Secondary Metabolites of the Rice Blast Fungus Pyricularia oryzae: Biosynthesis and Biological Function

Chemical Biology Research Group, RIKEN CSRS, Wako, Saitama 351-0198, Japan
Int. J. Mol. Sci. 2020, 21(22), 8698; https://doi.org/10.3390/ijms21228698
Submission received: 31 August 2020
/
Revised: 10 November 2020
/
Accepted: 17 November 2020
/
Published: 18 November 2020
(This article belongs to the Special Issue Molecular Biology and Chemistry of Mycotoxins and Phytotoxins)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Plant pathogenic fungi produce a wide variety of secondary metabolites with unique and complex structures. However, most fungal secondary metabolism genes are poorly expressed under laboratory conditions. Moreover, the relationship between pathogenicity and secondary metabolites remains unclear. To activate silent gene clusters in fungi, successful approaches such as epigenetic control, promoter exchange, and heterologous expression have been reported. Pyricularia oryzae, a well-characterized plant pathogenic fungus, is the causal pathogen of rice blast disease. P. oryzae is also rich in secondary metabolism genes. However, biosynthetic genes for only four groups of secondary metabolites have been well characterized in this fungus. Biosynthetic genes for two of the four groups of secondary metabolites have been identified by activating secondary metabolism. This review focuses on the biosynthesis and roles of the four groups of secondary metabolites produced by P. oryzae. These secondary metabolites include melanin, a polyketide compound required for rice infection; pyriculols, phytotoxic polyketide compounds; nectriapyrones, antibacterial polyketide compounds produced mainly by symbiotic fungi including endophytes and plant pathogens; and tenuazonic acid, a well-known mycotoxin produced by various plant pathogenic fungi and biosynthesized by a unique NRPS-PKS enzyme.
1. Introduction
Filamentous fungi, including plant pathogenic fungi, produce a wide variety of secondary metabolites with unique and complex structures. However, the relationship between pathogenicity and secondary metabolites remains unclear in most cases. Filamentous fungi are a rich source of secondary metabolites for drug development. Whole-genome sequencing analyses have revealed that filamentous fungi possess many more secondary metabolism genes than expected, suggesting that most secondary metabolite biosynthetic genes are silent under laboratory conditions. To utilize fungal secondary metabolite production ability, secondary metabolism genes have been activated through many approaches, including epigenetic control, manipulation of global regulators, ribosome engineering, overexpression of pathway-specific transcription factors, co-culture, and heterologous expression of secondary metabolite gene clusters [1,2,3,4].
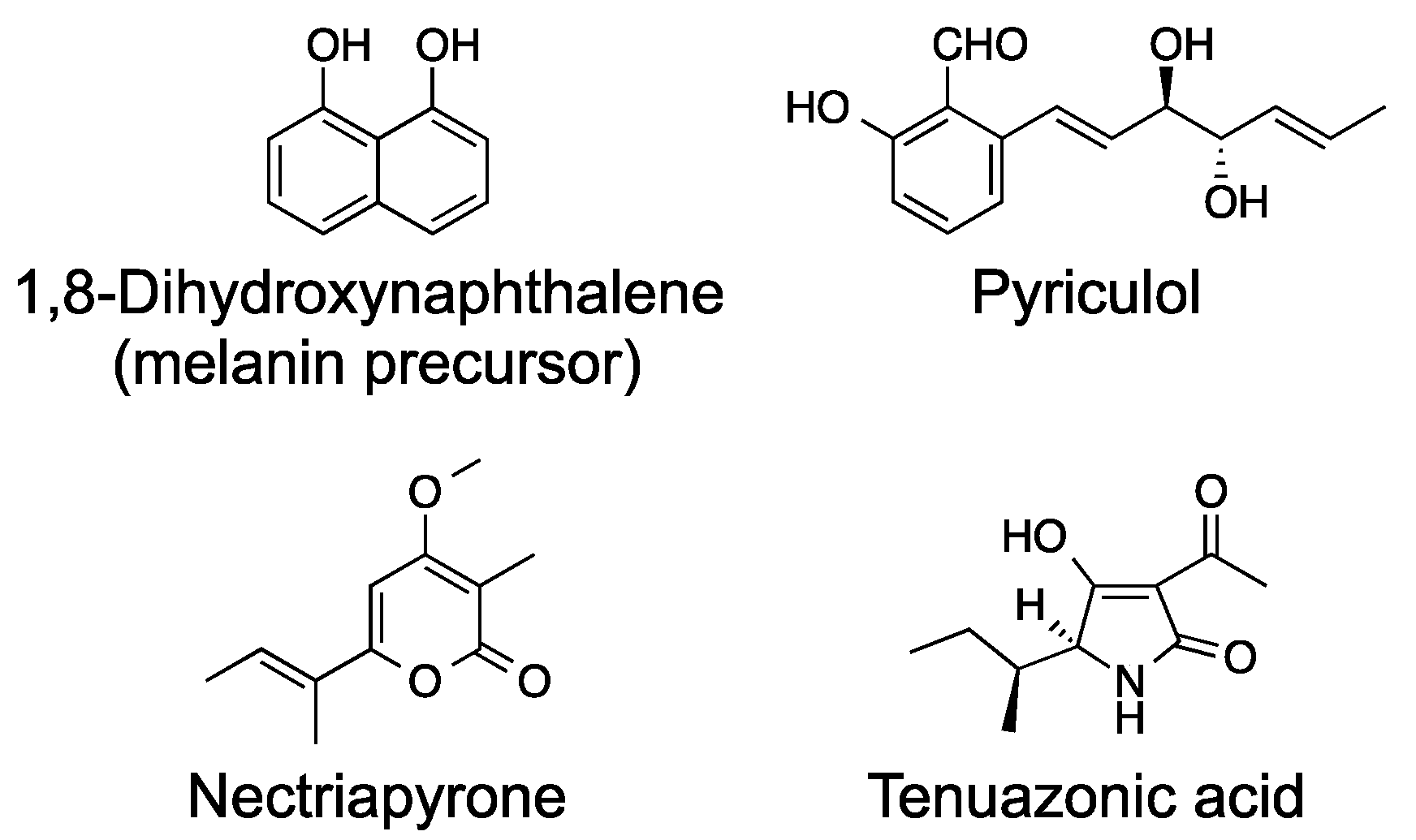
Pyricularia oryzae (syn. Magnaporthe oryzae) is the causal pathogen of rice blast disease and is a well-characterized plant pathogen. P. oryzae infects rice plants through an infection-specific organ, the appressorium, and proliferates inside the rice plant via filamentous growth and causes rice blast disease [5]. P. oryzae is also rich in secondary metabolism genes and shown to have 22 polyketide synthase (PKS) genes and eight non-ribosomal peptide synthetase (NRPS) genes [6,7]. Biosynthetic genes for only four groups of secondary metabolites (melanin, pyriculols, nectriapyrones, and tenuazonic acid) have been well characterized in P. oryzae (Figure 1). Biosynthetic genes for two (nectryapyrones and tenuazonic acid) of the four groups of secondary metabolites have been identified by activating secondary metabolism.
Here, I review the biosynthesis and biological roles of secondary metabolites in the rice blast fungus P. oryzae. This review mainly focuses on the four groups of secondary metabolites shown in Figure 1.
2. Melanin
P. oryzae produces the black pigment melanin (Figure 1), which is essential for rice infection [8]. Melanin is not a toxin, but this secondary metabolite is essential for infection by the mechanism shown below. P. oryzae forms an infection-specific organ, appressorium, and infects rice plants through this organ. Appressorium formation and appressorium melanin formation are essential for rice infection. The invasion of rice plants is achieved by an infection peg that is formed at the base of an appressorium, which adheres tightly to the host surface. For successful penetration from the infection peg, mechanical force exerted by appressoria is necessary [8]. An appressorial melanin layer between the cell wall and cell membrane is required for the generation of the mechanical force. The turgor forces are focused toward the epidermal surfaces of the rice plant, and the pressure inside the appressoria has been estimated to be as high as 8 MPa [9,10]. This pressure can be produced by 3.2 M glycerol formed inside the appressorium [11]. Melanin was proposed to function as a semipermeable membrane that passes water but not glycerol and as a structural support for this very high pressure.
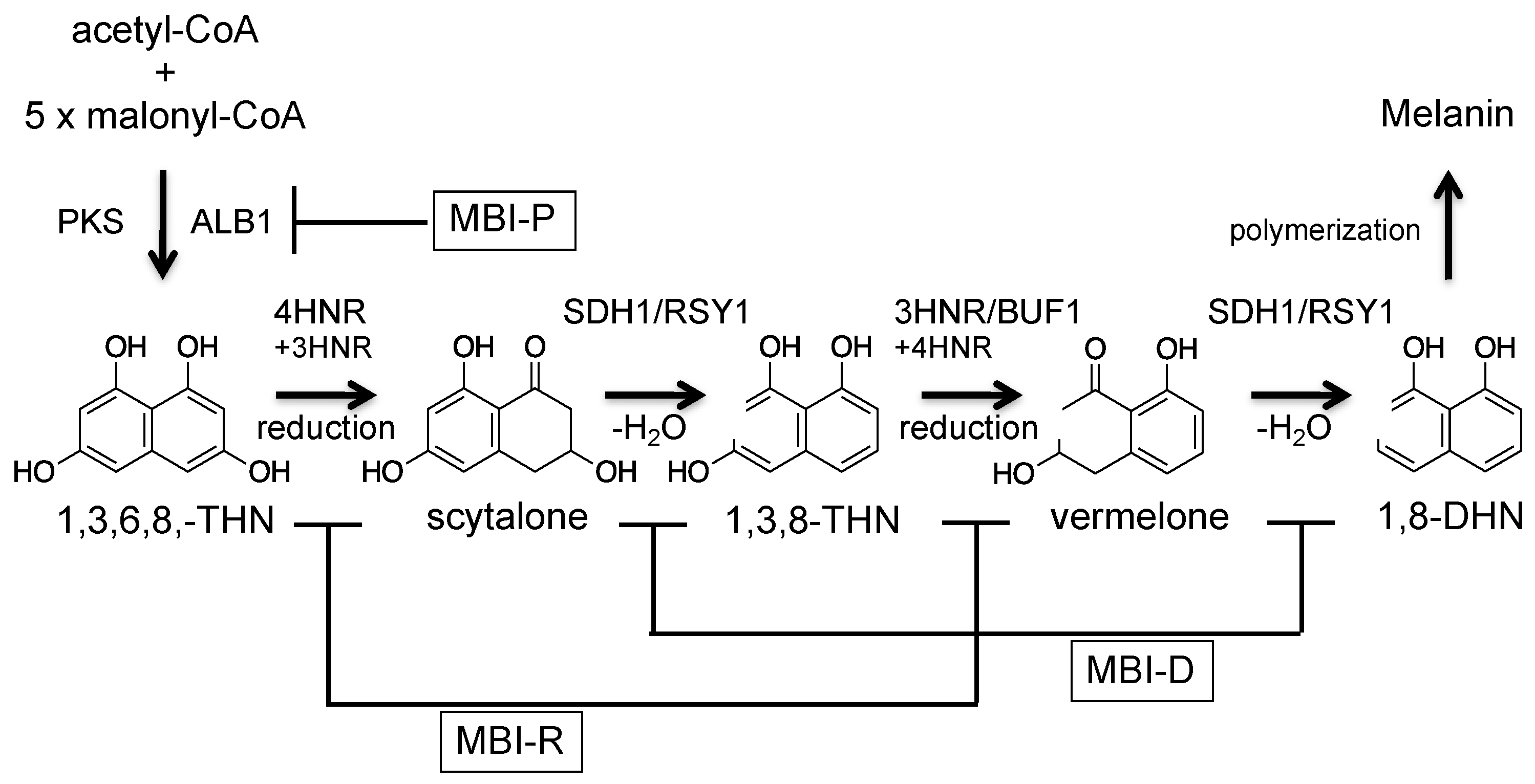
Melanin is a well-known black pigment of biological origin. One type of fungal melanin is dihydroxynaphthalene (DHN)-melanin, which is biosynthesized by polymerizing the polyketide compound 1,8-dihydroxynaphthalene (1,8-DHN) [12,13]. P. oryzae produces DHN-melanin and biosynthetic genes have been identified, and the biosynthetic pathway has been elucidated (Figure 2) [14,15,16,17]. The PKS enzyme ALB1/MGG_07219 biosynthesizes the backbone compound 1,3,6,8-tetrahydroxynaphthalene (1,3,6,8-THN). Melanin was originally proposed as a pentaketide compound. However, from the analysis of an ALB1 homolog in a closely related fungus, Colletotrichum lagenarium, it has been shown that melanin is a hexaketide compound and the backbone (1,3,6,8-THN) is biosynthesized using an acetyl-CoA and five malonyl-CoA [18]. Then, 1,3,6,8-THN is converted to 1,8-DHN by using three enzymes: 1,3,6,8-THN reductase (4HNR), scytalone dehydratase (SDH1/RSY1), and 1,3,8-trihydroxynaphthalene (1,3,8-THN) reductase (3HNR/BUF1). Finally, 1,8-DHN is polymerized to form DHN-melanin. Melanin biosynthesis can be induced by epigenetic control [19].
