JMJD6 Regulates Splicing of Its Own Gene Resulting in Alternatively Spliced Isoforms with Different Nuclear Targets

 , , , and
, , , and 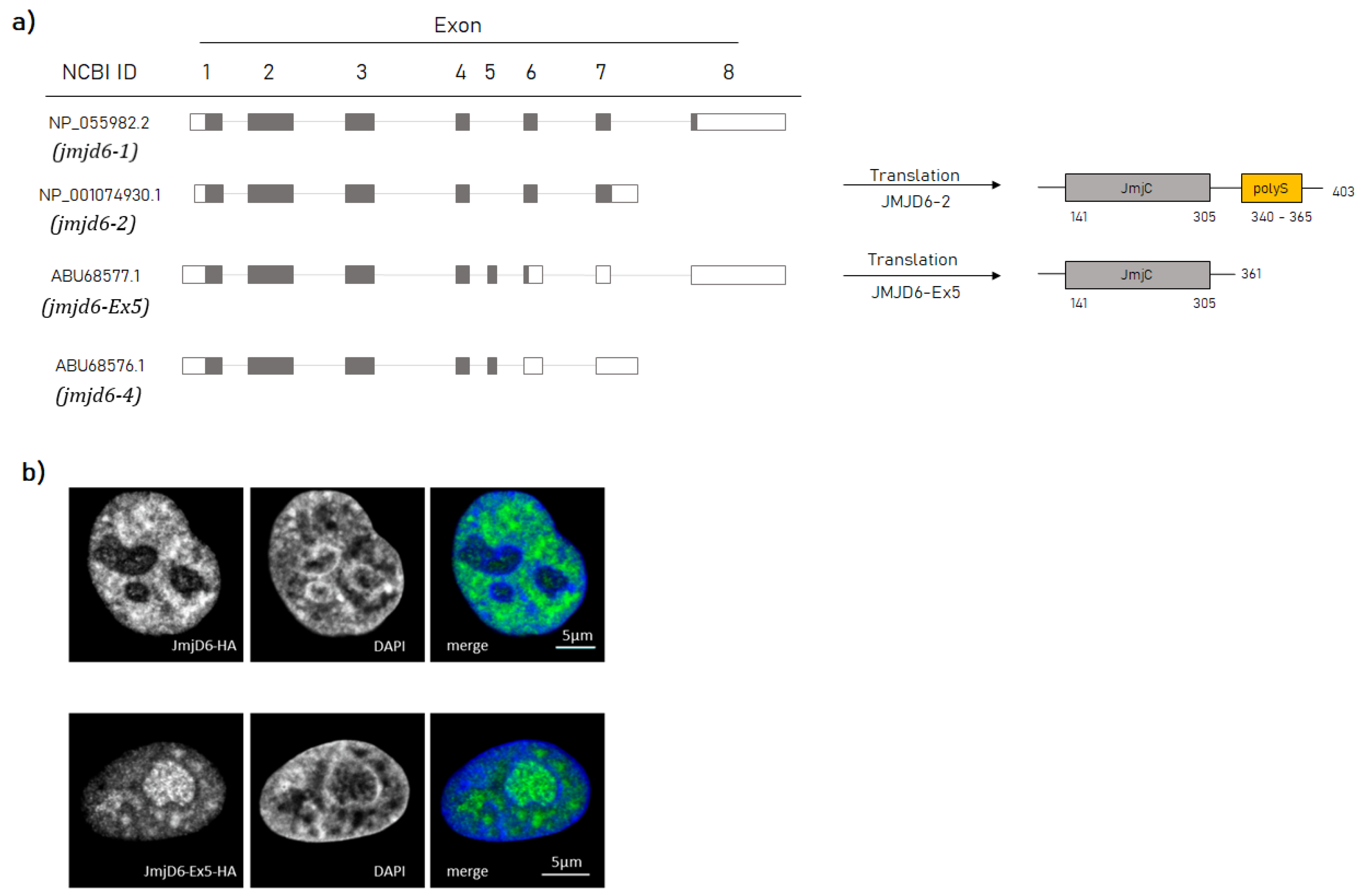{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
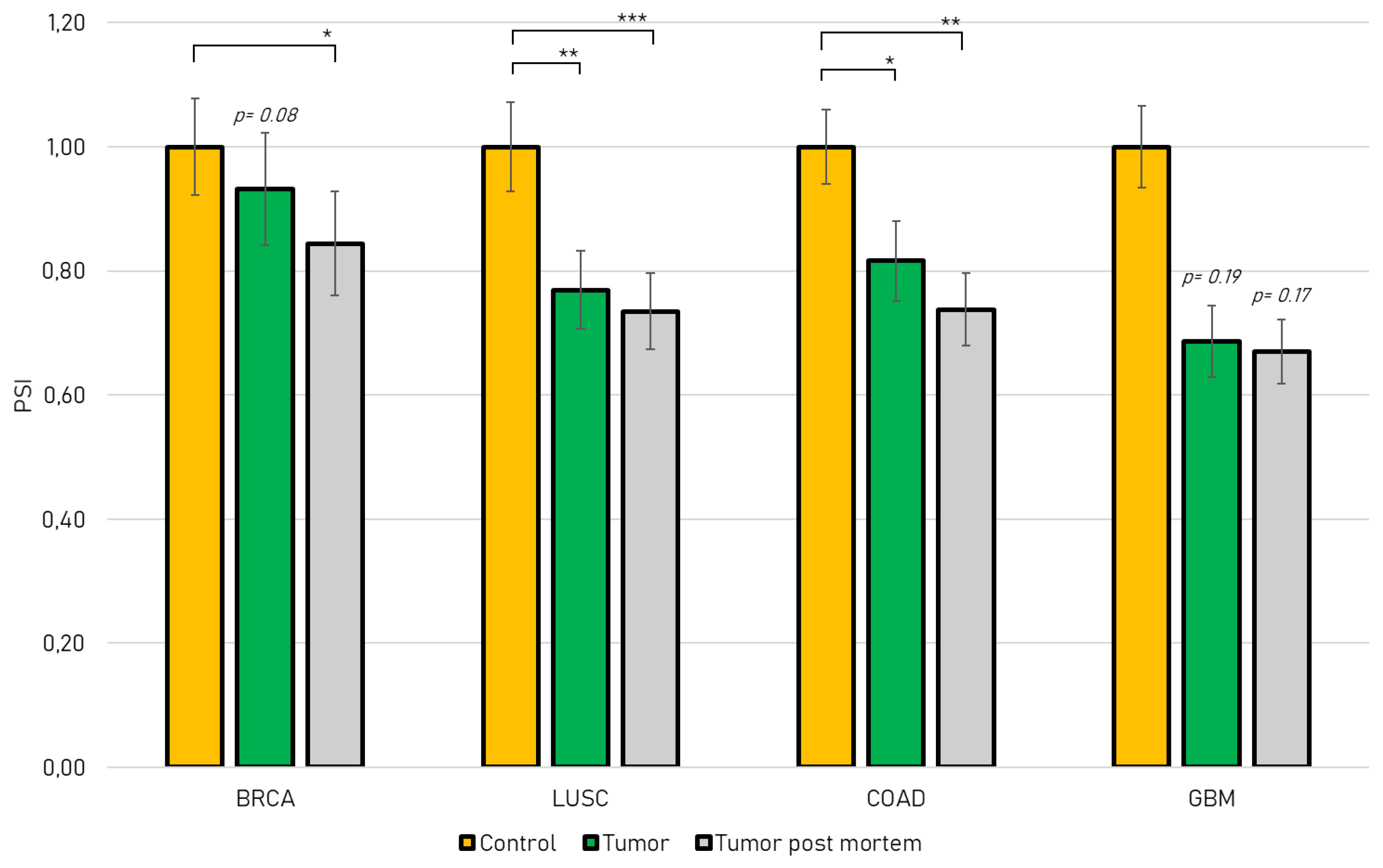
2.1. Jmjd6-Ex5 Is Significantly Decreased in JMJD6 Relevant Cancer Types
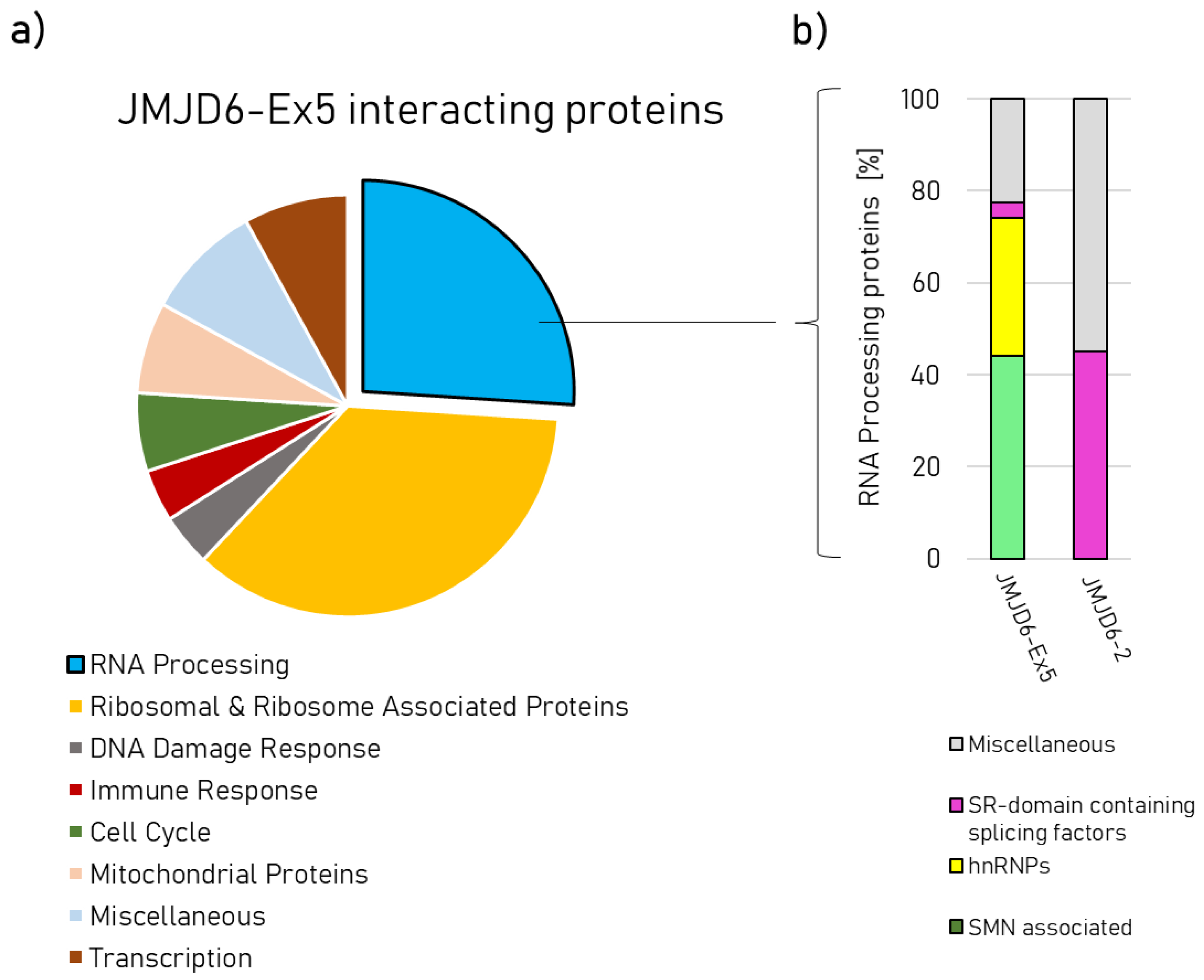
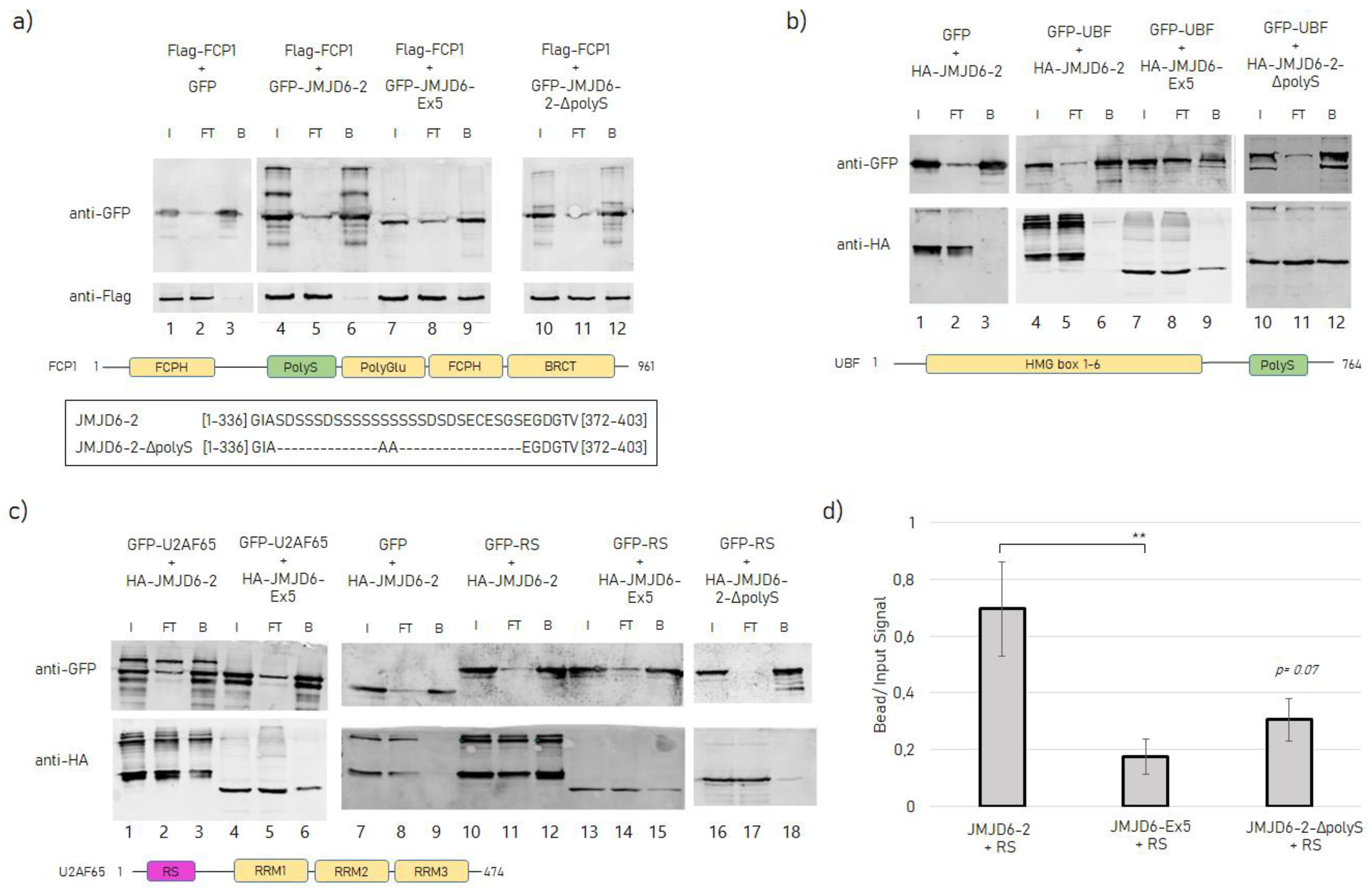
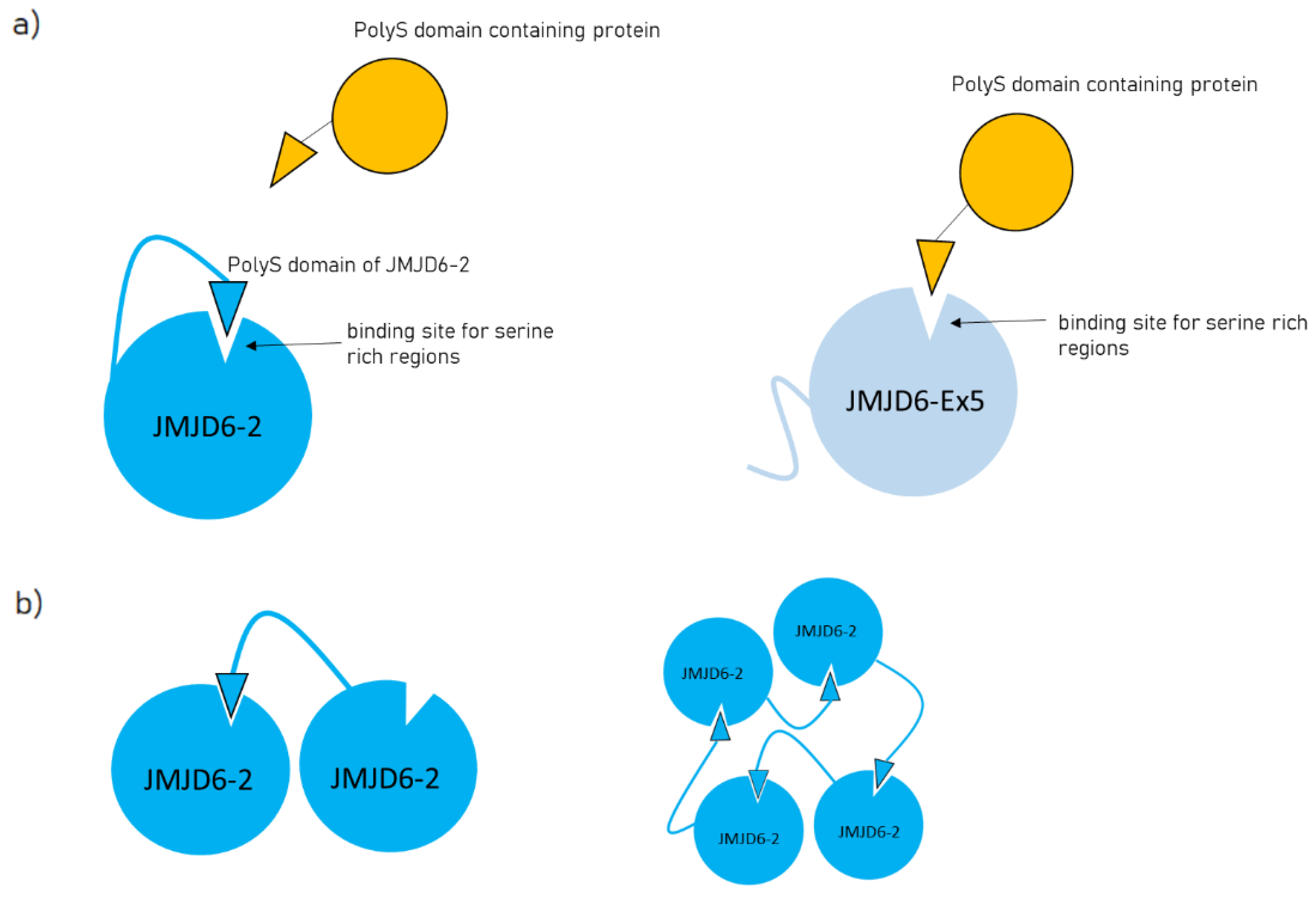
2.2. Protein–Protein Interactions of JMJD6-2 and JMJD6-Ex-5 and the Role of the polyS-domain
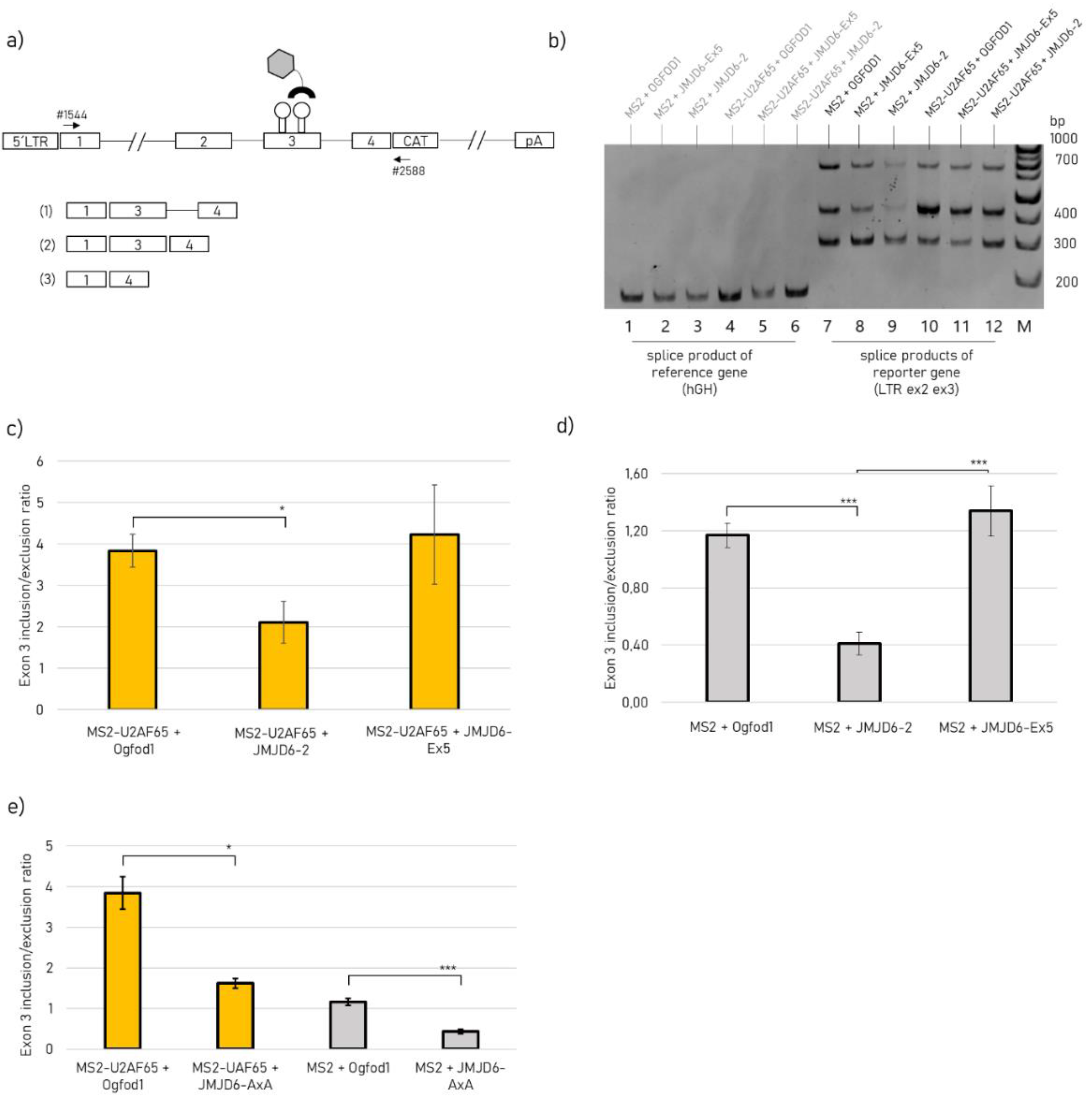
2.3. JMJD6-Ex5 Is not Involved in Splice Site Selection of a Reporter Gene
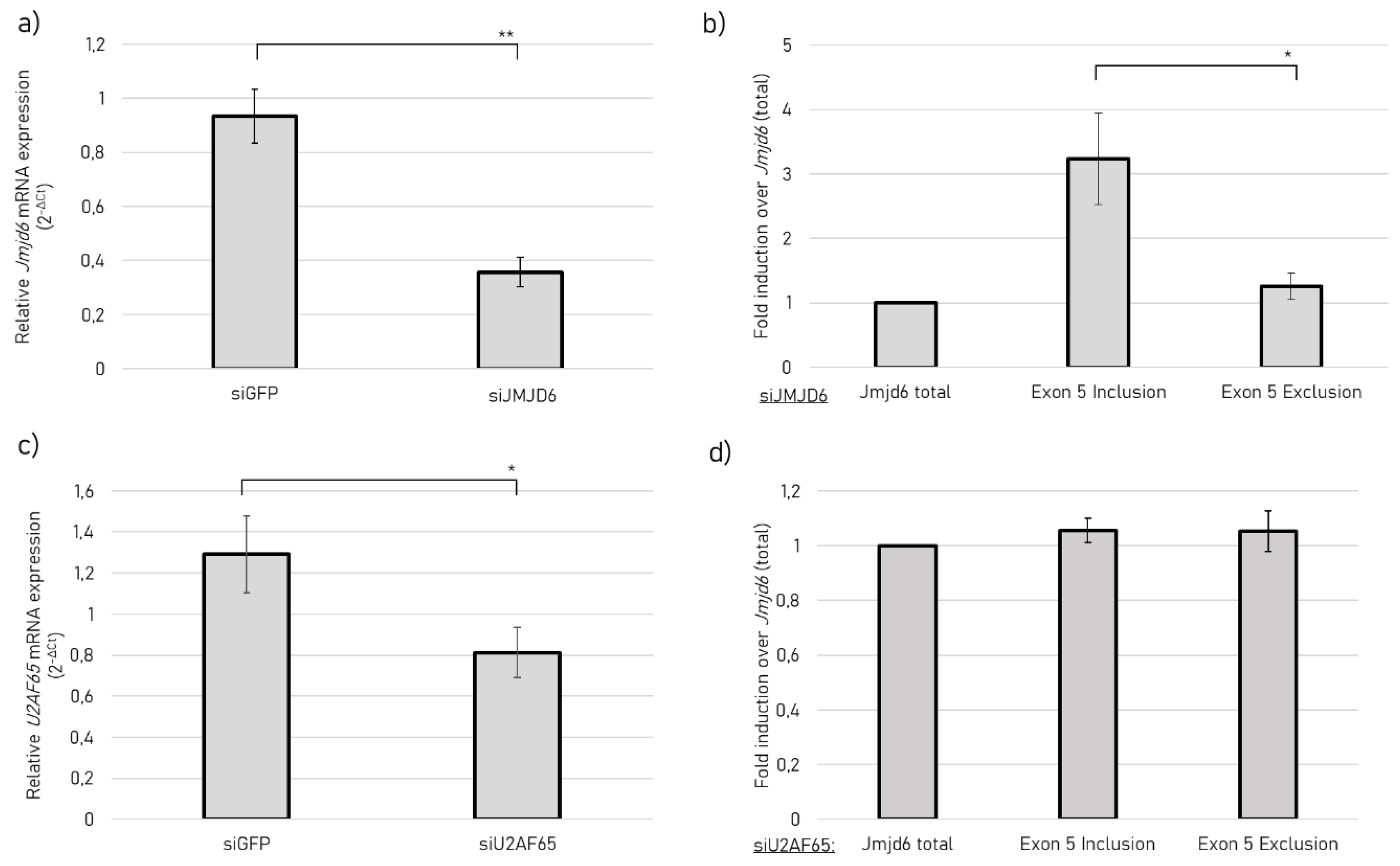
2.4. JMJD6 Regulates Splicing of Its Own Gene to Promote Exon 5 Exclusion
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
4.2. Co-Immunoprecipitation Experiments
4.3. Interactome Analysis
4.4. Proteomic Sample Preparation
4.5. LC-MS/MS Measurement and Analysis
4.6. HIV-1-Based Splicing Reporter Assay
4.7. RNA Interference
4.8. qRT-PCR from esiRNA Experiments
4.9. TCGA SpliceSeq Database
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Böse, J.; Gruber, A.D.; Helming, L.; Schiebe, S.; Wegener, L.; Hafner, M.; Beales, M.; Köntgen, F.; Lengeling, A. Of Biology BioMed Central The phosphatidylserine receptor has essential functions during embryogenesis but not in apoptotic cell removal. J. Biol. 2004, 3, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeckel, J.N.; Guarani, V.; Koyanagi, M.; Roexe, T.; Lengeling, A.; Schermuly, R.T.; Gellert, P.; Braun, T.; Zeiher, A.; Stefanie