Germinal Matrix-Intraventricular Hemorrhage of the Preterm Newborn and Preclinical Models: Inflammatory Considerations


Abstract
:1. The preterm newborn
1.1. Etiology and Consequences of Prematurity
1.2. Neurological Complications of the PTNB
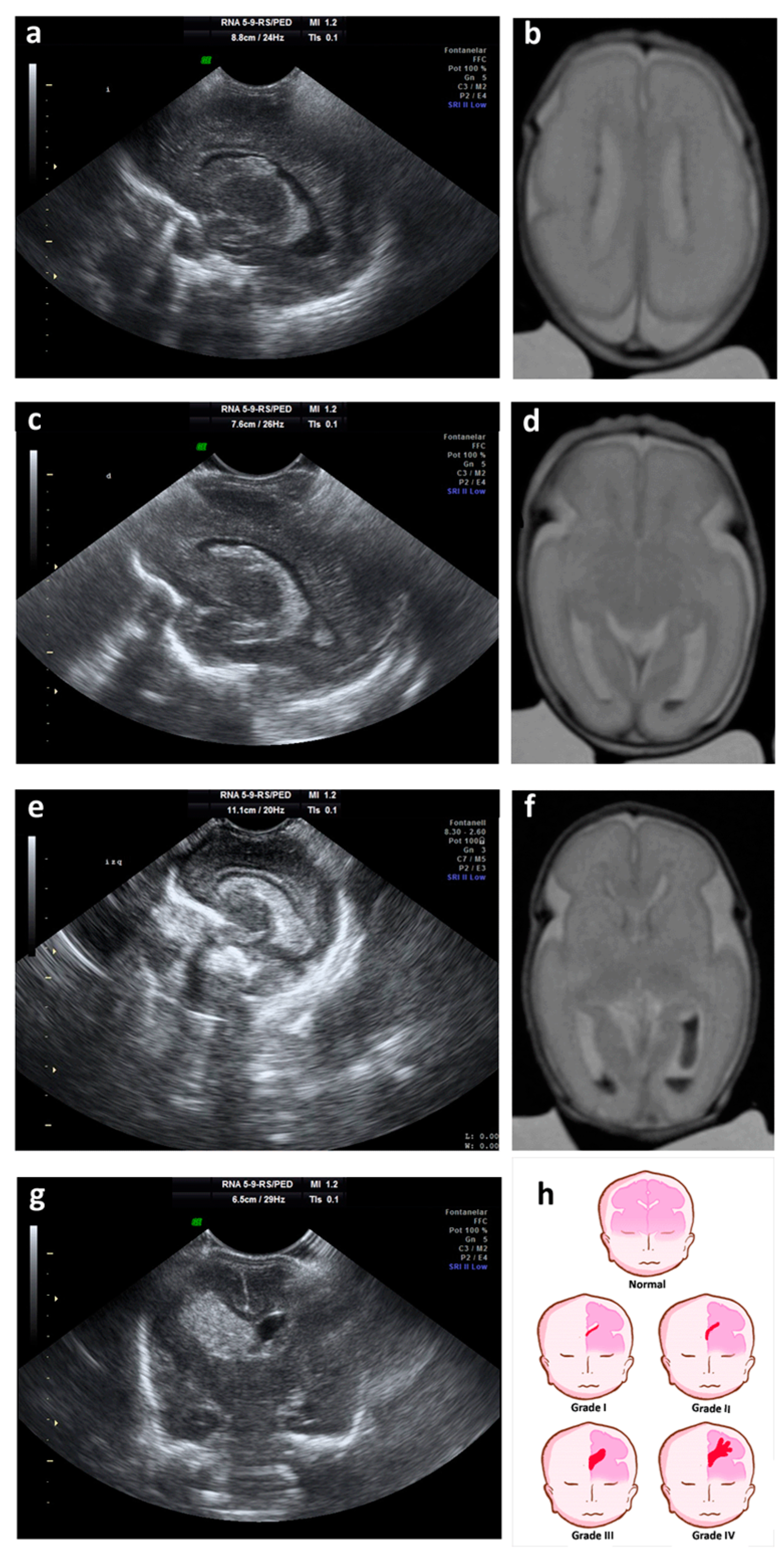
2. Germinal Matrix-Intraventricular Hemorrhage
- -
- Grade I: hemorrhage localized only in the subependymal GM (caudo thalamic groove).
- -
- Grade II: intraventricular hemorrhage without ventricular dilation.
- -
- Grade III: intraventricular hemorrhage with ventricular dilation.
- -
- Grade IV: parenchymal hemorrhage that corresponds to periventricular venous infarctions with hemorrhagic evolution [5].
2.1. GM-IVH Neurodevelopmental Disabilities
2.2. GM-IVH Associated Brain Damage
2.3. Neuroinflammation and Microglia in the GM-IVH
3. Animal Models of GM-IVH
3.1. Genetically Modified Models
3.2. Lesion-Induced Models
3.2.1. Glycerol-Induced GM-IVH in Rabbits
3.2.2. Blood and Blood Derivates-Induced GM-IVH in Rodents
3.2.3. Collagenase-Induced GM-IVH in Rodents
3.3. Neuroinflammation and Microglia in Animal Models of GM-IVH
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BBB | Bloodbrain barrier |
| CNS | central nervous system |
| CSF | cerebrospinal fluid |
| GM | germinal matrix |
| GM-IVH | germinal matrix-intraventricular hemorrhage |
| PTNB | preterm newborn |
| TLR | Toll-like receptor |
| VEGF | Vascular endothelial growth factor |
References
- Rellan, S.; Garcia de Ribera, C.; Aragon, M.P. El recien nacido pretermino. Protoc. Diagn. Ter. AEP Neonatol. 2008, 8, 69–75. [Google Scholar]
- Blencowe, H.; Cousens, S.; Chou, D.; Oestergaard, M.; Say, L.; Moller, A.; Kinney, M.; Lawn, J. Born Too Soon: The global epidemiology of 15 million preterm births. Reprod. Health 2013, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hee Chung, E.; Chou, J.; Brown, K.A. Neurodevelopmental outcomes of preterm infants: A recent literature review. Transl. Pediatr 2020, 9, S3–S8. [Google Scholar] [CrossRef] [PubMed]
- Tucker, J.; McGuire, W. Epidemiology of preterm birth. BMJ 2004, 329, 675–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerisola, A.; Baltar, F.; Ferrán, C.; Turcatti, E. Mecanismos de lesión cerebral en niños prematuros. MEDICINA (Buenos Aires) 2019, 79, 10–14. [Google Scholar]
- Hollanders, J.J.; Schaefer, N.; van der Pal, S.M.; Oosterlaan, J.; Rotteveel, J.; Finken, M.J.J.; on behalf of the Dutch, P.-C.S.G. Long-Term Neurodevelopmental and Functional Outcomes of Infants Born Very Preterm and/or with a Very Low Birth Weight. Neonatology 2019, 115, 310–319. [Google Scholar] [CrossRef]
- da Silva, L.S.; Ribeiro, G.E.; Montovani, J.C.; Silva, D. The effect of peri-intraventricular hemorrhage on the auditory pathway of infants. Int. J. Pediatr. Otorhinolaryngol. 2018, 112, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Hochart, A.; Nuytten, A.; Pierache, A.; Bauters, A.; Rauch, A.; Wibaut, B.; Susen, S.; Goudemand, J. Hemostatic profile of infants with spontaneous prematurity: Can we predict intraventricular hemorrhage development? Ital. J. Pediatr. 2019, 45, 113. [Google Scholar] [CrossRef]
- Walani, S.R. Global burden of preterm birth. Int. J. Gynaecol. Obstet. 2020, 150, 31–33. [Google Scholar] [CrossRef]
- Gotardo, J.W.; Volkmer, N.F.V.; Stangler, G.P.; Dornelles, A.D.; Bohrer, B.B.A.; Carvalho, C.G. Impact of peri-intraventricular haemorrhage and periventricular leukomalacia in the neurodevelopment of preterms: A systematic review and meta-analysis. PLoS ONE 2019, 14, e0223427. [Google Scholar] [CrossRef]
- Mendoza, L.A.; Claros, D.I.; Mendoza, L.I.; Arias, M.D.; Peñaranda, C.B. Epidemiología de la prematuridad, sus determinantes y prevención del parto prematuro. Rev. Chil. Obstet. Ginecol. 2016, 81, 330–342. [Google Scholar] [CrossRef] [Green Version]
- Stecher, J.F.; Pavlovic, M.; Contreras, C.; Carvajal, J. Prematuros tardíos: Estudio clínico retrospectivo de sus causas y consecuencias. Rev. Chil. Obstet. Ginecol. 2015, 80, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Gyamfi-Bannerman, C.; Fuchs, K.M.; Young, O.M.; Hoffman, M.K. Nonspontaneous late preterm birth: Etiology and outcomes. Am. J. Obstet. Gynecol. 2011, 205, 456.e1–456.e6. [Google Scholar] [CrossRef] [PubMed]
- Holland, M.G.; Refuerzo, J.S.; Ramin, S.M.; Saade, G.R.; Blackwell, S.C. Late preterm birth: How often is it avoidable? Am. J. Obstet. Gynecol. 2009, 201, 404.e1–404.e4. [Google Scholar] [CrossRef]
- Cormack, B.E.; Harding, J.E.; Miller, S.P.; Bloomfield, F.H. The Influence of Early Nutrition on Brain Growth and Neurodevelopment in Extremely Preterm Babies: A Narrative Review. Nutrients 2019, 11, 2029. [Google Scholar] [CrossRef] [Green Version]
- Twilhaar, E.S.; Wade, R.M.; de Kieviet, J.F.; van Goudoever, J.B.; van Elburg, R.M.; Oosterlaan, J. Cognitive Outcomes of Children Born Extremely or Very Preterm Since the 1990s and Associated Risk Factors: A Meta-analysis and Meta-regression. JAMA Pediatr. 2018, 172, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.P.; Chawanpaiboon, S.; Moller, A.B.; Watananirun, K.; Bonet, M.; Lumbiganon, P. The global epidemiology of preterm birth. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 52, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Benitz, W.E.; Committee on Fetus and Newborn. Patent Ductus Arteriosus in Preterm Infants. Pediatrics 2016, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harkin, P.; Marttila, R.; Pokka, T.; Saarela, T.; Hallman, M. Survival analysis of a cohort of extremely preterm infants born in Finland during 2005–2013. Matern. Fetal Neonatal Med. 2019, 1–7. [Google Scholar] [CrossRef]
- D’Apremont, I.; Marshall, G.; Musalem, C.; Mariani, G.; Musante, G.; Bancalari, A.; Fabres, J.; Mena, P.; Zegarra, J.; Tavosnanska, J.; et al. Trends in Perinatal Practices and Neonatal Outcomes of Very Low Birth Weight Infants during a 16-year Period at NEOCOSUR Centers. J. Pediatr. 2020, 225, 44–50.e1. [Google Scholar] [CrossRef]
