Biotechnological Approaches: Gene Overexpression, Gene Silencing, and Genome Editing to Control Fungal and Oomycete Diseases in Grapevine



Abstract
:1. Introduction
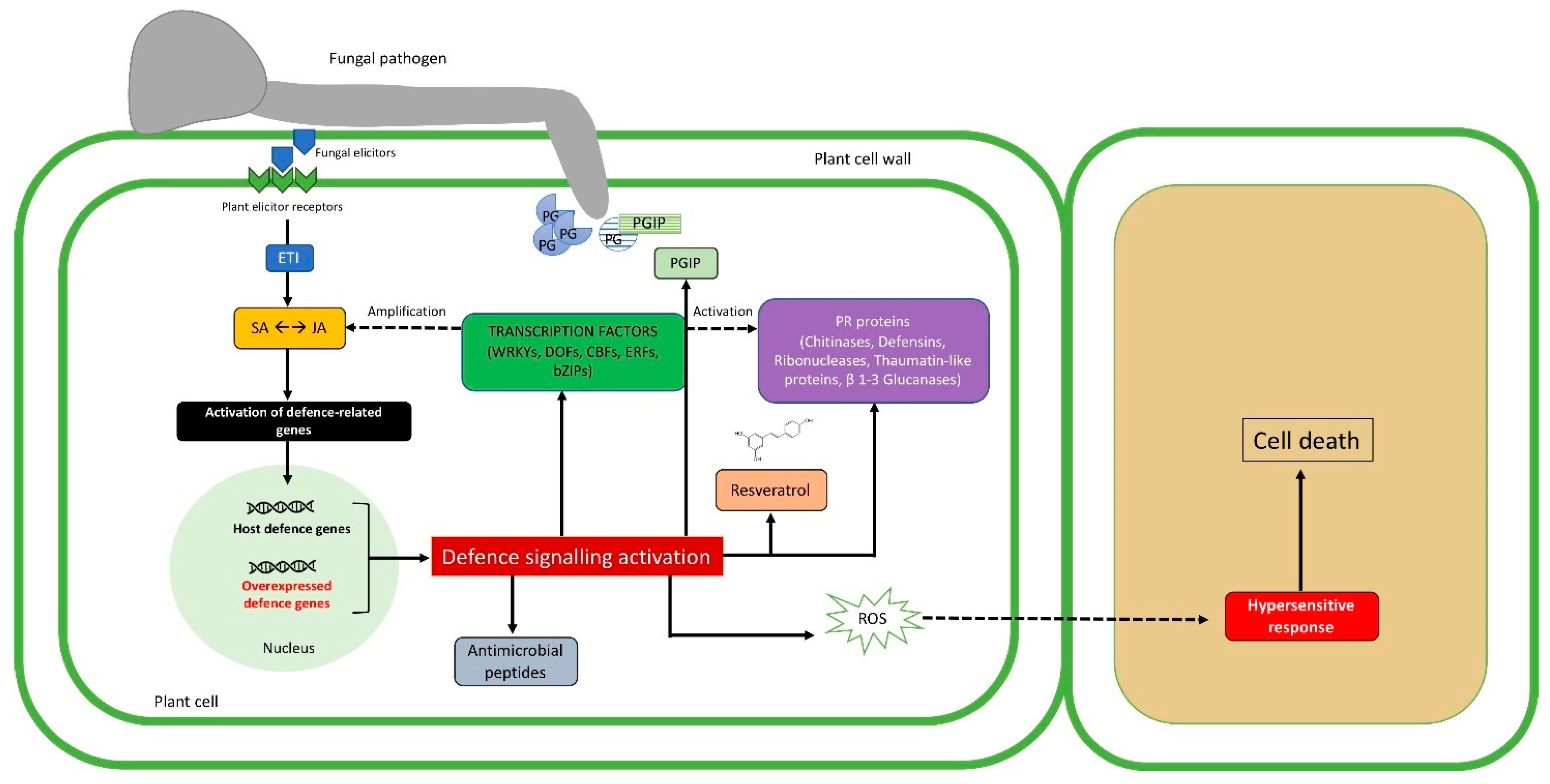
2. Plant Response Mechanisms to Pathogenic Attacks
3. Genetic Engineering for the Expression of Candidate Genes Involved in Fungal-Oomycete Resistance
3.1. Overexpression of Pathogenesis-Related Proteins
3.2. Gene Expression of Antimicrobial Peptides
3.3. Overexpression of Transcriptional Factors
3.4. Overexpression of Secondary Metabolites Generally Induced by Biotic Stress
3.5. Overexpression or Gene Expression of Other Defense-Related Genes
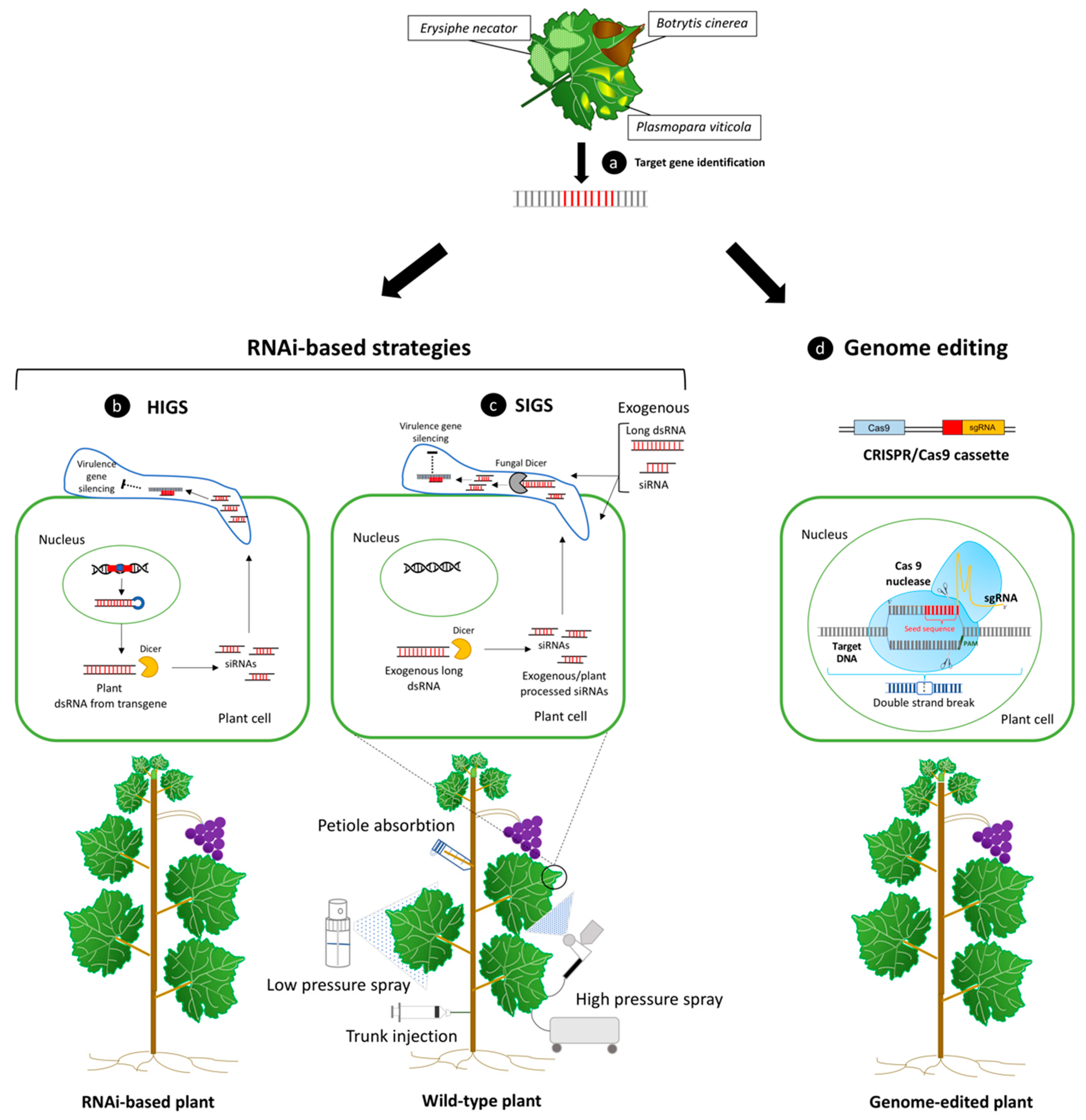
4. RNAi: Host- or Spray-Induced Gene Silencing against Fungi and Oomycetes
5. Genome Editing
6. Biosafety Considerations and Overview of Breeding Technologies Applied to Enhance Resistance against Fungal and Oomycetes Disease in Grapevine
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fournier-Level, A.; Lacombe, T.; Le Cunff, L.; Boursiquot, J.M.; This, P. Evolution of the VvMybA gene family, the major determinant of berry colour in cultivated grapevine (Vitis vinifera L.). Heredity 2010, 104, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzulli, S.; Zulini, L.; Stefanini, M. Genetics-assisted breeding for downy/powdery mildew and phylloxera resistance at fem. BIO Web Conf. 2019, 12, 01020. [Google Scholar] [CrossRef] [Green Version]
- De la Fuente Lloreda, M. Use of hybrids in viticulture. A challenge for the OIV. OENO One 2018, 52, 231–234. [Google Scholar] [CrossRef]
- Real, A.C.; Borges, J.; Cabral, J.S.; Jones, G.V. Partitioning the grapevine growing season in the Douro Valley of Portugal: Accumulated heat better than calendar dates. Int. J. Biometeorol. 2015, 59, 1045–1059. [Google Scholar] [CrossRef] [PubMed]
- Armijo, G.; Espinoza, C.; Loyola, R.; Restovic, F.; Santibáñez, C.; Schlechter, R.; Agurto, M.; Arce-Johnson, P. Grapevine Biotechnology: Molecular Approaches Underlying Abiotic and Biotic Stress Responses. In Grape and Wine Biotechnology; IntechOpen: Rijeka, Croatia, 2016. [Google Scholar] [CrossRef] [Green Version]
- Bois, B.; Zito, S.; Calonnec, A. Climate vs grapevine pests and diseases worldwide: The first results of a global survey. OENO One 2017, 51, 133–139. [Google Scholar] [CrossRef]
- Cortiñas Rodríguez, J.A.; González-Fernández, E.; Fernández-González, M.; Vázquez-Ruiz, R.A.; Aira, M.J. Fungal Diseases in Two North-West Spain Vineyards: Relationship with Meteorological Conditions and Predictive Aerobiological Model. Agronomy 2020, 10, 219. [Google Scholar] [CrossRef] [Green Version]
- Boso, S.; Gago, P.; Santiago, J.L.; La Fuente, M.D.; Martínez, M.C. Factors affecting the vineyard populational diversity of Plasmopara viticola. Plant Pathol. J. 2019, 35, 125–136. [Google Scholar] [CrossRef]
- Martínez-Bracero, M.; Alcázar, P.; Velasco-Jiménez, M.J.; Galán, C. Fungal spores affecting vineyards in Montilla-Moriles Southern Spain. Eur. J. Plant Pathol. 2019, 153, 1–13. [Google Scholar] [CrossRef]
- Toffolatti, S.L.; De Lorenzis, G.; Costa, A.; Maddalena, G.; Passera, A.; Bonza, M.C.; Pindo, M.; Stefani, E.; Cestaro, A.; Casati, P.; et al. Unique resistance traits against downy mildew from the center of origin of grapevine (Vitis vinifera). Sci. Rep. 2018, 8, 12523. [Google Scholar] [CrossRef]
- Dalla Costa, L.; Malnoy, M.; Gribaudo, I. Breeding next generation tree fruits: Technical and legal challenges. Hortic. Res. 2017, 4, 17067. [Google Scholar] [CrossRef] [Green Version]
- Guimier, S.; Delmotte, F.; Miclot, A.S.; Fabre, F.; Mazet, I.; Couture, C.; Schneider, C.; Delière, L. OSCAR, a national observatory to support the durable deployment of disease-resistant grapevine cultivars. Acta Hortic. 2019, 1248, 21–33. [Google Scholar] [CrossRef]
- Hussain, S.; Siddique, T.; Saleem, M.; Arshad, M.; Khalid, A. Chapter 5 Impact of Pesticides on Soil Microbial Diversity, Enzymes, and Biochemical Reactions. Adv. Agron. 2009, 102, 159–200. [Google Scholar] [CrossRef]
- Parmar, N.; Singh, K.H.; Sharma, D.; Singh, L.; Kumar, P.; Nanjundan, J.; Khan, Y.J.; Chauhan, D.K.; Thakur, A.K. Genetic engineering strategies for biotic and abiotic stress tolerance and quality enhancement in horticultural crops: A comprehensive review. 3 Biotech 2017, 7, 239. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Lawrence, C.B.; Chambers, O.; Davies, H.M.; Everett, N.P.; Li, Q.Q. Increased pathogen resistance and yield in transgenic plants expressing combinations of the modified antimicrobial peptides based on indolicidin and magainin. Planta 2006, 223, 1024–1032. [Google Scholar] [CrossRef]
- Wally, O.; Punja, Z.K. Genetic engineering for increasing fungal and bacterial disease resistance in crop plants. GM Crop. 2010, 1, 199–206. [Google Scholar] [CrossRef]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212–213, 29–37. [Google Scholar] [CrossRef]
- Gentile, A.; La Malfa, S. New Breeding Techniques for Sustainable Agriculture. In Innovations in Sustainable Agriculture; Springer International Publishing: Cham, Germany, 2019; pp. 411–437. [Google Scholar] [CrossRef]
- Holme, I.B.; Wendt, T.; Holm, P.B. Intragenesis and cisgenesis as alternatives to transgenic crop development. Plant Biotechnol. J. 2013, 11, 395–407. [Google Scholar] [CrossRef]
- Limera, C.; Sabbadini, S.; Sweet, J.B.; Mezzetti, B. New biotechnological tools for the genetic improvement of major woody fruit species. Front. Plant Sci. 2017, 8, 1418. [Google Scholar] [CrossRef]
- Taning, C.N.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Mezzetti, B.; Sabbadini, S.; Sorteberg, H.; Sweet, J.; Ventura, V.; et al. RNA-based biocontrol compounds: Current status and perspectives to reach the market. Pest Manag. Sci. 2020, 76, 841–845. [Google Scholar] [CrossRef]
- Kortekamp, A.; Zyprian, E. Leaf Hairs as a Basic Protective Barrier against Downy Mildew of Grape. J. Phytopathol. 1999, 147, 453–459. [Google Scholar] [CrossRef]
- Voigt, C.A. Callose-mediated resistance to pathogenic intruders in plant defense-related papillae. Front. Plant Sci. 2014, 5, 168. [Google Scholar] [CrossRef] [PubMed]
- Ellinger, D.; Naumann, M.; Falter, C.; Zwikowics, C.; Jamrow, T.; Manisseri, C.; Somerville, S.C.; Voigt, C.A. Elevated early callose deposition results in complete penetration resistance to powdery mildew in Arabidopsis. Plant Physiol. 2013, 161, 1433–1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.A.; Sundelin, T.; Nielsen, J.T.; Erbs, G. MAMP (microbe-associated molecular pattern) triggered immunity in plants. Front. Plant Sci. 2013, 4, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boutrot, F.; Zipfel, C. Function, Discovery, and Exploitation of Plant Pattern Recognition Receptors for Broad-Spectrum Disease Resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef] [PubMed]
