Phosphodiesterase Inhibitors: Could They Be Beneficial for the Treatment of COVID-19?

 , , and
, , and 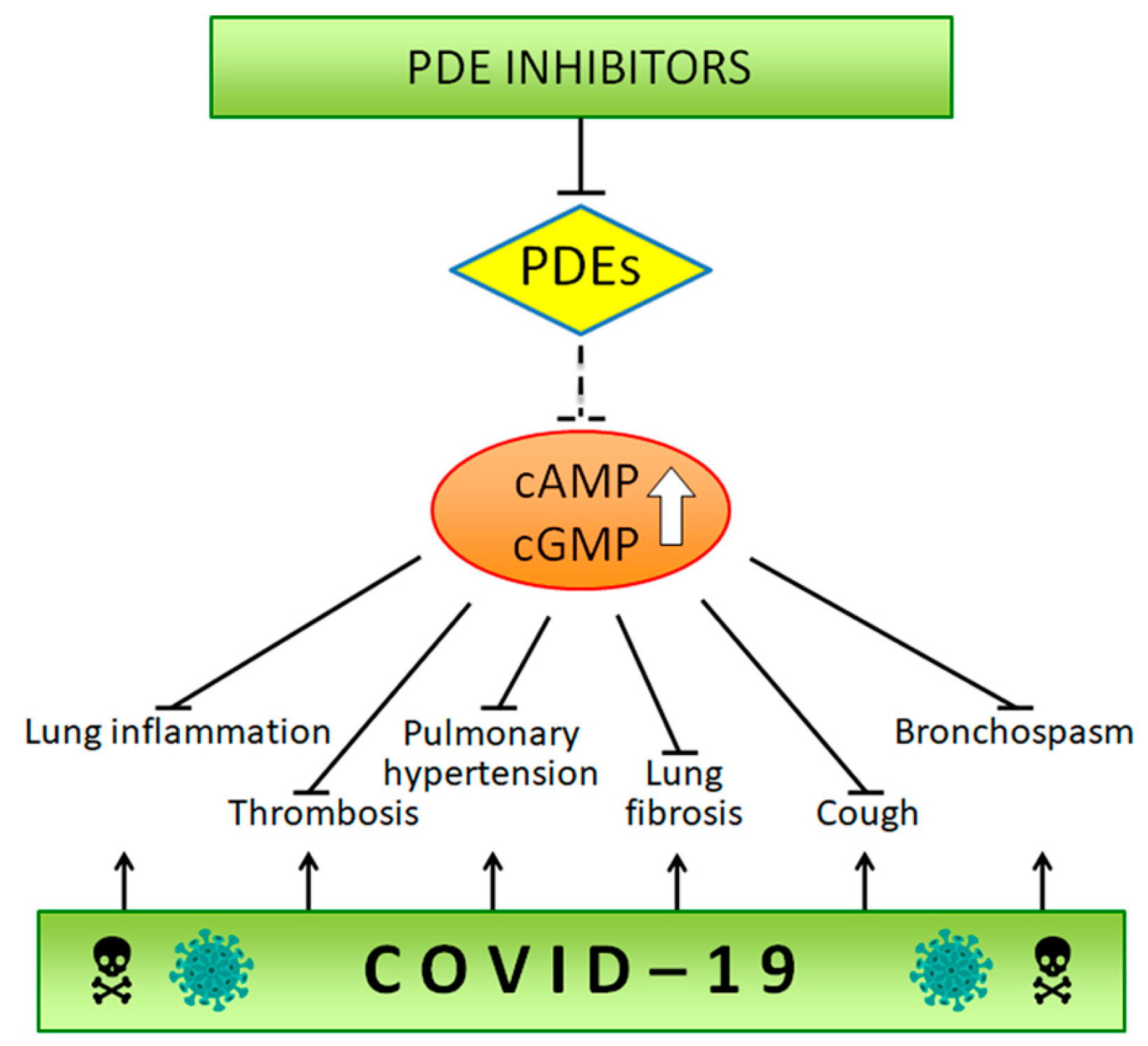{kind=link}
Abstract
:1. Introduction
2. Cyclic Nucleotide Pathway in Inflammation and Fibrosis
3. Cyclic Nucleotide Pathway in Vascular Resistance, Thrombosis and Stroke
4. Phosphodiesterase Inhibitors in COVID-19
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cyranoski, D. Mystery deepens over animal source of Coronavirus. Nature 2020, 579, 18–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Tai, W.; He, L.; Zhang, X.; Pu, J.; Voronin, D.; Jiang, S.; Zhou, Y.; Du, L. Characterization of the receptor-binding domain (RBD) of 2019 novel Coronavirus: Implication for development of RBD protein as a viral attachment inhibitor and vaccine. Cell Mol. Immunol. 2020, 17, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Baden, L.R.; Rubin, E.J. COVID-19: The search for effective therapy. N. Engl. J. Med. 2020, 382, 1851–1852. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wang, B.; Mao, J. The pathogenesis and treatment of the ‘Cytokine Storm’ in COVID-19. J. Infect 2020, 80, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, J.; Yang, Y.; Ma, H.; Li, Z.; Zhang, J.; Cheng, J.; Zhang, Y.Z.; Zhao, Y.; Xia, Z.; et al. The potential role of IL-6 in monitoring severe case of Coronavirus disease 2019. medRxiv 2020, preprint. [Google Scholar] [CrossRef]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; MacAry, P.A.; Ng, L.F.P. The trinity of COVID-19: Immunity, inflammation and intervention. Nat. Rev. Immun. 2020, 20, 363–374. [Google Scholar] [CrossRef]
- Tabas, I.; Glass, C.K. Anti-inflammatory therapy in chronic disease: Challenges and opportunities. Science 2013, 339, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Siebert, S.; Tsoukas, A.; Robertson, J.; Mcinnes, I. Cytokines as therapeutic targets in rheumatoid arthritis and other inflammatory diseases. Pharmacol. Rev. 2015, 67, 280–309. [Google Scholar] [CrossRef] [PubMed]
- Kazatchkine, M.D.; Kaveri, S.V. Immunomodulation of autoimmune and inflammatory diseases with intravenous immune globulin. N. Engl. J. Med. 2001, 345, 747–755. [Google Scholar] [CrossRef]
- Uguccioni, M.; Teixeira, M.M.; Locati, M.; Mantovani, A. Editorial: Regulation of inflammation, its resolution and therapeutic targeting. Front. Immunol. 2017, 8, 415. [Google Scholar] [CrossRef] [Green Version]
- Oxley, T.J.; Mocco, J.; Majidi, S.; Keller, C.P.; Shoirah, H.; Singh, I.P.; De Leacy, R.A.; Shigematsu, T.; Ladner, T.V.; Yaeger, K.A.; et al. Large-vessel stroke as a presenting feature of COVID-19 in the young. N. Engl. J. Med. 2020, 382, e60. [Google Scholar] [CrossRef]