Melanin biosynthetic enzymes are targets of agrochemical development, and three types of commercial melanin biosynthesis inhibitors (MBIs) have been developed (Figure 2). These inhibitors are classified into three groups: MBI-R (tricyclazole, pyroquilon, and phthalide), MBI-D (carpropamid, diclocymet, and fenoxanil), and MBI-P (tolprocarb) [20,21,22]. The targets of MBI-R, MBI-D, and MBI-P are 1,3,8-trihydroxynaphthalene reductase (3HNR/BUF1), scytalone dehydratase (SDH1/RSY1), and polyketide synthase (ALB1), respectively. MBIs are environmentally friendly agrochemicals because MBIs inhibit fungal infection without inhibiting fungal growth.
3. Pyriculols
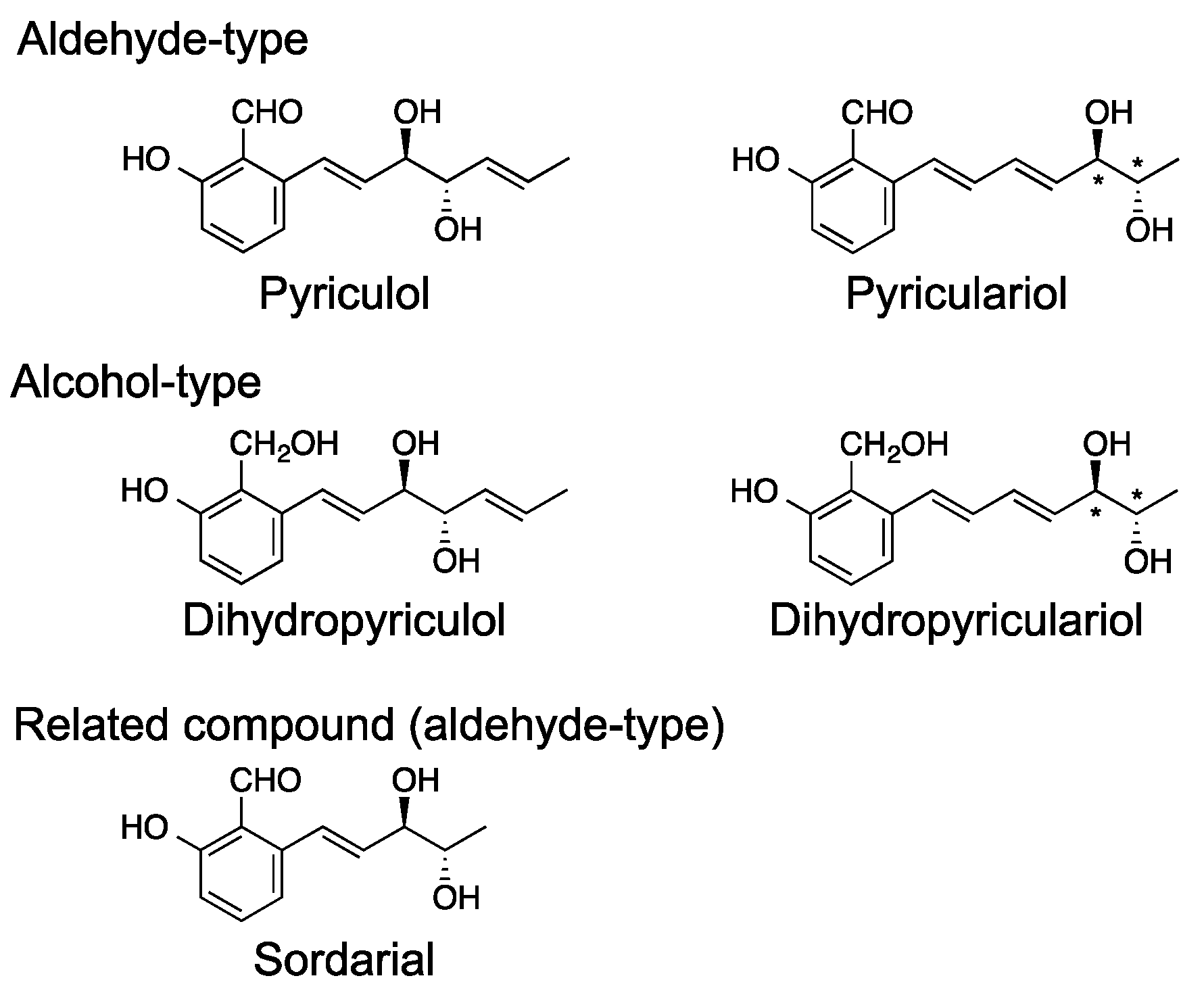
Pyriculol (Figure 1) is a well-known secondary metabolite of the rice blast fungus and is known as a phytotoxin [23]. Several analogs of pyriculol have been reported. The main analogs are dihydropyriculol [24], pyriculariol [25], and dihydropyriculariol [26] (Figure 3). It is also shown that griseaketides, analogs of pyriculol, are produced by an isolate of rice blast fungus [27]. Pyriculols are classified into two groups: Alcohol-type (dihydropyriculol and dihydropyriculariol) and aldehyde-type (pyriculol and pyriculariol). It has been shown that aldehyde derivatives induce lesion-like necrosis on rice leaves, while alcohol derivatives are inactive [24,26,28]. Four analogs are produced simultaneously [28] and interconversion between oxidized aldehyde analogs and reduced alcohol derivatives is expected. Currently, it is not clear why and how P. oryzae produces both alcohol and aldehyde analogs. Identification of the genes responsible for this oxidoreductive conversion will help to answer this question.
Pyriculols are polyketide compounds, and the biosynthetic gene cluster has been recently identified [28]. The PKS gene (MGG_10912/MoPKS19) and other genes predicted to be responsible for the biosynthesis of pyriculols have been identified [28]. It has been suggested that aldehyde-type analogs are produced first and converted to alcohol-type analogs by the reduction reaction [26]. The gene (MGG_10961/MoC19OXR1) responsible for the oxidation of alcohol-type analogs to aldehyde-type analogs has been reported [9], although the gene catalyzing the reverse reductive reaction has not yet been identified.
Neurospora crassa produces the structurally related salicylaldehyde sordarial (Figure 3). The biosynthetic mechanism of sordarial has been proposed [29]. In sordarial biosynthesis, it has been predicted that an aldehyde-type intermediate is released from PKS (SrdA) and cyclized by SrdC/D/E. The aldehyde-type intermediate is predicted to be modified by SrbB and SrdG to yield sordarial. In this pathway, an alcohol-type intermediate is thought to be produced from an aldehyde congener by an endogenous reductase; however, the gene responsible for the reduction of the aldehyde moiety has not yet been identified. SrdI, a homolog of MGG_10961/MoC19OXR1, is predicted to be involved in the oxidation of this alcohol-type intermediate [29]. The biological functions of sordarial are not known.
The extract of the PKS gene knockout strain fails to induce phytotoxic lesions on rice leaves, indicating that pyriculols are the sole lesion-inducing compounds produced by the wild-type strain under the culture condition used [28]. Interestingly, the PKS gene knockout strain is as pathogenic as the wild-type strain, demonstrating that pyriculols are not required for infection [28]. Further research is required to elucidate the biological roles of pyriculols.
4. Nectriapyrones
Nectriapyrone (Figure 1) is known as a polyketide compound produced by various fungi [30,31,32,33,34,35,36,37,38,39,40,41,42]. Interestingly, producers are mainly symbiotic fungi, including endophytes [34,36,37,38,41,43], plant pathogens [32,35], and sponge-associated fungi [30,31,33]. We recently found that nectriapyrone production can be induced in the rice blast fungus P. oryzae by disturbing the two-component signal transduction system [44]. We identified the nectriapyrone biosynthetic gene cluster and analyzed its physiological function.
Secondary metabolite production may be strictly regulated to produce under specific environmental conditions. Thus, we predicted that secondary metabolite production may be activated by disturbing signal transduction pathways involved in environmental responses. A two-component system (TCS) is a signal transduction system that regulates various cellular functions in response to environmental signals and is found in bacteria, archaea, plants, slime molds, and fungi [45,46]. The P. oryzae TCS was disturbed by disrupting OSM1 and PoYPD1, encoding a HOG MAP kinase and a His-containing phosphotransfer (HPt) protein, respectively. This genetic modification induced the production of two polyketide compounds, nectriapyrone and its hydroxylated analog.