- Poltronieri, P.; Marrazzo, M.T.; Cipriani, G. Grapevine: Resistance genes, sRNAs and immunity. In Applied Plant Biotechnology for Improving Resistance to Biotic Stress; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 151–179. [Google Scholar] [CrossRef]
- Gong, P.; Riemann, M.; Dong, D.; Stoeffler, N.; Gross, B.; Markel, A.; Nick, P. Two grapevine metacaspase genes mediate ETI-like cell death in grapevine defence against infection of Plasmopara viticola. Protoplasma 2019, 256, 951–969. [Google Scholar] [CrossRef]
- Le Henanff, G.; Farine, S.; Kieffer-Mazet, F.; Miclot, A.S.; Heitz, T.; Mestre, P.; Bertsch, C.; Chong, J. Vitis vinifera VvNPR1.1 is the functional ortholog of AtNPR1 and its overexpression in grapevine triggers constitutive activation of PR genes and enhanced resistance to powdery mildew. Planta 2011, 234, 405–417. [Google Scholar] [CrossRef]
- Yamamoto, T.; Iketani, H.; Ieki, H.; Nishizawa, Y.; Notsuka, K.; Hibi, T.; Hayashi, T.; Matsuta, N. Transgenic grapevine plants expressing a rice chitinase with enhanced resistance to fungal pathogens. Plant Cell Rep. 2000, 19, 639–646. [Google Scholar] [CrossRef]
- Bornhoff, B.A.; Harst, M.; Zyprian, E.; Töpfer, R. Transgenic plants of Vitis vinifera cv. Seyval blanc. Plant Cell Rep. 2005, 24, 433–438. [Google Scholar] [CrossRef]
- Nirala, N.K.; Das, D.K.; Srivastava, P.S.; Sopory, S.K.; Upadhyaya, K.C. Expression of a rice chitinase gene enhances antifungal potential in transgenic grapevine (Vitis vinifera L.). Vitis 2010, 49, 181–187. [Google Scholar]
- Nookaraju, A.; Agrawal, D.C. Enhanced tolerance of transgenic grapevines expressing chitinase and β-1,3-glucanase genes to downy mildew. Plant Cell Tissue Organ Cult. 2012, 111, 15–28. [Google Scholar] [CrossRef]
- Dai, L.; Wang, D.; Xie, X.; Zhang, C.; Wang, X.; Xu, Y.; Wang, Y.; Zhang, J. The Novel Gene VpPR4-1 from Vitis pseudoreticulata Increases Powdery Mildew Resistance in Transgenic Vitis vinifera L. Front. Plant Sci. 2016, 7, 695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhekney, S.A.; Li, Z.T.; Gray, D.J. Grapevines engineered to express cisgenic Vitis vinifera thaumatin-like protein exhibit fungal disease resistance. In Vitro Cell. Dev. Biol.-Plant 2011, 47, 458–466. [Google Scholar] [CrossRef]
- Yan, X.; Qiao, H.; Zhang, X.; Guo, C.; Wang, M.; Wang, Y.; Wang, X. Analysis of the grape (Vitis vinifera L.) thaumatin-like protein (TLP) gene family and demonstration that TLP29 contributes to disease resistance. Sci. Rep. 2017, 7, 4269. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Wu, J.; Zhang, Y.; Agüero, C.B.; Li, X.; Liu, S.; Wang, C.; Walker, M.A.; Lu, J. Overexpression of a thaumatin-like protein gene from Vitis amurensis improves downy mildew resistance in Vitis vinifera grapevine. Protoplasma 2017, 254, 1579–1589. [Google Scholar] [CrossRef]
- Su, H.; Jiao, Y.T.; Wang, F.F.; Liu, Y.E.; Niu, W.L.; Liu, G.T.; Xu, Y. Overexpression of VpPR10.1 by an efficient transformation method enhances downy mildew resistance in V. vinifera. Plant Cell Rep. 2018, 37, 819–832. [Google Scholar] [CrossRef]
- Vidal, J.J.; Kikkert, J.R.; Malnoy, M.A.; Wallace, P.G.; Barnard, J.; Reisch, B.I. Evaluation of transgenic “Chardonnay” (Vitis vinifera) containing magainin genes for resistance to crown gall and powdery mildew. Transgenic Res. 2006, 15, 69–82. [Google Scholar] [CrossRef]
- Rosenfield, C.-L.; Samuelian, S.; Vidal, J.R.; Reisch, B.I. Transgenic Disease Resistance in Vitis vinifera: Potential Use and Screening of Antimicrobial Peptides. Am. J. Enol. Vitic. 2010, 61, 348–357. [Google Scholar]
- Mzid, R.; Marchive, C.; Blancard, D.; Deluc, L.; Barrieu, F.; Corio-Costet, M.F.; Drira, N.; Hamdi, S.; Lauvergeat, V. Overexpression of VvWRKY2 in tobacco enhances broad resistance to necrotrophic fungal pathogens. Physiol. Plant. 2007, 131, 434–447. [Google Scholar] [CrossRef]
- Merz, P.R.; Moser, T.; Höll, J.; Kortekamp, A.; Buchholz, G.; Zyprian, E.; Bogs, J. The transcription factor VvWRKY33 is involved in the regulation of grapevine (Vitis vinifera) defense against the oomycete pathogen Plasmopara viticola. Physiol. Plant. 2015, 153, 365–380. [Google Scholar] [CrossRef]
- Zhu, Z.; Shi, J.; Cao, J.; He, M.; Wang, Y. VpWRKY3, a biotic and abiotic stress-related transcription factor from the Chinese wild Vitis pseudoreticulata. Plant Cell Rep. 2012, 31, 2109–2120. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Shi, J.; Xu, W.; Li, H.; He, M.; Xu, Y.; Xu, T.; Yang, Y.; Cao, J.; Wang, Y. Three ERF transcription factors from Chinese wild grapevine Vitis pseudoreticulata participate in different biotic and abiotic stress-responsive pathways. J. Plant Physiol. 2013, 170, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Folta, K.M.; Xie, Y.; Jiang, W.; Lu, J.; Zhang, Y. Overexpression of Muscadinia rotundifolia CBF2 gene enhances biotic and abiotic stress tolerance in Arabidopsis. Protoplasma 2017, 254, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.H.; Jiao, Z.L.; Bian, L.; Wan, Y.T.; Yu, K.K.; Zhang, G.H.; Guo, D.L. Overexpression of Vitis vinifera VvbZIP60 enhances Arabidopsis resistance to powdery mildew via the salicylic acid signaling pathway. Sci. Hortic. 2019, 256, 108640. [Google Scholar] [CrossRef]
- Yu, Y.H.; Bian, L.; Wan, Y.T.; Jiao, Z.L.; Yu, K.K.; Zhang, G.H.; Guo, D.L. Grape (Vitis vinifera) VvDOF3 functions as a transcription activator and enhances powdery mildew resistance. Plant Physiol. Biochem. 2019, 143, 183–189. [Google Scholar] [CrossRef]
- Yu, Y.; Wan, Y.; Jiao, Z.; Bian, L.; Yu, K.; Zhang, G.; Guo, D. Functional Characterization of Resistance to Powdery Mildew of VvTIFY9 from Vitis vinifera. Int. J. Mol. Sci. 2019, 20, 4286. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.H.; Li, X.Z.; Wu, Z.J.; Chen, D.X.; Li, G.R.; Li, X.Q.; Zhang, G.H. VvZFP11, a Cys2His2-type zinc finger transcription factor, is involved in defense responses in Vitis vinifera. Biol. Plant. 2016, 60, 292–298. [Google Scholar] [CrossRef]
- Coutos-Thévenot, P.; Poinssot, B.; Bonomelli, A.; Yean, H.; Breda, C.; Buffard, D.; Esnault, R.; Hain, R.; Boulay, M. In vitro tolerance to Botrytis cinerea of grapevine 41B rootstock in transgenic plants expressing the stilbene synthase Vst1 gene under the control of a pathogen-inducible PR 10 promoter. J. Exp. Bot. 2001, 52, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Dabauza, M.; Velasco, L.; Pazos-Navarro, M.; Pérez-Benito, E.; Hellín, P.; Flores, P.; Gómez-Garay, A.; Martínez, M.C.; Lacasa, A. Enhanced resistance to Botrytis cinerea in genetically-modified Vitis vinifera L. plants over-expressing the grapevine stilbene synthase gene. Plant Cell Tissue Organ Cult. 2015, 120, 229–238. [Google Scholar] [CrossRef]
- Fan, C.; Pu, N.; Wang, X.; Wang, Y.; Fang, L.; Xu, W.; Zhang, J. Agrobacterium-mediated genetic transformation of grapevine (Vitis vinifera L.) with a novel stilbene synthase gene from Chinese wild Vitis pseudoreticulata. Plant Cell Tissue Organ Cult. 2008, 92, 197–206. [Google Scholar] [CrossRef]
