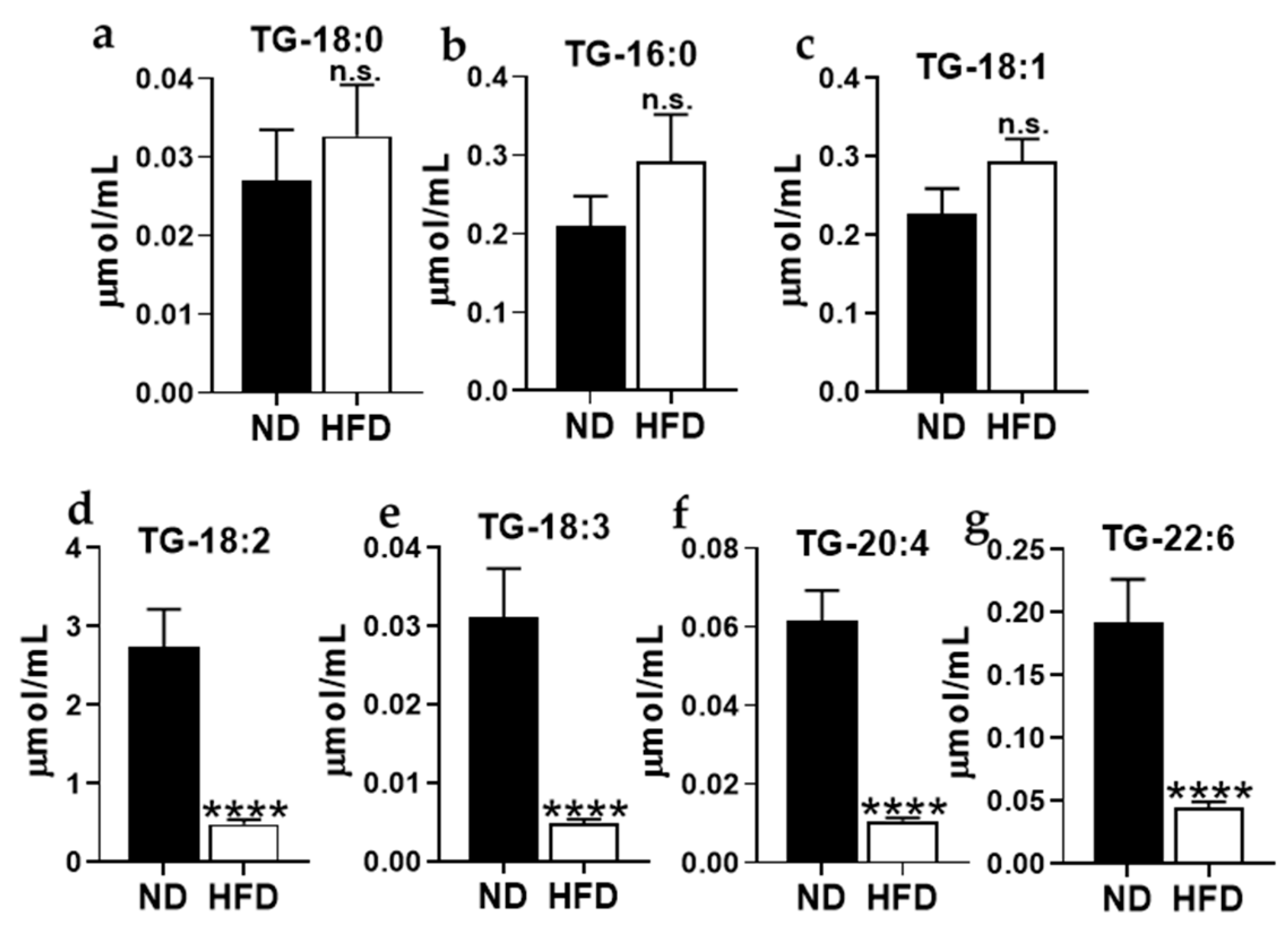
Appendix C. Total Fasting Levels of Various Plasma Triglycerides Species

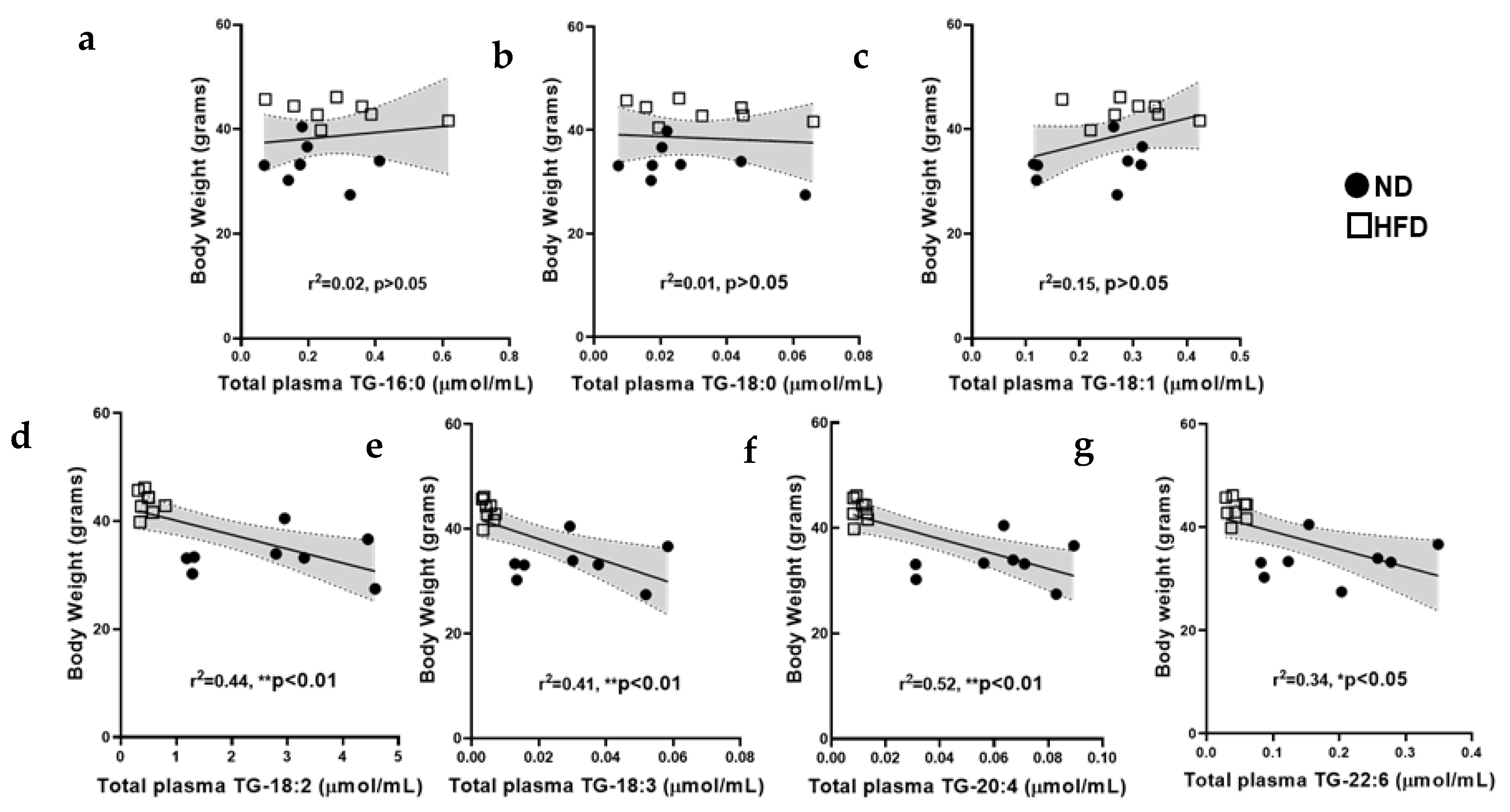
Appendix D. Correlations between Mouse Body Weight and Total Levels of Plasma Triglycerides Species

References
- Stein, C.J.; Colditz, G.A. The Epidemic of Obesity. J. Clin. Endocrinol. Metab. 2004, 89, 2522–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.C.; Lawlor, D.A.; Kimm, S.Y. Childhood obesity. Lancet 2010, 375, 1737–1748. [Google Scholar] [CrossRef]
- Nathan, B.M.; Moran, A. Metabolic complications of obesity in childhood and adolescence: More than just diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2008, 15, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Borlaug, B.A.; Reddy, Y.N. Getting at the heart of central obesity and the metabolic syndrome. Am. Heart Assoc. 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Office of the General Surgeon of the United States; The Offcie of Disease Prevention; Health Promotion; Centers for Disease Control and Prevention. The Surgeon General’s Call to Action to Prevent and Decrease Overweight and Obesity; US Government Printing Office: Washington, DC, USA, 2001.
