Polyethylene Glycol 35 as a Perfusate Additive for Mitochondrial and Glycocalyx Protection in HOPE Liver Preservation

 , , ,
, , ,
Abstract
:1. Introduction
2. Static Cold Storage (SCS) and PEG
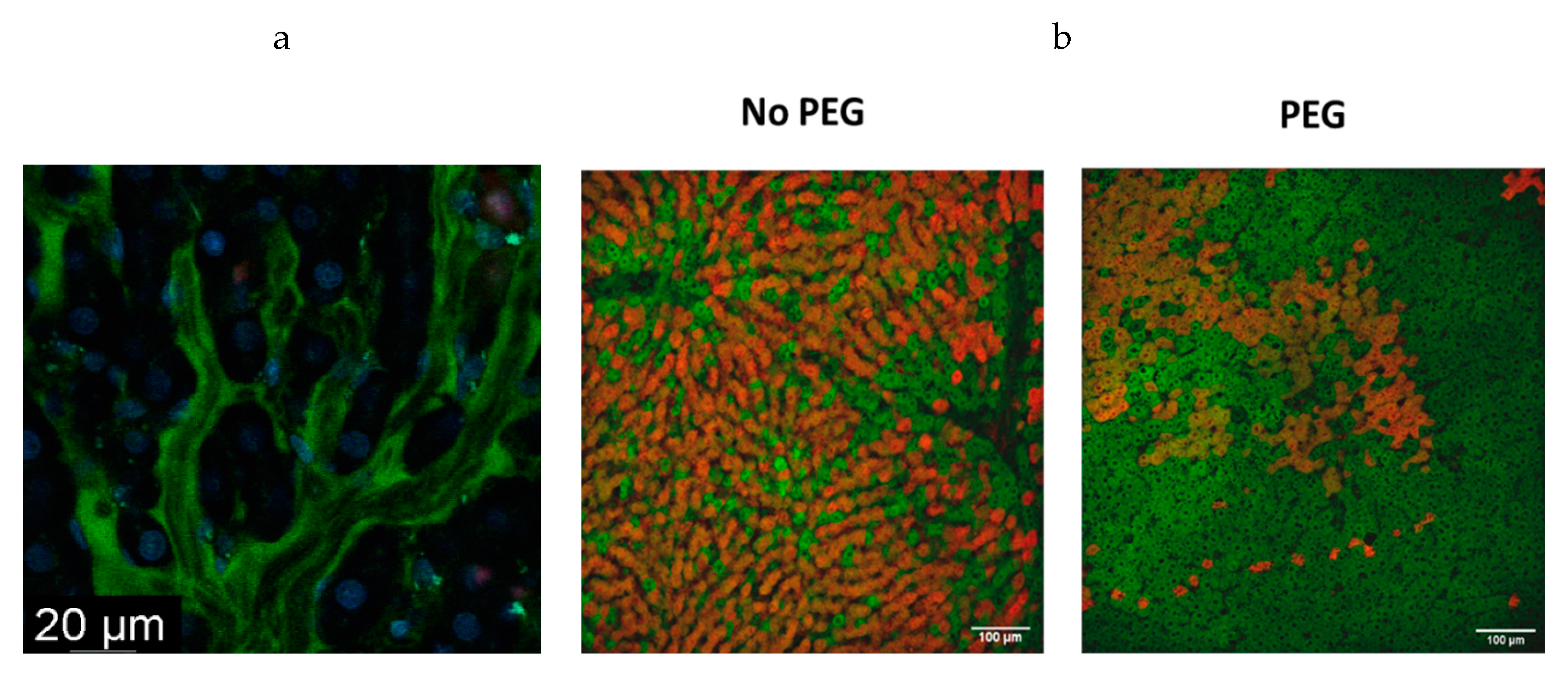
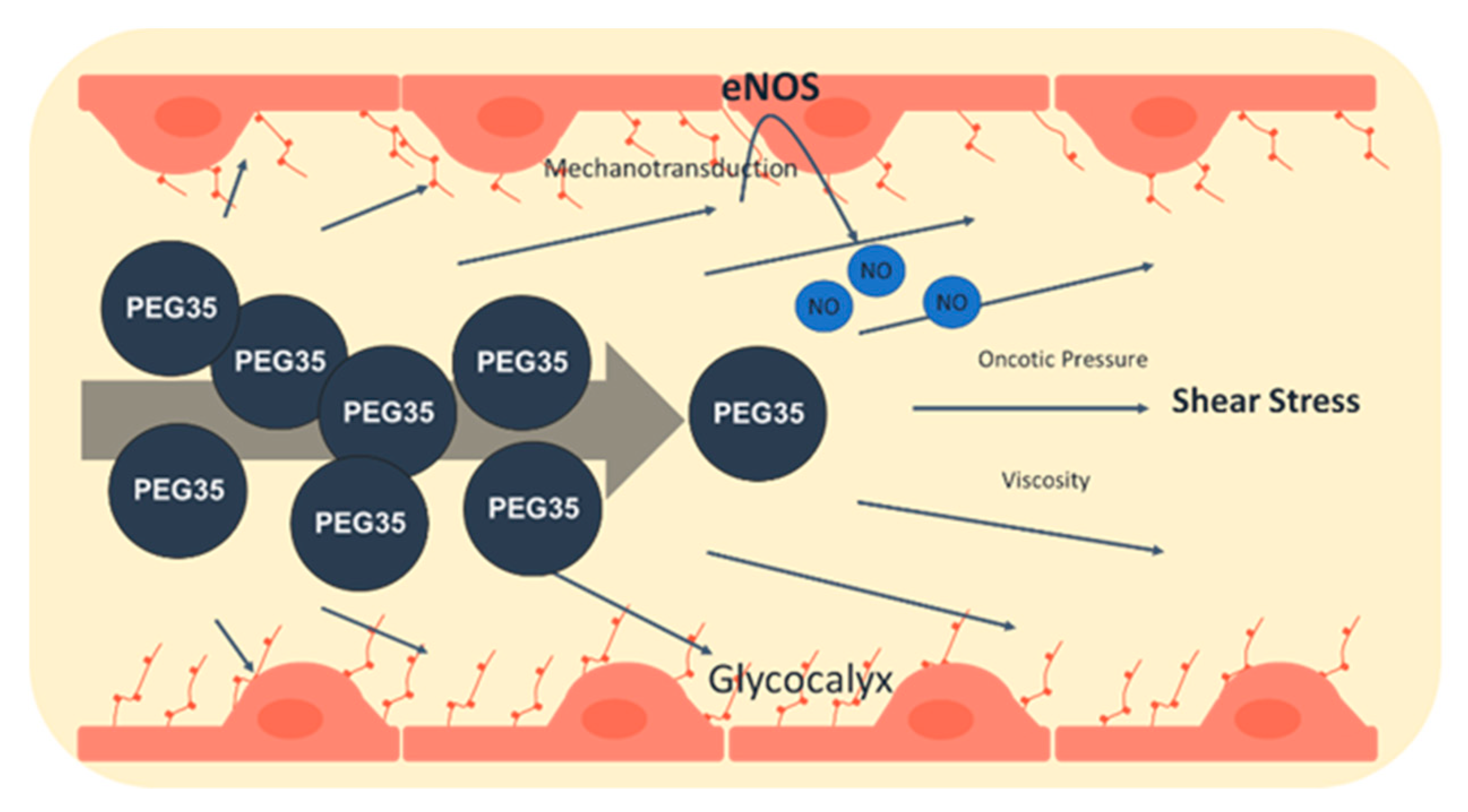
3. Endothelial Glycocalyx: Fluid Dynamics and PEG35 Effects
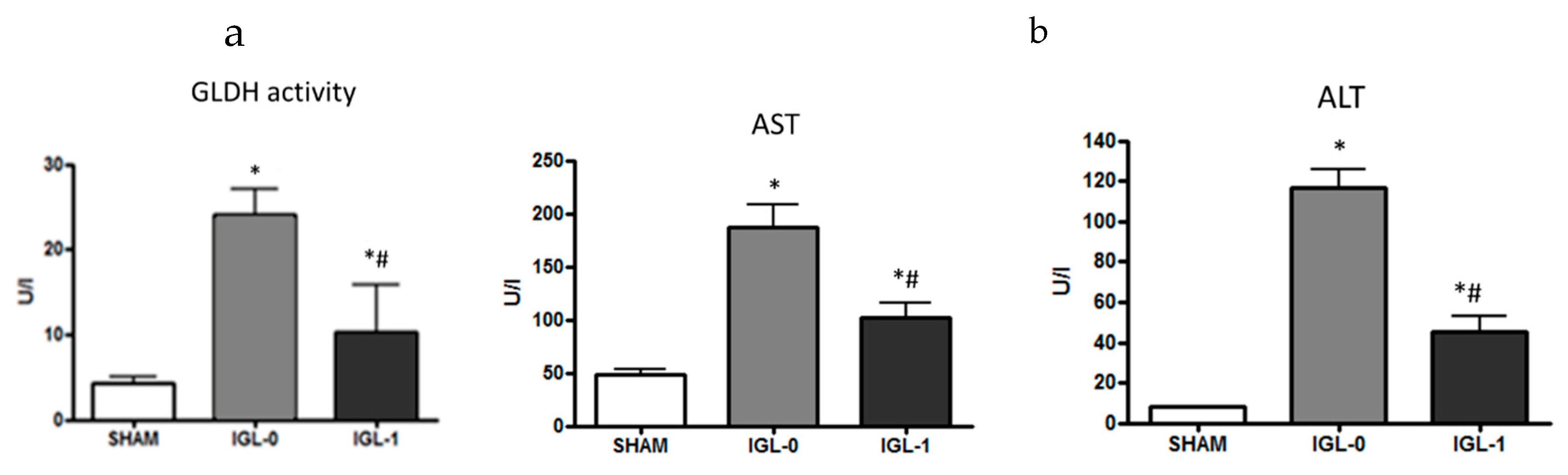
4. HOPE and PEG35 Perfusates
5. New Biomarkers for Dynamic Preservation
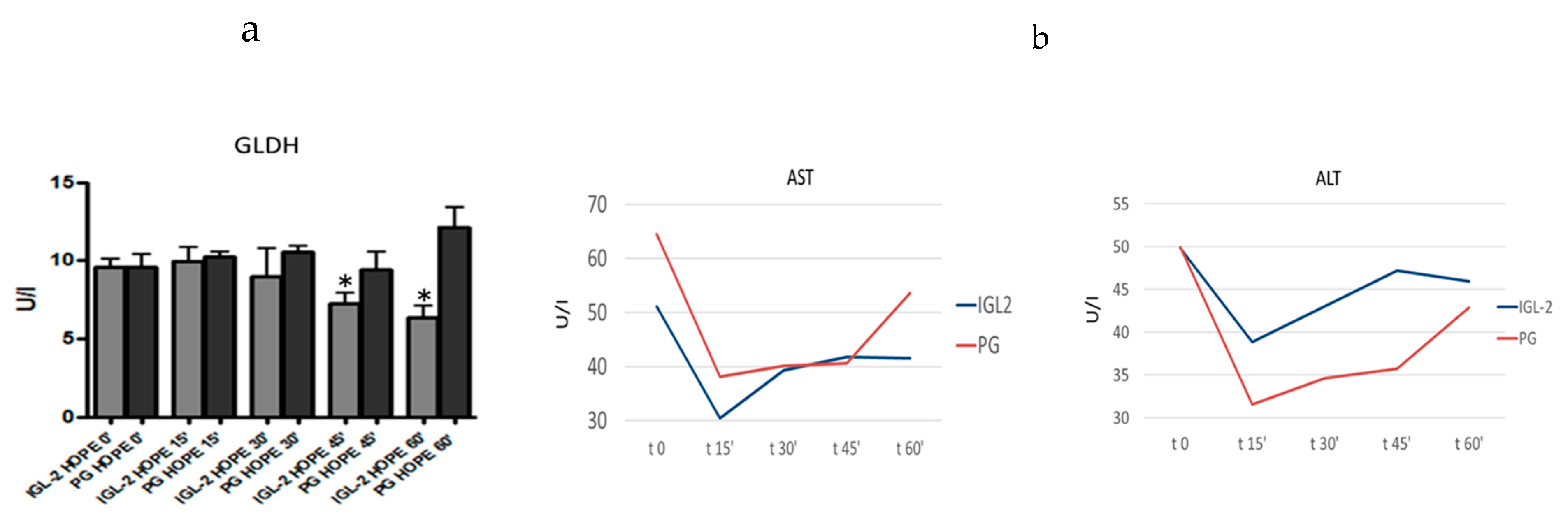
6. HOPE-COR-NMP and PEG35
7. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 4H NE | 4-Hydroxynonenal |
| ALDH2 | Aldehyde dehydrogenase-2 |
| ALT | Alanine aminotransferase |
| AMPK | AMP-activated protein kinase |
| AST | Aspartate aminotransferase |
| Belzer-MPS | Belzer Machine Perfusion Solution |
| DAMPs | Damage associated molecular patterns |
| DCD | Donor after cardiac death |
| EC | Endothelial cells |
| ECD | Extended criteria donors |
| eNOS | Endothelial nitric oxide synthase |
| GCX | Glycocalyx |
| GLDH | Glutamate dehydrogenase |
| PEG | Polyethylene glycol |
| HES | Hydroxyethyl starch |
| HIF | Hypoxia inducible factor |
| HOPE | Hypothermic oxygenated perfusion |
| HTK | Histidine-tryptophan-ketoglutarate |
| i.v. | Intravenous |
| IGL-1 | Institut Georges Lopez 1 |
| IRI | Ischemia–reperfusion injury |
| MP | Machine perfusion |
| MPTP | Mitochondrial permeability transition pore |
| NO | Nitric oxide |
| ROS | Radical oxygen species |
| SCS | Static cold storage |
| SS | Shear stress |
| TX | Liver transplantation |
| UCP1 | Uncoupling protein 1 |
| UCP2 | Uncoupling protein 2 |
| UW | University of Wisconsin |
References
- Dutkowski, P.; De Rougemont, O.; Clavien, P.A. Alexis Carrel: Genius, innovator and ideologist. Am. J. Transplant. 2008, 8, 1998–2003. [Google Scholar] [CrossRef] [PubMed]
- Zaouali, M.A.; Abdennebi, B.H.; Padrissa-Altes, S. Pharmacological strategies against cold sichemia reperfusion injury. Expert. Opin. Pharm. 2010, 11, 537–555. [Google Scholar] [CrossRef] [PubMed]
- Guibert, E.E.; Petrenko, A.Y.; Balaban, C.L.; Somov, A.Y.; Rodriguez, J.V.; Fuller, B.J. Organ Preservation: Current Concepts and New Strategies for the Next Decade. Transfus. Med. Hemother. 2011, 38, 125–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belzer, F.O.; Southard, J.H. Principles of solid-organ preservation by cold storage. Transplantation 1988, 45, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Southard, J.H.; Belzer, F.O. Organ preservation. Annu. Rev. Med. 1995, 4, 235–247. [Google Scholar] [CrossRef]
- Ben Mosbah, I.; Roselló-Catafau, J.; Franco-Gou, R.; Abdennebi, H.B.; Saidane, D.; Ramella-Virieux, S.; Boillot, O.; Peralta, C. Preservation of steatotic livers in IGL-1 solution. Liver Transplant. 2006, 12, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Menasche, P.; Termignon, J.L.; Pradier, F.; Grousset, C.; Mouas, C.; Alberici, G.; Weiss, M.; Piwnica, A.; Bloch, G. Experimental evaluation of Celsior, a new heart preservation solution. Eur. J. Cardiothorac. Surg. 1994, 8, 207–213. [Google Scholar] [CrossRef]