Dimmeler, S. Jumonji domain-containing protein 6 (Jmjd6) is required for angiogenic sprouting and regulates splicing of VEGF-receptor 1. Proc. Natl. Acad. Sci. USA 2011, 108, 3276–3281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Si, W.; Liu, X.; He, L.; Ren, J.; Yang, Z.; Yang, J.; Li, W.; Liu, S.; Pei, F.; et al. JMJD6 promotes melanoma carcinogenesis through regulation of the alternative splicing of PAK1, a key MAPK signaling component. Mol. Cancer 2017, 16, 175. [Google Scholar] [CrossRef] [PubMed]
- Li, M.O.; Sarkisian, M.R.; Mehal, W.Z.; Rakic, P.; Flavell, R.A. Phosphatidylserine Receptor Is Required for Clearance of Apoptotic Cells. Science 2003, 80, 1560–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunisaki, Y.; Masuko, S.; Noda, M.; Inayoshi, A.; Sanui, T.; Harada, M.; Sasazuki, T.; Fukui, Y. Defective fetal liver erythropoiesis and T lymphopoiesis in mice lacking the phosphatidylserine receptor. Blood 2004, 103, 3362–3364. [Google Scholar] [CrossRef]
- Webby, C.J.; Wolf, A.; Gromak, N.; Dreger, M.; Kramer, H.; Kessler, B.M.; Nielsen, M.L.; Schmitz, C.; Butler, D.S.; Yates, J.R.; et al. Jmjd6 catalyses lysyl-hydroxylation of U2AF65, a protein associated with RNA splicing. Science 2009, 80, 90–93. [Google Scholar] [CrossRef]
- Islam, S.; McDonough, M.A.; Chowdhury, R.; Gault, J.; Khan, A.; Pires, E.; Schofield, C.J. Biochemical and structural investigations clarify the substrate selectivity of the 2-oxoglutarate oxygenase JMJD6. J. Biol. Chem. 2019, 294, 11637–11652. [Google Scholar] [CrossRef] [Green Version]
