Human Semenogelin 1 Promotes Sperm Survival in the Mouse Female Reproductive Tract

 , , , and
, , , and 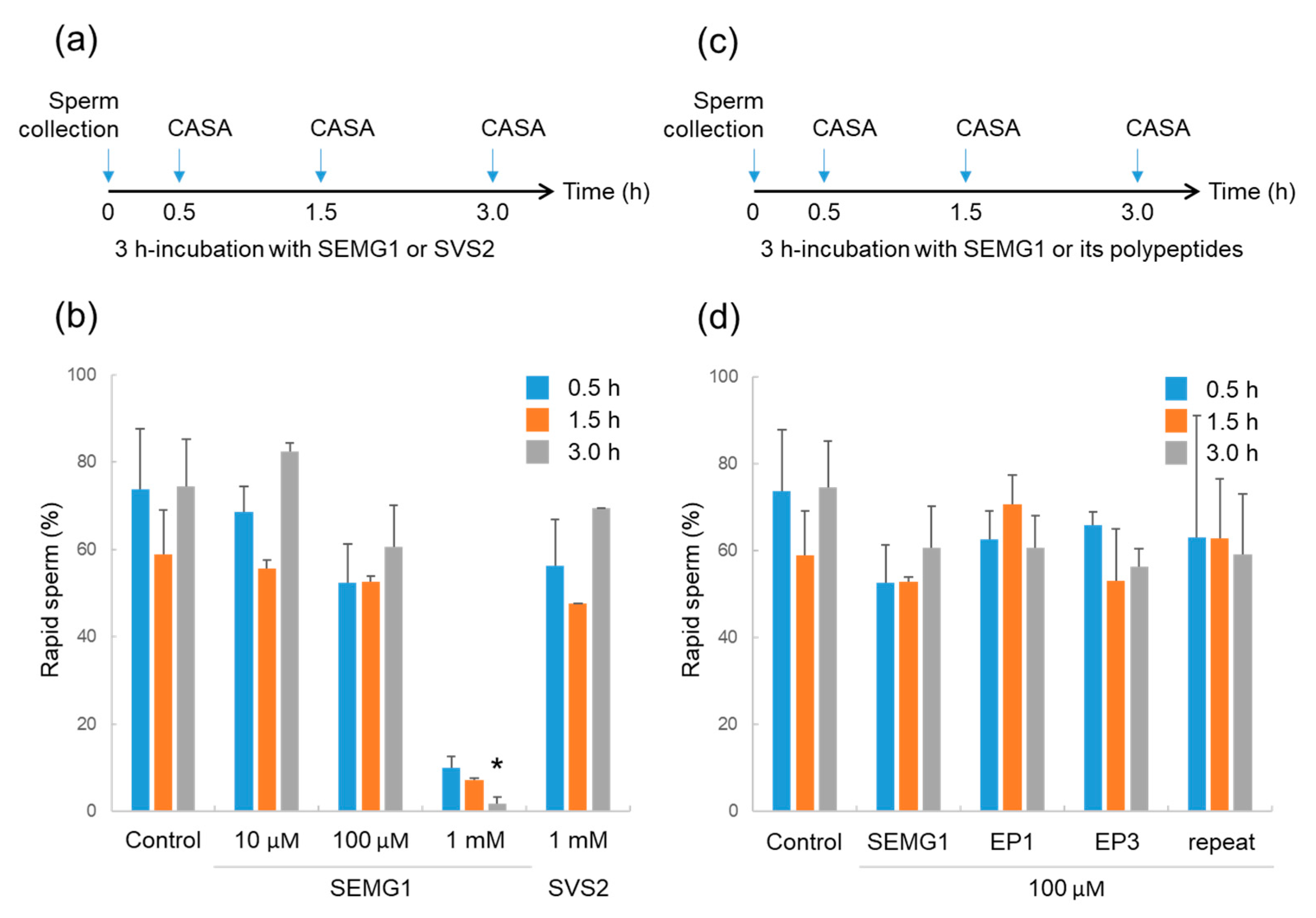{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Human SEMG1 on Mouse Sperm
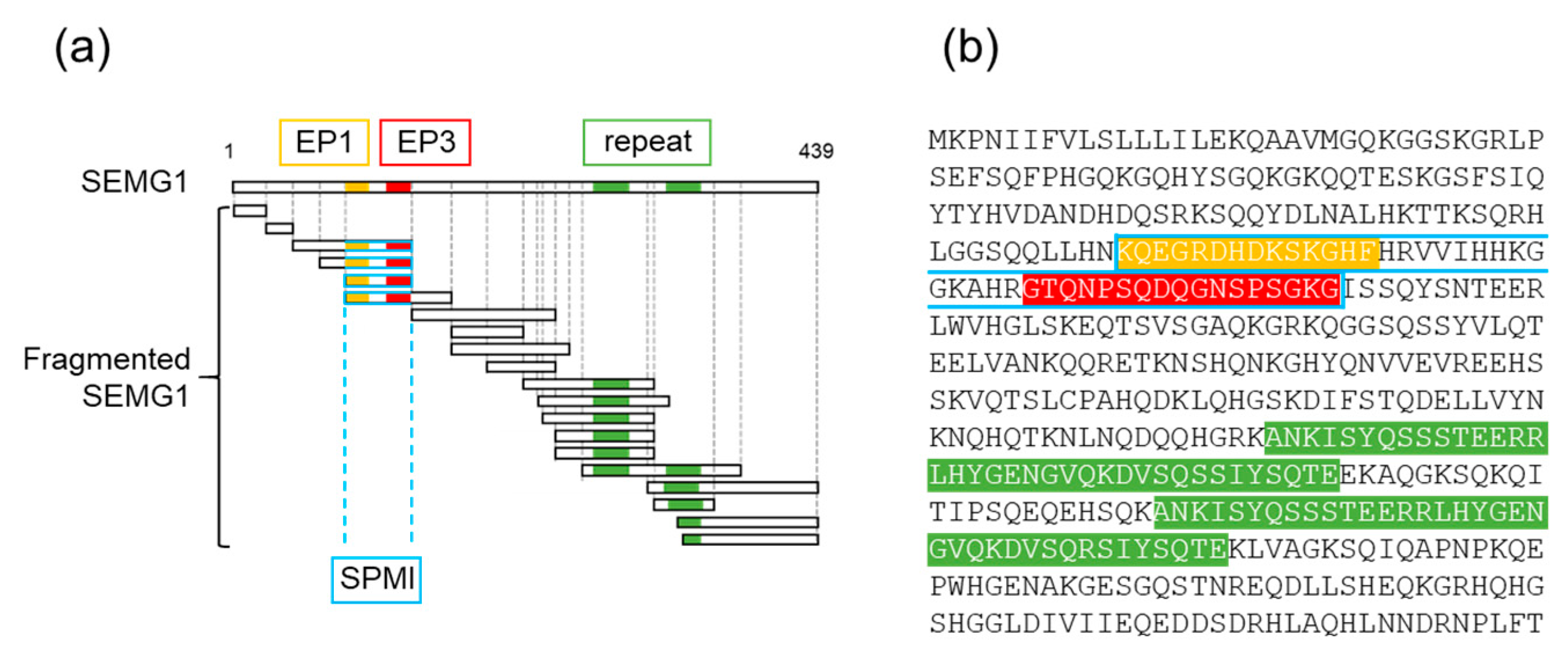
2.2. Effects of SEMG1-Derived Polypeptides on Mouse Sperm
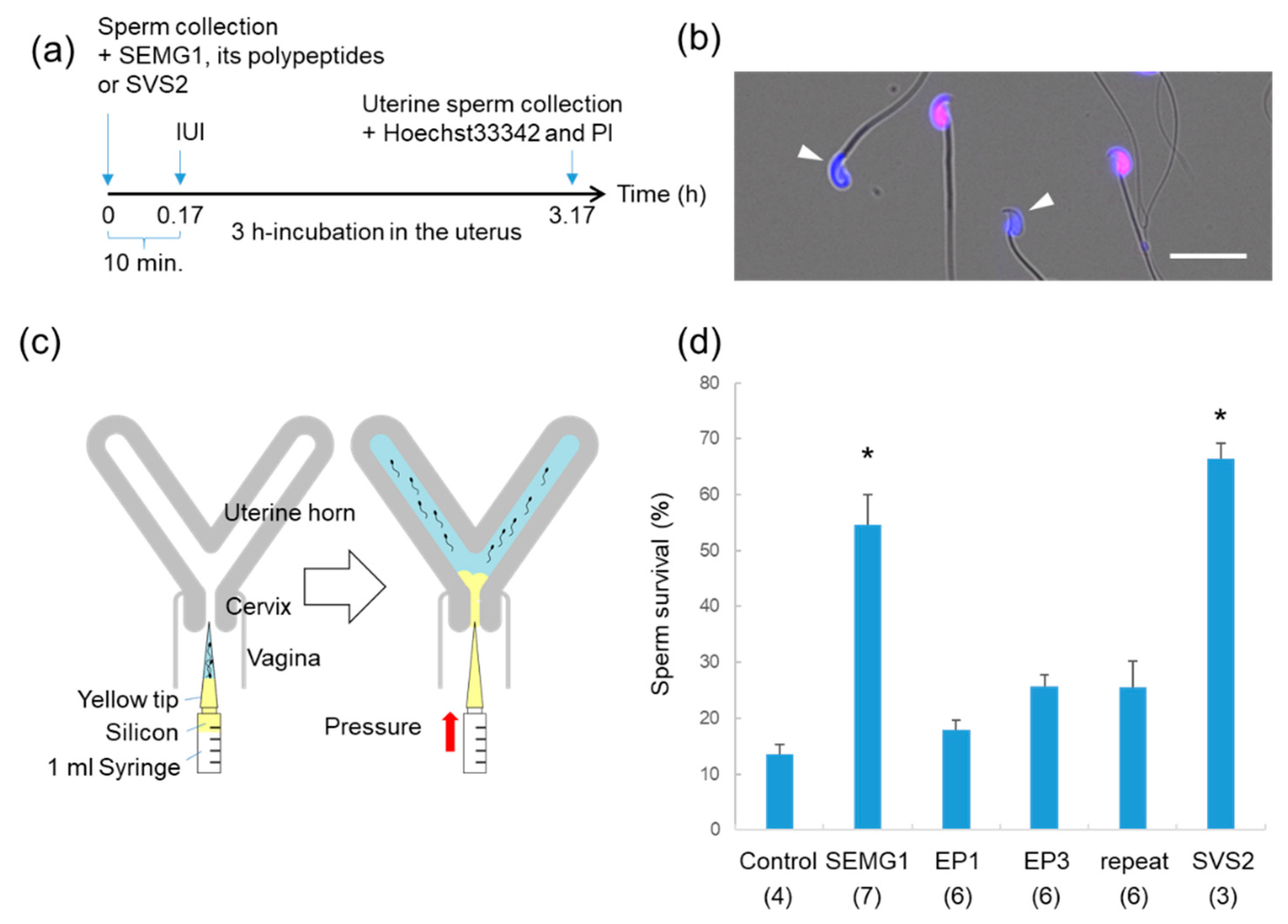
2.3. Evaluation of Sperm Survival After IUI
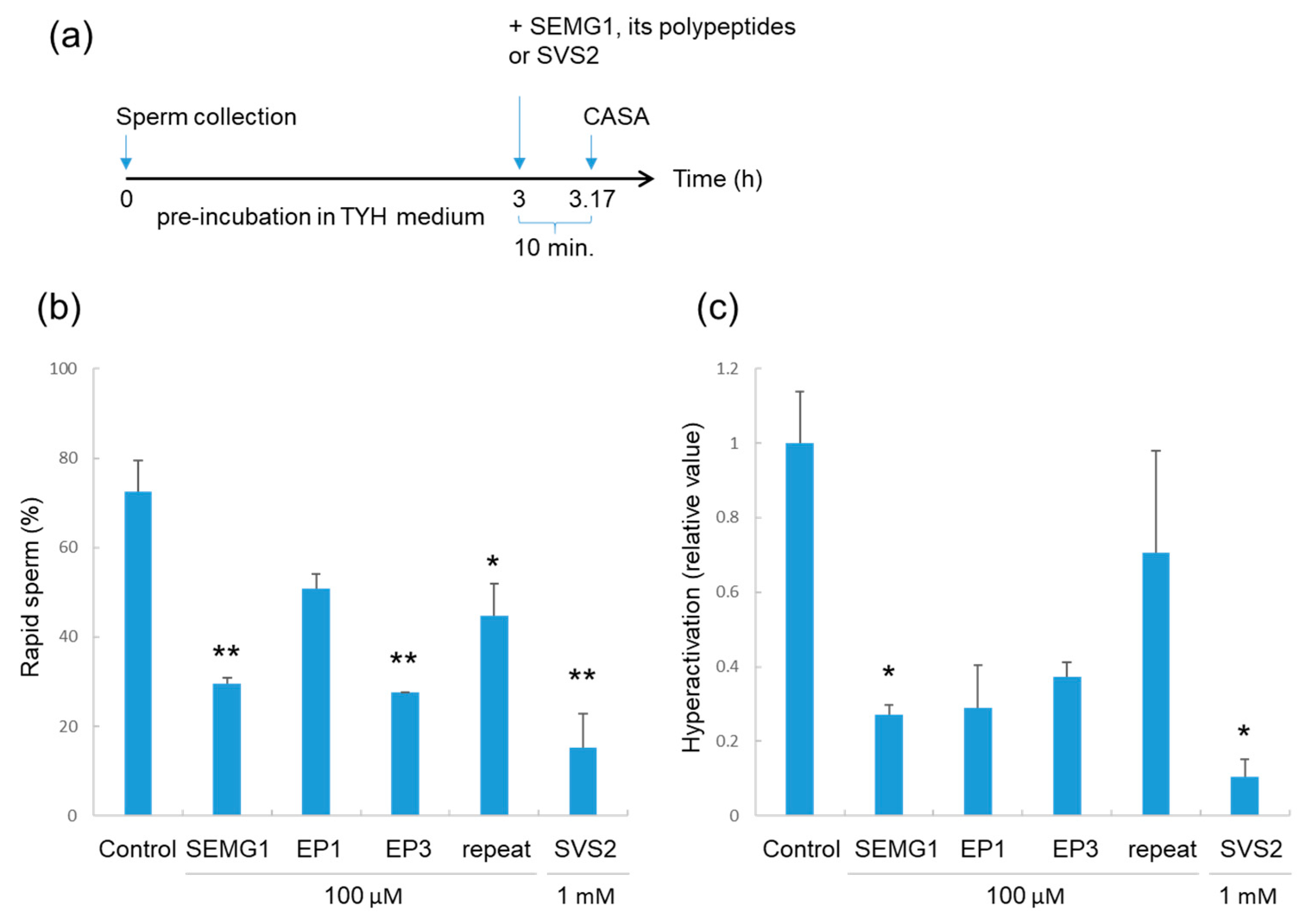
2.4. Effects of SEMG1 and its Polypeptides on Pre-Incubated Sperm
3. Discussion
4. Materials and Methods
4.1. Animal Care
4.2. Materials
4.3. Measurement of Sperm Motility
4.4. Sperm Survival Assay after IUI
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chang, M.C. Fertilizing capacity of spermatozoa deposited into the fallopian tubes. Nature 1951, 168, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.R. The ’Capacitation’ of the Mammalian Sperm. Nature 1952, 170, 326. [Google Scholar] [CrossRef] [PubMed]
- Shadan, S.; James, P.S.; Howes, E.A.; Jones, R. Cholesterol efflux alters lipid raft stability and distribution during capacitation of boar spermatozoa. Biol. Reprod. 2004, 71, 253–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, J.L. Factors regulating sperm capacitation. Syst. Biol. Reprod. Med. 2010, 56, 334–348. [Google Scholar] [CrossRef]