- Stewart, Z.A.; Cameron, A.M.; Singer, A.L.; Montgomery, R.A.; Segev, D.L. Histidine-Tryptophan-Ketoglutarate (HTK) is associated with reduced graft survival in deceased donor livers, especially those donated after cardiac death. Am. J. Transplant. 2009, 9, 286–293. [Google Scholar] [CrossRef]
- Adam, R.; Delvart, V.; Karam, V.; Ducerf, C.; Navarro, F.; Letoublon, C.; Belghiti, J.; Pezet, D.; Castaing, D.; Le Treut, Y.P.; et al. ELTR contributing centers, the European Liver, Intestine Transplant Association (ELITA). Compared efficacy of preservation solutions in liver transplantation: A long-term graft outcome study from the European Liver Transplant Registry. Am. J. Transplant. 2015, 15, 395–406. [Google Scholar] [CrossRef]
- Boni, L.T.; Hah, J.S.; Hui, S.W.; Mukherjee, P.; Ho, J.T.; Jung, C.Y. Aggregation and fusion of unilamellar vesicles by polyethylene glycol. Biochim. Biophys. Acta 1984, 775, 409–418. [Google Scholar] [CrossRef]
- Pasut, G.; Panisello, A.; Folch-Puy, E.; Lopez, A.; Castro-Benítez, C.; Calvo, M.; Carbonell, T.; García-Gil, A.; Adam, R.; Roselló-Catafau, J. Polyethylene glycols: An effective strategy for limiting liver ischemia reperfusion injury. World J. Gastroenterol. 2016, 22, 6501–6508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.R. Control of water content of non-metabolizing kidney slices by sodium chloride and polyethylene glycol (PEG 6000). J. Physiol. 1971, 213, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.R. Colloid osmotic pressure as a cause of pathological swelling of cells. Pathobiol. Cell Membr. 1975, 1, 173–289. [Google Scholar]
- Robinson, J.R. Control of water content of respiring kidney slices by sodium chloride and polyethylene glycol. J. Physiol. 1978, 282, 285–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, M.R.; Wakerley, C.L. Factors influencing the survival of cell monolayers during storage at 4 degrees. Br. J. Exp. Pathol. 1976, 57, 137–177. [Google Scholar] [PubMed]
- Ganote, C.E.; Worstell, J.; Iannotti, J.P.; Kaltenbach, J.P. Cellular swelling and irreversible myocardial injury. Effects of polyethylene glycol and mannitol in perfused rat hearts. Am. J. Pathol. 1977, 88, 95–118. [Google Scholar]
- Stefanovich, P.; Ezzell, R.M.; Sheehan, S.J.; Tompkins, R.G.; Yarmush, M.L.; Toner, M. Effects of hypothermia on the function, membrane integrity, and cytoskeletal structure of hepatocytes. Cryobiology 1995, 32, 389–403. [Google Scholar] [CrossRef]
- Wicomb, W.N.; Hill, J.D.; Avery, J.; Collins, G.M. Optimal cardioplegia and 24- hour heart storage with simplified UW solution containing polyethylene glycol. Transplantation 1990, 49, 261–263. [Google Scholar] [CrossRef]
- Zaouali, M.A.; Bejaoui, M.; Calvo, M.; Folch-Puy, E.; Pantazi, E.; Pasut, G.; Rimola, A.; Abdennebi, H.B.; Adam, R.; Roselló-Catafau, J. Polyethylene glycol rinse solution: An effective way to prevent ischemia-reperfusion injury. World J. Gastroenterol. 2015, 20, 16203–16214. [Google Scholar] [CrossRef]
- Karam, V.; Tedeschi, M.; Golse, N.; Allard, M.A.; Ciacio, O.; Pittau, G.; Vibert, E.; Cunha, S.A.; Saliba, F.; Ichai, P.; et al. Predictive Factords of Early allograft dysfunction and its impact on Long-term graft sursvival in ECD (abstract). Transplantation 2019, 103, 377–378. [Google Scholar]