- Chang, B.; Chen, Y.; Zhao, Y.; Bruick, R.K. JMJD6 Is a Histone Arginine Demethylase. Science 2007, 80, 444–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulard, C.; Rambaud, J.; Hussein, N.; Corbo, L.; Le Romancer, M. JMJD6 regulates ERα methylation on arginine. PLoS ONE 2014, 9, e87982. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Long, Y.-H.; Wang, S.-Q.; Zhang, Y.-Y.; Li, Y.-F.; Mi, J.-S.; Yu, C.-H.; Li, D.-Y.; Zhang, J.-H.; Zhang, X.-J. JMJD6 regulates histone H2A.X phosphorylation and promotes autophagy in triple-negative breast cancer cells via a novel tyrosine kinase activity. Oncogene 2019, 38, 980–997. [Google Scholar] [CrossRef]
- Lee, S.; Liu, H.; Hill, R.; Chen, C.; Hong, X.; Crawford, F.; Kingsley, M.C.; Zhang, Q.; Liu, X.; Chen, Z.; et al. JMJD6 cleaves MePCE to release positive transcription elongation factor b (P-TEFb) in higher eukaryotes. Elife 2020, 9, e53930. [Google Scholar] [CrossRef] [PubMed]
- Heim, A.; Grimm, C.; Müller, U.; Häußler, S.; Mackeen, M.M.; Merl, J.; Hauck, S.M.; Kessler, B.M.; Schofield, C.J.; Wolf, A.; et al. Jumonji domain containing protein 6 (Jmjd6) modulates splicing and specifically interacts with arginine-serine-rich (RS) domains of SR- and SR-like proteins. Nucleic Acids Res. 2014, 42, 7833–7850. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Shen, H.-F.; Qiu, J.-S.; Huang, M.-F.; Zhang, W.-J.; Ding, J.-C.; Zhu, X.-Y.; Zhou, Y.; Fu, X.