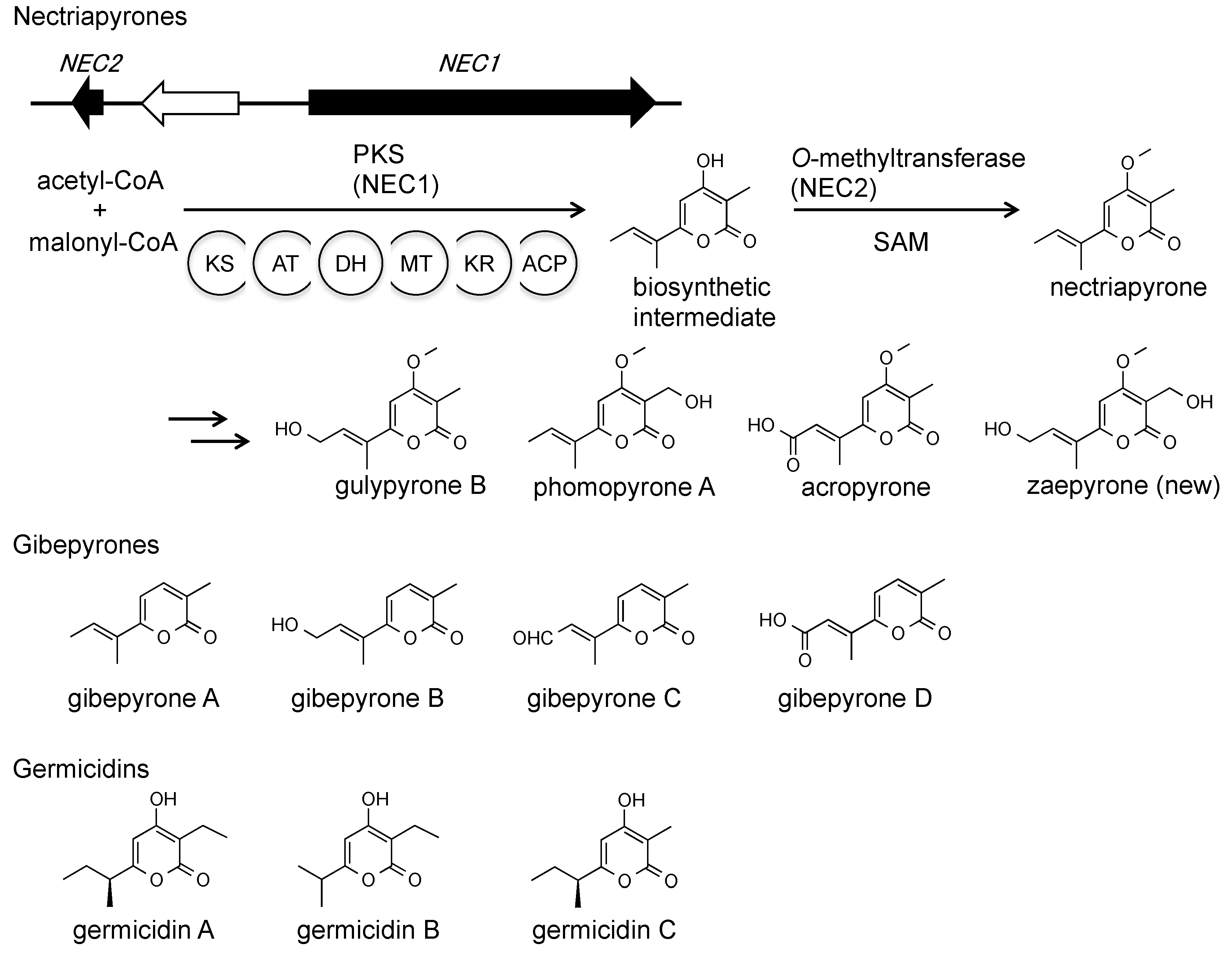
We identified the nectriapyrone biosynthetic gene cluster consisting of a PKS gene (NEC1/MGG_00806) and an O-methyltransferase gene (NEC2/MGG_14657) (Figure 4). Overexpression of the two genes caused overproduction of nectriapyrone and five nectriapyrone analogs, including a new derivative, zaepyrone (Figure 4) [47,48,49,50]. Nectriapyrone shows similarity to the gibepyrones (Figure 4) from Fusarium spp. Gebepyrones do not have a methoxy group, and the O-methyltransferase gene is absent from the gibepyrone biosynthetic gene cluster [51]. Nectriapyrone also shows similarity to germicidins [52,53,54] from Streptomyces spp. (Figure 4). A type III PKS biosynthesizes the germicidin backbone [55]. In contrast, a type I PKS (NEC1) biosynthesizes the nectriapyrone backbone.
Nectriapyrones belong to the class α-pyrone. α-Pyrones have a wide range of biological activities [56,57]. For example, photopyrones are bacterial signaling molecules that control cell clumping [58]. Germicidins, produced by some Streptomyces strains, act as autoregulators of spore germination [52,53]. Some biological activities of nectriapyrone have been reported, although the functions of nectriapyrone in its producers are unknown. Nectriapyrone is toxic to bacteria, tumor cells, and plants [35,36,42,59]. It stimulates the formation of DOPA melanin in B16-F1 melanoma cells [39]. Nectriapyrone also inhibits monoamine oxidase in the mouse brain [60].
Identification of the nectriapyrone biosynthetic gene cluster allowed us to analyze the biological functions of nectriapyrones in the fungi that produce them [44]. While many nectriapyrone producers have been identified from plant pathogens, our data have indicated that nectriapyrones are not involved in rice infection and have different functions. The structure of nectriapyrone is similar to that of the germicidins produced by Streptomyces spp. Our data have indicated that nectriapyrone can control growth and pigment formation in S. griseus and has a growth-promoting effect on P. oryzae in interactions with S. griseus. Therefore, nectriapyrones may be involved in microbe-microbe interactions with other environmental organisms, including bacteria such as endophytic Streptomyces strains. To identify the active nectriapyrone analogs in this interaction, we analyzed the bioactivity of each analog (Figure 4) and found that nectriapyrone was the active analog, suggesting that other nectriapyrone analogs may be inactivated (detoxified) compounds of nectriapyrone.
5. Tenuazonic Acid
Tenuazonic acid (TeA, Figure 1), a tetramic acid derivative, is a well-known mycotoxin first isolated from the culture broth of Alternaria tenius in 1957 [61]. Alternaria, a ubiquitous plant pathogenic fungus, causes spoilage of various fruits and food crops in the field and post-harvest decay [62]. TeA has been detected in various Alternaria-contaminated vegetables, fruits, and crops [63,64,65]. The plant pathogenic fungi P. oryzae and Phoma sorghina (sorghum pathogen) are known as TeA producers [66,67]. P. oryzae also produces a related tetramic acid derivative from valine as a very minor product [68]. Among the Alternaria toxins, TeA is the most toxic and shows acute toxicity to mammals. The oral median lethal doses for male and female mice are 182 or 225 mg kg−1 and 81 mg kg−1 body weight, respectively [69,70]. The European Food Safety Authority evaluates TeA toxicity and determines its threshold of toxicological concern to be 1500 ng kg−1 body weight day [71]. TeA inhibits protein biosynthesis by inhibiting the release of the polypeptide from the ribosome [72]. TeA also has antitumor, antiviral, antibacterial, phytotoxic, and plant disease-controlling activities [69,73,74,75]. TeA has been shown to inhibit photosynthesis [76,77,78], and the potential use of TeA as a herbicide targeting photosystem II (PSII) has been proposed [79]. A recent paper [80] has shown that TeA inhibits plant plasma membrane (PM) H+-ATPase at micromolar concentrations. Inhibition of the PM H+-ATPase results in depolarization of the membrane potential and eventually necrosis. However, it is still unclear whether TeA is required for plant infection or not. We previously induced TeA production in P. oryzae and identified the biosynthetic gene TAS1, encoding the first reported fungal NRPS-PKS hybrid enzyme [81,82]. We also revealed the biosynthetic and induction mechanisms of TeA [83,84]. In this section, we review the data on the induction and biosynthesis of TeA.
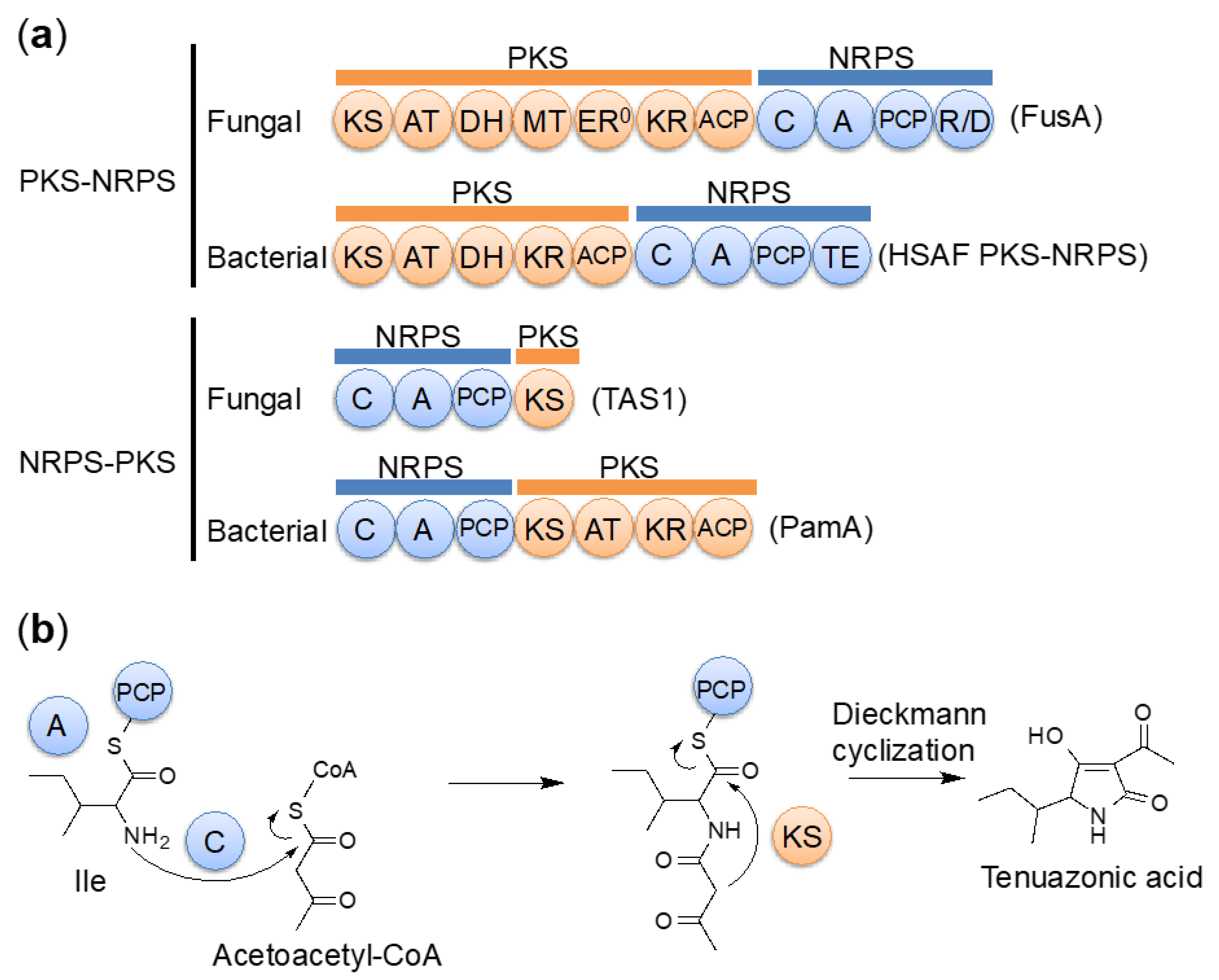
The major fungal secondary metabolites, polyketides and nonribosomal peptides, are biosynthesized by PKSs and NRPSs, respectively. Fungal PKSs can be classified into three types. The first type is iterative type I PKSs, which consist of multiple catalytic domains, including ketosynthase (KS), acyltransferase (AT), and acyl carrier protein (ACP) main domains, along with several optional β-keto modifying domains, such as β-ketoacyl reductase (KR), dehydratase (DH), and trans-acting enoyl reductase (ER) domains [85]. The second type is the type III PKSs, which consist of a homodimeric KS [86]. The third type is fungal PKS-NRPS hybrid enzymes, which consist of an iterative type I PKS followed by a single module NRPS. These enzymes are involved in producing a wide variety of structurally diverse polyketide-amino acid hybrid compounds [87,88,89,90,91]. In fungal PKS-NRPS, the PKS part consists of KS, AT, and ACP domains, along with several modifying domains such as KR, DH, and methyltransferase (MT) domains. The NRPS part consists of adenylation (A), peptidyl carrier protein (PCP), condensation (C), and terminal release or cyclization (R, reductase or DKC, Dieckmann cyclization) domains [90]. PKS-NRPS hybrid enzymes have also been observed in bacteria [92,93]. Furthermore, a different type of hybrid enzymes, NRPS–PKS hybrid enzymes (which begin with an NRPS module), are also known. However, this type of enzyme has only been found in bacteria [94,95,96,97,98,99].