- Adam, R.; Karam, V.; Sebagh, M.; Vibert, E.; Cunha, S.A.; Saliba, F.; Cherqui, D.; Samuel, D.; Castaing, D. Impact of preservation solution on early allograft dysfunction (EAD) and graft survival after liver transplantation using Expanded-Criteria-Donors (abstract). Transplantation 2017, 101, 161. [Google Scholar]
- Cano, N. Bench-to-bedside review: Glucose production from the kidney. Crit. Care 2002, 6, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Bessems, M.; Doorschodt, B.M.; van Marle, J.; Vreeling, H.; Meijer, A.J.; van Gulik, T.M. Improved machine perfusion preservation of the non-heart-beating donor rat liver using polysol: A new machine perfusion preservation solution. Liver Transplant. 2005, 11, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Panisello-Roselló, A.; Castro, C.; Rosello-Catafau, J.; Adam, R. IGL2 a new perfusate for HOPE strategies. In Proceedings of the ILTS Meeting 699, Istabul, Turkey, 30 May 2020. [Google Scholar]
- Bejaoui, M.; Pantazi, E.; Calvo, M.; Folch-Puy, E.; Serafín, A.; Pasut, G.; Panisello, A.; Adam, R.; Roselló-Catafau, J. Polyethylene Glycol Preconditioning: An Effective Strategy to Prevent Liver Ischemia Reperfusion Injury. Oxid Med. Cell Longev. 2016, 2016, 9096549. [Google Scholar] [CrossRef] [Green Version]
- Ferrero-Andrés, A.; Panisello-Roselló, A.; Serafín, A.; Roselló-Catafau, J.; Folch-Puy, E. Polyethylene Glycol 35 (PEG35) Protects against Inflammation in Experimental Acute Necrotizing Pancreatitis and Associated Lung Injury. Int. J. Mol. Sci. 2020, 21, 917. [Google Scholar] [CrossRef] [Green Version]
- Bejaoui, M.; Pantazi, E.; Folch-Puy, E.; Panisello, A.; Calvo, M.; Pasut, G.; Rimola, A.; Navasa, M.; Adam, R.; Roselló-Catafau, J. Protective Effect of Intravenous High Molecular Weight Polyethylene Glycol on Fatty Liver Preservation. Biomed. Res. Int. 2015, 2015, 794287. [Google Scholar] [CrossRef] [Green Version]
- Hauet, T.; Eugene, M. A new approach in organ preservation: Potential role of new polymers. Kidney Int. 2008, 74, 998–1003. [Google Scholar] [CrossRef] [Green Version]
- Mack, J.E.; Kerr, J.A.; Vreugdenhil, P.K.; Belzer, F.O.; Southard, J.H. Effect of polyethylene glycol on lipid peroxidation in cold-stored rat hepatocytes. Cryobiology 1991, 28, 1–7. [Google Scholar] [CrossRef]
- Dutheil, D.; Underhaug Gjerde, A.; Petit-Paris, I.; Mauco, G.; Holmsen, H. Polyethylene glycols interact with membrane glycerophospholipids: Is this part of their mechanism for hypothermic graft protection? J. Chem. Biol. 2009, 2, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Ohno, H.; Sakai, T.; Tsuchida, E.; Honda, K.; Sasakawa, S. Interaction of human erythrocyte ghosts or liposomes with polyethylene glycol detected by fluorescence polarization. Biochem. Biophys. Res. Commun. 1981, 102, 426–431. [Google Scholar] [CrossRef]
- Marsh, D.C.; Lindell, S.L.; Fox, L.E.; Belzer, F.O.; Southard, J.H. Hypothermic preservation of hepatocytes: Role of Cell Swelling. Cryobiology 1989, 26, 524–534. [Google Scholar] [CrossRef]
- Bote, G.; Bruinsma, T.; Berendsen, T.A.; Izamis, M.L.; Yeh, H.; Yarmush, M.L.; Uygun, K. Supercooling Preservation Of The Rat Liver For Transplantation. Nat. Protoc. 2015, 10, 484–494. [Google Scholar]