- Aleynova-Shumakova, O.A.; Dubrovina, A.S.; Manyakhin, A.Y.; Karetin, Y.A.; Kiselev, K.V. VaCPK20 gene overexpression significantly increased resveratrol content and expression of stilbene synthase genes in cell cultures of Vitis amurensis Rupr. Appl. Microbiol. Biotechnol. 2014, 98, 5541–5549. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Zhou, Q.; Li, R.; Du, Y.; He, J.; Wang, D.; Cheng, S.; Zhang, J.; Wang, Y. Establishment of a picloram-induced somatic embryogenesis system in Vitis vinifera cv. chardonnay and genetic transformation of a stilbene synthase gene from wild-growing Vitis species. Plant Cell Tissue Organ Cult. 2015, 121, 397–412. [Google Scholar] [CrossRef]
- Jiao, Y.; Xu, W.; Duan, D.; Wang, Y.; Nick, P. A stilbene synthase allele from a Chinese wild grapevine confers resistance to powdery mildew by recruiting salicylic acid signalling for efficient defence. J. Exp. Bot. 2016, 67, 5841–5856. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, D.; Wang, F.; Huang, L.; Tian, X.; van Nocker, S.; Gao, H.; Wang, X. Expression of the Grape VaSTS19 Gene in Arabidopsis Improves Resistance to Powdery Mildew and Botrytis cinerea but Increases Susceptibility to Pseudomonas syringe pv Tomato DC3000. Int. J. Mol. Sci. 2017, 18, 2000. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Ma, F.; Wu, F.; Jiang, C.; Wang, Y. Expression of stilbene synthase VqSTS6 from wild Chinese Vitis quinquangularis in grapevine enhances resveratrol production and powdery mildew resistance. Planta 2019, 250, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yan, X.; Zhang, S.; Zhu, Y.; Zhang, X.; Qiao, H.; van Nocker, S.; Li, Z.; Wang, X. The jasmonate-ZIM domain gene VqJAZ4 from the Chinese wild grape Vitis quinquangularis improves resistance to powdery mildew in Arabidopsis thaliana. Plant Physiol. Biochem. 2019, 143, 329–339. [Google Scholar] [CrossRef]
- Agüero, C.B.; Uratsu, S.L.; Greve, C.; Powell, A.L.T.; Labavitch, J.M.; Meredith, C.P.; Dandekar, A.M. Evaluation of tolerance to Pierce’s disease and Botrytis in transgenic plants of Vitis vinifera L. expressing the pear PGIP gene. Mol. Plant Pathol. 2005, 6, 43–51. [Google Scholar] [CrossRef]
- Rubio, J.; Montes, C.; Castro, Á.; Álvarez, C.; Olmedo, B.; Muñoz, M.; Tapia, E.; Reyes, F.; Ortega, M.; Sánchez, E.; et al. Genetically engineered Thompson Seedless grapevine plants designed for fungal tolerance: Selection and characterization of the best performing individuals in a field trial. Transgenic Res. 2015, 24, 43–60. [Google Scholar] [CrossRef]
- Yu, Y.; Xu, W.; Wang, J.; Wang, L.; Yao, W.; Yang, Y.; Xu, Y.; Ma, F.; Du, Y.; Wang, Y. Erysiphe necator-induced RING finger protein 1 (EIRP1) ahe Chinese wild grapevine (Vitis pseudoreticulata) E3 ubiquitin ligase ctivates plant defense responses by inducing proteolysis of the VpWRKY11 transcription factor. New Phytol. 2013, 200, 834–846. [Google Scholar] [CrossRef]
- Wang, J.; Yao, W.; Wang, L.; Ma, F.; Tong, W.; Wang, C.; Bao, R.; Jiang, C.; Yang, Y.; Zhang, J.; et al. Overexpression of VpEIFP1, a novel F-box/Kelch-repeat protein from wild Chinese Vitis pseudoreticulata, confers higher tolerance to powdery mildew by inducing thioredoxin z proteolysis. Plant Sci. 2017, 263, 142–155. [Google Scholar] [CrossRef]
- Yu, Y.; Bian, L.; Jiao, Z.; Yu, K.; Wan, Y.; Zhang, G.; Guo, D. Molecular cloning and characterization of a grapevine (Vitis vinifera L.) serotonin N-acetyltransferase (VvSNAT2) gene involved in plant defense. BMC Genom. 2019, 20, 880. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, Y.; Hou, F.; Wan, D.; Cheng, Y.; Han, Y.; Gao, Y.; Liu, J.; Guo, Y.; Xiao, S.; et al. Ectopic expression of Arabidopsis broad-spectrum resistance gene RPW8.2 improves the resistance to powdery mildew in grapevine (Vitis vinifera). Plant Sci. 2018, 267, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Dai, L.; Cheng, S.; He, J.; Wang, D.; Zhang, J.; Wang, Y. A circulatory system useful both for long-term somatic embryogenesis and genetic transformation in Vitis vinifera L. cv. Thompson Seedless. Plant Cell Tissue Organ Cult. 2014, 118, 157–168. [Google Scholar] [CrossRef]
- Jiao, L.; Zhang, Y.; Lu, J. Overexpression of a stress-responsive U-box protein gene VaPUB affects the accumulation of resistance related proteins in Vitis vinifera ‘Thompson Seedless. Plant Physiol. Biochem. 2017, 112, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, C.; Chao, N.; Lu, J.; Zhang, Y. Cloning, characterization, and functional investigation of VaHAESA from Vitis amurensis inoculated with Plasmopara viticola. Int. J. Mol. Sci. 2018, 19, 1204. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Van Loon, L.C.; Van Kammen, A. Polyacrylamide disc electrophoresis of the soluble leaf proteins from Nicotiana tabacum var. “Samsun” and “Samsun NN”. II. Changes in protein constitution after infection with tobacco mosaic virus. Virology 1970, 40, 199–211. [Google Scholar] [CrossRef]
- Ali, S.; Mir, A.; Tyagi, A.; Bhat, J.A.; Chandrashekar, N.; Papolu, P.K.; Rawat, S.; Grover, A. Identification and comparative analysis of Brassica juncea pathogenesis-related genes in response to hormonal, biotic and abiotic stresses. Acta Physiol. Plant. 2017, 39, 268. [Google Scholar] [CrossRef]
- Antoniw, J.F.; Ritter, C.E.; Pierpoint, W.S.; Van Loon, L.C. Comparison of three pathogenesis-related proteins from plants of two cultivars of tobacco infected with TMV. J. Gen. Virol. 1980, 47, 79–87. [Google Scholar] [CrossRef]
- Van Loon, L.C. Regulation of Changes in Proteins and Enzymes Associated with Active Defence against Virus Infection. In Active Defense Mechanisms in Plants; Springer: Boston, MA, USA, 1982; pp. 247–273. [Google Scholar] [CrossRef]
- Métraux, J.P.; Streit, L.; Staub, T. A pathogenesis-related protein in cucumber is a chitinase. Physiol. Mol. Plant Pathol. 1988, 33, 1–9. [Google Scholar] [CrossRef]
- Melchers, L.S.; Apotheker-de-Groot, M.; van der Knaap, J.A.; Ponstein, A.S.; Sela-Buurlage, M.B.; Bol, J.F.; Cornelissen, B.J.C.; van den Elzen, P.J.M.; Linthorst, H.J.M. A new class of tobacco chitinases homologous to bacterial exo-chitinases displays antifungal activity. Plant J. 1994, 5, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Green, T.R.; Ryan, C.A. Wound-induced proteinase inhibitor in plant leaves: A possible defense mechanism against insects. Science 1972, 175, 776–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagrimini, L.M.; Burkhart, W.; Moyer, M.; Rothstein, S. Molecular cloning of complementary DNA encoding the lignin-forming peroxidase from tobacco: Molecular analysis and tissue-specific expression. Proc. Natl. Acad. Sci. USA 1987, 84, 7542–7546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somssich, I.E.; Schmelzer, E.; Bollmann, J.; Hahlbrock, K. Rapid activation by fungal elicitor of genes encoding “pathogenesis-related” proteins in cultured parsley cells. Proc. Natl. Acad. Sci. USA 1986, 83, 2427–2430. [Google Scholar] [CrossRef] [Green Version]