- Cabañas, F.; Pellicer, A. Lesion cerebral en el niño prematuro. Protoc. Diagn. Ter. AEP Neonatol. 2008, 27, 253–269. [Google Scholar]
- Sancak, S.; Gursoy, T.; Karatekin, G.; Ovali, F. Effect of Intraventricular Hemorrhage on Cerebellar Growth in Preterm Neonates. Cerebellum 2017, 16, 89–94. [Google Scholar] [CrossRef]
- Bolisetty, S.; Dhawan, A.; Abdel-Latif, M.; Bajuk, B.; Stack, J.; Oei, J.-L.; Lui, K.; New South Wales and Australian Capital Territory Neonatal Intensive Care Units’ Data Collection. Intraventricular hemorrhage and neurodevelopmental outcomes in extreme preterm infants. Pediatrics 2014, 133, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Hinojosa-Rodriguez, M.; Harmony, T.; Carrillo-Prado, C.; Van Horn, J.D.; Irimia, A.; Torgerson, C.; Jacokes, Z. Clinical neuroimaging in the preterm infant: Diagnosis and prognosis. NeuroImage Clin. 2017, 16, 355–368. [Google Scholar] [CrossRef]
- Dorner, R.A.; Burton, V.J.; Allen, M.C.; Robinson, S.; Soares, B.P. Preterm neuroimaging and neurodevelopmental outcome: A focus on intraventricular hemorrhage, post-hemorrhagic hydrocephalus, and associated brain injury. J. Perinatol. Off. J. Calif. Perinat. Assoc. 2018, 38, 1431–1443. [Google Scholar] [CrossRef] [PubMed]
- Radic, J.A.; Vincer, M.; McNeely, P.D. Outcomes of intraventricular hemorrhage and posthemorrhagic hydrocephalus in a population-based cohort of very preterm infants born to residents of Nova Scotia from 1993 to 2010. J. Neurosurg. Pediatr. 2015, 15, 580–588. [Google Scholar] [CrossRef] [Green Version]
- Valdez Sandoval, P.; Hernandez Rosales, P.; Quinones Hernandez, D.G.; Chavana Naranjo, E.A.; Garcia Navarro, V. Intraventricular hemorrhage and posthemorrhagic hydrocephalus in preterm infants: Diagnosis, classification, and treatment options. Child’s Nerv. Syst. Off. J. Int. Soc. Pediatr. Neurosurg. 2019, 35, 917–927. [Google Scholar] [CrossRef]
- Cheng, X.R.; Xia, P.G.; Shi, Z.Y.; Xu, Q.Y.; Luo, C.H.; Lei, M.Y.; Zhang, Q. Increased risk of intracranial hemorrhage in preterm infants with OPRM1 gene A118G polymorphism. Ann. Transl. Med. 2019, 7, 478. [Google Scholar] [CrossRef]
- Mukerji, A.; Shah, V.; Shah, P.S. Periventricular/Intraventricular Hemorrhage and Neurodevelopmental Outcomes: A Meta-analysis. Pediatrics 2015, 136, 1132–1143. [Google Scholar] [CrossRef] [Green Version]
- Romantsik, O.; Agyemang, A.A.; Sveinsdottir, S.; Rutardottir, S.; Holmqvist, B.; Cinthio, M.; Morgelin, M.; Gumus, G.; Karlsson, H.; Hansson, S.R.; et al. The heme and radical scavenger alpha1-microglobulin (A1M) confers early protection of the immature brain following preterm intraventricular hemorrhage. J. Neuroinflammation 2019, 16, 122. [Google Scholar] [CrossRef] [Green Version]
- Christian, E.A.; Jin, D.L.; Attenello, F.; Wen, T.; Cen, S.; Mack, W.J.; Krieger, M.D.; McComb, J.G. Trends in hospitalization of preterm infants with intraventricular hemorrhage and hydrocephalus in the United States, 2000–2010. J. Neurosurg. Pediatr. 2016, 17, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Segado-Arenas, A.; Infante-Garcia, C.; Benavente-Fernandez, I.; Sanchez-Sotano, D.; Ramos-Rodriguez, J.J.; Alonso-Ojembarrena, A.; Lubian-Lopez, S.; Garcia-Alloza, M. Cognitive Impairment and Brain and Peripheral Alterations in a Murine Model of Intraventricular Hemorrhage in the Preterm Newborn. Mol. Neurobiol. 2018, 55, 4896–4910. [Google Scholar] [CrossRef]
- Morita, T.; Morimoto, M.; Yamada, K.; Hasegawa, T.; Morioka, S.; Kidowaki, S.; Moroto, M.; Yamashita, S.; Maeda, H.; Chiyonobu, T.; et al. Low-grade intraventricular hemorrhage disrupts cerebellar white matter in preterm infants: Evidence from diffusion tensor imaging. Neuroradiology 2015, 57, 507–514. [Google Scholar] [CrossRef]
- Gilard, V.; Chadie, A.; Ferracci, F.X.; Brasseur-Daudruy, M.; Proust, F.; Marret, S.; Curey, S. Post hemorrhagic hydrocephalus and neurodevelopmental outcomes in a context of neonatal intraventricular hemorrhage: An institutional experience in 122 preterm children. BMC Pediatr. 2018, 18, 288. [Google Scholar] [CrossRef]
- Matijevic, V.; Barbaric, B.; Kraljevic, M.; Milas, I.; Kolak, J. Gender Differences in Neurodevelopmental Outcomes among Full-Term Infants with Intraventricular Hemorrhage. Acta Clin. Croat. 2019, 58, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Reubsaet, P.; Brouwer, A.J.; van Haastert, I.C.; Brouwer, M.J.; Koopman, C.; Groenendaal, F.; de Vries, L.S. The Impact of Low-Grade Germinal Matrix-Intraventricular Hemorrhage on Neurodevelopmental Outcome of Very Preterm Infants. Neonatology 2017, 112, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Szpecht, D.; Frydryszak, D.; Miszczyk, N.; Szymankiewicz, M.; Gadzinowski, J. The incidence of severe intraventricular hemorrhage based on retrospective analysis of 35939 full-term newborns-report of two cases and review of literature. Child’s Nerv. Syst. Off. J. Int. Soc. Pediatr. Neurosurg. 2016, 32, 2447–2451. [Google Scholar] [CrossRef] [Green Version]
- Szpecht, D.; Szymankiewicz, M.; Nowak, I.; Gadzinowski, J. Intraventricular hemorrhage in neonates born before 32 weeks of gestation-retrospective analysis of risk factors. Child’s Nerv. Syst. Off. J. Int. Soc. Pediatr. Neurosurg. 2016, 32, 1399–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampe, R.; Rieger-Fackeldey, E.; Sidorenko, I.; Turova, V.; Botkin, N.; Eckardt, L.; Alves-Pinto, A.; Kovtanyuk, A.; Schundeln, M.; Felderhoff-Muser, U. Assessing key clinical parameters before and after intraventricular hemorrhage in very preterm infants. Eur. J. Pediatr. 2020. [Google Scholar] [CrossRef] [Green Version]