- de Vries, R.J.; Tessier, S.N.; Banik, P.D.; Nagpal, S.; Cronin, S.E.J.; Ozer, S.; Hafiz, E.O.A.; van Gulik, T.M.; Yarmush, M.L.; Markmann, J.F.; et al. Supercooling extends preservation time of human livers. Nat. Biotechnol. 2019, 37, 1131–1136. [Google Scholar] [CrossRef] [PubMed]
- Puts, C.F.; Berendsen, T.A.; Bruinsma, B.G.; Ozer, S.; Luitje, M.; Usta, O.B.; Yarmush, M.L.; Uygun, K. Polyethylene glycol protects primary hepatocytes during supercooling preservation. Criobiology 2015, 71, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Dutkowski, P.; Guarrera, J.V.; De Jonge, J.; Martins, P.N.; Porte, R.J.; Clavien, P.A. Evolving trends in machine perfusion for liver transpalntation. Gastroenterology 2019, 156, 1542–1547. [Google Scholar] [CrossRef] [Green Version]
- Brüggenwirth, I.M.; van Leeuwen, O.B.; de Vries, Y.; Bodewes, S.B.; Adelmeijer, J.; Wiersema-Buist, J.; Lisman, T.; Martins, P.N.; de Meijer, V.E.; Porte, R.J. Extended hypothermic oxygenated machine perfusion enables ex situ preservation of porcine livers for up to 24 h. JHep Rep. 2020, 2, 100092. [Google Scholar] [CrossRef] [Green Version]
- Schlegel, A.; Muller, X.; Kalisvaart, M.; Muellhaupt, B.; Perera, M.T.P.R.; Isaac, J.; Clavien, P.A.; Muiesan, P.; Dutkowski, P. Outcomes of liver transplantations from donation after circulatory death (DCD) treated by hypothermic oxygenated perfusion (HOPE) before implantation. J. Hepatol. 2019, 70, 50–57. [Google Scholar] [CrossRef]
- Czigany, Z.; Lurje, I.; Schmelzle, M.; Schöning, W.; Öllinger, R.; Raschzok, N.; Sauer, I.M.; Tacke, F.; Strnad, P.; Trautwein, C.; et al. Ischemia-Reperfusion Injury in Marginal Liver Grafts and the Role of Hypothermic Machine Perfusion: Molecular Mechanisms and Clinical Implications. J. Clin. Med. 2020, 9, 846. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Li, Y.S.; Chien, S. Shear stress-initiated signaling and its regulation of endothelial function. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2191–2198. [Google Scholar] [CrossRef] [Green Version]
- Rafetto, J.D.; Manello, F. Pathophysiology of chronic venous disease. Int. Angiol. 2014, 33, 212–221. [Google Scholar]
- Zeng, Y.; Zhang, X.F.; Fu, B.M.; Tarbel, J.M. The Role of Endothelial Surface Glycocalyx in Mechanosensing and Transduction. Adv. Exp. Med. Biol. 2018, 1097, 1–27. [Google Scholar] [PubMed]
- Morariu, A.M.; vd Plaats, A.; v Oeveren, W.; A’t Hart, N.; Leuvenink, H.G.; Graaff, R.; Ploeg, R.J.; Rakhorst, G. Hyperaggregating effect of hydroxyethyl starch components and UW solution on human red blood cells: A risk of impaired graft perfusion in organ procurement? Transplantation 2003, 76, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Mosbah, I.B.; Franco-Gou, R.; Abdennebi, H.B.; Hernandez, R.; Escolar, G.; Saïdane, D.; Rosello-Catafau, J.; Peralta, C. Effects of polyethylene glycol and hydroxyethyl starch in University of Wisconsin preservation solution on human red blood cell aggregation and viscosity. Transplant. Proc. 2006, 38, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, R.P.; Shapter, O.; Aitken, E.; Stevenson, K.; Shiels, P.G.; Kingsmorea, D.B. Has