- Terras, F.R.; Eggermont, K.; Kovaleva, V.; Raikhel, N.V.; Osborn, R.W.; Kester, A.; Rees, S.B.; Torrekens, S.; Van Leuven, F.; Vanderleyden, J. Small cysteine-rich antifungal proteins from radish: Their role in host defense. Plant Cell 1995, 7, 573–588. [Google Scholar] [CrossRef] [Green Version]
- Epple, P.; Apel, K.; Bohlmann, H. An Arabidopsis thaliana thionin gene is inducible via a signal transduction pathway different from that for pathogenesis-related proteins. Plant Physiol. 1995, 109, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Casañal, A.; Zander, U.; Muñoz, C.; Dupeux, F.; Luque, I.; Angel Botella, M.; Schwab, W.; Valpuesta, V.; Marquez, J.A. The Strawberry Pathogenesis-related 10 (PR-10) Fra a Proteins Control Flavonoid Biosynthesis by Binding to Metabolic Intermediates. J. Biol. Chem. 2013, 288, 35322. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wei, J.; Zou, Y.; Xu, K.; Wang, Y.; Cui, L.; Xu, Y. Molecular Characteristics and Biochemical Functions of VpPR10s from Vitis pseudoreticulata Associated with Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2014, 15, 19162–19182. [Google Scholar] [CrossRef] [Green Version]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [Green Version]
- Hightower, R.; Baden, C.; Penzes, E.; Dunsmuir, P. The expression of cecropin peptide in transgenic tobacco does not confer resistance to Pseudomonas syringae pv tabaci. Plant Cell Rep. 1994, 13, 295–299. [Google Scholar] [CrossRef]
- Eulgem, T. Regulation of the Arabidopsis defense transcriptome. Trends Plant Sci. 2005, 10, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.B.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Marchive, C.; Léon, C.; Kappel, C.; Coutos-Thévenot, P.; Corio-Costet, M.F.; Delrot, S.; Lauvergeat, V. Over-Expression of VvWRKY1 in Grapevines Induces Expression of Jasmonic Acid Pathway-Related Genes and Confers Higher Tolerance to the Downy Mildew. PLoS ONE 2013, 8, e54185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Costenaro da Silva, D.; da Silveira Falavigna, V.; Fasoli, M.; Buffon, V.; Denardi Porto, D.; Joannis Pappas Jr, G.; Pezzotti, M.; Pasquali, G.; Fernando Revers, L. Transcriptome analyses of the Dof-like gene family in grapevine reveal its involvement in berry, flower and seed development. Hortic. Res. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.M.; Bae, H. An overview of stress-induced resveratrol synthesis in grapes: Perspectives for resveratrol-enriched grape products. Molecules 2017, 22, 294. [Google Scholar] [CrossRef]
- Adrian, M.; Jeandet, P. Effects of resveratrol on the ultrastructure of Botrytis cinerea conidia and biological significance in plant/pathogen interactions. Fitoterapia 2012, 83, 1345–1350. [Google Scholar] [CrossRef]
- Gray, D.J.; Li, Z.T.; Dhekney, S.A. Precision breeding of grapevine (Vitis vinifera L.) for improved traits. Plant Sci. 2014, 228, 3–10. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef]
- Xiao, S.; Ellwood, S.; Calis, O.; Patrick, E.; Li, T.; Coleman, M.; Turner, J.G. Broad-spectrum mildew resistance in Arabidopsis thaliana mediated by RPW8. Science 2001, 291, 118–120. [Google Scholar] [CrossRef]
- Bhat, A.; Ryu, C.-M. Plant Perceptions of Extracellular DNA and RNA. Mol. Plant 2016, 9, 956–958. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Wang, H.; Hu, P.; Hamby, R.; Jin, H. Small RNAs—Big Players in Plant-Microbe Interactions. Cell Host Microbe 2019, 26, 173–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Zhai, Y.; Feng, L.; Karimi, H.Z.; Rutter, B.D.; Zeng, L.; Choi, D.S.; Zhang, B.; Gu, W.; Chen, X.; et al. A Phytophthora Effector Suppresses Trans-Kingdom RNAi to Promote Disease Susceptibility. Cell Host Microbe 2019, 25, 153–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalakouras, A.; Wassenegger, M.; Dadami, E.; Ganopoulos, I.; Pappas, M.L.; Papadopoulou, K. Update on Exogenous Application of RNAi Genetically Modified Organism-Free RNA Interference: Exogenous Application of RNA Molecules in Plants 1[OPEN]. Plant Physiol. Ò 2020, 182, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.-M.; Palmquist, J.; Huang, S.-D.; Jin, H. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.H.J.; Da Huang, H.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef]
- Nowara, D.; Schweizer, P.; Gay, A.; Lacomme, C.; Shaw, J.; Ridout, C.; Douchkov, D.; Hensel, G.; Kumlehn, J. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell 2010, 22, 3130–3141. [Google Scholar] [CrossRef] [Green Version]
- Pliego, C.; Nowara, D.; Bonciani, G.; Gheorghe, D.M.; Xu, R.; Surana, P.; Whigham, E.; Nettleton, D.; Bogdanove, A.J.; Wise, R.P.; et al. Host-induced gene silencing in barley powdery mildew reveals a class of ribonuclease-like effectors. Mol. Plant-Microbe Interact. 2013, 26, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Whigham, E.; Qi, S.; Mistry, D.; Surana, P.; Xu, R.; Fuerst, G.; Pliego, C.; Bindschedler, L.V.; Spanu, P.D.; Dickerson, J.A.; et al. Broadly conserved fungal effector BEC1019 suppresses host cell death and enhances pathogen virulence in powdery mildew of barley (Hordeum vulgare L.). Mol. Plant-Microbe Interact. 2015, 28, 968–983. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.; Kumar, N.; Weber, L.; Keller, H.; Imani, J.; Kogel, K.H. Host-induced gene silencing of cytochrome P450 lanosterol C14α-demethylase-encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. USA 2013, 110, 19324–19329. [Google Scholar] [CrossRef] [Green Version]
- Ghag, S.B.; Shekhawat, U.K.S.; Ganapathi, T.R. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium wilt in banana. Plant Biotechnol. J. 2014, 12, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Song, X.-S.; Li, H.-P.; Cao, L.-H.; Sun, K.; Qiu, X.-L.; Xu, Y.-B.; Yang, P.; Huang, T.; Zhang, J.-B.; et al. Host-induced gene silencing of an essential chitin synthase gene confers durable resistance to Fusarium head blight and seedling blight in wheat. Plant Biotechnol. J. 2015, 13, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Panwar, V.; McCallum, B.; Bakkeren, G. Host-induced gene silencing of wheat leaf rust fungus Puccinia triticina pathogenicity genes mediated by the Barley stripe mosaic virus. Plant Mol. Biol. 2013, 81, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jin, Y.; Zhao, J.-H.; Gao, F.; Zhou, B.-J.; Fang, Y.-Y.; Guo, H.-S. Host-Induced Gene Silencing of the Target Gene in Fungal Cells Confers Effective Resistance to the Cotton Wilt Disease Pathogen Verticillium dahliae. Mol. Plant 2016, 9, 939–942. [Google Scholar] [CrossRef] [Green Version]
- Vega-Arreguín, J.C.; Jalloh, A.; Bos, J.I.; Moffett, P. Recognition of an Avr3a homologue plays a major role in mediating nonhost resistance to phytophthora capsici in Nicotiana species. Mol. Plant-Microbe Interact. 2014, 27, 770–780. [Google Scholar] [CrossRef] [Green Version]