-D.; Liu, W. JMJD6 and U2AF65 co-regulate alternative splicing in both JMJD6 enzymatic activity dependent and independent manner. Nucleic Acids Res. 2017, 45, 3503–3518. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Zang, J.; White, J.; Wang, C.; Pan, C.-H.; Zhao, R.; Murphy, R.C.; Dai, S.; Henson, P.; Kappler, J.; et al. Interaction of JMJD6 with single-stranded RNA. Proc. Natl. Acad. Sci. USA 2010, 107, 14568–14572. [Google Scholar] [CrossRef] [Green Version]
- Wolf, A.; Mantri, M.; Heim, A.; Müller, U.; Fichter, E.; Mackeen, M.M.; Schermelleh, L.; Dadie, G.; Leonhardt, H.; Vénien-Bryan, C.; et al. The polyserine domain of the lysyl-5 hydroxylase Jmjd6 mediates subnuclear localization. Biochem. J. 2013, 453, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Chen, S.; Yang, Y.; Ma, X.; Shao, B.; Yang, S.; Wei, Y.; Wei, X. Jumonji domain-containing protein 6 protein and its role in cancer. Cell Prolif. 2020, 53, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Ma, Q.; Wong, K.; Li, W.; Ohgi, K.; Zhang, J.; Aggarwal, A.K.; Rosenfeld, M.G. Brd4 and JMJD6-associated anti-pause enhancers in regulation of transcriptional pause release. Cell 2013, 155, 1581–1595. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.; Sun, Y.; Xi, Z.; Milazzo, G.; Poulos, R.C.; Bartenhagen, C.; Bell, J.L.; Mayoh, C.; Ho, N.; Tee, A.E.; et al. JMJD6 is a tumorigenic factor and therapeutic target in neuroblastoma. Nat. Commun. 2019, 10, 3319. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; He, L.; Huangyang, P.; Liang, J.; Si, W.; Yan, R.; Han, X.; Liu, S.; Gui, B.; Li, W.; et al. JMJD6 Promotes Colon Carcinogenesis through Negative Regulation of p53 by Hydroxylation. PLoS Biol. 2014, 12, e1001819. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.C.; Cleland, J.; Kim, R.; Wong, W.C.; Weinstein, J.N. SpliceSeq: A resource for analysis and visualization of RNA-Seq data on alternative splicing and its functional impacts. Bioinformatics 2012, 2385–2387. [Google Scholar] [CrossRef] [Green Version]
- Hahn, P.; Böse, J.; Edler, S.; Lengeling, A. Genomic structure and expression of Jmjd6 and evolutionary analysis in the context of related JmjC domain containing proteins. BMC Genom. 2008, 9, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermann, C.J. PolyASite 2.0: A consolidated atlas of polyadenylation sites from 3′end sequencing. Nucleic Acids Res. 2020, 48, D174–D179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.F.; Miller, L.D.; Chan, X.B.; Black, M.B.; Pang, B.; Ong, C.W.; Salto-Tellez, M.; Liu, E.T.; Desai, K.V. JMJD6 is a driver of cellular proliferation and motility and a marker of poor prognosis in breast cancer. Breast Cancer Res. 2012, 14, R85. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ni, S.S.; Zhao, W.L.; Dong, X.C.; Wang, J.L. High expression of JMJD6 predicts unfavorable survival in lung adenocarcinoma. Tumor Biol. 2013, 34, 2397–2401. [Google Scholar] [CrossRef]
- Miller, T.E.; Liau, B.B.; Wallace, L.C.; Morton, A.R.; Xie, Q.; Dixit, D.; Factor, D.C.; Kim, L.J.Y.; Morrow, J.J.; Wu, Q.; et al. Transcription elongation factors represent in vivo cancer dependencies in glioblastoma. Nature 2017, 547, 355–359. [Google Scholar] [CrossRef]