- Helwich, E.; Rutkowska, M.; Bokiniec, R.; Gulczyńska, E.; Hożejowski, R. Intraventricular hemorrhage in premature infants with respiratory distress syndrome treated with surfactant: Incidence and risk factors in the prospective cohort study. Dev. Period Med. 2017, XXI, 328–335. [Google Scholar]
- He, L.; Zhou, W.; Zhao, X.; Liu, X.; Rong, X.; Song, Y. Development and validation of a novel scoring system to predict severe intraventricular hemorrhage in very low birth weight infants. Brain Dev. 2019, 41, 671–677. [Google Scholar] [CrossRef]
- Iyer, K.K.; Roberts, J.A.; Hellstrom-Westas, L.; Wikstrom, S.; Hansen Pupp, I.; Ley, D.; Breakspear, M.; Vanhatalo, S. Early Detection of Preterm Intraventricular Hemorrhage From Clinical Electroencephalography. Crit. Care Med. 2015, 43, 2219–2227. [Google Scholar] [CrossRef]
- Papile, L.A.; Burstein, J.; Burstein, R.; Koffler, H. Incidence and evolution of subependymal and intraventricular hemorrhage: A study of infants with birth weights less than 1500 gm. J. Pediatr. 1978, 92, 529–534. [Google Scholar] [CrossRef]
- Volpe, J.; Inder, T.; Darras, B.; de Vries, L.S.; du Plessis, A.; Neil, J.; Perlman, J. Volpe’s Neurology of the Newborn, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Bowerman, R.A.; Donn, S.M.; Silver, T.M.; Jaffe, M.H. Natural history of neonatal periventricular/intraventricular hemorrhage and its complications: Sonographic observations. AJR Am. J. Roentgenol. 1984, 143, 1041–1052. [Google Scholar] [CrossRef]
- Badhiwala, J.H.; Hong, C.J.; Nassiri, F.; Hong, B.Y.; Riva-Cambrin, J.; Kulkarni, A.V. Treatment of posthemorrhagic ventricular dilation in preterm infants: A systematic review and meta-analysis of outcomes and complications. J. Neurosurg. Pediatr. 2015, 16, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Provenzi, L.; Guida, E.; Montirosso, R. Preterm behavioral epigenetics: A systematic review. Neurosci. Biobehav. Rev. 2018, 84, 262–271. [Google Scholar] [CrossRef]
- Holwerda, J.C.; Van Braeckel, K.; Roze, E.; Hoving, E.W.; Maathuis, C.G.B.; Brouwer, O.F.; Martijn, A.; Bos, A.F. Functional outcome at school age of neonatal post-hemorrhagic ventricular dilatation. Early Hum. Dev. 2016, 96, 15–20. [Google Scholar] [CrossRef]
- Larroque, B.; Ancel, P.Y.; Marret, S.; Marchand, L.; André, M.; Arnaud, C.; Pierrat, V.; Rozé, J.C.; Messer, J.; Thiriez, G.; et al. Neurodevelopmental disabilities and special care of 5-year-old children born before 33 weeks of gestation (the EPIPAGE study): A longitudinal cohort study. Lancet 2008, 371, 813–820. [Google Scholar] [CrossRef]
- Cizmeci, M.N.; de Vries, L.S.; Ly, L.G.; van Haastert, I.C.; Groenendaal, F.; Kelly, E.N.; Traubici, J.; Whyte, H.E.; Leijser, L.M. Periventricular Hemorrhagic Infarction in Very Preterm Infants: Characteristic Sonographic Findings and Association with Neurodevelopmental Outcome at Age 2 Years. J. Pediatr. 2020, 217, 79–85.e71. [Google Scholar] [CrossRef]
- Klebermass-Schrehof, K.; Czaba, C.; Olischar, M.; Fuiko, R.; Waldhoer, T.; Rona, Z.; Pollak, A.; Weninger, M. Impact of low-grade intraventricular hemorrhage on long-term neurodevelopmental outcome in preterm infants. Childs Nerv. Syst. 2012, 28, 2085–2092. [Google Scholar] [CrossRef]
- You, J.; Shamsi, B.H.; Hao, M.C.; Cao, C.H.; Yang, W.Y. A study on the neurodevelopment outcomes of late preterm infants. BMC Neurol. 2019, 19, 108. [Google Scholar] [CrossRef] [Green Version]
- Hollebrandse, N.L.; Spittle, A.J.; Burnett, A.C.; Anderson, P.J.; Roberts, G.; Doyle, L.W.; Cheong, J.L.Y. School-age outcomes following intraventricular haemorrhage in infants born extremely preterm. Arch. Dis. Child Fetal Neonatal Ed. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gram, M.; Sveinsdottir, S.; Ruscher, K.; Hansson, S.R.; Cinthio, M.; Akerström, B.; Ley, D. Hemoglobin induces inflammation after preterm intraventricular hemorrhage by methemoglobin formation. J. Neuroinflamm. 2013, 10, 100. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, S.; Di Martino, E.; Mukai, T.; Tsuji, S.; Murakami, T.; Harris, R.A.; Blomgren, K.; Aden, U. Aggravated brain injury after neonatal hypoxic ischemia in microglia-depleted mice. J. Neuroinflamm. 2020, 17, 111. [Google Scholar] [CrossRef] [Green Version]
- Klebe, D.; McBride, D.; Flores, J.J.; Zhang, J.H.; Tang, J. Modulating the Immune Response Towards a Neuroregenerative Peri-injury Milieu After Cerebral Hemorrhage. J. Neuroimmune Pharmacol. 2015, 10, 576–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supramaniam, V.; Vontell, R.; Srinivasan, L.; Wyatt-Ashmead, J.; Hagberg, H.; Rutherford, M. Microglia activation in the extremely preterm human brain. Pediatr. Res. 2013, 73, 301–309. [Google Scholar] [CrossRef]
- Fernandez-Lopez, D.; Faustino, J.; Klibanov, A.L.; Derugin, N.; Blanchard, E.; Simon, F.; Leib, S.L.; Vexler, Z.S. Microglial Cells Prevent Hemorrhage in Neonatal Focal Arterial Stroke. J. Neurosci. 2016, 36, 2881–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, J.J.; Klebe, D.; Rolland, W.B.; Lekic, T.; Krafft, P.R.; Zhang, J.H. PPARgamma-induced upregulation of CD36 enhances hematoma resolution and attenuates long-term neurological deficits after germinal matrix hemorrhage in neonatal rats. Neurobiol. Dis. 2016, 87, 124–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Li, L.; Jiang, B.; Feng, Z.; Yang, L.; Tang, J.; Chen, Q.; Zhang, J.; Tan, Q.; Feng, H.; et al. Cannabinoid receptor-2 stimulation suppresses neuroinflammation by regulating microglial M1/M2 polarization through the cAMP/PKA pathway in an experimental GMH rat model. Brain Behav. Immun. 2016, 58, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ding, Y.; Lu, T.; Zhang, Y.; Xu, N.; Yu, L.; McBride, D.W.; Flores, J.J.; Tang, J.; Zhang, J.H. Bliverdin reductase-A improves neurological function in a germinal matrix hemorrhage rat model. Neurobiol. Dis. 2018, 110, 122–132. [Google Scholar] [CrossRef]