- Sanju, S.; Siddappa, S.; Thakur, A.; Shukla, P.K.; Srivastava, N.; Pattanayak, D.; Sharma, S.; Singh, B.P. Host-mediated gene silencing of a single effector gene from the potato pathogen Phytophthora infestans imparts partial resistance to late blight disease. Funct. Integr. Genom. 2015, 15, 697–706. [Google Scholar] [CrossRef]
- Jahan, S.N.; Åsman, A.K.M.; Corcoran, P.; Fogelqvist, J.; Vetukuri, R.R.; Dixelius, C. Plant-mediated gene silencing restricts growth of the potato late blight pathogen Phytophthora infestans. J. Exp. Bot. 2015, 66, 2785–2794. [Google Scholar] [CrossRef] [Green Version]
- Govindarajulu, M.; Epstein, L.; Wroblewski, T.; Michelmore, R.W. Host-induced gene silencing inhibits the biotrophic pathogen causing downy mildew of lettuce. Plant Biotechnol. J. 2015, 13, 875–883. [Google Scholar] [CrossRef] [Green Version]
- Panstruga, R. Serpentine plant MLO proteins as entry portals for powdery mildew fungi. Biochem. Soc. Trans. 2005, 33, 389–392. [Google Scholar] [CrossRef]
- Pessina, S.; Lenzi, L.; Perazzolli, M.; Campa, M.; Dalla Costa, L.; Urso, S.; Valè, G.; Salamini, F.; Velasco, R.; Malnoy, M. Knockdown of MLO genes reduces susceptibility to powdery mildew in grapevine. Hortic. Res. 2016, 3, 16016. [Google Scholar] [CrossRef] [Green Version]
- Sabbadini, S.; Capriotti, L.; Molesini, B.; Pandolfini, T.; Navacchi, O.; Limera, C.; Ricci, A.; Mezzetti, B. Comparison of regeneration capacity and Agrobacterium-mediated cell transformation efficiency of different cultivars and rootstocks of Vitis spp. via organogenesis. Sci. Rep. 2019, 9, 582. [Google Scholar] [CrossRef] [PubMed]
- Dubrovina, A.; Aleynova, O.; Kalachev, A.; Suprun, A.; Ogneva, Z.; Kiselev, K. Induction of Transgene Suppression in Plants via External Application of Synthetic dsRNA. Int. J. Mol. Sci. 2019, 20, 1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, A.; Biedenkopf, D.; Furch, A.; Weber, L.; Rossbach, O.; Abdellatef, E.; Linicus, L.; Johannsmeier, J.; Goesmann, J.L.; Cardoza, V.; et al. An RNAi-based Control of Fusarium graminearum Infections Through Spraying of Long dsRNAs involves a plant passage and is controlled by the fungal silencing machinery. PLoS Pathog. 2016, 12, e1005901. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, A.G.; Wytinck, N.; Walker, P.L.; Girard, I.J.; Rashid, K.Y.; De Kievit, T.; Fernando, W.G.D.; Whyard, S.; Belmonte, M.F. Identification and application of exogenous dsRNA confers plant protection against Sclerotinia sclerotiorum and Botrytis cinerea. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Gu, K.X.; Song, X.S.; Xiao, X.M.; Duan, X.X.; Wang, J.X.; Duan, Y.B.; Hou, Y.P.; Zhou, M.G. A β 2 -tubulin dsRNA derived from Fusarium asiaticum confers plant resistance to multiple phytopathogens and reduces fungicide resistance. Pestic. Biochem. Physiol. 2019, 153, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Cagliari, D.; Avila dos Santos, E.; Dias, N.; Smagghe, G.; Zotti, M. Nontransformative Strategies for RNAi in Crop Protection. In Modulating Gene Expression—Abridging the RNAi and CRISPR-Cas9 Technologies [Working Title]; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Sang, H.; Kim, J. Il Advanced strategies to control plant pathogenic fungi by host-induced gene silencing (HIGS) and spray-induced gene silencing (SIGS). Plant Biotechnol. Rep. 2020, 14, 1–8. [Google Scholar] [CrossRef]
- Dalakouras, A.; Jarausch, W.; Buchholz, G.; Bassler, A.; Braun, M.; Manthey, T.; Krczal, G.; Wassenegger, M. Delivery of Hairpin RNAs and Small RNAs Into Woody and Herbaceous Plants by Trunk Injection and Petiole Absorption. Front. Plant Sci. 2018, 9, 1253. [Google Scholar] [CrossRef] [Green Version]
- Dubrovina, A.S.; Kiselev, K.V. Exogenous RNAs for Gene Regulation and Plant Resistance. Int. J. Mol. Sci. 2019, 20, 2282. [Google Scholar] [CrossRef] [Green Version]
- Das, P.R.; Sherif, S.M. Application of Exogenous dsRNAs-induced RNAi in Agriculture: Challenges and Triumphs. Front. Plant Sci. 2020, 11, 946. [Google Scholar] [CrossRef]
- Wang, M.; Dean, R.A. Movement of small RNAs in and between plants and fungi. Mol. Plant Pathol. 2020, 21, 589–601. [Google Scholar] [CrossRef]
- Song, X.S.; Gu, K.X.; Duan, X.X.; Xiao, X.M.; Hou, Y.P.; Duan, Y.B.; Wang, J.X.; Zhou, M.G. A myosin5 dsRNA that reduces the fungicide resistance and pathogenicity of Fusarium asiaticum. Pestic. Biochem. Physiol. 2018, 150, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dalakouras, A.; Wassenegger, M.; McMillan, J.N.; Cardoza, V.; Maegele, I.; Dadami, E.; Runne, M.; Krczal, G.; Wassenegger, M. Induction of silencing in plants by high-pressure spraying of In vitro-synthesized small RNAs. Front. Plant Sci. 2016, 7, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerva, L.; Sandrini, M.; Gambino, G.; Chitarra, W. Double-Stranded RNAs (dsRNAs) as a Sustainable Tool against Gray Mold (Botrytis cinerea) in Grapevine: Effectiveness of Different Application Methods in an Open-Air Environment. Biomolecules 2020, 10, 200. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.; Deikman, J.; Hendrix, B.; Iandolino, A. Barriers to Efficient Foliar Uptake of dsRNA and Molecular Barriers to dsRNA Activity in Plant Cells. Front. Plant Sci. 2020, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Demirci, Y.; Zhang, B.; Unver, T. CRISPR/Cas9: An RNA-guided highly precise synthetic tool for plant genome editing. J. Cell. Physiol. 2018, 233, 1844–1859. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tu, M.; Wang, D.; Liu, J.; Li, Y.; Li, Z.; Wang, Y.; Wang, X. CRISPR/Cas9-mediated efficient targeted mutagenesis in grape in the first generation. Plant Biotechnol. J. 2018, 16, 844–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, C.; Liu, X.; Zhang, Z.; Wang, Y.; Duan, W.; Li, S.; Liang, Z. CRISPR/Cas9-mediated efficient targeted mutagenesis in Chardonnay (Vitis vinifera L.). Sci. Rep. 2016, 6, 32289. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A Robust CRISPR/Cas9 System for Convenient, High-Efficiency Multiplex Genome Editing in Monocot and Dicot Plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef]
- Pan, C.; Ye, L.; Qin, L.; Liu, X.; He, Y.; Wang, J.; Chen, L.; Lu, G. CRISPR/Cas9-mediated efficient and heritable targeted mutagenesis in tomato plants in the first and later generations. Sci. Rep. 2016, 6, 24765. [Google Scholar] [CrossRef]
- Nakajima, I.; Azuma, A.; Onoue, N.; Moriguchi, T.; Yamamoto, T.; Toki, S.; Endo, M. CRISPR/Cas9-mediated targeted mutagenesis in grape. PLoS ONE 2017, 12, e0177966. [Google Scholar] [CrossRef] [Green Version]