- Cho, H.; Kim, T.-K.; Mancebo, H.; Lane, W.S.; Flores, O.; Reinberg, D. A protein phosphatase functions to recycle RNA polymerase II. Genes Dev. 1999, 13, 1540–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.K.; Erkelenz, S.; Rattay, S.; Dehof, A.K.; Hildebrandt, A.; Schulze-Osthoff, K.; Schaal, H.; Schwerk, C. Human SAP18 mediates assembly of a splicing regulatory multiprotein complex via its ubiquitin-like fold. RNA 2010, 16, 2442–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, P.; Wegener, I.; Burrells, A.; Böse, J.; Wolf, A.; Erck, C.; Butler, D.; Schofield, C.J.; Böttger, A.; Lengeling, A. Analysis of jmjd6 cellular localization and testing for its involvement in histone demethylation. PLoS ONE 2010, 5, e13769. [Google Scholar] [CrossRef]
- Böttger, A.; Islam, M.S.; Chowdhury, R.; Schofield, C.J.; Wolf, A. The oxygenase Jmjd6-A case study in conflicting assignments. Biochem. J. 2015, 468, 191–202. [Google Scholar] [CrossRef]
- Hentze, M.W.; Kulozik, A.E. A perfect message: RNA surveillance and nonsense-mediated decay. Cell 1999, 96, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Cho, E.J.; Kobor, M.S.; Kim, M.; Greenblatt, J.; Buratowski, S. Opposing effects of Ctk1 kinase and Fcp1 phosphatase at Ser 2 of the RNA polymerase II C-terminal domain. Genes Dev. 2001, 15, 3319–3329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.S.; Dubois, M.F.; Dahmus, M.E. TFIIF-associating carboxyl-terminal domain phosphatase dephosphorylates phosphoserines 2 and 5 of RNA polymerase II. J. Biol. Chem. 2002, 277, 45949–45956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.