- Visconti, P.E.; Krapf, D.; de la Vega-Beltran, J.L.; Acevedo, J.J.; Darszon, A. Ion channels, phosphorylation and mammalian sperm capacitation. Asian J. Androl. 2011, 13, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Yanagimachi, R.; Noda, Y.D. Electron microscope studies of sperm incorporation into the golden hamster egg. Am. J. Anat. 1970, 128, 429–462. [Google Scholar] [CrossRef]
- Demott, R.P.; Suarez, S.S. Hyperactivated sperm progress in the mouse oviduct. Biol. Reprod. 1992, 46, 779–785. [Google Scholar] [CrossRef]
- Chang, M.C. A detrimental effect of seminal plasma on the fertilizing capacity of sperm. Nature 1957, 179, 258–259. [Google Scholar] [CrossRef]
- Bedford, J.M.; Chang, M.C. Removal of decapacitation factor from seminal plasma by high-speed centrifugation. Am. J. Physiol. 1962, 202, 179–181. [Google Scholar] [CrossRef]
- Pang, S.F.; Chow, P.H.; Wong, T.M. The role of the seminal vesicles, coagulating glands and prostate glands on the fertility and fecundity of mice. J. Reprod. Fertil. 1979, 56, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Kawano, N.; Araki, N.; Yoshida, K.; Hibino, T.; Ohnami, N.; Makino, M.; Kanai, S.; Hasuwa, H.; Yoshida, M.; Miyado, K.; et al. Seminal vesicle protein SVS2 is required for sperm survival in the uterus. Proc. Natl. Acad. Sci. USA 2014, 111, 4145–4150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bromfield, J.J.; Schjenken, J.E.; Chin, P.Y.; Care, A.S.; Jasper, M.J.; Robertson, S.A. Maternal tract factors contribute to paternal seminal fluid impact on metabolic phenotype in offspring. Proc. Natl. Acad. Sci. USA 2014, 111, 2200–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, M.; Gagnon, C. Purification and characterization of the active precursor of a human sperm motility inhibitor secreted by the seminal vesicles: Identity with semenogelin. Biol. Reprod. 