- Ren, F.; Ren, C.; Zhang, Z.; Duan, W.; Lecourieux, D.; Li, S.; Liang, Z. Efficiency Optimization of CRISPR/Cas9-Mediated Targeted Mutagenesis in Grape. Front. Plant Sci. 2019, 10, 612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Zhu, Q.; Chen, Y.; Liu, Y.G. CRISPR/Cas9 Platforms for Genome Editing in Plants: Developments and Applications. Mol. Plant 2016, 9, 961–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osakabe, Y.; Liang, Z.; Ren, C.; Nishitani, C.; Osakabe, K.; Wada, M.; Komori, S.; Malnoy, M.; Velasco, R.; Poli, M.; et al. CRISPR–Cas9-mediated genome editing in apple and grapevine. Nat. Protoc. 2018, 13, 2844–2863. [Google Scholar] [CrossRef] [PubMed]
- Bertini, E.; Tornielli, G.B.; Pezzotti, M.; Zenoni, S. Regeneration of plants from embryogenic callus-derived protoplasts of Garganega and Sangiovese grapevine (Vitis vinifera L.) cultivars. Plant Cell Tissue Organ Cult. 2019, 138, 239–246. [Google Scholar] [CrossRef]
- Malnoy, M.; Viola, R.; Jung, M.-H.; Koo, O.-J.; Kim, S.; Kim, J.-S.; Velasco, R.; Nagamangala Kanchiswamy, C. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7, 1904. [Google Scholar] [CrossRef]
- Giacomelli, L.; Zeilmaker, T.; Dalla Costa, L.; Malnoy, M.; Rouppe van der Voort, J.; Moser, C. Generation of mildew-resistant grapevine clones via genome editing: Potentials and hurdles. In LXII SIGA Annual Congress: Plant Development and Crop Productivity for Sustainable Agriculture; SIGA: Verona, Italy, 2018; p. 904570. [Google Scholar] [CrossRef]
- Zimny, T.; Sowa, S.; Tyczewska, A.; Twardowski, T. Certain new plant breeding techniques and their marketability in the context of EU GMO legislation—Recent developments. N Biotechnol. 2019, 51, 49–56. [Google Scholar] [CrossRef]
- Dalakouras, A.; Wassenegger, M.; Dadami, E.; Ganopoulos, I.; Pappas, M.; Papadopoulou, K.K. GMO-free RNAi: Exogenous application of RNA molecules in plants. Plant Physiol. 2019, 00570. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Fresnedo-Ramírez, J.; Wang, M.; Cote, L.; Schweitzer, P.; Barba, P.; Takacs, E.M.; Clark, M.; Luby, J.; Manns, D.C.; et al. A next-generation marker genotyping platform (AmpSeq) in heterozygous crops: A case study for marker-assisted selection in grapevine. Hortic. Res. 2016, 3, 16002. [Google Scholar] [CrossRef] [Green Version]
- Myles, S. Improving fruit and wine: What does genomics have to offer? Trends Genet. 2013, 29, 190–196. [Google Scholar] [CrossRef]
- Riaz, S.; Tenscher, A.C.; Graziani, R.; Krivanck, A.F.; Ramming, D.W.; Andrew Walkcrl, M.; Kahica, K.; Dervishian, G.; Ng, D. Using Marker-Assisted Selection to Breed Pierce’s Disease-Resistant Grapes. Am. J. Enol. Vitic. 2009, 60, 199–207. [Google Scholar]
- Hartung, F.; Schiemann, J. Precise plant breeding using new genome editing techniques: Opportunities, safety and regulation in the EU. Plant J. 2014, 78, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Schouten, H.J.; Krens, F.A.; Jacobsen, E. Cisgenic plants are similar to traditionally bred plants. EMBO Rep. 2006, 7, 750–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rommens, C.M.; Haring, M.A.; Swords, K.; Davies, H.V.; Belknap, W.R. The intragenic approach as a new extension to traditional plant breeding. Trends Plant Sci. 2007, 12, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.G.; Schaart, J.G.; Groenwold, R.; Jacobsen, E.; Schouten, H.J.; Krens, F.A. Functional analysis and expression profiling of HcrVf1 and HcrVf2 for development of scab resistant cisgenic and intragenic apples. Plant Mol. Biol. 2011, 75, 579–591. [Google Scholar] [CrossRef] [Green Version]
- Zotti, M.; dos Santos, E.A.; Cagliari, D.; Christiaens, O.; Taning, C.N.T.; Smagghe, G. RNA interference technology in crop protection against arthropod pests, pathogens and nematodes. Pest Manag. Sci. 2018, 74, 1239–1250. [Google Scholar] [CrossRef]
- Zischewski, J.; Fischer, R.; Bortesi, L. Detection of on-target and off-target mutations generated by CRISPR/Cas9 and other sequence-specific nucleases. Biotechnol. Adv. 2017, 35, 95–104. [Google Scholar] [CrossRef]
- Lusk, J.L.; Rozan, A. Consumer acceptance of ingenic foods. Biotechnol. J. 2006, 1, 1433–1434. [Google Scholar] [CrossRef]
- Frewer, L.J. Consumer acceptance and rejection of emerging agrifood technologies and their applications. Eur. Rev. Agric. Econ. 2017, 44, 683–704. [Google Scholar] [CrossRef]
- Delwaide, A.C.; Nalley, L.L.; Dixon, B.L.; Danforth, D.M.; Nayga, R.M.; Van Loo, E.J.; Verbeke, W. Revisiting GMOs: Are there differences in European consumers’ acceptance and valuation for cisgenically vs transgenically bred rice? PLoS ONE 2015, 10, e0126060. [Google Scholar] [CrossRef] [Green Version]
- Lucht, J. Public Acceptance of Plant Biotechnology and GM Crops. Viruses 2015, 7, 4254–4281. [Google Scholar] [CrossRef]
- Malyska, A.; Bolla, R.; Twardowski, T. The Role of Public Opinion in Shaping Trajectories of Agricultural Biotechnology. Trends Biotechnol. 2016, 34, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Telem, R.; Wani, S.; Singh, N.; Nandini, R.; Sadhukhan, R.; Bhattacharya, S.; Mandal, N. Cisgenics—A Sustainable Approach for Crop Improvement. Curr. Genom. 2013, 14, 468–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, E.; Schouten, H.J. Cisgenesis, a New Tool for Traditional Plant Breeding, Should be Exempted from the Regulation on Genetically Modified Organisms in a Step by Step Approach. Potato Res. 2008, 51, 75–88. [Google Scholar] [CrossRef]
- Arpaia, S.; Christiaens, O.; Giddings, K.; Jones, H.; Mezzetti, B.; Moronta-Barrios, F.; Perry, J.N.; Sweet, J.B.; Taning, C.N.T.; Smagghe, G.; et al. Biosafety of GM Crop Plants Expressing dsRNA: Data Requirements and EU Regulatory Considerations. Front. Plant Sci. 2020, 11, 940. [Google Scholar] [CrossRef]
- Casacuberta, J.M.; Devos, Y.; du Jardin, P.; Ramon, M.; Vaucheret, H.; Nogué, F. Biotechnological uses of RNAi in plants: Risk assessment considerations. Trends Biotechnol. 2015, 33, 145–147. [Google Scholar] [CrossRef] [Green Version]
- Mezzetti, B.; Smagghe, G.; Arpaia, S.; Christiaens, O.; Dietz-Pfeilstetter, A.; Jones, H.; Kostov, K.; Sabbadini, S.; Opsahl-Sorteberg, H.-G.; Ventura, V.; et al. RNAi: What is its position in agriculture? J. Pest Sci. 2020, 1, 3. [Google Scholar] [CrossRef]
- Papadopoulou, N.; Devos, Y.; Álvarez-Alfageme, F.; Lanzoni, A.; Waigmann, E. Risk Assessment Considerations for Genetically Modified RNAi Plants: EFSA’s Activities and Perspective. Front. Plant Sci. 2020, 11, 445. [Google Scholar] [CrossRef] [Green Version]
- Shew, A.M.; Danforth, D.M.; Nalley, L.L.; Nayga, R.M.; Tsiboe, F.; Dixon, B.L. New innovations in agricultural biotech: Consumer acceptance of topical RNAi in rice production. Food Control 2017, 81, 189–195. [Google Scholar] [CrossRef]
- El-Mounadi, K.; Morales-Floriano, M.L.; Garcia-Ruiz, H. Principles, Applications, and Biosafety of Plant Genome Editing Using CRISPR-Cas9. Front. Plant Sci. 2020, 11, 56. [Google Scholar] [CrossRef]
- Ishii, T.; Araki, M. Consumer acceptance of food crops developed by genome editing. Plant Cell Rep. 2016, 35, 1507–1518. [Google Scholar] [CrossRef]