- Himes, R.W.; Smith, C.W. Tlr2 is critical for diet-induced metabolic syndrome in a murine model. FASEB J. 2010, 24, 731–739. [Google Scholar] [CrossRef] [Green Version]
- Mensah, G.A.; Mokdad, A.H.; Ford, E.; Narayan, K.V.; Giles, W.H.; Vinicor, F.; Deedwania, P.C. Obesity, metabolic syndrome, and type 2 diabetes: Emerging epidemics and their cardiovascular implications. Cardiol. Clin. 2004, 22, 485–504. [Google Scholar] [CrossRef]
- Pietiläinen, K.H.; Sysi-Aho, M.; Rissanen, A.; Seppänen-Laakso, T.; Yki-Jarvinen, H.; Kaprio, J.; Orešič, M. Acquired obesity is associated with changes in the serum lipidomic profile independent of genetic effects–a monozygotic twin study. PLoS ONE 2007, 2, e218. [Google Scholar] [CrossRef]
- Burkhardt, R. Hyperlipidemia and cardiovascular disease: New insights on lipoprotein (a). Curr. Opin. Lipidol. 2019, 30, 260–261. [Google Scholar] [CrossRef]
- Rodwell, V.W.; Bender, D.A.; Botham, K.M.; Kennelly, P.J.; Weil, P.A. Harper’s Illustrated Biochemistry, 30th ed.; McGraw-Hill Education: New York, NY, USA, 2015. [Google Scholar]
- Kuriakose, R.K.; Braich, P.S. Dyslipidemia and its association with meibomian gland dysfunction: A systematic review. Int. Ophthalmol. 2018, 38, 1809–1816. [Google Scholar] [CrossRef]
- Osae, E.A.; Steven, P.; Redfern, R.; Hanlon, S.; Smith, C.W.; Rumbaut, R.E.; Burns, A. Dyslipidemia and Meibomian Gland Dysfunction: Utility of Lipidomics and Experimental Prospects with a Diet-Induced Obesity Mouse Model. Int. J. Mol. Sci. 2019, 20, 3505. [Google Scholar] [CrossRef] [Green Version]
- Knop, E.; Knop, N.; Millar, T.; Obata, H.; Sullivan, D.A. The international workshop on meibomian gland dysfunction: Report of the subcommittee on anatomy, physiology, and pathophysiology of the meibomian gland. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1938–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, T.J.; Schuett, B.S. The real reason for having a meibomian lipid layer covering the outer surface of the tear film—A review. Exp. Eye Res. 2015, 137, 125–138. [Google Scholar] [CrossRef]
- Mudgil, P. Antimicrobial role of human meibomian lipids at the ocular surface. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A. On the lipid composition of human meibum and tears: Comparative analysis of nonpolar lipids. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3779–3789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A. Fatty acid composition of cholesteryl esters of human meibomian gland secretions. Steroids 2010, 75, 726–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A. Lipidomics of human meibomian gland secretions: Chemistry, biophysics, and physiological role of meibomian lipids. Prog. Lipid Res. 2011, 50, 278–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green-Church, K.B.; Butovich, I.; Willcox, M.; Borchman, D.; Paulsen, F.; Barabino, S.; Glasgow, B.J. The international workshop on meibomian gland dysfunction: Report of the subcommittee on tear film lipids and lipid–protein interactions in health and disease. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1979–1993. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A.; Lu, H.; McMahon, A.; Eule, J.C. Toward an animal model of the human tear film: Biochemical comparison of the mouse, canine, rabbit, and human meibomian lipidomes. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6881–6896. [Google Scholar] [CrossRef]
- Sandra, K.; dos Santos Pereira, A.; Vanhoenacker, G.; David, F.; Sandra, P. Comprehensive blood plasma lipidomics by liquid chromatography/quadrupole time-of-flight mass spectrometry. J. Chromatogr. A 2010, 1217, 4087–4099. [Google Scholar] [CrossRef]