TeA was shown to be a hybrid of an isoleucine and two acetates [100]. Because TeA has a tetramic acid-containing structure, TeA was also expected to be a product of a PKS-NRPS hybrid enzyme [101]. We successfully induced TeA production by disruption of the OSM1 gene and 1% dimethyl sulfoxide (DMSO) treatment. OSM1 is the osmosensory mitogen-activated protein kinase (MAPK), which works downstream of the two-component signal transduction system involved in environmental responses. We identified the TeA biosynthetic gene TAS1/MGG_07803 from the induced genes under inducing conditions [81,82]. TAS1 (tenuazonic acid synthetase 1) was not a PKS-NRPS hybrid enzyme but a NRPS-PKS hybrid enzyme. TAS1 is the first reported fungal NRPS-PKS hybrid enzyme consisting of an NRPS module of domains C-A-PCP and a terminal PKS KS domain (Figure 5a). TAS1 is a novel NRPS-PKS hybrid enzyme that starts with an NRPS module (C-A-PCP). This domain structure is very different from that of conventional fungal PKS-NRPS enzymes, which start with a PKS module (Figure 5a). The PKS portion of TAS1 has only a KS domain, in contrast to other NRPS-PKS hybrid enzymes. This KS domain is indispensable for TAS1 activity and has a unique sequence. By phylogenetic analysis, this KS domain was classified as an independent clade close to the type I PKS KS domain. We revealed that TAS1 synthesizes TeA from isoleucine and acetoacetyl-CoA (diketide) (Figure 5b). The unique KS domain catalyzes the final Dieckmann cyclization step for tetramic acid ring formation and TeA release, although involvement in diketide biosynthesis has been previously predicted [100]. In other NRPSs, bacterial NRPSs use the terminal thioesterase (TE) domain to catalyze substrate cyclization [102], whereas fungal NRPSs use the terminal condensation-like domain for substrate cyclization [103]. In addition, fungal PKS–NRPSs use the terminal reductase-like cyclization (DKC) domain for substrate cyclization [90]. These data indicate that TAS1 is a unique type of biosynthetic enzyme and may be used for the production of various compounds.
The KS domains of PKS normally catalyze the decarboxylative Claisen condensation of acyl and malonyl blocks to extend the polyketide chain [104]. In contrast, the terminal KS domain in TAS1 from P. oryzae conducts substrate cyclization [81]. Nonconventional KS domains with noncanonical roles have also been reported in type I PKS systems. A KS domain, one His residue of the catalytic triad is mutated, has been reported to only catalyze substrate transfer to the next domain in FR901464 biosynthesis [105]. KS3 of RhiE is required for vinylogous chain branching without polyketide chain elongation in rhizoxin biosynthesis [106]. Furthermore, homodimers of KS domains catalyze polyketide extension and substrate cyclization in a single catalytic pocket in type III PKSs [107]. However, a KS domain without a polyketide chain extension role has only been reported in TAS1. We analyzed the unique features of the TAS1 KS domain [84]. We found that the TAS1 KS domain is uniquely monomeric like NRPSs [108] although KSs are usually dimeric [109,110,111]. The 1.68 Å resolution crystal structure suggested that the substrate cyclization is triggered by proton abstraction from the active methylene moiety in the substrate by the catalytic H322 residue. We also found that TAS1 KS shows broad substrate specificity and promiscuously accepts aminoacyl substrates. Furthermore, this promiscuity could be increased by a single amino acid substitution in the substrate-binding pocket. These data provide hints to the substrate cyclization mechanism performed by the KS domain in TeA biosynthesis. These data also provide insight into how the NRPS-PKS hybrid enzyme accepts bulky amino acid-containing substrates.
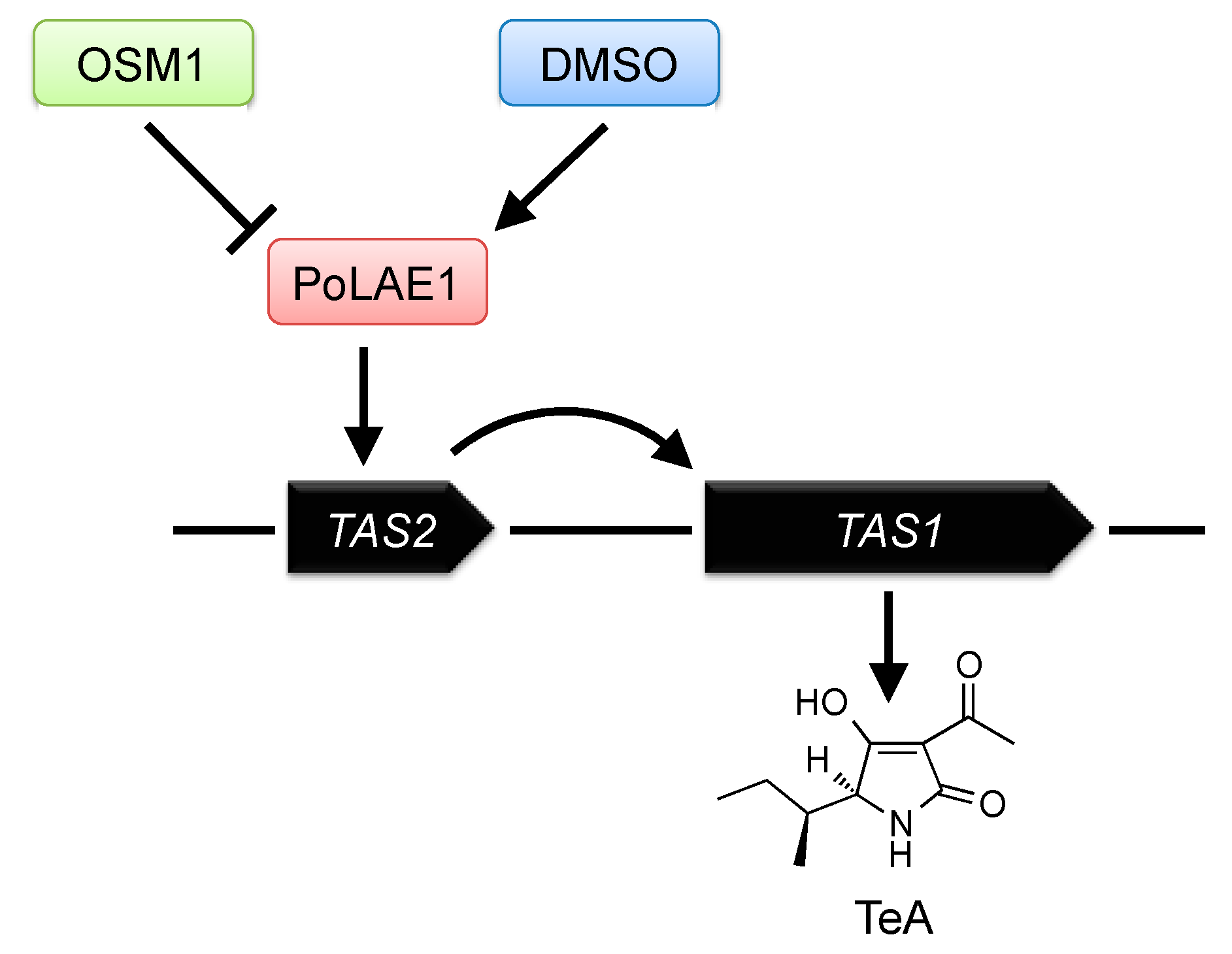
Transcription of secondary metabolism genes should be properly regulated in fungi in response to various environmental signals. Elucidation of the regulatory mechanism of these secondary metabolism genes is important for understanding the interaction between fungi and their environments. For example, elucidation of the regulatory mechanism of mycotoxin biosynthesis is important for protecting human and animal health by controlling mycotoxin production. As shown previously, TeA is biosynthesized in P. oryzae by TAS1, and its production is induced by osmo-sensory MAPK gene (OSM1) deletion or 1% DMSO treatment. However, the detailed regulatory mechanisms of TeA production have been unknown. We found two positive regulators of TeA production (Figure 6) [83]. In most cases, fungal secondary metabolites are produced using biosynthetic gene clusters. Many gene clusters have a gene for a cluster-specific DNA binding binuclear Zn(II)2Cys6-type transcription factor, which is known to be unique to fungi and activates the transcription of the clustered genes to produce a secondary metabolite [112]. These transcription factor genes include Aspergillus nidulans aflR for aflatoxin biosynthesis [113], A. fumigatus gliZ for gliotoxin biosynthesis [114], Monascus purpureus ctnA for citrinin biosynthesis [115], and Fusarium sporotrichioides tri6 for trichothecene biosynthesis [116]. We identified a Zn(II)2Cys6-type transcription factor, TAS2 (MGG_07800), which regulates TeA production. TAS2 is located in the upstream region of TAS1 (Figure 6). In fungi, secondary metabolite production is also regulated by upper-level regulators rather than by cluster-specific transcription factors. These upper-level regulators, called global regulators, are trans-acting positive or negative transcriptional factors of secondary metabolite gene clusters. LaeA (loss of aflR expression) is a well-known global regulator of secondary metabolism identified and characterized in Aspergillus spp. [117,118,119,120]. Orthologs of LaeA have been identified in other fungi, including Monascus pilosus, Cochliobolus heterostrophus, and Fusarium spp. [121]. We identified a LaeA ortholog, PoLAE1 (MGG_01233), from P. oryzae. Analysis of PoLAE1 deletion and overexpression strains showed that PoLAE1 positively regulated TeA production. We also revealed that two TeA-inducing signals, OSM1 deletion and 1% DMSO treatment, were transmitted via PoLAE1. These results indicated that TeA production was regulated by two specific regulators, TAS2 and PoLAE1, in P. oryzae (Figure 6). Recently, it has been shown that TeA production is also induced by mycovirus infection via upregulation of TAS2 [122].