- Klok, F.A.; Kruip, M.J.H.A.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.A.M.P.J.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Incidence of thrombotic complications in critically ill ICU patients with COVID-19. Thromb. Res. 2020, 191, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Klok, F.A.; Kruip, M.J.H.A.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.; Kant, K.M.; Kaptein, F.H.J.; van Paassens, J.; Stals, M.A.M.; Huisman, M.V.; et al. Confirmation of the high cumulative incidence of thrombotic complications in critically ill ICU patients with COVID-19: An updated analysis. Thromb. Res. 2020, 191, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, X.; Cai, Y.; Xia, J.; Zhou, X.; Xu, S.; Huang, H.; Zhang, L.; Zhou, X.; Du, C.; et al. Risk factors associated with acute respiratory distress syndrome and death in patients with Coronavirus disease 2019 pneumonia in Wuhan, China. JAMA Intern. Med. 2020. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, J.; Liu, H.; Han, N.; Ju, J.; Kou, Y.; Chen, L.; Jiang, M.; Pan, F.; Zheng, Y.; et al. Long-term bone and lung consequences associated with hospital-acquired severe acute respiratory syndrome: A 15-year follow-up from a prospective cohort study. Bone Res. 2020, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.M.; Lee, E.Y.; Singh, R.; Enani, M.A.; Al Dossari, K.; van Gorkom, K.; Larsson, S.G.; Langer, R.D. Follow-up chest radiographic findings in patients with MERS-CoV after recovery. Indian J. Radiol. Imaging 2017, 27, 342–349. [Google Scholar] [CrossRef]
- George, P.M.; Wells, A.U.; Jenkins, R.G. Pulmonary Fibrosis and COVID-19: The Potential Role for Antifibrotic Therapy. Lancet Respir. Med. 2020, 15, S2213–S2600. [Google Scholar] [CrossRef]
- Spagnolo, P.; Balestro, E.; Aliberti, S.; Cocconcelli, E.; Biondini, D.; Casa, G.D.; Sverzellati, N.; Maher, T.M. Pulmonary Fibrosis Secondary to COVID-19: A Call to Arms? Lancet Respir. Med. 2020. [Google Scholar] [CrossRef]
- Lugnier, C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: A new target for the development of specific therapeutic agents. Pharmacol. Ther. 2006, 109, 366–398. [Google Scholar] [CrossRef]
- Bender, A.T.; Beavo, J.A. Cyclic nucleotide phosphodiesterases: Molecular regulation to clinical use. Pharmacol. Rev. 2006, 58, 488–520. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Goldminz, A.M.; Kim, N.; Gottlieb, A.B. Phosphodiesterase 4-targeted treatments for autoimmune diseases. BMC Med. 2013, 11, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, S.H.; Blount, M.A.; Corbin, J.D. Mammalian cyclic nucleotide phosphodiesterases: Molecular mechanisms and physiological functions. Physiol. Rev. 2011, 91, 651–690. [Google Scholar] [CrossRef] [Green Version]
- Massimi, M.; Cardarelli, S.; Galli, F.; Giardi, M.F.; Panera, N.; Cinque, B.; Cifone, M.G.; Biagioni, S.; Giorgi, M. Increase of intracellular cyclic AMP by PDE4 inhibitors affects HepG2 cell cycle progression and survival. J. Cell Biochem. 2017, 118, 1401–1411. [Google Scholar] [CrossRef]
- Massimi, M.; Ragusa, F.; Cardarelli, S.; Giorgi, M. Targeting cyclic AMP signalling in hepatocellular carcinoma. Cells 2019, 8, 1511. [Google Scholar] [CrossRef] [Green Version]
- Chiricozzi, A.; Caposiena, D.; Garofalo, V.; Cannizzaro, M.V.; Chimenti, S.; Saraceno, R. A new therapeutic for the treatment of moderate-tosevere plaque psoriasis: Apremilast. Expert Rev. Clin. Immunol. 2016, 12, 237–249. [Google Scholar] [CrossRef]