1996, 55, 813–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilja, H. A kallikrein-like serine protease in prostatic fluid cleaves the predominant seminal vesicle protein. J. Clin. Investig. 1985, 76, 1899–1903. [Google Scholar] [CrossRef] [PubMed]
- Lilja, H.; Abrahamsson, P.A.; Lundwall, A. Semenogelin, the predominant protein in human semen. Primary structure and identification of closely related proteins in the male accessory sex glands and on the spermatozoa. J. Biol. Chem. 1989, 264, 1894–1900. [Google Scholar] [PubMed]
- Iwamoto, T.; Gagnon, C. A human seminal plasma protein blocks the motility of human spermatozoa. J. Urol. 1988, 140, 1045–1048. [Google Scholar] [CrossRef]
- Yoshida, K.; Yamasaki, T.; Yoshiike, M.; Takano, S.; Sato, I.; Iwamoto, T. Quantification of seminal plasma motility inhibitor/semenogelin in human seminal plasma. J. Androl. 2003, 24, 878–884. [Google Scholar] [CrossRef] [Green Version]
- Terai, K.; Yoshida, K.; Yoshiike, M.; Fujime, M.; Iwamoto, T. Association of seminal plasma motility inhibitors/semenogelins with sperm in asthenozoospermia-infertile men. Urol. Int. 2010, 85, 209–215. [Google Scholar] [CrossRef]