6. Other Secondary Metabolites
ACE1 (MGG_12447) is an avirulence gene, and isolates of P. oryzae carrying the ACE1 gene are specifically recognized by rice cultivars carrying the resistance gene Pi33 [87]. This recognition activates defense responses in resistant plants. ACE1 is a secondary metabolism gene and encodes a PKS-NRPS hybrid enzyme. The secondary metabolite whose synthesis is dependent on ACE1 is predicted to be recognized by resistant rice plants. ACE1 is located in an infection-specific gene cluster consisting of 15 genes [7]. Fourteen of the 15 genes are predicted to be involved in secondary metabolism as they code for proteins such as a second PKS-NRPS (SYN2), two enoyl reductases (RAP1 and RAP2), and a putative Zn(II)2Cys6-type transcription factor (BC2). The ACE1 gene cluster shows a rice-infection-specific expression pattern. Heterologous co-expression of ACE1 and RAP1 in Aspergillus oryzae causes production of an amide compound similar to the PKS-NRPS-derived backbone of cytochalasans [123]. Bioactivity analysis shows that the produced compound is not responsible for the observed ACE1-mediated avirulence. These data suggest that the active final product may be a cytochalasin-like compound.
Penicillin production in rice blast fungus has been recently shown [124]. Overexpression of a laeA homolog (MoLAEA/MGG_07964) increases the production of penicillin G compared to the wild type strain. In contrast, the silenced strain does not produce penicillin G. The putative NRPS gene (MGG_14767) for penicillin G biosynthesis is 3-fold upregulated in the overexpression strain, whereas it is 3.8-fold downregulated in the knockdown strain. Transcriptomic data show that MoLAEA regulates genes involved in secondary metabolism. This laeA homolog (MoLAEA/MGG_07964) is different from the laeA homolog (PoLAE1/MGG_01233) involved in TeA production. Multiple laeA homologs may be involved in the control of secondary metabolite production.
ABM (MGG_04777) is a monooxidase gene located in a putative secondary metabolite biosynthetic gene cluster containing a polyketide synthase gene (MGG_04775) [125]. While the role of Abm (antibiotic biosynthesis monooxygenase) in this gene cluster is unknown, Abm can convert endogenous free jasmonic acid (JA) into 12OH-JA in P. oryzae. Such fungal 12OH-JA is secreted during infection and helps evade the defense response by inhibiting the induction of JA signaling. In contrast, loss of Abm in P. oryzae causes accumulation of methyl JA (MeJA), which induces host defense and blocks fungal infection. Furthermore, Abm itself is secreted after infection and is predicted to convert plant JA into 12OH-JA to help host colonization. P. oryzae also produces other plant hormones, abscisic acid (ABA) [126], cytokinins (CKs) [127], and auxins [indole-3-acetic acid (IAA)] [128], and the biosynthetic genes for ABA and CKs have been identified and characterized [129,130]. Gene knockout of MoABA4/MGG_07514, homologous to the Botrytis cinerea ABA4 gene responsible for ABA biosynthesis, reduces ABA levels by two-fold [129]. The virulence of the ΔMoaba4 mutant is strongly compromised, suggesting that ABA contributes to the virulence of this fungus. CKS1/MGG_04857 encodes a putative tRNA-Isopentenyl transferase (tRNA-IPT) protein essential for CK biosynthesis [130]. The interaction between rice and the Δcsk1 strain has been characterized. This analysis has shown that P. oryzae-derived CKs are required for full virulence by affecting rice defenses, nutrient distribution, and fungal oxidative stress tolerance.
7. Conclusions
P. oryzae is rich in secondary metabolism genes, and some of the secondary metabolites are expected to be involved in rice infection. Here, this review focused on the biosynthesis and biological roles of secondary metabolites in P. oryzae. Five groups of secondary metabolites (melanin, pyriculols, nectriapyrones, TeA, and penicillin G) have been shown to be produced by P. oryzae. Biosynthetic genes for three (nectryapyrone, TeA, and penicillin G) of the five groups of secondary metabolites have been identified by activating secondary metabolism. Nectryapyrones and TeA were induced by manipulating the factors involved in the TCS. Penicillin G was induced by overexpression of a laeA homolog. Activation of secondary metabolism is a useful method for the identification of secondary metabolites. Melanin is a well-characterized secondary metabolite that is essential for rice infection. Identification and gene manipulation of the biosynthetic genes revealed that three groups of secondary metabolites (pyriculols, nectriapyrones, and TeA) are not required for rice infection. Nectriapyrones and penicillin G show antibacterial activity and are predicted to be involved in interactions between bacteria. Controlling plant pathogenic fungi is important in agriculture. Unveiling the roles of secondary metabolites of plant pathogens will help in developing agrochemicals.
Funding
This research was funded by JSPS KAKENHI, grant numbers JP17H06412, JP17K07742, JP17K07784, JP18H03945, and JP20K05820.
Conflicts of Interest
The author declares no conflict of interest.
Abbreviations
| PKS | Polyketide synthase |
| NRPS | Nonribosomal peptide synthetase |
| 1,8-DHN | Dihydroxynaphthalene |
| 1,3,6,8-THN | 1,3,6,8-Tetrahydroxynaphthalene |
| 1,3,8-THN | 1,3,8-Trihydroxynaphthalene |
| MBIs | Melanin biosynthesis inhibitors |
| TCS | Two-component system |
| HPt | His-containing phosphotransfer |
| DOPA | 3,4-Dihydroxyphenylalanine |
| TeA | Tenuazonic acid |
| PSII | Photosystem II |
| PM | Plasma membrane |
| KS | Ketosynthase |
| AT | Acyltransferase |
| ACP | Acyl carrier protein |
| KR | β-Ketoacyl reductase |
| DH | Dehydratase |
| ER | Enoyl reductase |
| MT | Methyltransferase |
| A | Adenylation |
| PCP | Peptidyl carrier protein |
| C | Condensation |
| R | Reductase |
| DKC | Dieckmann cyclization |
| DMSO | Dimethylsulfoxide |
| MAPK | Mitogen-activated protein kinase |
| TE | Thioesterase |
| JA | Jasmonic acid |
| MeJA | Methyl jasmonic acid |
| ABA | abscisic acid |
| CKs | cytokinins |
| IAA | indole-3-acetic acid |
| tRNA-IPT | tRNA-Isopentenyl transferase |
References
- Ochi, K.; Hosaka, T. New strategies for drug discovery: Activation of silent or weakly expressed microbial gene clusters. Appl. Microbiol. Biotechnol. 2013, 97, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Netzker, T.; Fischer, J.; Weber, J.; Mattern, D.J.; Konig, C.C.; Valiante, V.; Schroeckh, V.; Brakhage, A.A. Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 2015, 6, 299. [Google Scholar] [CrossRef]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Reviews. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and Role of Fungal Secondary Metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Valent, B. Breaking and entering: Host penetration by the fungal rice blast pathogen Magnaporthe grisea. Annu. Rev. Microbiol. 1996, 50, 491–512. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.A.; Talbot, N.J.; Ebbole, D.J.; Farman, M.L.; Mitchell, T.K.; Orbach, M.J.; Thon, M.; Kulkarni, R.; Xu, J.R.; Pan, H.; et al. The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 2005, 434, 980–986. [Google Scholar] [CrossRef]
- Collemare, J.; Pianfetti, M.; Houlle, A.E.; Morin, D.; Camborde, L.; Gagey, M.J.; Barbisan, C.; Fudal, I.; Lebrun, M.H.; Bohnert, H.U. Magnaporthe grisea avirulence gene ACE1 belongs to an infection-specific gene cluster involved in secondary metabolism. New Phytol. 2008, 179, 196–208. [Google Scholar] [CrossRef]
- Howard, R.J.; Ferrari, M.A. Role of Melanin in Appressorium Function. Exp. Mycol. 1989, 13, 403–418. [Google Scholar] [CrossRef]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1991, 88, 11281–11284. [Google Scholar] [CrossRef] [Green Version]
- Money Nicholas, P.; Howard Richard, J. Confirmation of a link between fungal pigmentation, turgor pressure, and pathogenicity using a new method of turgor measurement. Fungal. Genet. Biol. 1996, 20, 217–227. [Google Scholar] [CrossRef]
- De Jong, J.C.; McCormack, B.J.; Smirnoff, N.; Talbot, N.J. Glycerol generates turgor in rice blast. Nature 1997, 389, 244–245. [Google Scholar] [CrossRef]
- Bell, A.A.; Wheeler, M.H. Biosynthesis and functions of fungal melanins. Ann. Rev. Phytopathol. 1986, 24, 411–451. [Google Scholar] [CrossRef]
- Butler, M.J.; Day, A.W. Fungal melanins: A review. Can. J. Microbiol. 1998, 44, 1115–1136. [Google Scholar] [CrossRef]