the Expansion in Extended Criteria Deceased Donors Led to a Different Type of Delayed Graft Function and Poorer Outcomes? Transplant. Proc. 2018, 50, 3160–3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, J.; Dong, J.; Eng, M.; Landsberg, D.; Gill, J.S. Pulsatile Perfusion Reduces the Risk of Delayed Graft Function in Deceased Donor Kidney Transplants, Irrespective of Donor Type and Cold Ischemic Time. Transplantation 2014, 97, 668–674. [Google Scholar] [CrossRef]
- Panisello-Roselló, A.; Alva, N.; Flores, M.; Lopez, A.; Castro Benítez, C.; Folch-Puy, E.; Rolo, A.; Palmeira, C.; Adam, R.; Carbonell, T.; et al. Aldehyde Dehydrogenase 2 (ALDH2) in Rat Fatty Liver Cold Ischemia Injury. Int. J. Mol. Sci. 2018, 22, 2479. [Google Scholar] [CrossRef] [Green Version]
- Kulek, A.R.; Anzell, A.; Wider, J.M.; Sanderson, T.H.; Przyklenk, K. Mitochondrial quality control: Role in cardiac models of lethal ischemia-reperfusion injury. Cells 2020, 9, 214. [Google Scholar] [CrossRef] [Green Version]
- Chiang, E.T.; Camp, S.M.; Dudek, S.M.; Brown, M.E.; Usatyuk, P.V.; Zaborina, O.; Alverdy, J.C.; Garcia, J.G. Protective effects of high molecular weight Polyethylene Glycol (PEG) in human lung endothelial cell barrier regulation: Role of actin cytoskeletal rearrangement. Microvasc. Res. 2009, 77, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Zaouali, M.A.; Ben Mosbah, I.; Boncompagni, E.; Ben Abdennebi, H.; Mitjavila, M.T.; Bartrons, R.; Freitas, I.; Rimola, A.; Roselló-Catafau, J. Hypoxia inducible factor-1 alpha accumulation in steatotic liver preservation: Role of nitric oxide. World J. Gastroenterol. 2010, 16, 3499–3509. [Google Scholar] [CrossRef] [Green Version]
- Zaoualí, M.A.; Mosbah, I.B.; Abdennebi, H.B.; Calvo, M.; Boncompagni, E.; Boillot, O.; Peralt, C.; Roselló-Catafau, J. New insights into fatty liver preservation using Institute Georges Lopez preservation solution. Transplant. Proc. 2010, 42, 159–161. [Google Scholar] [CrossRef] [Green Version]
- Bouma, H.R.; Ketelaar, M.E.; Yard, B.A.; Ploeg, R.J.; Henning, R.H. AMP-activated protein kinase as a target for preconditioning in transplantation an medicine. Transplantation 2010, 90, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Mosbah, I.; Massip-Salcedo, M.; Fernández-Monteiro, I.; Xaus, C.; Bartrons, R.; Boillot, O.; Roselló-Catafau, J.; Peralta, C. Addition of adenosine monophosphate-activated protein kinase activators to University of Wisconsin solution: A way of protecting rat steatotic livers. Liver Transplant. 2007, 13, 410–425. [Google Scholar] [CrossRef] [PubMed]
- Abdennebi, H.B.; Zaoualí, M.A.; Alfany-Fernandez, I.; Tabka, D.; Roselló-Catafau, J. How to protect liver graft with nitric oxide. World J. Gastroenterol. 2011, 17, 2879–2889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho, F.S.; Fernandez-Monteiro, I.; Rosello-Catafau, J.; Peralta, C. Hepatic microcirculatory failure. Acta Cir. Bras. 2006, 21, 48–53. [Google Scholar] [CrossRef]
- Carrasco-Chaumel, E.; Roselló-Catafau, J.; Bartrons, R.; Franco-Gou, R.; Xaus, C.; Casillas, A.; Gelpí, E.; Rodés, J.; Peralta, C. Adenosine monophosphate activated protein kinase and nitric oxide in rat steatotic liver transplantation. J. Hepatol. 2005, 43, 97–104. [Google Scholar] [CrossRef]