- Voytas, D.F.; Gao, C. Precision Genome Engineering and Agriculture: Opportunities and Regulatory Challenges. PLoS Biol. 2014, 12, e1001877. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene Name | Gene Source | Description | Biological Effect | Host | References |
|---|---|---|---|---|---|
| Pathogenesis-Related Proteins | |||||
| Non-expressor of Pathogenesis Related 1 (VvNPR1.1) | Vitis vinifera | Key signal in salicylic acid pathway and local basal resistance to biotrophs | Enhanced resistance to Erysiphe necator | Vitis vinifera cv. Chardonnay | [30] |
| Rice chitinase (RCC2) | Oryza sativa | PR protein Class I Chitinase | Major resistance to Erysiphe necator and slight resistance to Elsinoe ampelina | Vitis vinifera cv. Neo Muscat | [31] |
| Chitinase and ribosome-inactivating protein (RIP) | Hordeum vulgare | These genes encode for two antifungal proteins | Susceptibility to Erysiphe necator and Plasmopara viticola infection equal to that of the control | Vitis vinifera cv. Seyval blanc | [32] |
| Rice Chitinase (Chi 11) | Oryza sativa | Pathogenesis-related protein | Late and reduced manifestation of Erysiphe necator symptoms | Vitis vinifera cv. Pusa Seedless | [33] |
| Chitinase and β-1,3-glucanase | Scab-infected Sumai 3 wheat | Pathogenesis-related proteins | Chitinase was more effective than glucanase in conferring tolerance to Plasmopara viticola | Vitis vinifera cv. Crimson Seedless | [34] |
| VpPR4-1 | Vitis pseudoreticulata | PR4 proteins are considered chitin-binding proteins | Improved tolerance to Erysiphe necator | Vitis vinifera cv. Red Globe | [35] |
| Thaumatin-like protein (Vvtl-1) | Vitis vinifera cv. Chardonnay | Pathogenesis-related protein 5 | Increased resistance to Erysiphe necator and Elsinoe ampelina | Vitis vinifera cv. Thompson Seedless | [36] |
| Thaumatin-like protein (VqTLP29) | Vitis quinquangularis cv. Shang-24 | Pathogenesis-related protein 5 | Increased resistance to powdery mildew, but decreased resistance to Botrytis cinerea | Arabidopsis thaliana | [37] |
| Thaumatin-like protein (VaTLP) | Vitis amurensis Rupr. “Zuoshan-1” | PR5 proteins have endo-β-1,3-glucanase activity; binding β-1,3-glucan | Reinforced resistance to Plasmopara viticola | Vitis vinifera cv. Thompson Seedless | [38] |
| VpPR10.1 | Vitis pseudoreticulata | PR10 have in vitro ribonuclease activity | Increased tolerance to Plasmopara viticola | Vitis vinifera cv. Thompson Seedless | [39] |
| Antimicrobial Peptides (AMPs) | |||||
| Natural Magainin-2 (Mag2)/synthetic derivate (MS199) | Magainin extracts from the skin of Xenopus laevis frog | Magainins with broad-spectrum in vitro antimicrobial activity against bacteria and fungi | Resistance to bacterial diseases such as crown gall diseases, minor susceptibility against Erysiphe necator | Vitis vinifera cv. Chardonnay | [40] |
| Magainin-2 (mag2) +PGL | Magainin extracts from the skin of Xenopus laevis frog | AMP belonging to the Magainins family | PGL protein seems to inhibit Botrytis cinerea spore germination | Vitis vinifera cv. Chardonnay | [41] |
| Transcription Factors | |||||
| VvWRKY2 | Vitis vinifera cv. Cabernet Sauvignon | WRKY protein isolated after Plasmopara viticola infection | Increased tolerance to Botrytis cinerea and broad-spectrum fungal resistance | Nicotiana tabacum cv. Xanthi | [42] |
| VvWRKY33 | Vitis vinifera | WRKY protein | Enhanced resistance to Plasmopara viticola | Vitis vinifera cv. Shiraz | [43] |
| VpWRKY3 | Vitis pseudoreticulata accession “Baihe-35-1” | WRKY protein isolated after Erysiphe necator infection | Improved tolerance to Ralstonia solanacearum | Nicotiana tabacum cv. NC89 | [44] |
| Ethylene response factors (VpERF2 and VpERF3) | Vitis pseudoreticulata | Transcription factor isolated after Erysiphe necator infection | Enhanced resistance to Ralstonia solanacearum and Phytophtora parasitica var. nicotianae Tucker | Nicotiana tabacum cv. NC89 | [45] |
| C-repeat-binding factor dehydration-responsive element-binding factor 1C (MrCBF2/DREB1C) | Muscadinia rotundifolia “Noble” | Transcription factor isolated after Plasmopara viticola inoculation | Enhanced resistance to Peronospora parasitica | Arabidopsis thaliana “COL0” | [46] |
| bZIP transcription factor (VvbZIP60) | Vitis vinifera cv. Jing Xiu | Transcription factor that activates the accumulation of salicylic acid and the expression of PR1 protein | Enhanced resistance to powdery mildew | Arabidopsis thaliana | [47] |
| DOF protein (VvDOF3) | Vitis vinifera | Protein involved in plant growth, development, and plant defense | Enhanced resistance to powdery mildew | Arabidopsis thaliana | [48] |
| Tify protein (VvTIFY9) | Vitis vinifera | Protein highly expressed in leaves. Play an active role in SA pathway | Increased resistance to powdery mildew | Arabidopsis thaliana | [49] |
| C2H2-type zinc finger protein (VvZFP11) | Vitis vinifera | This protein expression is regulated by salicylic acid and methyl jasmonate | Enhanced resistance to powdery mildew | Arabidopsis thaliana | [50] |
| Secondary stress-related metabolites | |||||
| PR10 promoter- Stilbene synthase (Vst1) | Vitis vinifera cv. Optima | Stilbenes production | Decreased susceptibility to Botrytis cinerea | 41B rootstock (Vitis vinifera cv. Chasselas x Vitis berlandieri) | [51] |
| Stilbene synthase (Vst1) | Vitis vinifera | Stilbenes production | Reinforced resistant against Botrytis cinerea | Vitis vinifera cv. Sugraone | [52] |
| Stilbene synthase (STS) | Vitis pseudoreticulata | Stilbenes synthesis | Transgenic plants with high resveratrol content | Vitis vinifera cv. Thompson Seedless | [53] |
| Calcium-dependent protein kinase (CDPK) (VaCPK20) | Vitis amurensis | Regulator of the biosynthetic pathways of resveratrol | Increased expression of STS7 gene, enhanced resveratrol production | Cell cultures of Vitis amurensis rupr. | [54] |
| Stilbene synthase (VpSTSgDNA2) | Vitis pseudoreticulata | Stilbenes production | Improved tolerance against Erysiphe necator | Vitis vinifera cv. Chardonnay | [55] |
| Stilbene synthase (VpSTS) | Vitis pseudoreticulata | Stilbenes production | Improved resistance to powdery mildew | Arabidopsis thaliana | [56] |
| Stilbene synthase (VaSTS19) | Vitis amurensis | Stilbenes production | Improved resistance to Botrytis cinerea and powdery mildew | Arabidopsis thaliana | [57] |
| Stilbene synthase (VqSTS6) | Vitis quinquangularis | Stilbenoids accumulation | Improved resistance to Erysiphe necator | Vitis vinifera cv. Thompson Seedless | [58] |
| Defense-related genes | |||||
| Jasmonate-ZIM domain protein (VqJAZ4) | Vitis quinquangularis clone Shang-24 | This gene is upregulated after Erisiphe necator inoculation | Improved resistance to powdery mildew and enhanced susceptibility to Botrytis cinerea. | Arabidopsis thaliana | [59] |
| Polygalacturonase-inhibiting proteins (pPGIPs) | Pear fruit | PGIPs are plant cell wall proteins that specifically inhibit fungal endo-polygalacturonases (PGs). | Increased resistance to Botrytis cinerea and slight tolerance to Xylella fastidiosa | Vitis vinifera cv. Thompson Seedless and Chardonnay | [60] |