- Lundqvist, T.; Rice, J.; Hodge, C.N.; Basarab, G.S.; Pierce, J.; Lindqvist, Y. Crystal structure of scytalone dehydratase--a disease determinant of the rice pathogen, Magnaporthe grisea. Structure 1994, 2, 937–944. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Cros, A.; Viviani, F.; Labesse, G.; Boccara, M.; Gaudry, M. Polyhydroxynaphthalene reductase involved in melanin biosynthesis in Magnaporthe grisea. Purification, cDNA cloning and sequencing. Eur. J. Biochem. 1994, 219, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.E.; Fahnestock, S.; Farrall, L.; Liao, D.I.; Valent, B.; Jordan, D.B. The second naphthol reductase of fungal melanin biosynthesis in Magnaporthe grisea: Tetrahydroxynaphthalene reductase. J. Biol. Chem. 2000, 275, 34867–34872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliahu, N.; Igbaria, A.; Rose, M.S.; Horwitz, B.A.; Lev, S. Melanin biosynthesis in the maize pathogen Cochliobolus heterostrophus depends on two mitogen-activated protein kinases, Chk1 and Mps1, and the transcription factor Cmr1. Eukaryot. Cell 2007, 6, 421–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagstad, A.L.; Hill, E.A.; Labonte, J.W.; Townsend, C.A. Characterization of a fungal thioesterase having Claisen cyclase and deacetylase activities in melanin biosynthesis. Chem. Biol. 2012, 19, 1525–1534. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Izawa, M.; Nakajima, Y.; Jin, Q.; Hirose, T.; Nakamura, T.; Koshino, H.; Kanamaru, K.; Ohsato, S.; Kamakura, T.; et al. Increased metabolite production by deletion of an HDA1-type histone deacetylase in the phytopathogenic fungi, Magnaporthe oryzae (Pyricularia oryzae) and Fusarium asiaticum. Lett. Appl. Microbiol. 2017, 65, 446–452. [Google Scholar] [CrossRef]
- Motoyama, T.; Yamaguchi, I. Fungicides, Melanin Biosynthesis Inhibitors. In Encyclopedia of Agrochemicals; Plimmer, J.R., Gammon, D.W., Ragsdale, N.N., Eds.; Wiley: Hoboken, NJ, USA, 2003; Volume 2, pp. 584–592. [Google Scholar]
- Banba, S.; Hamada, T.; Araki, N.; Ebihara, K. Synthesis and activities of tolprocarb derivatives against Pyricularia oryzae: Relationships among the activities for polyketide synthase, melanin biosynthesis, and rice blast. J. Pestic Sci. 2017, 42, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, H.; Ezaki, R.; Hamada, T.; Tsuda, M.; Ebihara, K. Development of a novel fungicide, tolprocarb. J. Pestic Sci. 2019, 44, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, S.; Muro, H.; Sasaki, K.; Nozoe, S.; Okuda, S.; Sato, Z. Isolations of phytotoxic substances produced by pyricularia oryzae cavara. Tetrahedron Lett. 1973, 14, 3537–3542. [Google Scholar] [CrossRef]
- Kono, Y.; Sekido, S.; Yamaguchi, I.; Kondo, H.; Suzuki, Y.; Neto, G.C.; Sakurai, A.; Yaegashi, H. Structures of Two Novel Pyriculol-related Compounds and Identification of Naturally Produced Epipyriculol from Pyricularia oryzae. Agric. Biol. Chem. 1991, 55, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Nukina, M.; Sassa, T.; Ikeda, M.; Umezawa, T.; Tasaki, H. Pyriculariol, a New Phytotoxic Metabolite of Pyricularia oryzae Cavara. Agric. Biol. Chem. 1981, 45, 2161–2162. [Google Scholar] [CrossRef]
- Tanaka, K.; Sasaki, A.; Cao, H.-Q.; Yamada, T.; Igarashi, M.; Komine, I.; Nakahigashi, H.; Minami, N.; Kuwahara, S.; Nukina, M.; et al. Synthesis and Biotransformation of Plausible Biosynthetic Intermediates of Salicylaldehyde-Type Phytotoxins of Rice Blast Fungus, Magnaporthe grisea. Eur. J. Org. Chem. 2011, 2011, 6276–6280. [Google Scholar] [CrossRef]
- Yang, Y.H.; Yang, D.S.; Lei, H.M.; Li, C.Y.; Li, G.H.; Zhao, P.J. Griseaketides A-D, New Aromatic Polyketides from the Pathogenic Fungus Magnaporthe grisea. Molecules 2019, 25. [Google Scholar] [CrossRef] [Green Version]
- Jacob, S.; Grötsch, T.; Foster, A.J.; Schüffler, A.; Rieger, P.H.; Sandjo, L.P.; Liermann, J.C.; Opatz, T.; Thines, E. Unravelling the biosynthesis of pyriculol in the rice blast fungus Magnaporthe oryzae. Microbiol. (Read. Engl.) 2017, 163, 541–553. [Google Scholar] [CrossRef]
- Zhao, Z.; Ying, Y.; Hung, Y.S.; Tang, Y. Genome Mining Reveals Neurospora crassa Can Produce the Salicylaldehyde Sordarial. J. Nat. Prod. 2019, 82, 1029–1033. [Google Scholar] [CrossRef]
- He, W.-J.; Mai, Y.-X.; Lin, X.-P.; Liao, S.-R.; Yang, B.; Wang, J.-F.; Liu, Y.; He, W.-J.; Zhou, X.-J.; Qin, X.-C.; et al. Quinone/hydroquinone meroterpenoids with antitubercular and cytotoxic activities produced by the sponge-derived fungus Gliomastix sp. ZSDS1-F7. Nat. Prod. Res. 2017, 31, 604–609. [Google Scholar] [CrossRef]
- Gong, T.; Zhen, X.; Li, B.-J.; Yang, J.-L.; Zhu, P. Two new monoterpenoid α-pyrones from a fungus Nectria sp. HLS206 associated with the marine sponge Gelliodes carnosa. J. Asian Nat. Prod. Res. 2015, 17, 633–637. [Google Scholar] [CrossRef]
- Andolfi, A.; Boari, A.; Evidente, M.; Cimmino, A.; Vurro, M.; Ash, G.; Evidente, A. Gulypyrones A and B and Phomentrioloxins B and C Produced by Diaporthe gulyae, a Potential Mycoherbicide for Saffron Thistle (Carthamus lanatus). J. Nat. Prod. 2015, 78, 623–629. [Google Scholar] [CrossRef]
- Zhu, H.; Hua, X.-X.; Gong, T.; Pang, J.; Hou, Q.; Zhu, P. Hypocreaterpenes A and B, cadinane-type sesquiterpenes from a marine-derived fungus, Hypocreales sp. Phytochem. Lett. 2013, 6, 392–396. [Google Scholar] [CrossRef]
- Xi, J.; Yang, Z.; Xu, J.; Ge, M.; Chen, D. Study on the metabolites of endophytic fungus Colletotrichum sp. from Elaeagnus umbellata Thunb. Xibei Yaoxue Zazhi 2012, 27, 523–525. [Google Scholar] [CrossRef]
- Evidente, A.; Rodeva, R.; Andolfi, A.; Stoyanova, Z.; Perrone, C.; Motta, A. Phytotoxic polyketides produced by Phomopsis foeniculi, a strain isolated from diseased Bulgarian fennel. Eur. J. Plant. Pathol. 2011, 130, 173–182. [Google Scholar] [CrossRef]
- Guimaraes, D.O.; Borges, W.S.; Kawano, C.Y.; Ribeiro, P.H.; Goldman, G.H.; Nomizo, A.; Thiemann, O.H.; Oliva, G.; Lopes, N.P.; Pupo, M.T. Biological activities from extracts of endophytic fungi isolated from Viguiera arenaria and Tithonia diversifolia. Fems Immunol. Med. Microbiol. 2008, 52, 134–144. [Google Scholar] [CrossRef]
- Meister, J.; Weber, D.; Martino, V.; Sterner, O.; Anke, T. Phomopsidone, a novel depsidone from an endophyte of the medicinal plant Eupatorium arnottianum. Z. Naturforsch. C J. Biosci. 2007, 62, 11–15. [Google Scholar] [CrossRef]
- Weber, D.; Gorzalczany, S.; Martino, V.; Acevedo, C.; Sterner, O.; Anke, T. Metabolites from endophytes of the medicinal plant Erythrina crista-galli. Z. Naturforsch., C: J. Biosci. 2005, 60, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Thines, E.; Anke, H.; Sterner, O. Scytalols A, B, C, and D and other modulators of melanin biosynthesis from Scytalidium sp. 36–93. J. Antibiot. 1998, 51, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Avent, A.G.; Hanson, J.R.; Truneh, A. Two pyrones from Gliocladium vermoesenii. Phytochemistry 1992, 31, 1065–1066. [Google Scholar] [CrossRef]
- Claydon, N.; Grove, J.F.; Pople, M. Elm bark beetle boring and feeding deterrents from Phomopsis oblonga. Phytochemistry 1985, 24, 937–943. [Google Scholar] [CrossRef]
- Nair, M.S.R.; Carey, S.T. Metabolites of pyrenomycetes. II. Nectriapyrone, an antibiotic monoterpenoid. Tetrahedron Lett. 1975, 1655–1658. [Google Scholar] [CrossRef]
- Lu, X.; Xu, N.; Dai, H.-F.; Mei, W.-L.; Yang, Z.-X.; Pei, Y.-H. Three new compounds from endophytic fungus L10 of Cephalotaxus hainanensis. J. Asian Nat. Prod. Res. 2009, 11, 397–400. [Google Scholar] [CrossRef]
- Motoyama, T.; Nogawa, T.; Hayashi, T.; Hirota, H.; Osada, H. Induction of Nectriapyrone Biosynthesis in the Rice Blast Fungus Pyricularia oryzae by Disturbance of the Two-Component Signal Transduction System. Chembiochem 2019, 20, 693–700. [Google Scholar] [CrossRef]
- West, A.H.; Stock, A.M. Histidine kinases and response regulator proteins in two-component signaling systems. Trends Biochem. Sci. 2001, 26, 369–376. [Google Scholar] [CrossRef]
- Catlett, N.L.; Yoder, O.C.; Turgeon, B.G. Whole-genome analysis of two-component signal transduction genes in fungal pathogens. Eukaryot. Cell 2003, 2, 1151–1161. [Google Scholar] [CrossRef] [Green Version]
- Abramson, H.N.; Wormser, H.C. Synthesis of nectriapyrone. J. Heterocycl. Chem. 1981, 18, 363–366. [Google Scholar] [CrossRef]
- Hammerschmidt, L.; Debbab, A.; Ngoc, T.D.; Wray, V.; Hemphil, C.P.; Lin, W.; Broetz-Oesterhelt, H.; Kassack, M.U.; Proksch, P.; Aly, A.H. Polyketides from the mangrove-derived endophytic fungus Acremonium strictum. Tetrahedron Lett. 2014, 55, 3463–3468. [Google Scholar] [CrossRef]
- Cai, R.; Chen, S.; Liu, Z.; Tan, C.; Huang, X.; She, Z. A new alpha-pyrone from the mangrove endophytic fungus Phomopsis sp. HNY29-2B. Nat. Prod. Res. 2017, 31, 124–130. [Google Scholar] [CrossRef]
- Burkhardt, I.; Dickschat, J.S. Synthesis and Absolute Configuration of Natural 2-Pyrones. Eur. J. Org. Chem. 2018, 2018, 3144–3157. [Google Scholar] [CrossRef]
- Janevska, S.; Arndt, B.; Niehaus, E.M.; Burkhardt, I.; Rosler, S.M.; Brock, N.L.; Humpf, H.U.; Dickschat, J.S.; Tudzynski, B. Gibepyrone Biosynthesis in the Rice Pathogen Fusarium fujikuroi Is Facilitated by a Small Polyketide Synthase Gene Cluster. J. Biol. Chem. 2016, 291, 27403–27420. [Google Scholar] [CrossRef] [Green Version]
- Petersen, F.; Zahner, H.; Metzger, J.W.; Freund, S.; Hummel, R.P. Germicidin, an autoregulative germination inhibitor of Streptomyces viridochromogenes NRRL B-1551. J. Antibiot. 1993, 46, 1126–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, Y.; Matsumoto, D.; Kawaide, H.; Natsume, M. Physiological role of germicidins in spore germination and hyphal elongation in Streptomyces coelicolor A3(2). J. Antibiot. 2011, 64, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ding, L.; Hertweck, C. A branched extender unit shared between two orthogonal polyketide pathways in an endophyte. Angew. Chem. (Int. Ed. Engl.) 2011, 50, 4667–4670. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Barona-Gomez, F.; Corre, C.; Xiang, L.; Udwary, D.W.; Austin, M.B.; Noel, J.P.; Moore, B.S.; Challis, G.L. Type III polyketide synthase beta-ketoacyl-ACP starter unit and ethylmalonyl-CoA extender unit selectivity discovered by Streptomyces coelicolor genome mining. J. Am. Chem. Soc. 2006, 128, 14754–14755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGlacken, G.P.; Fairlamb, I.J. 2-Pyrone natural products and mimetics: Isolation, characterisation and biological activity. Nat. Prod. Rep. 2005, 22, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Schaberle, T.F. Biosynthesis of alpha-pyrones. Beilstein J. Org. Chem. 2016, 12, 571–588. [Google Scholar] [CrossRef]