-F.; Krieger, K.L.; Lagundzin, D.; Li, X.; Cheung, R.S.; Taniguchi, T.; Johnson, K.R.; Bessho, T.; Monteiro, A.N.A.; Woods, N.T. CTDP1 regulates breast cancer survival and DNA repair through BRCT-specific interactions with FANCI. Cell Death Discov. 2019, 5, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Contreras, R.; Cloutier, P.; Shkreta, L.; Fisette, J.-F.; Revil, T.; Chabot, B. hnRNP proteins and splicing control. Adv. Exp. Med. Biol. 2007, 623, 123–147. [Google Scholar] [PubMed]
- Matlin, A.; Moore, M. Spliceosome assembly and composition. Adv. Exp. Med. Biol. 2007, 623, 14–35. [Google Scholar]
- Erkelenz, S.; Theiss, S.; Otte, M.; Widera, M.; Peter, J.O.; Schaal, H. Genomic HEXploring allows landscaping of novel potential splicing regulatory elements. Nucleic Acids Res. 2014, 42, 10681–10697. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Hauck, S.M.; Dietter, J.; Kraner, R.L.; Hofmaier, F.; Zipplies, J.K.; Amann, B.; Feuchtinger, A.; Deeg, C.A.; Ueffing, M. Deciphering membrane-associated molecular processes in target tissue of autoimmune uveitis by label-free quantitative mass spectrometry. Mol. Cell. Proteom. 2010, 9, 2292–2305. [Google Scholar] [CrossRef] [Green Version]
- Merl, J.; Ueffing, M.; Hauck, S.M.; Vvon Toerne, C. Direct comparison of MS-based label-free and SILAC quantitative proteome profiling strategies in primary retinal Müller cells. Proteomics 2012, 12, 1902–1911. [Google Scholar] [CrossRef]