| Two endochitinases (ech42 and ech33) and one N-acetyl-β-d-hexosaminidase (nag70) | Trichoderma harzianum, Trichoderma virens | Extracellular endochitinases of biocontrol agents and chitinolytic genes | Enhanced resistance to Botrytis cinerea. Tolerance to Erysiphe necator in ech42-nag70 expressing transgenic plants | Vitis vinifera cv. Thompson Seedless | [61] |
| E3 ubiquitin ligase Erysiphe necator-induced RING finger protein 1 (VpEIRP1) | Vitis pseudoreticulata Baihe 31-1 accession | This protein activates plant defense response through the proteolysis of VpWRKY11 transcription factor | Enhanced resistance to powdery mildew | Arabidopsis thaliana | [62] |
| F-box/Kelch-repeat protein (VpEIFP1) | Vitis pseudoreticulata | Transcription of EIFP protein is induced after powdery mildew infection and activation of PR genes | Enhanced tolerance to Erysiphe necator | Vitis vinifera cv. Red Globe and Arabidopsis thaliana | [63] |
| Metacaspases (VrMC2 andVrMC5) | Vitis rupestris | Executors of hypersensitive response (HR), isolated after Plasmopara viticola infection | Programmed cell death (PCD) activation | Nicotiana tabacum cv. Bright Yellow 2 and Vitis vinifera cell cultures | [29] |
| Serotonin N-acetyltransferase (VvSNAT2) | Vitis vinifera | Protein essential for melatonin production and for SA and JA signaling pathways activation | Improved resistance to powdery mildew | Arabidopsis thaliana | [64] |
| Resistance to Powdery Mildew 8 locus (RPW8.2) | Arabidopsis thaliana | Protein that encodes for small basic protein, with weak homology with NB-LRR protein | Erysiphe necator hyphal growth and sporulation were significantly restricted | Vitis vinifera cv. Thompson Seedless | [65] |
| Ubiquitin ligase (VpPUB23) | Vitis pseudoreticulata | Type E3 ubiquitin ligase is involved in many immune regulation responses | Decreased resistance to Erysiphe necator | Vitis vinifera cv. Thompson Seedless | [66] |
| Ubiquitin ligase (VaPUB) | Vitis amurensis | U-box protein E3 ligase causes downregulation of PR10 | Transgenic plants were susceptible as control to Plasmopara viticola | Vitis vinifera cv. Thompson Seedless | [67] |
| VaHAESA | Vitis amurensis cv. Shuanghong | Pattern recognition receptor (PRR) that belongs to leucine-rich repeat receptor-like protein kinase | Induce H2O2, NO, and callose accumulation. Leaves showed less spores and Plasmopara viticola infected areas than control | Vitis vinifera cv. Thompson Seedless | [68] |
| Technology | Type of Modification | Target Origin and Description | Time Needed | Classification According to EU-Legislation | Side Effects | Biosafety Concerns | Consumer Acceptance (Proposed) |
|---|---|---|---|---|---|---|---|
| Traditional breeding | Breeding and several backcrosses generation, introgression breeding, induced mutagenesis, and somatic hybridization | Genes found in crossable, sexually compatible organisms | At least 10-15 years | Non-GMO | Altered clone identity, partial resistance to biotic stresses | No biosafety concerns and basic regulation needed [149] | High [153] |
| Transgenesis | Genetic transformation | Overexpression of genes also from non-sexually compatible organisms, presence of gene sequences (i.e., promoter, selectable marker gene) from non-compatible organisms | Around 1 or 2 years | GMO | Release in the environment of genes of different origins; expression of new protein products with possible allergen/toxic effects | Expression of unknown protein/enzyme; use of antibiotic/herbicide resistance markers, lack of coexistence with non-GM, organic cultivations | Low [147,154,155,156,157] |
| Cisgenesis/Intragenesis | Genetic transformation | Expression/overexpression of a gene originating from the recipient plant itself (cisgenesis), or expression of full/partial coding sequence originating from a sexually compatible plant (intragenesis) | Around 1 or 2 years | GMO | Scarce availability of efficient cisgenic selectable marker genes | cisgenic/intragenic plants solve the current biosafety problems regarding the presence of foreign genes in the plant host genome [158] | Medium/High [148,149,159] |
| Gene silencing-HIGS | Genetic transformation | Overexpression of non-coding dsRNAs downregulating exogenous or endogenous gene expression | Around 1 or 2 years | GMO | Efficacy of gene silencing varies with the genes and target organisms. Possible off-target effects in non-target organisms (NTOs) | Reduced off-target effects by designing RNAi sequences with high specificity and verified with bioinformatic studies. Minimal biosafety concerns when HIGS is applied only to rootstocks by trans-grafting technique [160] | Medium [161,162,163] |
| Gene silencing-SIGS | No genetic modification | External application of non-coding dsRNAs downregulating exogenous or endogenous gene expression | Few months | Non-GMO | Efficacy of gene silencing depends on the efficiency and specificity of the RNAi sequence and on the degree of adsorption showed by plants and pathogens cells | RNAi sequence should be selected in order to avoid off-target effects [143]. The absence of negative effects, that can be caused by the nanotechnology-based delivery method used, on the environment and human health needs to be demonstrated | Medium/High [21,164] |
| Genome editing | Genetic transformation/plasmid-free protoplast transformation | Artificially gene modification/target random mutation | About 1 or 2 years. More time necessary if transgene segregation is required from T0 plants, or if protoplast in vitro regeneration is required | Non-GMO/GMO in Europe (ECJ-2018) | Possible appearance of off-target mutations; difficulties in plant regeneration from protoplasts | Transgene integration, effect of the expression of Cas9 protein, specificity and fidelity of Cas9 protein [165] | Medium/High [166,167] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capriotti, L.; Baraldi, E.; Mezzetti, B.; Limera, C.; Sabbadini, S. Biotechnological Approaches: Gene Overexpression, Gene Silencing, and Genome Editing to Control Fungal and Oomycete Diseases in Grapevine. Int. J. Mol. Sci. 2020, 21, 5701. https://doi.org/10.3390/ijms21165701
Capriotti L, Baraldi E, Mezzetti B, Limera C, Sabbadini S. Biotechnological Approaches: Gene Overexpression, Gene Silencing, and Genome Editing to Control Fungal and Oomycete Diseases in Grapevine. International Journal of Molecular Sciences. 2020; 21(16):5701. https://doi.org/10.3390/ijms21165701
Chicago/Turabian StyleCapriotti, Luca, Elena Baraldi, Bruno Mezzetti, Cecilia Limera, and Silvia Sabbadini. 2020. "Biotechnological Approaches: Gene Overexpression, Gene Silencing, and Genome Editing to Control Fungal and Oomycete Diseases in Grapevine" International Journal of Molecular Sciences 21, no. 16: 5701. https://doi.org/10.3390/ijms21165701