- Shindo, M.; Inui, M.; Kang, W.; Tamano, M.; Tingwei, C.; Takada, S.; Hibino, T.; Yoshida, M.; Yoshida, K.; Okada, H.; et al. Deletion of a Seminal Gene Cluster Reinforces a Crucial Role of SVS2 in Male Fertility. Int. J. Mol. Sci. 2019, 20, 4557. [Google Scholar] [CrossRef] [Green Version]
- Dorus, S.; Evans, P.D.; Wyckoff, G.J.; Choi, S.S.; Lahn, B.T. Rate of molecular evolution of the seminal protein gene SEMG2 correlates with levels of female promiscuity. Nat. Genet. 2004, 36, 1326–1329. [Google Scholar] [CrossRef]
- de Lamirande, E.; Yoshida, K.; Yoshiike, T.M.; Iwamoto, T.; Gagnon, C. Semenogelin, the main protein of semen coagulum, inhibits human sperm capacitation by interfering with the superoxide anion generated during this process. J. Androl. 2001, 22, 672–679. [Google Scholar] [PubMed]
- Silva, E.J.; Hamil, K.G.; O’Rand, M.G. Interacting proteins on human spermatozoa: Adaptive evolution of the binding of semenogelin I to EPPIN. PLoS ONE 2013, 8, e82014. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.J.; Hamil, K.G.; Richardson, R.T.; O’Rand, M.G. Characterization of EPPIN’s semenogelin I binding site: A contraceptive drug target. Biol. Reprod. 2012, 87, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, K.; Yoshida, K.; Yoshiike, M.; Shimada, K.; Nishiyama, H.; Takamizawa, S.; Yanagida, K.; Iwamoto, T. Relationship between Semenogelins bound to human sperm and other semen parameters and pregnancy outcomes. Basic Clin. Androl. 2017, 27, 15. [Google Scholar] [CrossRef] [Green Version]
- Kawano, N.; Yoshida, M. Semen-coagulating protein, SVS2, in mouse seminal plasma controls sperm fertility. Biol. Reprod. 2007, 76, 353–361. [Google Scholar] [CrossRef]