- Van Ginneken, V.; Verhey, E.; Poelmann, R.; Ramakers, R.; Van Dijk, K.W.; Ham, L.; Voshol, P.; Havekes, L.; Van Eck, M.; Van Der Greef, J. Metabolomics (liver and blood profiling) in a mouse model in response to fasting: A study of hepatic steatosis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2007, 1771, 1263–1270. [Google Scholar] [CrossRef]
- Shine, W.E.; Silvany, R.; McCulley, J.P. Relation of cholesterol-stimulated Staphylococcus aureus growth to chronic blepharitis. Investig. Ophthalmol. Vis. Sci. 1993, 34, 2291–2296. [Google Scholar]
- McCulley, J.P.; Shine, W.E. Meibomian secretions in chronic blepharitis. Adv. Exp. Med. Biol. 1998, 438, 319–326. [Google Scholar] [PubMed]
- Van Haeringen, N.; Glasius, E. Cholesterol in human tear fluid. Exp. Eye Res. 1975, 20, 271–274. [Google Scholar] [CrossRef]
- Hargrave, A.; Mehta, P.; Landry, P.; Amanda, H.; Dupre, M.; Magadi, S.; Li, Z.; Smith, C.W.; Burns, A.R. Metabolic syndrome affects mouse corneal epithelium and nerve morphology. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3076. [Google Scholar]
- Özbalci, C.; Sachsenheimer, T.; Brügger, B. Quantitative analysis of cellular lipids by nano-electrospray ionization mass spectrometry. In Membrane Biogenesis; Springer: Totota, NJ, USA, 2013; pp. 3–20. [Google Scholar]
- Voet, D.; Voet, J.G.; Pratt, C.W. Fundamentals of biochemistry. In Life at the Molecular Level, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Poutahidis, T.; Kleinewietfeld, M.; Smillie, C.; Levkovich, T.; Perrotta, A.; Bhela, S.; Varian, B.J.; Ibrahim, Y.M.; Lakritz, J.R.; Kearney, S.M.; et al. Microbial reprogramming inhibits Western diet-associated obesity. PLoS ONE 2013, 8, e68596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Vedova, M.C.; Muñoz, M.D.; Santillan, L.D.; Plateo-Pignatari, M.G.; Germanó, M.J.; Tosi, M.E.R.; Garcia, S.; Gomez, N.N.; Fornes, M.W.; Mejiba, S.E.G.; et al. A mouse model of diet-induced obesity resembling most features of human metabolic syndrome. Nutr. Metab. Insights 2016, 9. [Google Scholar] [CrossRef] [Green Version]
- Liou, A.P.; Paziuk, M.; Luevano, J.-M.; Machineni, S.; Turnbaugh, P.J.; Kaplan, L.M. Conserved shifts in the gut microbiota due to gastric bypass reduce host weight and adiposity. Sci. Transl. Med. 2013, 5, 178ra41. [Google Scholar] [CrossRef] [Green Version]
- Lai, S.-W.; Ng, K.; Lin, H.-F.; Chen, H.-L. Association between obesity and hyperlipidemia among children. Yale J. Biol. Med. 2001, 74, 205. [Google Scholar]
- Jung, U.J.; Choi, M.-S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [Green Version]
- Reins, R.Y.; Lema, C.; Courson, J.; Kunnen, C.M.; Redfern, R.L. MyD88 deficiency protects against dry eye–induced damage. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2967–2976. [Google Scholar] [CrossRef] [Green Version]
- Miyake, H.; Oda, T.; Katsuta, O.; Seno, M.; Nakamura, M. A novel model of Meibomian gland dysfunction induced with complete Freund’s adjuvant in rabbits. Vision 2017, 1, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, N.; Saban, D.R. Pathogenesis of meibomian gland dysfunction (MGD) requires the T cell-neutrophil axis, in the allergy setting. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1431. [Google Scholar]
- Picardo, M.; Ottaviani, M.; Camera, E.; Mastrofrancesco, A. Sebaceous gland lipids. Dermato-Endocrinology 2009, 1, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, A. Variations in the composition of the skin surface lipid associated with dietary carbohydrates. Proc. Nutr. Soc. 1967, 26, 11. [Google Scholar]
- Macdonald, I. Changes in the fatty acid composition of sebum associated with high carbohydrate diets. Nature 1964, 203, 1067–1068. [Google Scholar] [CrossRef]