- Brosch, M.; Yu, L.; Hubbard, T.; Choudhary, J. UKPMC Funders Group Author Manuscript Accurate and Sensitive Peptide Identification with Mascot Percolator. Proteome 2009, 8, 3176–3181. [Google Scholar]
- Hillebrand, F.; Peter, J.O.; Brillen, A.-L.; Otte, M.; Schaal, H.; Erkelenz, S. Differential hnRNP D isoform incorporation may confer plasticity to the ESSV-mediated repressive state across HIV-1 exon 3. Biochim. Biophys. Acta-Gene Regul. Mech. 2017, 1860, 205–217. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raguz, N.; Heim, A.; Engal, E.; Wesche, J.; Merl-Pham, J.; Hauck, S.M.; Erkelenz, S.; Schaal, H.; Bensaude, O.; Wolf, A.; et al. JMJD6 Regulates Splicing of Its Own Gene Resulting in Alternatively Spliced Isoforms with Different Nuclear Targets. Int. J. Mol. Sci. 2020, 21, 6618. https://doi.org/10.3390/ijms21186618
Raguz N, Heim A, Engal E, Wesche J, Merl-Pham J, Hauck SM, Erkelenz S, Schaal H, Bensaude O, Wolf A, et al. JMJD6 Regulates Splicing of Its Own Gene Resulting in Alternatively Spliced Isoforms with Different Nuclear Targets. International Journal of Molecular Sciences. 2020; 21(18):6618. https://doi.org/10.3390/ijms21186618
Chicago/Turabian StyleRaguz, Nikoleta, Astrid Heim, Eden Engal, Juste Wesche, Juliane Merl-Pham, Stefanie M. Hauck, Steffen Erkelenz, Heiner Schaal, Olivier Bensaude, Alexander Wolf, and et al. 2020. "JMJD6 Regulates Splicing of Its Own Gene Resulting in Alternatively Spliced Isoforms with Different Nuclear Targets" International Journal of Molecular Sciences 21, no. 18: 6618. https://doi.org/10.3390/ijms21186618