- Kawano, N.; Yoshida, K.; Iwamoto, T.; Yoshida, M. Ganglioside GM1 Mediates Decapacitation Effects of SVS2 on Murine Spermatozoa. Biol. Reprod. 2008, 79, 1153–1159. [Google Scholar] [CrossRef] [Green Version]
- Araki, N.; Trencsenyi, G.; Krasznai, Z.T.; Nizsaloczki, E.; Sakamoto, A.; Kawano, N.; Miyado, K.; Yoshida, K.; Yoshida, M. Seminal vesicle secretion 2 acts as a protectant of sperm sterols and prevents ectopic sperm capacitation in mice. Biol. Reprod. 2015, 92, 8, 1–10. [Google Scholar] [CrossRef]
- Robert, M.; Gibbs, B.F.; Jacobson, E.; Gagnon, C. Characterization of prostate-specific antigen proteolytic activity on its major physiological substrate, the sperm motility inhibitor precursor/semenogelin I. Biochemistry 1997, 36, 3811–3819. [Google Scholar] [CrossRef]
- Yoshida, K.; Kawano, N.; Yoshiike, M.; Yoshida, M.; Iwamoto, T.; Morisawa, M. Physiological roles of semenogelin I and zinc in sperm motility and semen coagulation on ejaculation in humans. Mol. Hum. Reprod. 2008, 14, 151–156. [Google Scholar] [CrossRef] [Green Version]
- de Lamirande, E. Semenogelin, the main protein of the human semen coagulum, regulates sperm function. Semin. Thromb. Hemost. 2007, 33, 60–68. [Google Scholar] [CrossRef]
- Carlson, B.M. Human Embryology and Developmental Biology, 5th ed.; Saunders: Philadelphia, PA, USA, 2013; p. 520. [Google Scholar]