- Habiyaremye, G.; Morales, D.M.; Morgan, C.D.; McAllister, J.P.; CreveCoeuR, T.S.; Han, R.H.; Gabir, M.; Baksh, B.; Mercer, D.; Limbrick, D.D. Chemokine and cytokine levels in the lumbar cerebrospinal fluid of preterm infants with post-hemorrhagic hydrocephalus. Fluids Barriers CNS 2017, 14. [Google Scholar] [CrossRef] [Green Version]
- Dohare, P.; Zia, M.T.; Ahmed, E.; Ahmed, A.; Yadala, V.; Schober, A.L.; Ortega, J.A.; Kayton, R.; Ungvari, Z.; Mongin, A.A.; et al. AMPA-Kainate Receptor Inhibition Promotes Neurologic Recovery in Premature Rabbits with Intraventricular Hemorrhage. J. Neurosci. 2016, 36, 3363–3377. [Google Scholar] [CrossRef] [Green Version]
- Kallankari, H.; Kaukola, T.; Ojaniemi, M.; Herva, R.; Perhomaa, M.; Vuolteenaho, R.; Kingsmore, S.F.; Hallman, M. Chemokine CCL18 predicts intraventricular hemorrhage in very preterm infants. Ann. Med. 2010, 42, 416–425. [Google Scholar] [CrossRef]
- Nelson, K.B.; Grether, J.K.; Dambrosia, J.M.; Walsh, E.; Kohler, S.; Satyanarayana, G.; Nelson, P.G.; Dickens, B.F.; Phillips, T.M. Neonatal cytokines and cerebral palsy in very preterm infants. Pediatric Res. 2003, 53, 600–607. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Kim, H.J.; Yang, J.H.; Choi, J.s.; Lim, J.E.; Oh, M.J.; Na, J.Y. Neonatal Brain Damage Following Prolonged Latency after Preterm Premature Rupture of Membranes. JKMS 2006, 21, 485–489. [Google Scholar] [CrossRef]
- Kenet, G.; Kuperman, A.A.; Strauss, T.; Brenner, B. Neonatal IVH--mechanisms and management. Thromb. Res. 2011, 127 (Suppl. 3), S120–S122. [Google Scholar] [CrossRef]
- Ballabh, P.; Xu, H.; Hu, F.; Braun, A.; Smith, K.; Rivera, A.; Lou, N.; Ungvari, Z.; Goldman, S.A.; Csiszar, A.; et al. Angiogenic inhibition reduces germinal matrix hemorrhage. Nat. Med. 2007, 13, 477–485. [Google Scholar] [CrossRef]
- Goddard, J.; Lewis, R.M.; Alcala, H.; Zeller, R.S. Intraventricular hemorrhage--an animal model. Biol. Neonate 1980, 37, 39–52. [Google Scholar] [CrossRef]
- Wheeler, A.S.; Sadri, S.; Gutsche, B.B.; DeVore, J.S.; David-Mian, Z.; Latyshevsky, H. Intracranial hemorrhage following intravenous administration of sodium bicarbonate or saline solution in the newborn lamb asphyxiated in utero. Anesthesiology 1979, 51, 517–521. [Google Scholar] [CrossRef]
- Reynolds, M.L.; Evans, C.A.; Reynolds, E.O.; Saunders, N.R.; Durbin, G.M.; Wigglesworth, J.S. Intracranial haemorrhage in the preterm sheep fetus. Early Hum. Dev. 1979, 3, 163–186. [Google Scholar] [CrossRef]
- Lekic, T.; Manaenko, A.; Rolland, W.; Tang, J.P.; Zhang, J.H. A Novel Preclinical Model of Germinal Matrix Hemorrhage Using Neonatal Rats. Acta Neurochir. Suppl. 2011, 111, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Mayfrank, L.; Kissler, J.; Raoofi, R.; Delsing, P.; Weis, J.; Kuker, W.; Gilsbach, J.M. Ventricular dilatation in experimental intraventricular hemorrhage in pigs. Characterization of cerebrospinal fluid dynamics and the effects of fibrinolytic treatment. Stroke 1997, 28, 141–148. [Google Scholar] [CrossRef]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106–107, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lodygensky, G.A.; Vasung, L.; Sizonenko, S.V.; Huppi, P.S. Neuroimaging of cortical development and brain connectivity in human newborns and animal models. J. Anat. 2010, 217, 418–428. [Google Scholar] [CrossRef]
- Bockhorst, K.H.; Narayana, P.A.; Liu, R.; Ahobila-Vijjula, P.; Ramu, J.; Kamel, M.; Wosik, J.; Bockhorst, T.; Hahn, K.; Hasan, K.M.; et al. Early postnatal development of rat brain: In vivo diffusion tensor imaging. J. Neurosci. Res. 2008, 86, 1520–1528. [Google Scholar] [CrossRef] [Green Version]
- Craig, A.; Ling Luo, N.; Beardsley, D.J.; Wingate-Pearse, N.; Walker, D.W.; Hohimer, A.R.; Back, S.A. Quantitative analysis of perinatal rodent oligodendrocyte lineage progression and its correlation with human. Exp. Neurol. 2003, 181, 231–240. [Google Scholar] [CrossRef]
- Catalani, A.; Sabbatini, M.; Consoli, C.; Cinque, C.; Tomassoni, D.; Azmitia, E.; Angelucci, L.; Amenta, F. Glial fibrillary acidic protein immunoreactive astrocytes in developing rat hippocampus. Mech. Ageing Dev. 2002, 123, 481–490. [Google Scholar] [CrossRef]
- Kakita, A.; Goldman, J.E. Patterns and dynamics of SVZ cell migration in the postnatal forebrain: Monitoring living progenitors in slice preparations. Neuron 1999, 23, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Levers, T.E.; Edgar, J.M.; Price, D.J. The fates of cells generated at the end of neurogenesis in developing mouse cortex. J. Neurobiol. 2001, 48, 265–277. [Google Scholar] [CrossRef]
- Christopherson, K.S.; Ullian, E.M.; Stokes, C.C.; Mullowney, C.E.; Hell, J.W.; Agah, A.; Lawler, J.; Mosher, D.F.; Bornstein, P.; Barres, B.A. Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell 2005, 120, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Russo, I.; Gavello, D.; Menna, E.; Vandael, D.; Veglia, C.; Morello, N.; Corradini, I.; Focchi, E.; Alfieri, A.; Angelini, C.; et al. p140Cap Regulates GABAergic Synaptogenesis and Development of Hippocampal Inhibitory Circuits. Cereb. Cortex 2019, 29, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goritz, C.; Mauch, D.H.; Pfrieger, F.W. Multiple mechanisms mediate cholesterol-induced synaptogenesis in a CNS neuron. Mol. Cell. Neurosci. 2005, 29, 190–201. [Google Scholar] [CrossRef]
- Clancy, B.; Kersh, B.; Hyde, J.; Darlington, R.B.; Anand, K.J.S.; Finlay, B.L. Web-based method for translating neurodevelopment from laboratory species to humans. Neuroinformatics 2007, 5, 79–94. [Google Scholar] [CrossRef]
- Tartaglione, A.M.; Armida, M.; Potenza, R.L.; Pezzola, A.; Popoli, P.; Calamandrei, G. Aberrant self-grooming as early marker of motor dysfunction in a rat model of Huntington’s disease. Behav. Brain Res. 2016, 313, 53–57. [Google Scholar] [CrossRef]