- Maurice, D.H.; Ke, H.; Ahmad, F.; Wang, Y.; Chung, J.; Manganiello, V.C. Advances in targeting cyclic 710 nucleotide phosphodiesterases. Nat. Rev. Drug Discov. 2014, 13, 290–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.S.; Wynn, T.A. Pulmonary fibrosis: Pathogenesis, etiology and regulation. Mucosal Immunol. 2009, 2, 103–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambers, C.; Boehm, P.M.; Karabacak, Y.; Samaha, E.; Benazzo, A.; Jaksch, P.; Roth, M. Combined activation of guanylate cyclase and cyclic AMP in lung fibroblasts as a novel therapeutic concept for lung fibrosis. Biomed. Res. Int. 2019, 2019, 10. [Google Scholar] [CrossRef]
- Li, H.; Zuo, J.; Tang, W. Phosphodiesterase-4 inhibitors for the treatment of inflammatory fiseases. Front. Pharmacol. 2018, 9, 1048. [Google Scholar] [CrossRef] [Green Version]
- Insel, P.A.; Murray, F.; Yokoyama, U.; Romano, S.; Yun, H.; Brown, L.; Snead, A.; Lu, D.; Aroonsakool, N. cAMP and epac in the regulation of tissue fibrosis. Br. J. Pharmacol. 2012, 166, 447–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billington, C.K.; Oluwaseun, O.; Raymond, B.P.; Satoru, I. cAMP regulation of airway smooth muscle function. Pulm. Pharmacol. Ther. 2013, 26, 112–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójcik-Pszczoła, K.; Chłoń-Rzepa, G.; Jankowska, A.; Ślusarczyk, M.; Ferdek, P.E.; Kusiak, A.A.; Świerczek, A.; Pociecha, K.; Koczurkiewicz-Adamczyk, P.; Wyska, E.; et al. A novel, Pan-PDE inhibitor exerts anti-fibrotic effects in human lung fibroblasts via inhibition of TGF-β signaling and activation of cAMP/PKA signaling. Int. J. Mol. Sci. 2020, 21, 4008. [Google Scholar] [CrossRef] [PubMed]
- Page, C. Phosphodiesterase inhibitors for the treatment of asthma and chronic obstructive pulmonary disease. Int. Arch. Allergy Immunol. 2014, 165, 152–164. [Google Scholar] [CrossRef]
- Zuo, H.; Faiz, A.; van den Berge, M.; Mudiyanselage, S.N.H.R.; Borghuis, T.; Timens, W.; Nikolaev, V.O.; Burgess, J.K.; Schmidt, M. Cigarette smoke exposure alters phosphodiesterases in human structural lung cells. Am. J. Physiol. Cell. Mol. Physiol. 2019, 318, L59–L64. [Google Scholar] [CrossRef]
- Barnes, P.J. Theophylline. Am. J. Respir. Crit. Care Med. 2013, 188, 901–906. [Google Scholar] [CrossRef]
- Oñatibia-Astibia, A.; Martínez-Pinilla, E.; Franco, R. The potential of methylxanthine-based therapies in pediatric respiratory tract diseases. Respir. Med. 2016, 112, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.E. Inhaled phosphodiesterase 4 (PDE4) inhibitors for inflammatory respiratory diseases. Front. Pharmacol. 2020, 11, 259. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Beeh, K.M.; Colgan, B.; Kornmann, O.; Leaker, B.; Watz, H.; Lucci, G.; Geraci, S.; Emirova, A.; Govoni, M.; et al. Effect of the inhaled PDE4 inhibitor CHF6001 on biomarkers of inflammation in COPD. Respir. Res. 2019, 20, 180. [Google Scholar] [CrossRef] [Green Version]
- Oldenburger, A.; Maarsingh, H.; Schmidt, M. Multiple facets of cAMP signalling and physiological impact: cAMP compartmentalization in the lung. Pharmaceuticals 2012, 5, 1291–1331. [Google Scholar] [CrossRef] [Green Version]