- Padrissa-Altés, S.; Zauali, M.A.; Roselló-Catafau, J. AMP-activated protein kinase as target for preconditioning in transplantation medicine. Transplantation 2010, 90, 12. [Google Scholar] [CrossRef]
- Tarbell, J.M.; Cancel, L.M. The glycocalyx and its significance in human medicine. J. Intern. Med. 2016, 80, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Alphonsus, C.S.; Rodseth, R.N. The endothelial glycocalyx: A review of the vascular barrier. Anaestesia 2014, 69, 777–784. [Google Scholar] [CrossRef]
- Ebong, E.E.; Lopez-Quintero, S.V.; Tarbell, J.M. Shear induced endothelial NOS activation and remodeling via heparan sulfate, glypican-1, and syndecan-1. Integr. Biol. 2014, 6, 338–347. [Google Scholar] [CrossRef] [Green Version]
- Weinbaum, S.; Zhang, X.; Cowin, S.C. Mechanotransduction and flow across the endothelial glycocalyx. Proc. Natl. Acad. Sci. USA 2003, 100, 7988–7995. [Google Scholar] [CrossRef] [Green Version]
- Pahakis, M.Y.; Kosky, J.R.; Tarbell, J.M. The role of endothelial glycocalyx components in mechanotransduction of fluid shear stress. Biochem. Biophys. Res. Commun. 2007, 355, 228–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiefer, J.; Faybik, P.; Koch, M.D.S.; Tudor, B.; Kollmann, D.; Kuessel, L.; Krenn, C.G.; Berlakovich, G.; Baron, D.M.; Baron-Stefaniak, J. Glycocalyx Damage within human Liver Grafts Correlates with Graft Injury and Postoperative Graft Function after orthotopic Liver Transplantation. Transplantation 2020, 104, 72–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, A.; Panisello-Rosello, A.; Castro-Benitez, C.; Adam, R. Glycocalyx preservation and NO production in fatty livers—The protective role of high molecular polyethylene glycol in cold ischemia Injury. Int. J. Mol. Sci. 2018, 19, 2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessheimer, A.J.; Fondevila, C.; García-Valdecasas, J.C. Extracorporeal perfusion for resuscitation of marginal grafts. In Transplantation of the Liver, 3rd ed.; Elsevier Inc.: Philadelphia, PA, USA, 2015. [Google Scholar]
- Schlegel, A.; Kron, P.; Graf, R.; Dutkowski, P.; Clavien, P.A. Warm vs. cold perfusion techniques to rescue rodent liver grafts. J. Hepatol. 2014, 61, 1267–1275. [Google Scholar] [CrossRef]
- Kron, P.; Schlegel, A.; Mancina, L.; Clavien, P.A.; Dutkowski, P. Hypothermic oxygenated perfusion (HOPE) for fatty liver grafts in rats and humans. J. Hepatol. 2018, 68, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Dutkowski, P.; Furrer, K.; Tian, Y.; Graf, R.; Clavien, P.A. Novel short-term hypothermic oxygenated perfusion (HOPE) system prevents injury in rat liver graft from non-heart beating donor. Ann. Surg. 2006, 244, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Bessems, M.; Doorschodt, B.M.; Hooijschuur, O.; Van Vliet, A.K.; Van Gulik, T.M. Optimization of a new preservation solution for machine perfusion of the liver: Which is the preferred colloid? Transplant. Proc. 2005, 37, 329–331. [Google Scholar] [CrossRef]