- Fox, W.M. Reflex-Ontogeny and Behavioural Development of Mouse. Anim. Behav. 1965, 13, 234–241. [Google Scholar] [CrossRef]
- Hillar, C.; Onnis, G.; Rhea, D.; Tecott, L. Active State Organization of Spontaneous Behavioral Patterns. Sci. Rep. 2018, 8, 1064. [Google Scholar] [CrossRef]
- Aquilina, K.; Chakkarapani, E.; Love, S.; Thoresen, M. Neonatal rat model of intraventricular haemorrhage and post-haemorrhagic ventricular dilatation with long-term survival into adulthood. Neuropathol. Appl. Neurobiol. 2011, 37, 156–165. [Google Scholar] [CrossRef]
- Cherian, S.S.; Love, S.; Silver, I.A.; Porter, H.J.; Whitelaw, A.G.; Thoresen, M. Posthemorrhagic ventricular dilation in the neonate: Development and characterization of a rat model. J. Neuropathol. Exp. Neurol. 2003, 62, 292–303. [Google Scholar] [CrossRef] [Green Version]
- Hagberg, H.; Peebles, D.; Mallard, C. Models of white matter injury: Comparison of infectious, hypoxic-ischemic, and excitotoxic insults. Ment. Retard. Dev. Disabil. Res. Rev. 2002, 8, 30–38. [Google Scholar] [CrossRef]
- McCarty, J.H.; Monahan-Earley, R.A.; Brown, L.F.; Keller, M.; Gerhardt, H.; Rubin, K.; Shani, M.; Dvorak, H.F.; Wolburg, H.; Bader, B.L.; et al. Defective associations between blood vessels and brain parenchyma lead to cerebral hemorrhage in mice lacking alphav integrins. Mol. Cell. Biol. 2002, 22, 7667–7677. [Google Scholar] [CrossRef] [Green Version]
- Gould, D.B.; Phalan, F.C.; Breedveld, G.J.; van Mil, S.E.; Smith, R.S.; Schimenti, J.C.; Aguglia, U.; van der Knaap, M.S.; Heutink, P.; John, S.W. Mutations in Col4a1 cause perinatal cerebral hemorrhage and porencephaly. Science 2005, 308, 1167–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Baumann, J.M.; Sun, Y.Y.; Tang, M.; Dunn, R.S.; Akeson, A.L.; Kernie, S.G.; Kallapur, S.; Lindquist, D.M.; Huang, E.J.; et al. Overexpression of vascular endothelial growth factor in the germinal matrix induces neurovascular proteases and intraventricular hemorrhage. Sci. Transl. Med. 2013, 5, 193ra190. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.T.; Li, W.Y.; Kaartinen, V. Tissue-specific expression of Cre recombinase from the Tgfb3 locus. Genesis 2008, 46, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, S.L.; Boylen, K.P.; Einheber, S.; Milner, T.A.; Ramos, D.M.; Pytela, R. Synaptic and glial localization of the integrin alphavbeta8 in mouse and rat brain. Brain Res. 1998, 791, 271–282. [Google Scholar] [CrossRef]
- Yamamoto, H.; Ehling, M.; Kato, K.; Kanai, K.; van Lessen, M.; Frye, M.; Zeuschner, D.; Nakayama, M.; Vestweber, D.; Adams, R.H. Integrin β1 controls VE-cadherin localization and blood vessel stability. Nat. Commun. 2015, 6, 6429. [Google Scholar] [CrossRef] [Green Version]
- Milner, R.; Relvas, J.B.; Fawcett, J.; ffrench-Constant, C. Developmental regulation of alphav integrins produces functional changes in astrocyte behavior. Mol. Cell. Neurosci. 2001, 18, 108–118. [Google Scholar] [CrossRef]
- Bader, B.L.; Rayburn, H.; Crowley, D.; Hynes, R.O. Extensive vasculogenesis, angiogenesis, and organogenesis precede lethality in mice lacking all alpha v integrins. Cell 1998, 95, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Liu, D.; Huang, Y.; Jovin, I.; Shai, S.Y.; Kyriakides, T.; Ross, R.S.; Giordano, F.J. Endothelial expression of beta1 integrin is required for embryonic vascular patterning and postnatal vascular remodeling. Mol. Cell Biol. 2008, 28, 794–802. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Motejlek, K.; Wang, D.; Zang, K.; Schmidt, A.; Reichardt, L.F. beta8 integrins are required for vascular morphogenesis in mouse embryos. Development 2002, 129, 2891–2903. [Google Scholar]
- Ma, S.; Santhosh, D.; Kumar, P.; Huang, Z. A Brain-Region-Specific Neural Pathway Regulating Germinal Matrix Angiogenesis. Dev. Cell 2017, 41, 366–381.e364. [Google Scholar] [CrossRef] [Green Version]
- Bilguvar, K.; DiLuna, M.L.; Bizzarro, M.J.; Bayri, Y.; Schneider, K.C.; Lifton, R.P.; Gunel, M.; Ment, L.R.; Pacifier; Breastfeeding Trial, G. COL4A1 mutation in preterm intraventricular hemorrhage. J. Pediatr. 2009, 155, 743–745. [Google Scholar] [CrossRef] [Green Version]
- Krum, J.M.; Mani, N.; Rosenstein, J.M. Angiogenic and astroglial responses to vascular endothelial growth factor administration in adult rat brain. Neuroscience 2002, 110, 589–604. [Google Scholar] [CrossRef]
- El Shimi, M.S.; Hassanein, S.M.; Mohamed, M.H.; Abdou, R.M.; Roshdy, A.; Atef, S.H.; Aly, H. Predictive value of vascular endothelial growth factor in preterm neonates with intraventricular haemorrhage. J. Matern. Fetal Neonatal Med. 2012, 25, 1586–1590. [Google Scholar] [CrossRef]
- Takashima, S.; Tanaka, K. Microangiography and vascular permeability of the subependymal matrix in the premature infant. Can. J. Neurol. Sci. 1978, 5, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Fischer, E.G.; Lorenzo, A.V.; Landis, W.J.; Welch, K.; Ofori-Kwakye, S.K.; Dorval, B.; Hodgens, K.J.; Kerr, C.S. Vasculature to the germinal matrix in rabbit pups. J. Neurosurg. 1986, 64, 650–656. [Google Scholar] [CrossRef]
- Georgiadis, P.; Xu, H.; Chua, C.; Hu, F.; Collins, L.; Huynh, C.; Lagamma, E.F.; Ballabh, P. Characterization of acute brain injuries and neurobehavioral profiles in a rabbit model of germinal matrix hemorrhage. Stroke 2008, 39, 3378–3388. [Google Scholar] [CrossRef] [Green Version]
- Dohare, P.; Cheng, B.; Ahmed, E.; Yadala, V.; Singla, P.; Thomas, S.; Kayton, R.; Ungvari, Z.; Ballabh, P. Glycogen synthase kinase-3beta inhibition enhances myelination in preterm newborns with intraventricular hemorrhage, but not recombinant Wnt3A. Neurobiol. Dis. 2018, 118, 22–39. [Google Scholar] [CrossRef]
- Lee, J.H.; Chang, Y.S.; Ahn, S.Y.; Sung, S.I.; Park, W.S. Dexamethasone does not prevent hydrocephalus after severe intraventricular hemorrhage in newborn rats. PLoS ONE 2018, 13, e0206306. [Google Scholar] [CrossRef] [PubMed]