- Brachmann, A.O.; Brameyer, S.; Kresovic, D.; Hitkova, I.; Kopp, Y.; Manske, C.; Schubert, K.; Bode, H.B.; Heermann, R. Pyrones as bacterial signaling molecules. Nat. Chem. Biol. 2013, 9, 573–578. [Google Scholar] [CrossRef]
- Turkkan, M.; Andolfi, A.; Zonno, M.C.; Erper, I.; Perrone, C.; Cimmino, A.; Vurro, M.; Evidente, A. Phytotoxins produced by Pestalotiopsis guepinii, the causal agent of hazelnut twig blight. Phytopathol. Mediterr. 2011, 50, 154–158. [Google Scholar]
- Lee, I.-K.; Yun, B.-S.; Oh, S.; Kim, Y.-H.; Lee, M.-K.; Yoo, I.-D. 5-Methylmellein and nectriapyrone, two new monoamine oxidase inhibitors. Med. Sci. Res. 1999, 27, 463–465. [Google Scholar]
- Rosett, T.; Sankhala, R.H.; Stickings, C.E.; Taylor, M.E.U.; Thomas, R. Studies in the biochemistry of micro-organisms. 103. Metabolites of Alternaria tenuis Auct.: Culture filtrate products. Biochem. J. 1957, 67, 390–400. [Google Scholar] [CrossRef] [Green Version]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Siegel, D.; Rasenko, T.; Koch, M.; Nehls, I. Determination of the Alternaria mycotoxin tenuazonic acid in cereals by high-performance liquid chromatography-electrospray ionization ion-trap multistage mass spectrometry after derivatization with 2,4-dinitrophenylhydrazine. J. Chromatogr. A 2009, 1216, 4582–4588. [Google Scholar] [CrossRef] [PubMed]
- Gross, M.; Curtui, V.; Ackermann, Y.; Latif, H.; Usleber, E. Enzyme immunoassay for tenuazonic acid in apple and tomato products. J. Agric. Food Chem. 2011, 59, 12317–12322. [Google Scholar] [CrossRef]
- Lohrey, L.; Marschik, S.; Cramer, B.; Humpf, H.U. Large-scale synthesis of isotopically labeled 13C2-tenuazonic acid and development of a rapid HPLC-MS/MS method for the analysis of tenuazonic acid in tomato and pepper products. J. Agric. Food Chem. 2013, 61, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Umetsu, N.; Kaji, J.; Tamari, K. Investigation on the toxin production by several blast fungus strains and isolation of tenuazonic acid as a novel toxin. Agr. Biol. Chem. 1972, 36, 859–866. [Google Scholar] [CrossRef]
- Steyn, P.S.; Rabie, C.J. Characterization of magnesium and calcium tenuazonate from Phoma sorghina. Phytochemistry 1976, 15, 1977–1979. [Google Scholar] [CrossRef]
- Lebrun, M.H.; Dutfoy, F.; Gaudemer, F.; Kunesch, G.; Gaudemer, A. Detection and quantification of the fungal phytotoxin tenuazonic acid produced by Pyricularia oryzae. Phytochemistry 1990, 29, 3777–3783. [Google Scholar] [CrossRef]
- Miller, F.A.; Rightsel, W.A.; Sloan, B.J.; Ehrlich, J.; French, J.C.; Bartz, Q.R. Antiviral activity of tenuazonic acid. Nature 1963, 200, 1338–1339. [Google Scholar] [CrossRef]
- Smith, E.R.; Fredrickson, T.N.; Hadidian, Z. Toxic effects of the sodium and the N,N′-dibenzylethylenediamine salts of tenuazonic acid. Cancer Chemother. Rep. 1968, 52, 579–585. [Google Scholar]
- Asam, S.; Rychlik, M. Potential health hazards due to the occurrence of the mycotoxin tenuazonic acid in infant food. Eur. Food Res. Technol. 2013, 236, 491–497. [Google Scholar] [CrossRef]
- Shigeura, H.T.; Gordon, C.N. The biological activity of tenuazonic acidBIOLOGICAL ACTIVITY OF TENUAZONIC ACID. Biochemistry 1963, 2, 1132–1137. [Google Scholar] [CrossRef]
- Gitterman, C.O. Antitumor, cytotoxic, and antibacterial activities of tenuazonic acid and congeneric tetramic acids. J. Med. Chem. 1965, 8, 483–486. [Google Scholar] [CrossRef]
- Lebrun, M.H.; Nicolas, L.; Boutar, M.; Gaudemer, F.; Ranomenjanahary, S.; Gaudemer, A. Relationships between the structure and the phytotoxicity of the fungal toxin tenuazonic acid. Phytochemistry 1988, 27, 77–84. [Google Scholar] [CrossRef]
- Aver’yanov, A.A.; Lapikova, V.P.; Lebrun, M.H. Tenuazonic acid, toxin of rice blast fungus, induces disease resistance and reactive oxygen production in plants. Russ. J. Plant. Physiol. 2007, 54, 749–754. [Google Scholar] [CrossRef]
- Chen, S.; Xu, X.; Dai, X.; Yang, C.; Qiang, S. Identification of tenuazonic acid as a novel type of natural photosystem II inhibitor binding in Q(B)-site of Chlamydomonas reinhardtii. Biochim. Biophys. Acta 2007, 1767, 306–318. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Yin, C.; Qiang, S.; Zhou, F.; Dai, X. Chloroplastic oxidative burst induced by tenuazonic acid, a natural photosynthesis inhibitor, triggers cell necrosis in Eupatorium adenophorum Spreng. Biochim. Biophys. Acta 2010, 1797, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Kim, C.; Lee, J.M.; Lee, H.A.; Fei, Z.; Wang, L.; Apel, K. Blocking the QB-binding site of photosystem II by tenuazonic acid, a non-host-specific toxin of Alternaria alternata, activates singlet oxygen-mediated and EXECUTER-dependent signalling in Arabidopsis. Plant. Cell Environ. 2015, 38, 1069–1080. [Google Scholar] [CrossRef]
- Chen, S.; Qiang, S. Recent advances in tenuazonic acid as a potential herbicide. Pestic Biochem. Physiol. 2017, 143, 252–257. [Google Scholar] [CrossRef]
- Bjørk, P.K.; Rasmussen, S.A.; Gjetting, S.K.; Havshøi, N.W.; Petersen, T.I.; Ipsen, J.; Larsen, T.O.; Fuglsang, A.T. Tenuazonic acid from Stemphylium loti inhibits the plant plasma membrane H(+) -ATPase by a mechanism involving the C-terminal regulatory domain. New Phytol. 2020, 226, 770–784. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.S.; Motoyama, T.; Osada, H. Biosynthesis of the mycotoxin tenuazonic acid by a fungal NRPS-PKS hybrid enzyme. Nat. Commun. 2015, 6, 8758. [Google Scholar] [CrossRef] [Green Version]
- Motoyama, T.; Osada, H. Biosynthetic approaches to creating bioactive fungal metabolites: Pathway engineering and activation of secondary metabolism. Bioorg Med. Chem. Lett. 2016, 26, 5843–5850. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.S.; Motoyama, T.; Osada, H. Regulatory Mechanism of Mycotoxin Tenuazonic Acid Production in Pyricularia oryzae. Acs Chem. Biol. 2017, 12, 2270–2274. [Google Scholar] [CrossRef]
- Yun, C.S.; Nishimoto, K.; Motoyama, T.; Shimizu, T.; Hino, T.; Dohmae, N.; Nagano, S.; Osada, H. Unique features of the ketosynthase domain in a non-ribosomal peptide synthetase-polyketide synthase hybrid enzyme, tenuazonic acid synthetase 1. J. Biol. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, M.A.; Walsh, C.T. Assembly-line enzymology for polyketide and nonribosomal Peptide antibiotics: Logic, machinery, and mechanisms. Chem. Rev. 2006, 106, 3468–3496. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Nonaka, T.; Fujii, I. Fungal type III polyketide synthases. Nat. Prod. Rep. 2014, 31, 1306–1317. [Google Scholar] [CrossRef] [PubMed]
- Böhnert, H.U.; Fudal, I.; Dioh, W.; Tharreau, D.; Notteghem, J.-L.; Lebrun, M.-H. A putative polyketide synthase/peptide synthetase from Magnaporthe grisea signals pathogen attack to resistant rice. Plant. Cell 2004, 16, 2499–2513. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Cox, R.J.; Lazarus, C.M.; Simpson, T.T. Fusarin C biosynthesis in Fusarium moniliforme and Fusarium venenatum. Chembiochem 2004, 5, 1196–1203. [Google Scholar] [CrossRef]
- Eley, K.L.; Halo, L.M.; Song, Z.; Powles, H.; Cox, R.J.; Bailey, A.M.; Lazarus, C.M.; Simpson, T.J. Biosynthesis of the 2-pyridone tenellin in the insect pathogenic fungus Beauveria bassiana. Chembiochem 2007, 8, 289–297. [Google Scholar] [CrossRef]
- Boettger, D.; Hertweck, C. Molecular diversity sculpted by fungal PKS-NRPS hybrids. Chembiochem 2013, 14, 28–42. [Google Scholar] [CrossRef]
- Fisch, K.M. Biosynthesis of natural products by microbial iterative hybrid PKS–NRPS. RSC Adv. 2013, 3, 18228–18247. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Zaleta-Rivera, K.; Zhu, X.; Huffman, J.; Millet, J.C.; Harris, S.D.; Yuen, G.; Li, X.-C.; Du, L. Structure and biosynthesis of heat-stable antifungal factor (HSAF), a broad-spectrum antimycotic with a novel mode of action. Antimicrob. Agents Chemother. 2007, 51, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blodgett, J.A.; Oh, D.C.; Cao, S.; Currie, C.R.; Kolter, R.; Clardy, J. Common biosynthetic origins for polycyclic tetramate macrolactams from phylogenetically diverse bacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 11692–11697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silakowski, B.; Schairer, H.U.; Ehret, H.; Kunze, B.; Weinig, S.; Nordsiek, G.; Brandt, P.; Blöcker, H.; Höfle, G.; Beyer, S.; et al. New lessons for combinatorial biosynthesis from myxobacteria. The myxothiazol biosynthetic gene cluster of Stigmatella aurantiaca DW4/3-1. J. Biol. Chem. 1999, 274, 37391–37399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Sánchez, C.; Shen, B. Hybrid peptide-polyketide natural products: Biosynthesis and prospects toward engineering novel molecules. Metab. Eng. 2001, 3, 78–95. [Google Scholar] [CrossRef]