- Wójcik-Pszczoła, K.; Chłoń-Rzepa, G.; Jankowska, A.; Ellen, E.; Świerczek, A.; Pociecha, K.; Koczurkiewicz, P.; Piska, K.; Gawędzka, A.; Wyska, E.; et al. Novel phosphodiesterases inhibitors from the group of purine-2,6-dione derivatives as potent modulators of airway smooth muscle cell remodelling. Eur. J. Pharmacol. 2019, 865, 172779. [Google Scholar] [CrossRef]
- Grace, M.S.; Baxter, M.; Dubuis, E.; Birrell, M.A.; Belvisi, M.G. Transient receptor potential (TRP) channels in the airway: Role in airway disease. Br. J. Pharmacol. 2014, 171, 2593–2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Li, S. Transient receptor potential ankyrin 1 (TRPA1) channel and neurogenic inflammation in pathogenesis of asthma. Med. Sci. Monit. 2016, 22, 2917–2923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belvisi, M.G.; Birrell, M.A. The emerging role of transient receptor potential channels in chronic lung disease. Eur. Respir. J. 2017, 50, 1601357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, A. Modulators of transient receptor potential (TRP) channels as therapeutic options in lung disease. Pharm. Basel 2019, 12, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Chen, X.; Zhu, B.; Ramírez-Alcántara, V.; Canzoneri, J.C.; Lee, K.; Sigler, S.; Gary, B.; Li, Y.; Zhang, W.; et al. Suppression of β-catenin/TCF transcriptional activity and colon tumor cell growth by dual inhibition of PDE5 and 10. Oncotarget 2015, 29, 27403–27415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazza, G.A.; Chen, X.; Ward, A.; Coley, A.; Zhou, G.; Buchsbaum, D.J.; Maxuitenko, Y.; Keeton, A.B. Targeting cGMP/PKG signaling for the treatment or prevention of colorectal cancer with novel sulindac derivatives lacking cyclooxygenase inhibitory activity. Oncol. Signal. 2020, 3, 1–6. [Google Scholar] [CrossRef]
- Piazza, G.A.; Ward, A.; Chen, X.; Maxuitenko, Y.; Coley, A.; Aboelella, N.S.; Buchsbaum, D.J.; Boyd, M.R.; Keeton, A.B.; Zhou, G. PDE5 and PDE10 inhibition activates cGMP/PKG signaling to block Wnt/ b-catenin transcription, cancer cell growth, and tumor immunity. Drug Discov. Today 2020. [Google Scholar] [CrossRef]
- Ghofrani, H.A.; Osterloh, I.H.; Grimminger, F. Sildenafil: From angina to erectile dysfunction to pulmonary hypertension and beyond. Nat. Rev. Drug Disc. 2006, 5, 689–702. [Google Scholar] [CrossRef]
- Galie, N.; Ghofrani, H.A.; Torbicki, A.; Barst, R.J.; Rubin, L.J. Sildenafil citrate therapy for pulmonary arterial hypertension. N. Engl. J. Med. 2005, 353, 2148–2157. [Google Scholar] [CrossRef] [Green Version]
- Isidori, A.M.; Cornacchione, M.; Barbagallo, F.; Di Grazia, A.; Barrios, F.; Fassina, L.; Monaco, L.; Giannetta, E.; Gianfrilli, D.; Garofalo, S.; et al. Inhibition of type 5 phosphodiesterase counteracts 2-adrenergic signalling in beating cardiomyocytes. Cardiovasc. Res. 2015, 106, 408–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannetta, E.; Feola, T.; Gianfrilli, D.; Pofi, R.; Dall’Armi, V.; Badagliacca, R.; Barbagallo, F.; Lenzi, A.; Isidori, A.M. Is chronic inhibition of phosphodiesterase type 5 cardioprotective and safe? A meta-analysis of randomized controlled trials. BMC Med. 2014, 12, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, A.; Xi, L.; Kukreja, R.C. Phosphodiesterase-5 inhibitor sildenafil preconditions adult cardiac myocytes against necrosis and apoptosis. Essential role of nitric oxide signaling. J. Biol. Chem. 2005, 280, 12944–12955. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Salloum, F.N.; Xi, L.; Rao, Y.J.; Kukreja, R.C. Erk phosphorylation mediates sildenafil-induced myocardial protection against ischemia-reperfusion injury in mice. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H1236–H1243. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Ockaili, R.; Salloum, F.; Kukreja, R.C. Protein kinase c plays an essential role in sildenafil-induced cardioprotection in rabbits. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H1455–H1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fibbi, B.; Morelli, A.; Marini, M.; Zhang, X.H.; Mancina, R.; Vignozzi, L.; Filippi, S.; Chavalmane, A.; Silvestrini, E.; Colli, E.; et al. Atorvastatin but not elocalcitol increases sildenafil responsiveness spontaneously hypertensive rats by regulating the RhoA/ROCK pathway. J. Androl. 2008, 29, 70–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, M.J.; Ersboll, M.; Axelsson, A.; Gustafsson, F.; Hassager, C.; Kober, L. Sildenafil and diastolic dysfunction after acute myocardial infarction in patients with preserved ejection fraction: SIDAMI. Circulation 2013, 127, 1200–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salloum, F.N.; Abbate, A.; Das, A.; Houser, J.E.; Mudrick, C.A.; Qureshi, I.Z. Sildenafil (Viagra) attenuates ischemic cardiomyopathy and improves left ventricular function in mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1398–H1406. [Google Scholar] [CrossRef]
- Geng, Y.; Li, E.; Mu, Q.; Zhang, Y.; Wei, X.; Li, H.; Cheng, L.; Zhang, B. Hydrogen sulfide inhalation decreases early blood-brain barrier permeability and brain edema induced by cardiac arrest and resuscitation. J. Cereb. Blood Flow Metab. 2015, 35, 494–500. [Google Scholar] [CrossRef] [Green Version]
- Zarychanski, R.; Abou-Setta, A.M.; Turgeon, A.F.; Houston, B.L. Association of hydroxyethyl starch administration with mortality and acute kidney injury in critically ill patients requiring volume resuscitation: A systematic review and meta-analysis. JAMA 2013, 309, 678–688. [Google Scholar] [CrossRef]
- Ntontsi, P.; Detta, A.; Bakakos, P.; Loukides, S.; Hillas, G. Experimental and investigational phosphodiesterase inhibitors in development for asthma. Expert Opin. Investig. Drugs 2019, 28, 261–266. [Google Scholar] [CrossRef]
- Paul, T.; Salazar-Degracia, A.; Peinado, V.I.; Tura-Ceide, O. Soluble guanylate cyclase stimulation reduces oxidative stress in experimental chronic obstructive pulmonary disease. PLoS ONE 2018, 13, e0190628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glynos, C.; Dupont, L.L.; Vassilakopoulos, T.; Papapetropoulos, A.; Brouckaert, P.; Giannis, A. The role of soluble guanylyl cyclase in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2013, 188, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.; Cattani-Cavalieri, I.; Musheshe, N.; Nikolaev, V.O.; Schmidt, M. Phosphodiesterases as therapeutic targets for respiratory diseases. Pharmacol. Ther. 2019, 197, 225–242. [Google Scholar] [CrossRef]