- Strahle, J.M.; Garton, T.; Bazzi, A.A.; Kilaru, H.; Garton, H.J.; Maher, C.O.; Muraszko, K.M.; Keep, R.F.; Xi, G. Role of hemoglobin and iron in hydrocephalus after neonatal intraventricular hemorrhage. Neurosurgery 2014, 75, 696–705; discussion 706. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.Y.; Chang, Y.S.; Sung, D.K.; Sung, S.I.; Yoo, H.S.; Lee, J.H.; Oh, W.I.; Park, W.S. Mesenchymal stem cells prevent hydrocephalus after severe intraventricular hemorrhage. Stroke 2013, 44, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Zhao, G.; Ding, Y.; Wang, T.; Flores, J.; Ocak, U.; Wu, P.; Zhang, T.; Mo, J.; Zhang, J.H.; et al. Rh-IFN-alpha attenuates neuroinflammation and improves neurological function by inhibiting NF-kappaB through JAK1-STAT1/TRAF3 pathway in an experimental GMH rat model. Brain Behav. Immun. 2019, 79, 174–185. [Google Scholar] [CrossRef]
- Leroux, P.; Omouendze, P.L.; Roy, V.; Dourmap, N.; Gonzalez, B.J.; Brasse-Lagnel, C.; Carmeliet, P.; Leroux-Nicollet, I.; Marret, S. Age-dependent neonatal intracerebral hemorrhage in plasminogen activator inhibitor 1 knockout mice. J. Neuropathol. Exp. Neurol. 2014, 73, 387–402. [Google Scholar] [CrossRef] [Green Version]
- Sotrel, A.; Lorenzo, A.V. Ultrastructure of blood vessels in the ganglionic eminence of premature rabbits with spontaneous germinal matrix hemorrhages. J. Neuropathol. Exp. Neurol. 1989, 48, 462–482. [Google Scholar] [CrossRef]
- Lorenzo, A.V.; Welch, K.; Conner, S. Spontaneous germinal matrix and intraventricular hemorrhage in prematurely born rabbits. J. Neurosurg. 1982, 56, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, A.V.; Welch, K. Preterm rabbit model of intraventricular hemorrhage. J. Neurosurg. 1986, 64, 688–689. [Google Scholar] [CrossRef]
- Chua, C.O.; Chahboune, H.; Braun, A.; Dummula, K.; Chua, C.E.; Yu, J.; Ungvari, Z.; Sherbany, A.A.; Hyder, F.; Ballabh, P. Consequences of intraventricular hemorrhage in a rabbit pup model. Stroke 2009, 40, 3369–3377. [Google Scholar] [CrossRef] [Green Version]
- Vinukonda, G.; Dohare, P.; Arshad, A.; Zia, M.T.; Panda, S.; Korumilli, R.; Kayton, R.; Hascall, V.C.; Lauer, M.E.; Ballabh, P. Hyaluronidase and Hyaluronan Oligosaccharides Promote Neurological Recovery after Intraventricular Hemorrhage. J. Neurosci. 2016, 36, 872–889. [Google Scholar] [CrossRef]
- Vinukonda, G.; Csiszar, A.; Hu, F.; Dummula, K.; Pandey, N.K.; Zia, M.T.; Ferreri, N.R.; Ungvari, Z.; LaGamma, E.F.; Ballabh, P. Neuroprotection in a rabbit model of intraventricular haemorrhage by cyclooxygenase-2, prostanoid receptor-1 or tumour necrosis factor-alpha inhibition. Brain J. Neurol. 2010, 133, 2264–2280. [Google Scholar] [CrossRef] [Green Version]
- Gram, M.; Sveinsdottir, S.; Cinthio, M.; Sveinsdottir, K.; Hansson, S.R.; Morgelin, M.; Akerstrom, B.; Ley, D. Extracellular hemoglobin - mediator of inflammation and cell death in the choroid plexus following preterm intraventricular hemorrhage. J. Neuroinflamm. 2014, 11, 200. [Google Scholar] [CrossRef] [Green Version]
- Park, W.S.; Sung, S.I.; Ahn, S.Y.; Sung, D.K.; Im, G.H.; Yoo, H.S.; Choi, S.J.; Chang, Y.S. Optimal Timing of Mesenchymal Stem Cell Therapy for Neonatal Intraventricular Hemorrhage. Cell Transpl. 2016, 25, 1131–1144. [Google Scholar] [CrossRef] [Green Version]
- Vinukonda, G.; Liao, Y.; Hu, F.; Ivanova, L.; Purohit, D.; Finkel, D.A.; Giri, P.; Bapatla, L.; Shah, S.; Zia, M.T.; et al. Human Cord Blood-Derived Unrestricted Somatic Stem Cell Infusion Improves Neurobehavioral Outcome in a Rabbit Model of Intraventricular Hemorrhage. Stem Cells Transl. Med. 2019, 8, 1157–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulding, D.S.; Vogel, R.C.; Gensel, J.C.; Morganti, J.M.; Stromberg, A.J.; Miller, B.A. Acute brain inflammation, white matter oxidative stress, and myelin deficiency in a model of neonatal intraventricular hemorrhage. J. Neurosurg. Pediatr. 2020, 1–11. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Chang, Y.S.; Sung, D.K.; Sung, S.I.; Yoo, H.S.; Im, G.H.; Choi, S.J.; Park, W.S. Optimal Route for Mesenchymal Stem Cells Transplantation after Severe Intraventricular Hemorrhage in Newborn Rats. PLoS ONE 2015, 10, e0132919. [Google Scholar] [CrossRef]
- Mukai, T.; Mori, Y.; Shimazu, T.; Takahashi, A.; Tsunoda, H.; Yamaguchi, S.; Kiryu, S.; Tojo, A.; Nagamura-Inoue, T. Intravenous injection of umbilical cord-derived mesenchymal stromal cells attenuates reactive gliosis and hypomyelination in a neonatal intraventricular hemorrhage model. Neuroscience 2017, 355, 175–187. [Google Scholar] [CrossRef]
- Dawes, W.J.; Zhang, X.; Fancy, S.P.J.; Rowitch, D.; Marino, S. Moderate-Grade Germinal Matrix Haemorrhage Activates Cell Division in the Neonatal Mouse Subventricular Zone. Dev. Neurosci. 2016, 38, 430–444. [Google Scholar] [CrossRef] [PubMed]
- Aquilina, K.; Hobbs, C.; Tucker, A.; Whitelaw, A.; Thoresen, M. Do drugs that block transforming growth factor beta reduce posthaemorrhagic ventricular dilatation in a neonatal rat model? Acta Paediatr. 2008, 97, 1181–1186. [Google Scholar] [CrossRef]
- Ko, H.R.; Ahn, S.Y.; Chang, Y.S.; Hwang, I.; Yun, T.; Sung, D.K.; Sung, S.I.; Park, W.S.; Ahn, J.Y. Human UCB-MSCs treatment upon intraventricular hemorrhage contributes to attenuate hippocampal neuron loss and circuit damage through BDNF-CREB signaling. Stem Cell Res. Ther. 2018, 9, 326. [Google Scholar] [CrossRef]
- Garton, T.P.; He, Y.; Garton, H.J.; Keep, R.F.; Xi, G.; Strahle, J.M. Hemoglobin-induced neuronal degeneration in the hippocampus after neonatal intraventricular hemorrhage. Brain Res. 2016, 1635, 86–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kim, Y.E.; Hong, S.; Kim, K.T.; Sung, D.K.; Lee, Y.; Park, W.S.; Chang, Y.S.; Song, M.R. Reactive microglia and astrocytes in neonatal intraventricular hemorrhage model are blocked by mesenchymal stem cells. Glia 2020, 68, 178–192. [Google Scholar] [CrossRef]