- Downing, D.T.; Strauss, J.S.; Pochi, P.E. Changes in skin surface lipid composition induced by severe caloric restriction in man. Am. J. Clin. Nutr. 1972, 25, 365–367. [Google Scholar] [CrossRef]
- Pochi, P.E.; Downing, D.T.; Strauss, J.S. Sebaceous gland response in man to prolonged total caloric deprivation. J. Investig. Dermatol. 1970, 55, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Shine, W.E.; McCulley, J.P. Polar lipids in human meibomian gland secretions. Curr. Eye Res. 2003, 26, 89–94. [Google Scholar] [CrossRef]
- Haffner, S.M. Diabetes, hyperlipidemia, and coronary artery disease. Am. J. Cardiol. 1999, 83, 17–21. [Google Scholar] [CrossRef]
- De Souza, R.J.; Mente, A.; Maroleanu, A.; Cozma, A.I.; Ha, V.; Kishibe, T.; Uleryk, E.; Budylowski, P.; Schunemann, H.J.; Beyene, J.; et al. Intake of saturated and trans unsaturated fatty acids and risk of all cause mortality, cardiovascular disease, and type 2 diabetes: Systematic review and meta-analysis of observational studies. BMJ 2015, 351, h3978. [Google Scholar] [CrossRef] [Green Version]
- Borchman, D.; Yappert, M.C.; Foulks, G.N. Changes in human meibum lipid with meibomian gland dysfunction using principal component analysis. Exp. Eye Res. 2010, 91, 246–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sledge, S.; Henry, C.; Borchman, D.; Yappert, M.C.; Bhola, R.; Ramasubramanian, A.; Blackburn, R.; Austin, J.; Massey, K.; Sayied, S.; et al. Human Meibum Age, Lipid–Lipid Interactions and Lipid Saturation in Meibum from Infants. Int. J. Mol. Sci. 2017, 18, 1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchman, D.; Foulks, G.N.; Yappert, M.C.; Milliner, S.E. Differences in human meibum lipid composition with meibomian gland dysfunction using NMR and principal component analysis. Investig. Ophthalmol. Vis. Sci. 2012, 53, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Nencheva, Y.; Ramasubramanian, A.; Eftimov, P.; Yokoi, N.; Borchman, D.; Georgiev, G.A. Effects of lipid saturation on the surface properties of human meibum films. Int. J. Mol. Sci. 2018, 19, 2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inaba, T.; Tanaka, Y.; Tamaki, S.; Ito, T.; Ntambi, J.M.; Tsubota, K. Compensatory increases in tear volume and mucin levels associated with meibomian gland dysfunction caused by stearoyl-CoA desaturase-1 deficiency. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- He, X.; Zhao, Z.; Wang, S.; Kang, J.; Zhang, M.; Bu, J.; Cai, X.; Jia, C.; Li, Y.; Li, K.; et al. High-fat diet-induced functional and pathological changes in lacrimal gland. Am. J. Pathol. 2020. [Google Scholar] [CrossRef]
- Hargrave, A.; Courson, J.A.; Pham, V.; Landry, P.; Magadi, S.; Shankar, P.; Hanlon, S.; Das, A.; Rumbaut, R.E.; Smith, C.W.; et al. Corneal dysfunction precedes the onset of hyperglycemia in a mouse model of diet-induced obesity. PLoS ONE 2020, 15, e0238750. [Google Scholar] [CrossRef]
- Stern, M.E.; Beuerman, R.W.; Fox, R.I.; Gao, J.; Mircheff, A.K.; Pflugfelder, S.C. A unified theory of the role of the ocular surface in dry eye. In Lacrimal Gland, Tear Film, and Dry Eye Syndromes 2; Springer: Boston, MA, USA, 1998; pp. 643–651. [Google Scholar]
- Gipson, I.K. The ocular surface: The challenge to enable and protect vision: The Friedenwald lecture. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4391–4398. [Google Scholar] [CrossRef] [Green Version]