- Bieber, M.; Schuhmann, M.K.; Volz, J. Description of a novel phosphodiesterase (PDE)-3 inhibitor protecting mice from ischemic stroke independent from platelet function. Stroke 2019, 50, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Okuda, Y.K.Y.; Yamashita, K. Cilostazol. Cardiovasc. Drug Rev. 1993, 11, 451–465. [Google Scholar] [CrossRef]
- Criqui, M.H.; Fronek, A.; Barrett-Connor, E.; Klauber, M.R.; Gabriel, S.; Goodman, D. The prevalence of peripheral arterial disease in a defined population. Circulation 1985, 71, 510–515. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, Y.; Katayama, Y.; Uchiyama, S.; Yamaguchi, T.; Handa, S.; Matsuoka, K.; Ohashi, Y.; Tanahashi, N.; Yamamoto, H.; Genka, C.; et al. Cilostazol for prevention of secondary stroke (CSPS 2): An aspirin-controlled, double-blind, randomised non-inferiority trial. Lancet Neurol. 2010, 9, 959–968. [Google Scholar] [CrossRef]
- Han, Y.; Li, Y.; Wang, S.; Jing, Q.; Wang, Z.; Wang, D.; Shu, Q.; Tang, X. Cilostazol in addition to aspirin and clopidogrel improves long-term outcomes after percutaneous coronary intervention in patients with acute coronary syndromes: A randomized, controlled study. Am. Heart J. 2009, 157, 733–739. [Google Scholar] [CrossRef]
- Suh, J.W.; Lee, S.P.; Park, K.W.; Lee, H.Y.; Kang, H.J.; Koo, B.K.; Cho, Y.S.; Youn, T.J.; Chae, I.H.; Rha, S.W.; et al. Multicenter randomized trial evaluating the efficacy of cilostazol on ischemic vascular complications after drug-eluting stent implantation for coronary heart disease: Results of the CILON-T (influence of cilostazol-based triple antiplatelet therapy on ischemic complication after drug-eluting stenT implantation) trial. J. Am. Coll Cardiol 2011, 57, 280–289. [Google Scholar] [CrossRef] [Green Version]
- Yamagami, H.; Sakai, N.; Matsumaru, Y.; Sakai, C.; Kai, Y.; Sugiu, K.; Fujinaka, T.; Matsumoto, Y.; Miyachi, S.; Yoshimura, S.; et al. Periprocedural cilostazol treatment and restenosis after carotid artery stenting: The retrospective study of in-stent restenosis after carotid artery stenting (ReSISteR-CAS). J. Stroke Cerebrovasc. Dis. 2012, 21, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Jennings, D.L.; Kalus, J.S. Addition of cilostazol to aspirin and a thienopyridine for prevention of restenosis after coronary artery stenting: A meta-analysis. J. Clin. Pharmacol. 2010, 50, 415–421. [Google Scholar] [CrossRef]
- Schrör, K. The pharmacology of cilostazol. Diabetes Obes. Metab. 2002, 4, S14–S19. [Google Scholar] [CrossRef] [PubMed]
- Gresele, P.; Momi, S.; Falcinelli, E. Anti-platelet therapy: Phosphodiesterase inhibitors. Br. J. Clin. Pharmacol. 2011, 72, 634–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisert, W.G. Dipyridamole. In Platelets; Michelson, A.D., Ed.; Academic: San Diego, CA, USA, 2007; pp. 1165–1179. [Google Scholar]