- Klebe, D.; Krafft, P.R.; Hoffmann, C.; Lekic, T.; Flores, J.J.; Rolland, W.; Zhang, J.H. Acute and delayed deferoxamine treatment attenuates long-term sequelae after germinal matrix hemorrhage in neonatal rats. Stroke 2014, 45, 2475–2479. [Google Scholar] [CrossRef]
- Alles, Y.C.; Greggio, S.; Alles, R.M.; Azevedo, P.N.; Xavier, L.L.; DaCosta, J.C. A novel preclinical rodent model of collagenase-induced germinal matrix/intraventricular hemorrhage. Brain Res. 2010, 1356, 130–138. [Google Scholar] [CrossRef]
- Guo, J.; Chen, Q.; Tang, J.; Zhang, J.; Tao, Y.; Li, L.; Zhu, G.; Feng, H.; Chen, Z. Minocycline-induced attenuation of iron overload and brain injury after experimental germinal matrix hemorrhage. Brain Res. 2015, 1594, 115–124. [Google Scholar] [CrossRef]
- Lekic, T.; Manaenko, A.; Rolland, W.; Krafft, P.R.; Peters, R.; Hartman, R.E.; Altay, O.; Tang, J.P.; Zhang, J.H. Rodent neonatal germinal matrix hemorrhage mimics the human brain injury, neurological consequences, and post-hemorrhagic hydrocephalus. Exp. Neurol. 2012, 236, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Zhang, T.; Wu, G.; McBride, D.W.; Xu, N.; Klebe, D.W.; Zhang, Y.; Li, Q.; Tang, J.; Zhang, J.H. Astrogliosis inhibition attenuates hydrocephalus by increasing cerebrospinal fluid reabsorption through the glymphatic system after germinal matrix hemorrhage. Exp. Neurol. 2019, 320, 113003. [Google Scholar] [CrossRef]
- Tang, J.; Chen, Q.; Guo, J.; Yang, L.; Tao, Y.; Li, L.; Miao, H.; Feng, H.; Chen, Z.; Zhu, G. Minocycline Attenuates Neonatal Germinal-Matrix-Hemorrhage-Induced Neuroinflammation and Brain Edema by Activating Cannabinoid Receptor 2. Mol. Neurobiol. 2016, 53, 1935–1948. [Google Scholar] [CrossRef]
- Li, P.; Zhao, G.; Chen, F.; Ding, Y.; Wang, T.; Liu, S.; Lu, W.; Xu, W.; Flores, J.; Ocak, U.; et al. Rh-relaxin-2 attenuates degranulation of mast cells by inhibiting NF-κB through PI3K-AKT/TNFAIP3 pathway in an experimental germinal matrix hemorrhage rat model. J. Neuroinflamm. 2020, 17, 250. [Google Scholar] [CrossRef]
- Liu, S.P.; Huang, L.; Flores, J.; Ding, Y.; Li, P.; Peng, J.; Zuo, G.; Zhang, J.H.; Lu, J.; Tang, J.P. Secukinumab attenuates reactive astrogliosis via IL-17RA/(C/EBPβ)/SIRT1 pathway in a rat model of germinal matrix hemorrhage. CNS Neurosci. Ther. 2019, 25, 1151–1161. [Google Scholar] [CrossRef] [Green Version]
- Klebe, D.; Flores, J.J.; McBride, D.W.; Krafft, P.R.; Rolland, W.B.; Lekic, T.; Zhang, J.H. Dabigatran ameliorates post-haemorrhagic hydrocephalus development after germinal matrix haemorrhage in neonatal rat pups. J. Cereb. Blood Flow Metab. 2017, 37, 3135–3149. [Google Scholar] [CrossRef]
- Rolland, W.B.; Krafft, P.R.; Lekic, T.; Klebe, D.; LeGrand, J.; Weldon, A.J.; Xu, L.; Zhang, J.H. Fingolimod confers neuroprotection through activation of Rac1 after experimental germinal matrix hemorrhage in rat pups. J. Neurochem. 2017, 140, 776–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, M.; Balasubramaniam, J.; Buist, R.J.; Peeling, J.; Del Bigio, M.R. Periventricular/intraventricular hemorrhage in neonatal mouse cerebrum. J. Neuropathol. Exp. Neurol. 2003, 62, 1154–1165. [Google Scholar] [CrossRef] [Green Version]
- Hierro-Bujalance, C.; Carmen, I.-G.; Sanchez-Sotano, D.; del Marco, A.; Casado-Revuelta, A.; Mengual-Gonzalez, C.M.; Lucena-Porras, C.; Bernal-Martin, M.; Benavente-Fernandez, I.; Lubian-Lopez, S.; et al. Erytrhopoietin Improves Atrophy, Bleeding and Cognition in the Newborn Intraventricular Hemorrhage. Front. Cell. Dev. Biol. 2020, 8, 911. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Lee, N.K.; Na, D.L.; Chang, J.W. Optimal mesenchymal stem cell delivery routes to enhance neurogenesis for the treatment of Alzheimer’s disease: Optimal MSCs delivery routes for the treatment of AD. Histol. Histopathol. 2018, 33, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Abrantes De Lacerda Almeida, T.; Santos, M.V.; Da Silva Lopes, L.; Goel, G.; Leonardo De Freitas, R.; De Medeiros, P.; Crippa, J.A.; Machado, H.R. Intraperitoneal cannabidiol attenuates neonatal germinal matrix hemorrhage-induced neuroinflamation and perilesional apoptosis. Neurol. Res. 2019, 41, 980–990. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, N.; Ding, Y.; Zhang, Y.; Li, Q.; Flores, J.; Haghighiabyaneh, M.; Doycheva, D.; Tang, J.; Zhang, J.H. Chemerin suppresses neuroinflammation and improves neurological recovery via CaMKK2/AMPK/Nrf2 pathway after germinal matrix hemorrhage in neonatal rats. Brain Behav. Immun. 2018, 70, 179–193. [Google Scholar] [CrossRef]
- Xu, M.M.; Seyler, L.; Bauerle, T.; Kalinichenko, L.S.; Muller, C.P.; Huttner, H.B.; Schwab, S.; Manaenko, A. Serelaxin activates eNOS, suppresses inflammation, attenuates developmental delay and improves cognitive functions of neonatal rats after germinal matrix hemorrhage. Sci. Rep. 2020, 10, 8115. [Google Scholar] [CrossRef]
- Feng, Z.; Ye, L.; Klebe, D.; Ding, Y.; Guo, Z.N.; Flores, J.J.; Yin, C.; Tang, J.; Zhang, J.H. Anti-inflammation conferred by stimulation of CD200R1 via Dok1 pathway in rat microglia after germinal matrix hemorrhage. J. Cereb. Blood Flow Metab. 2019, 39, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, J.; Li, P.; Ding, Y.; Tang, J.; Chen, G.; Zhang, J.H. NT-4 attenuates neuroinflammation via TrkB/PI3K/FoxO1 pathway after germinal matrix hemorrhage in neonatal rats. J. Neuroinflamm. 2020, 17, 158. [Google Scholar] [CrossRef]