- Tang, G.L.; Cheng, Y.Q.; Shen, B. Leinamycin biosynthesis revealing unprecedented architectural complexity for a hybrid polyketide synthase and nonribosomal peptide synthetase. Chem. Biol. 2004, 11, 33–45. [Google Scholar] [CrossRef]
- Simunovic, V.; Zapp, J.; Rachid, S.; Krug, D.; Meiser, P.; Müller, R. Myxovirescin A biosynthesis is directed by hybrid polyketide synthases/nonribosomal peptide synthetase, 3-hydroxy-3-methylglutaryl-CoA synthases, and trans-acting acyltransferases. Chembiochem 2006, 7, 1206–1220. [Google Scholar] [CrossRef]
- Gerc, A.J.; Song, L.; Challis, G.L.; Stanley-Wall, N.R.; Coulthurst, S.J. The Insect Pathogen Serratia marcescens Db10 Uses a Hybrid Non-Ribosomal Peptide Synthetase-Polyketide Synthase to Produce the Antibiotic Althiomycin. PLoS ONE 2012, 7, e44673. [Google Scholar] [CrossRef] [Green Version]
- Müller, S.; Garcia-Gonzalez, E.; Mainz, A.; Hertlein, G.; Heid, N.C.; Mösker, E.; van den Elst, H.; Overkleeft, H.S.; Genersch, E.; Süssmuth, R.D. Paenilamicin: Structure and Biosynthesis of a Hybrid Nonribosomal Peptide/Polyketide Antibiotic from the Bee Pathogen Paenibacillus larvae. Angew. Chem. Int. Ed. 2014, 53, 10821–10825. [Google Scholar] [CrossRef]
- Stickings, C.E.; Townsend, R.J. Studies in the biochemistry of micro-organisms. 108. Metabolites of Alternaria tenuis Auct.: The biosynthesis of tenuazonic acid. Biochem. J. 1961, 78, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Collemare, J.; Billard, A.; Bohnert, H.U.; Lebrun, M.H. Biosynthesis of secondary metabolites in the rice blast fungus Magnaporthe grisea: The role of hybrid PKS-NRPS in pathogenicity. Mycol. Res. 2008, 112, 207–215. [Google Scholar] [CrossRef]
- Trauger, J.W.; Kohli, R.M.; Mootz, H.D.; Marahiel, M.A.; Walsh, C.T. Peptide cyclization catalysed by the thioesterase domain of tyrocidine synthetase. Nature 2000, 407, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Haynes, S.W.; Ames, B.D.; Wang, P.; Vien, L.P.; Walsh, C.T.; Tang, Y. Cyclization of fungal nonribosomal peptides by a terminal condensation-like domain. Nat. Chem. Biol. 2012, 8, 823–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. (Int. Ed. Engl.) 2009, 48, 4688–4716. [Google Scholar] [CrossRef] [PubMed]
- He, H.-Y.; Tang, M.-C.; Zhang, F.; Tang, G.-L. Cis-Double Bond Formation by Thioesterase and Transfer by Ketosynthase in FR901464 Biosynthesis. J. Am. Chem. Soc. 2014, 136, 4488–4491. [Google Scholar] [CrossRef]
- Bretschneider, T.; Heim, J.B.; Heine, D.; Winkler, R.; Busch, B.; Kusebauch, B.; Stehle, T.; Zocher, G.; Hertweck, C. Vinylogous chain branching catalysed by a dedicated polyketide synthase module. Nature 2013, 502, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Katsuyama, Y.; Ohnishi, Y. Type III polyketide synthases in microorganisms. Methods Enzymol. 2012, 515, 359–377. [Google Scholar] [CrossRef]
- Tanovic, A.; Samel, S.A.; Essen, L.O.; Marahiel, M.A. Crystal structure of the termination module of a nonribosomal peptide synthetase. Science 2008, 321, 659–663. [Google Scholar] [CrossRef] [Green Version]
- Kao, C.M.; Pieper, R.; Cane, D.E.; Khosla, C. Evidence for Two Catalytically Independent Clusters of Active Sites in a Functional Modular Polyketide Synthase. Biochemistry 1996, 35, 12363–12368. [Google Scholar] [CrossRef]
- Dutta, S.; Whicher, J.R.; Hansen, D.A.; Hale, W.A.; Chemler, J.A.; Congdon, G.R.; Narayan, A.R.H.; Håkansson, K.; Sherman, D.H.; Smith, J.L.; et al. Structure of a modular polyketide synthase. Nature 2014, 510, 512–517. [Google Scholar] [CrossRef] [Green Version]
- Weissman, K.J. Uncovering the structures of modular polyketide synthases. Nat. Prod. Rep. 2015, 32, 436–453. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Butchko, R.A.; Fernandes, M.; Keller, N.P.; Leonard, T.J.; Adams, T.H. Conservation of structure and function of the aflatoxin regulatory gene aflR from Aspergillus nidulans and A. flavus. Curr. Genet. 1996, 29, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Chung, D.; Balajee, S.A.; Marr, K.A.; Andes, D.; Nielsen, K.F.; Frisvad, J.C.; Kirby, K.A.; Keller, N.P. GliZ, a transcriptional regulator of gliotoxin biosynthesis, contributes to Aspergillus fumigatus virulence. Infect. Immun. 2006, 74, 6761–6768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Kinoshita, H.; Nihira, T. Identification and in vivo functional analysis by gene disruption of ctnA, an activator gene involved in citrinin biosynthesis in Monascus purpureus. Appl Environ. Microbiol. 2007, 73, 5097–5103. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P.; Desjardins, A.E. Tri6 encodes an unusual zinc finger protein involved in regulation of trichothecene biosynthesis in Fusarium sporotrichioides. Appl. Environ. Microbiol. 1995, 61, 1923–1930. [Google Scholar] [CrossRef] [Green Version]
- Perrin, R.M.; Fedorova, N.D.; Bok, J.W.; Cramer, R.A., Jr.; Wortman, J.R.; Kim, H.S.; Nierman, W.C.; Keller, N.P. Transcriptional Regulation of Chemical Diversity in Aspergillus fumigatus by LaeA. PLoS Pathog. 2007, 3, e50. [Google Scholar] [CrossRef] [Green Version]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.J.; Keller, N.P.; Yu, J.H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Yin, W.; Keller, N.P. Transcriptional regulatory elements in fungal secondary metabolism. J. Microbiol. 2011, 49, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef]
- Bok, J.W.; Keller, N.P. 2 Insight into Fungal Secondary Metabolism from Ten Years of LaeA Research. In Biochemistry and Molecular Biology; Hoffmeister, D., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 21–29. [Google Scholar] [CrossRef]
- Ninomiya, A.; Urayama, S.I.; Suo, R.; Itoi, S.; Fuji, S.I.; Moriyama, H.; Hagiwara, D. Mycovirus-Induced Tenuazonic Acid Production in a Rice Blast Fungus Magnaporthe oryzae. Front. Microbiol. 2020, 11, 1641. [Google Scholar] [CrossRef]
- Song, Z.; Bakeer, W.; Marshall, J.W.; Yakasai, A.A.; Khalid, R.M.; Collemare, J.; Skellam, E.; Tharreau, D.; Lebrun, M.H.; Lazarus, C.M.; et al. Heterologous expression of the avirulence gene ACE1 from the fungal rice pathogen Magnaporthe oryzae. Chem. Sci. 2015, 6, 4837–4845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, P.; Ghosh, S.; Roy-Barman, S. MoLAEA Regulates Secondary Metabolism in Magnaporthe oryzae. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patkar, R.N.; Benke, P.I.; Qu, Z.; Chen, Y.Y.; Yang, F.; Swarup, S.; Naqvi, N.I. A fungal monooxygenase-derived jasmonate attenuates host innate immunity. Nat. Chem. Biol. 2015, 11, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.J.; Shimono, M.; Sugano, S.; Kojima, M.; Yazawa, K.; Yoshida, R.; Inoue, H.; Hayashi, N.; Sakakibara, H.; Takatsuji, H. Abscisic acid interacts antagonistically with salicylic acid signaling pathway in rice-Magnaporthe grisea interaction. Mol. Plant. Microbe Interact. 2010, 23, 791–798. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.J.; Shimono, M.; Sugano, S.; Kojima, M.; Liu, X.; Inoue, H.; Sakakibara, H.; Takatsuji, H. Cytokinins act synergistically with salicylic acid to activate defense gene expression in rice. Mol. Plant. Microbe Interact. 2013, 26, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, E.; Koga, H.; Mori, M.; Mori, M. Auxin Production by the Rice Blast Fungus and its Localization in Host Tissue. J. Phytopathol. 2011, 159, 522–530. [Google Scholar] [CrossRef]
- Spence, C.A.; Lakshmanan, V.; Donofrio, N.; Bais, H.P. Crucial Roles of Abscisic Acid Biogenesis in Virulence of Rice Blast Fungus Magnaporthe oryzae. Front. Plant. Sci 2015, 6, 1082. [Google Scholar] [CrossRef] [Green Version]
- Chanclud, E.; Kisiala, A.; Emery, N.R.; Chalvon, V.; Ducasse, A.; Romiti-Michel, C.; Gravot, A.; Kroj, T.; Morel, J.B. Cytokinin Production by the Rice Blast Fungus Is a Pivotal Requirement for Full Virulence. PLoS Pathog. 2016, 12, e1005457. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Chemical structures of secondary metabolites from the rice blast fungus P. oryzae.

Figure 2.
Melanin biosynthetic pathway of the rice blast fungus P. oryzae.

Figure 3.
Structures of the pyriculols and a related compound.

Figure 4.
Nectriapyrones and related compounds.

Figure 5.
Tenuazonic acid (TeA) biosynthesis. (a) Comparison of domain structures. (b) Biosynthetic pathway of TeA.
Figure 5.
Tenuazonic acid (TeA) biosynthesis. (a) Comparison of domain structures. (b) Biosynthetic pathway of TeA.

Figure 6.
Regulation of TeA production in P. oryzae.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Motoyama, T. Secondary Metabolites of the Rice Blast Fungus Pyricularia oryzae: Biosynthesis and Biological Function. Int. J. Mol. Sci. 2020, 21, 8698. https://doi.org/10.3390/ijms21228698
AMA Style
Motoyama T. Secondary Metabolites of the Rice Blast Fungus Pyricularia oryzae: Biosynthesis and Biological Function. International Journal of Molecular Sciences. 2020; 21(22):8698. https://doi.org/10.3390/ijms21228698
Chicago/Turabian StyleMotoyama, Takayuki. 2020. "Secondary Metabolites of the Rice Blast Fungus Pyricularia oryzae: Biosynthesis and Biological Function" International Journal of Molecular Sciences 21, no. 22: 8698. https://doi.org/10.3390/ijms21228698
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.