- Maio, R.; Costa, P.; Figueiredo, N.; Santos, I. Evaluation of different preservation solutions utilized in the machine perfusion of kidneys retrieved under cardiac arrest. An experimental study. Rev. Port. Cir. Cardiotorac. Vasc. 2007, 14, 149–156. [Google Scholar]
- Schlegel, A.; Rougemont, O.; De Graf, R.; Clavien, P.A.; Dutkowski, P. Protective mechanisms of end-ischemic cold machine perfusion in DCD liver grafts. J. Hepatol. 2013, 58, 278–286. [Google Scholar] [CrossRef]
- van Golen, R.F.; Reiniers, M.J.; Vrisekoop, N.; Zuurbier, C.J.; Olthof, P.B.; van Rheenen, J.; van Gulik, T.M.; Parsons, B.J.; Heger, M. The mechanisms and physiological relevance of glycocalyx degradation in hepatic ischemia/reperfusion injury. Antioxid. Redox Signal 2014, 21, 1098–1118. [Google Scholar] [CrossRef] [Green Version]
- Panisello-Roselló, A.; Lopez, A.; Folch-Puy, E.; Carbonell, T.; Rolo, A.; Palmeira, C.; Adam, R.; Net, M.; Roselló-Catafau, J. Role of aldehyde dehydrogenase 2 in ischemia reperfusion injury: An update. World J. Gastroenterol. 2018, 24, 29. [Google Scholar] [CrossRef] [PubMed]
- Panisello-Rosello, A.; Castro, C.; Lopez, A.; Teixeira da Silva, R.; Rosello-Catafau, J.; Adam, R. Glycocalyx as useful marker of endothelial injury in liver transplantation: The role of preservation solution. Transplantation 2020, in press. [Google Scholar]
- Hoffman, J.; Otarashvili, G.; Meszaros, A.; Ebner, S.; Weissenbacher, A.; Cardini, B.; Oberhuber, R.; Resch, T.; Ofner, D.; Schneeberger, S.; et al. Restoring mitochondrial Function While Avoiding Redox Stress: The Key to Preventing Ischemia/Reperfusion Injury in Machine Perfused Liver Grafts? Int. J. Mol. Sci. 2020, 21, 3132. [Google Scholar] [CrossRef] [PubMed]
- Jochmans, I.; Akhtar, M.Z.; Nasralla, D.; Kocabayoglu, P.; Boffa, C.; Kaisar, M.; Brat, A.; O’Callaghan, J.; Pengel, L.H.M.; Knight, S.; et al. Past, Present, and Future of Dynamic Kidney and Liver Preservation and Resuscitation. Am. J. Transplant. 2016, 16, 2545–2555. [Google Scholar] [CrossRef] [Green Version]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijević, D.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, A.; Sosunov, S.; Niatsetskaya, Z.; Konrad, C.; Starkov, A.; Manfredi, G.; Wittig, I.; Ten, V.; Galkin, A. Redox-Dependent Loss of Flavin by Mitochondrial Complex I in Brain Ischemia/Reperfusion Injury. Antioxid. Redox Signal 2019, 20, 608–622. [Google Scholar] [CrossRef]
- Jia, P.; Wu, X.; Pan, T.; Xu, S.; Hu, J.; Din, X. Uncoupling protein 1 inhibits mitchondrial reactive oxygen species generation and alleviates acute kidney injury. EBioMedicine 2019, 49, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Wan, C.D.; Wang, C.Y.; Liu, T.; Cheng, R.; Wang, H.B. Alleviation on ischemia/reperfusion injury in ob/ob mice by inhibiting UCP2 expression in fatty liver. World J. Gastroenterol. 2008, 14, 590–594. [Google Scholar] [CrossRef]
- Mattiasson, G.; Sullivan, P.G. The emerging functions of UCP2 in health, disease, and therapeutics. Antioxid. Redox Signal 2006, 8, 1–38. [Google Scholar] [CrossRef]
- Ninomiya, M.; Shirabe, K.; Shimada, M.; Terashi, T.; Maehara, Y. Role of UCP2 expression after hepatic warm ischemia-reperfusion in the rat. Gut Liver 2011, 5, 486–492. [Google Scholar] [CrossRef]