- Li, Q.; Ding, Y.; Krafft, P.; Wan, W.; Yan, F.; Wu, G.; Zhang, Y.; Zhan, Q.; Zhang, J.H. Targeting Germinal Matrix Hemorrhage-Induced Overexpression of Sodium-Coupled Bicarbonate Exchanger Reduces Posthemorrhagic Hydrocephalus Formation in Neonatal Rats. J. Am. Heart Assoc. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Traudt, C.M.; McPherson, R.J.; Studholme, C.; Millen, K.J.; Juul, S.E. Systemic glycerol decreases neonatal rabbit brain and cerebellar growth independent of intraventricular hemorrhage. Pediatr. Res. 2014, 75, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Ley, D.; Romantsik, O.; Vallius, S.; Sveinsdottir, K.; Sveinsdottir, S.; Agyemang, A.A.; Baumgarten, M.; Morgelin, M.; Lutay, N.; Bruschettini, M.; et al. High Presence of Extracellular Hemoglobin in the Periventricular White Matter Following Preterm Intraventricular Hemorrhage. Front. Physiol. 2016, 7, 330. [Google Scholar] [CrossRef] [Green Version]
- Lekic, T.; Klebe, D.; Poblete, R.; Krafft, P.R.; Rolland, W.B.; Tang, J.; Zhang, J.H. Neonatal brain hemorrhage (NBH) of prematurity: Translational mechanisms of the vascular-neural network. Curr. Med. Chem. 2015, 22, 1214–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramaniam, J.; Xue, M.; Buist, R.J.; Ivanco, T.L.; Natuik, S.; Del Bigio, M.R. Persistent motor deficit following infusion of autologous blood into the periventricular region of neonatal rats. Exp. Neurol. 2006, 197, 122–132. [Google Scholar] [CrossRef]
- Xue, M.; Del Bigio, M.R. Comparison of brain cell death and inflammatory reaction in three models of intracerebral hemorrhage in adult rats. J. Stroke Cerebrovasc. Dis. 2003, 12, 152–159. [Google Scholar] [CrossRef]
- Sansing, L.H.; Kasner, S.E.; McCullough, L.; Agarwal, P.; Welsh, F.A.; Kariko, K. Autologous blood injection to model spontaneous intracerebral hemorrhage in mice. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [Green Version]
- Krafft, P.R.; Rolland, W.B.; Duris, K.; Lekic, T.; Campbell, A.; Tang, J.; Zhang, J.H. Modeling intracerebral hemorrhage in mice: Injection of autologous blood or bacterial collagenase. J. Vis. Exp. 2012, e4289. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Du, H.; Hua, Y.; Keep, R.F.; Strahle, J.; Xi, G. Role of red blood cell lysis and iron in hydrocephalus after intraventricular hemorrhage. J. Cereb. Blood Flow Metab. 2014, 34, 1070–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Bigio, M.R.; Yan, H.J.; Buist, R.; Peeling, J. Experimental intracerebral hemorrhage in rats. Magnetic resonance imaging and histopathological correlates. Stroke 1996, 27, 2312–2319; discussion 2319–2320. [Google Scholar] [CrossRef]
- Malaguit, J.; Casel, D.; Dixon, B.; Doycheva, D.; Tang, J.; Zhang, J.H.; Lekic, T. Intranasal Osteopontin for Rodent Germinal Matrix Hemorrhage. Acta Neurochir. Suppl. 2016, 121, 217–220. [Google Scholar] [CrossRef]
- Lekic, T.; Klebe, D.; McBride, D.W.; Manaenko, A.; Rolland, W.B.; Flores, J.J.; Altay, O.; Tang, J.; Zhang, J.H. Protease-activated receptor 1 and 4 signal inhibition reduces preterm neonatal hemorrhagic brain injury. Stroke 2015, 46, 1710–1713. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Tang, J.; Chen, Q.; Guo, J.; Li, L.; Yang, L.; Feng, H.; Zhu, G.; Chen, Z. Cannabinoid CB2 receptor stimulation attenuates brain edema and neurological deficits in a germinal matrix hemorrhage rat model. Brain Res. 2015, 1602, 127–135. [Google Scholar] [CrossRef]


{kind=link}
| GM-IVH Lesion | Brain Atrophy and Myelinization | Inflammation | Motor Activity and Cognitive Impairments |
|---|---|---|---|
| Glycerol | ↓ White matter, myelinization and myelin basic protein [117,118]. Ventriculomegaly [117]. Cortical thinning [117]. ↓ Cerebellar volume [118]. Neurodegeneration [119]. | ↑ Microglia burden (107). ↑ Pro-inflammatory cytokines [30,54,63,108,120,121]. | Hypertonia [117]. ↓ Walking speed [117]. Weakness [117]. Abnormal gait [117]. Impaired locomotion on 30° inclination [122]. |
| Blood and blood derivates | ↓ White matter, myelinization and myelin basic protein [89,109,111,123,124,125]. Ventricle dilatation [7,88,89,123,124,125,126,127]. Cell death and neuronal loss [109,111,126,128,129]. | ↑ Reactive gliosis, microglia and astrocyte burdens [89,109,111,123,125,130]. ↑ Proinflammatory cytokines [109]. ↑ Neutrophil infiltration [107]. | Altered negative geotaxis test [88,109,111,127]. Altered rotarod test [121,124] Altered grip traction test [89]. Altered open field test [125]. Memory alterations in the passive avoidance test and the Y-maze [128]. |
| Collagenase | White matter lesions [131]. ↓ Brain volume [32,132,133,134,135]. Ventricle enlargement [32,133,134,135]. Cortical thinning [32,131,136,137,138]. Neuronal death [32]. Hydrocephalus [135,136,139,140]. | ↑ Reactive gliosis, microglia and astrocyte burdens [32,112,141,142,143,144]. ↑ Proinflammatory cytokines [145]. | Delayed eye opening latency [146]. Altered righting reflex, negative geotaxis or rotarod tests [112,132,135,147,148]. Altered composite neuroscore and foot fault tests [72,140]. Altered open field test [72,146]. Altered Morris water maze test [32,72,135,140,149]. Altered new object discrimination test [32]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atienza-Navarro, I.; Alves-Martinez, P.; Lubian-Lopez, S.; Garcia-Alloza, M. Germinal Matrix-Intraventricular Hemorrhage of the Preterm Newborn and Preclinical Models: Inflammatory Considerations. Int. J. Mol. Sci. 2020, 21, 8343. https://doi.org/10.3390/ijms21218343
Atienza-Navarro I, Alves-Martinez P, Lubian-Lopez S, Garcia-Alloza M. Germinal Matrix-Intraventricular Hemorrhage of the Preterm Newborn and Preclinical Models: Inflammatory Considerations. International Journal of Molecular Sciences. 2020; 21(21):8343. https://doi.org/10.3390/ijms21218343
Chicago/Turabian StyleAtienza-Navarro, Isabel, Pilar Alves-Martinez, Simon Lubian-Lopez, and Monica Garcia-Alloza. 2020. "Germinal Matrix-Intraventricular Hemorrhage of the Preterm Newborn and Preclinical Models: Inflammatory Considerations" International Journal of Molecular Sciences 21, no. 21: 8343. https://doi.org/10.3390/ijms21218343