- Miyano, S.; Yoshida, K.; Yoshiike, M.; Miyamoto, C.; Furuichi, Y.; Iwamoto, T. A large deletion of the repeat site in semenogelin I is not involved in male infertility. Int. J. Mol. Med. 2003, 11, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Goodson, S.G.; Zhang, Z.; Tsuruta, J.K.; Wang, W.; O’Brien, D.A. Classification of mouse sperm motility patterns using an automated multiclass support vector machines model. Biol. Reprod. 2011, 84, 1207–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, M.; Gagnon, C. Semenogelin I: A coagulum forming, multifunctional seminal vesicle protein. Cell. Mol. Life Sci. 1999, 55, 944–960. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakaguchi, D.; Miyado, K.; Iwamoto, T.; Okada, H.; Yoshida, K.; Kang, W.; Suzuki, M.; Yoshida, M.; Kawano, N. Human Semenogelin 1 Promotes Sperm Survival in the Mouse Female Reproductive Tract. Int. J. Mol. Sci. 2020, 21, 3961. https://doi.org/10.3390/ijms21113961
Sakaguchi D, Miyado K, Iwamoto T, Okada H, Yoshida K, Kang W, Suzuki M, Yoshida M, Kawano N. Human Semenogelin 1 Promotes Sperm Survival in the Mouse Female Reproductive Tract. International Journal of Molecular Sciences. 2020; 21(11):3961. https://doi.org/10.3390/ijms21113961
Chicago/Turabian StyleSakaguchi, Daiki, Kenji Miyado, Teruaki Iwamoto, Hiroshi Okada, Kaoru Yoshida, Woojin Kang, Miki Suzuki, Manabu Yoshida, and Natsuko Kawano. 2020. "Human Semenogelin 1 Promotes Sperm Survival in the Mouse Female Reproductive Tract" International Journal of Molecular Sciences 21, no. 11: 3961. https://doi.org/10.3390/ijms21113961