- Uchino, S.; Yamaguchi, Y.; Furuhashi, T.; Wang, F.S.; Zhang, J.L.; Okabe, K.; Kihara, S.; Yamada, S.; Mor, K.; Ogawa, M. Steatotic liver allografts up-regulate UCP-2 expression and suffer necrosis in rats. J. Surg. Res. 2004, 120, 73–82. [Google Scholar] [CrossRef]
- van Leeuwen, O.B.; de Vries, Y.; Fujiyoshi, M.; Nijsten, M.W.; Ubbink, R.; Pelgrim, G.J.; Werner, M.J.; Reyntjens, K.M.; van den Berg, A.P.; de Boer, M.T.; et al. Transplantation of High-risk Donor Livers After Ex Situ Resuscitation and Assessment Using Combined Hypo- and Normothermic Machine Perfusion: A Prospective Clinical Trial. Ann. Surg. 2019, 270, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Von Horn, C.; Minor, T. Transient hyperthermia during oxygenated rewarming of isolated rat livers. Transplant. Int. 2020, 33, 272–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosello-Catafau, J. New trends in transient hyperthermia and liver transplantation. Transplant. Int. 2020, 33, 270–271. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preservation Solution | PEG (kB) | Concentration (g/L) |
|---|---|---|
| IGL-1 | 35 | 1 |
| IGL-2 | 35 | 5 |
| Polysol | 35 | 20 |
| SCOT | 20 | 15 |
| Preservation Solution | Belzer-MPS | IGL-2 |
|---|---|---|
| Electrolytes (mmol/L) | ||
| K + | 25 | 25 |
| Na + | 120 | 125 |
| Mg2 + | 5 | 5 |
| SO4 2- | 5 | 5 |
| Ca + | 0.5 | |
| Zn 2+ | 0.091 | |
| Buffers (mmol/L) | ||
| Phosphate | 25 | 25 |
| HEPES | 10 | |
| Histidine | 30 | |
| Impermeants (mmol/L) | ||
| Mannitol | 30 | 60 |
| Lactobionic acid | 80 | |
| Dextrose | 10 | |
| Ribose | 5 | |
| Gluconate | 85 | |
| Colloids (g/L) | ||
| Hydroxyethyle starch | 50 | |
| Polyethylene glycol-35 | 5 | |
| Antioxydants (mmol/L) | ||
| Glutathione | 3 | 9 |
| Metabolic precursors (mmol/L) | ||
| Adenosine | 5 | |
| Adenine | 5 | |
| NaNO2 (nmol/L) | 50 | |
| pH | 7.4 | 7.4 |
| Osmolarity (mosmol/L) | 320 | 320 |
| Viscosity (cP) | 2.4 | 1.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panisello Rosello, A.; Teixeira da Silva, R.; Castro, C.; G. Bardallo, R.; Calvo, M.; Folch-Puy, E.; Carbonell, T.; Palmeira, C.; Roselló Catafau, J.; Adam, R. Polyethylene Glycol 35 as a Perfusate Additive for Mitochondrial and Glycocalyx Protection in HOPE Liver Preservation. Int. J. Mol. Sci. 2020, 21, 5703. https://doi.org/10.3390/ijms21165703
Panisello Rosello A, Teixeira da Silva R, Castro C, G. Bardallo R, Calvo M, Folch-Puy E, Carbonell T, Palmeira C, Roselló Catafau J, Adam R. Polyethylene Glycol 35 as a Perfusate Additive for Mitochondrial and Glycocalyx Protection in HOPE Liver Preservation. International Journal of Molecular Sciences. 2020; 21(16):5703. https://doi.org/10.3390/ijms21165703
Chicago/Turabian StylePanisello Rosello, Arnau, Rui Teixeira da Silva, Carlos Castro, Raquel G. Bardallo, Maria Calvo, Emma Folch-Puy, Teresa Carbonell, Carlos Palmeira, Joan Roselló Catafau, and René Adam. 2020. "Polyethylene Glycol 35 as a Perfusate Additive for Mitochondrial and Glycocalyx Protection in HOPE Liver Preservation" International Journal of Molecular Sciences 21, no. 16: 5703. https://doi.org/10.3390/ijms21165703