- Butovich, I.A.; McMahon, A.; Wojtowicz, J.C.; Bhat, N.; Wilkerson, A. Effects of sex (or lack thereof) on meibogenesis in mice (Mus musculus): Comparative evaluation of lipidomes and transcriptomes of male and female tarsal plates. Ocul. Surf. 2019, 17, 793–808. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Schwamb, J.; Feldhaus, V.; Baumann, M.; Patz, M.; Brodesser, S.; Brinker, R.; Claasen, J.; Pallasch, C.P.; Hallek, M.; Wendtner, C.M.; et al. B-cell receptor triggers drug sensitivity of primary CLL cells by controlling glucosylation of ceramides. Blood 2012, 120, 3978–3985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteng, A.-B.; Bhattacharya, A.; Brodesser, S.; Qi, L.; Tan, N.S.; Kersten, S. Feeding Angptl4−/− mice trans fat promotes foam cell formation in mesenteric lymph nodes without leading to ascites. J. Lipid Res. 2017, 58, 1100–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arganda-Carreras, I.; Kaynig, V.; Rueden, C.; Eliceiri, K.W.; Schindelin, J.; Cardona, A.; Seung, H.S. Trainable Weka Segmentation: A machine learning tool for microscopy pixel classification. Bioinformatics 2017, 33, 2424–2426. [Google Scholar] [CrossRef] [PubMed]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction, 2nd ed.; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Chemical Formula | * C:D | Known * mp (°C) | Plasma | Meibum |
|---|---|---|---|---|---|
| Myristic acid | C14H28O2 | 14:0 | 53.9 | ✓ | ✓ |
| Pentadecylic acid | C15H30O2 | 15:0 | 51–53 | ✓ | |
| Palmitic acid | C16H32O2 | 16:0 | 61.8 | ✓ | ✓ |
| Margaric acid | C17H34O2 | 17:0 | 61.3 | ✓ | |
| Stearic acid | C18H36O2 | 18:0 | 69.6 | ✓ | ✓ |
| Nonadecylic | C19H38O2 | 19:0 | 68.0–70.0 | ✓ | |
| Arachidic acid | C20H40O2 | 20:0 | 77.0 | ✓ | ✓ |
| Heneicosylic acid | C21H42O2 | 21:0 | 74.0–75.0 | ✓ | |
| Behenic acid | C22H44O2 | 22:0 | 81.5 | ✓ | ✓ |
| Tricosylic acid | C23H46O2 | 23:0 | 77.0–79.0 | ✓ | |
| Lignoceric acid | C24H48O2 | 24:0 | 88.0 | ✓ | ✓ |
| Pentacosylic acid | C25H50O2 | 25:0 | * n/a | ✓ | |
| Cerotic acid | C26H52O2 | 26:0 | 88.5 | ✓ | ✓ |
| Myristoleic acid | C14H26O2 | 14:1 | −4.0 | ✓ | |
| Palmitoleic acid | C16H30O2 | 16:1 | −0.5–−0.1 | ✓ | ✓ |
| Oleic acid | C18H34O2 | 18:1 | 12.0 | ✓ | ✓ |
| Paullinic acid | C20H38O2 | 20:1 | 13.4 | ✓ | |
| Erucic acid | C22H42O2 | 22:1 | 33.8 | ✓ | ✓ |
| Nervonic acid | C24H46O2 | 24:1 | 39.0 | ✓ | ✓ |
| Ximenic acid | C26H50O2 | 26:1 | 50.5–50.9 | ✓ | ✓ |
| Linoleic acid | C18H32O2 | 18:2 | −5.0 | ✓ | ✓ |
| α- Linoleic acid | C18H30O2 | 18:3 | −11.0 | ✓ | ✓ |
| Eicosatrienoic acid | C20H34O2 | 20:3 | n/a | ✓ | ✓ |
| Arachidonic acid | C20H32O2 | 20:4 | −49.5 | ✓ | ✓ |
| Eicosapentanoic acid | C20H30O2 | 20:5 | −54.0 | ✓ | |
| Docosapentanoic acid | C22H34O2 | 22:5 | −78.0 | ✓ | |
| Docosahexaenoic acid | C22H32O2 | 22:6 | −44.0 | ✓ | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osae, E.A.; Bullock, T.; Chintapalati, M.; Brodesser, S.; Hanlon, S.; Redfern, R.; Steven, P.; Smith, C.W.; Rumbaut, R.E.; Burns, A.R. Obese Mice with Dyslipidemia Exhibit Meibomian Gland Hypertrophy and Alterations in Meibum Composition and Aqueous Tear Production. Int. J. Mol. Sci. 2020, 21, 8772. https://doi.org/10.3390/ijms21228772
Osae EA, Bullock T, Chintapalati M, Brodesser S, Hanlon S, Redfern R, Steven P, Smith CW, Rumbaut RE, Burns AR. Obese Mice with Dyslipidemia Exhibit Meibomian Gland Hypertrophy and Alterations in Meibum Composition and Aqueous Tear Production. International Journal of Molecular Sciences. 2020; 21(22):8772. https://doi.org/10.3390/ijms21228772
Chicago/Turabian StyleOsae, Eugene A., Tiffany Bullock, Madhavi Chintapalati, Susanne Brodesser, Samuel Hanlon, Rachel Redfern, Philipp Steven, C. Wayne Smith, Rolando E. Rumbaut, and Alan R. Burns. 2020. "Obese Mice with Dyslipidemia Exhibit Meibomian Gland Hypertrophy and Alterations in Meibum Composition and Aqueous Tear Production" International Journal of Molecular Sciences 21, no. 22: 8772. https://doi.org/10.3390/ijms21228772