- Aktas, B.; Utz, A.; Hoenig-Liedl, P.; Walter, U.; Geiger, J. Dipyridamole enhances NO/cGMP-mediated vasodilator stimulated phosphoprotein phosphorylation and signaling in human platelets: In vitro and in vivo/ex vivo studies. Stroke 2003, 34, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Klabunde, R.E. Dipyridamole inhibition of adenosine metabolism in human blood. Eur. J. Pharmacol. 1983, 93, 21–26. [Google Scholar] [CrossRef]
- Neri Serneri, G.G.; Masotti, G.; Poggesi, L.; Galanti, G.; Morettini, A. Enhanced prostacyclin production by dipyridamole in man. Eur. J. Clin. Pharmacol. 1981, 21, 9–15. [Google Scholar] [CrossRef]
- Solaimanzadeh, I. Acetazolamide, nifedipine and phosphodiesterase inhibitors: Rationale for their utilization as adjunctive countermeasures in the treatment of Coronavirus disease 2019 (COVID-19). Cureus 2020, 12, e7343. [Google Scholar] [CrossRef] [Green Version]
- Isidori, A.M.; Giannetta, E.; Pofi, R.; Venneri, M.A.; Gianfrilli, D.; Campolo, F.; Mastroianni, C.M.; Lenzi, A.; d’Ettorre, G. Targeting the NO-cGMP-PDE5 pathway in COVID-19 infection. Andrologia 2020. [Google Scholar] [CrossRef]
- Anderson, S.G.; Hutchings, D.C.; Woodward, M.; Rahimi, K.; Rutter, M.K.; Kirby, M.; Hackett, G.; Trafford, A.W.; Heald, A.H. Phosphodiesterase type-5 inhibitor use in type 2 diabetes is associated with a reduction in all-cause mortality. Heart 2016, 102, 1750–1756. [Google Scholar] [CrossRef] [Green Version]
- Dalamaga, M.; Karampela, I.; Mantzoros, C.S. Commentary: Phosphodiesterase 4 inhibitors as potential adjunct treatment targeting the cytokine storm in COVID-19. Metabolism 2020, 109, S0026–S0495. [Google Scholar] [CrossRef] [PubMed]
- Angelidi, A.M.; Belanger, M.J.; Mantzoros, C.S. COVID-19 and diabetes mellitus: What we know, how our patients should be treated now, and what should happen next. Metabolism 2020, 2020, 154245. [Google Scholar] [CrossRef] [PubMed]
- Bridgewood, C.; Damiani, G.; Sharif, K.; Abdulla Watad, A.; Bragazzi, N.L.; Quartuccio, L.; Savic, S.; McGonagle, D. Rationale for evaluating PDE4 inhibition for mitigating against severe inflammation in COVID-19 pneumonia and beyond. Isr. Med. Assoc. J. 2020, 22, 335–339. [Google Scholar] [PubMed]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giorgi, M.; Cardarelli, S.; Ragusa, F.; Saliola, M.; Biagioni, S.; Poiana, G.; Naro, F.; Massimi, M. Phosphodiesterase Inhibitors: Could They Be Beneficial for the Treatment of COVID-19? Int. J. Mol. Sci. 2020, 21, 5338. https://doi.org/10.3390/ijms21155338
Giorgi M, Cardarelli S, Ragusa F, Saliola M, Biagioni S, Poiana G, Naro F, Massimi M. Phosphodiesterase Inhibitors: Could They Be Beneficial for the Treatment of COVID-19? International Journal of Molecular Sciences. 2020; 21(15):5338. https://doi.org/10.3390/ijms21155338
Chicago/Turabian StyleGiorgi, Mauro, Silvia Cardarelli, Federica Ragusa, Michele Saliola, Stefano Biagioni, Giancarlo Poiana, Fabio Naro, and Mara Massimi. 2020. "Phosphodiesterase Inhibitors: Could They Be Beneficial for the Treatment of COVID-19?" International Journal of Molecular Sciences 21, no. 15: 5338. https://doi.org/10.3390/ijms21155338