Functional Analysis of Differentially Expressed Acetylated Spermatozoal Proteins in Infertile Men with Unilateral and Bilateral Varicocele


Abstract
:1. Introduction
2. Results
2.1. Semen Analysis and Advanced Sperm Function Tests
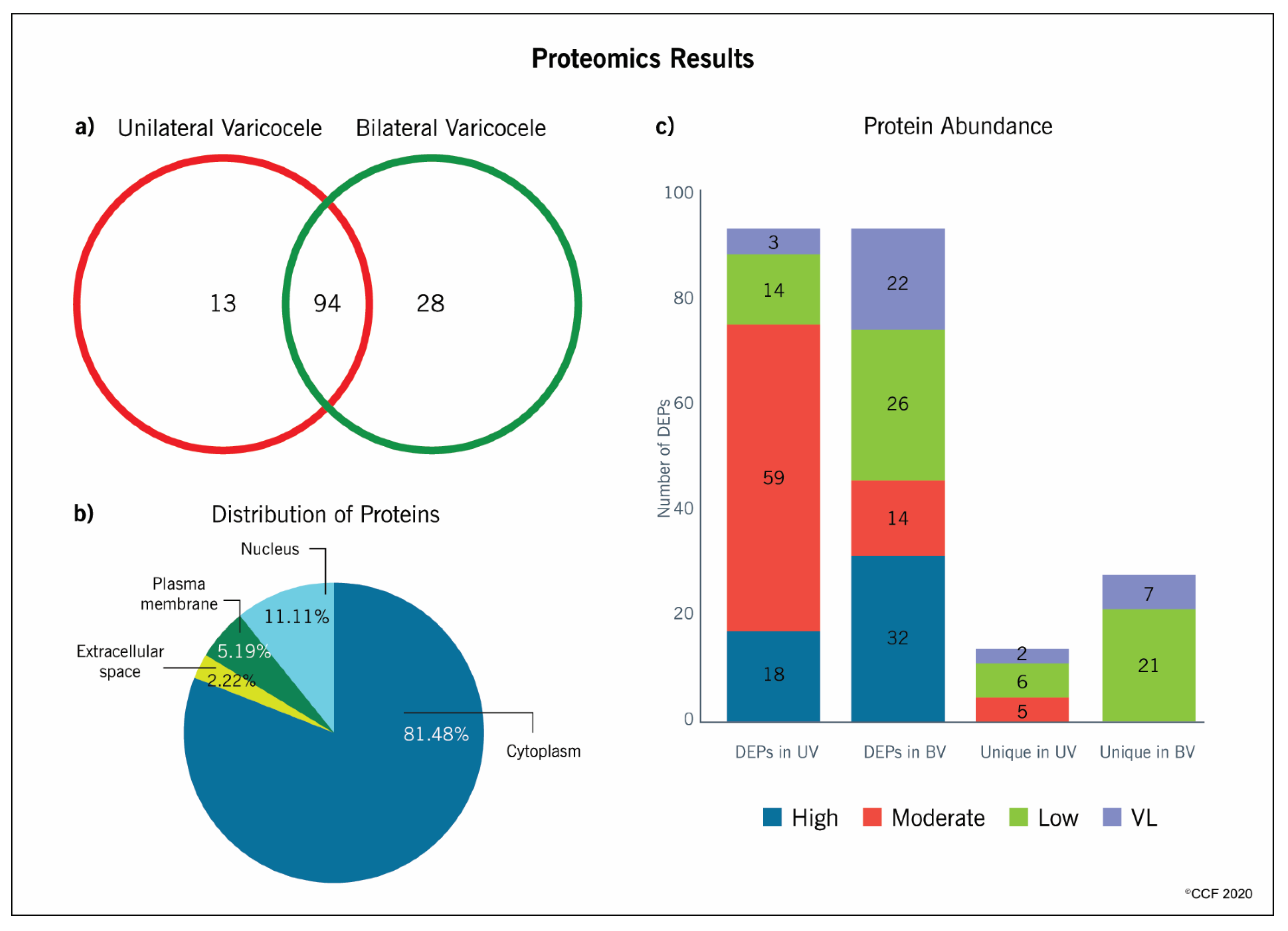
2.2. Proteomic Analysis
2.3. Distribution Pattern and Functional Annotation of Proteins
2.4. Classification of DEPs with Predicted Acetylation
2.5. Identification of DEPs Involved in Important Networks
2.6. Protein–Protein Interaction
2.7. Potential Protein Biomarkers and Western Blot Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Patient Characteristics
5.2. Proteomic Analysis
5.3. Database Searching and Protein Identification
5.4. Quantitative Proteomics
5.5. Large-Scale Identification of Acetylation Sites
5.6. Functional Analysis
5.7. Bioinformatic Analysis of Protein–Protein Interaction
5.8. Immunoprecipitation and Western Blotting of Acetylated Proteins
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Samanta, L.; Durairajanayagam, D. Introduction. In Proteomics in Human Reproduction: Biomarkers for Millennials; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–8. [Google Scholar]
- Marmar, J.L. The pathophysiology of varicoceles in the light of current molecular and genetic information. Hum. Reprod. Update 2001, 7, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Dahl, E.V.; Herrick, J.F. A vascular mechanism for maintaining testicular temperature by counter-current exchange. Surg. Gynecol. Obstet. 1959, 108, 697–705. [Google Scholar] [PubMed]
- Jarow, J.P.; Coburn, M.; Sigman, M. Incidence of varicoceles in men with primary and secondary infertility. Urology 1996, 47, 73–76. [Google Scholar] [CrossRef]
- Baazeem, A.; Belzile, E.; Ciampi, A.; Dohle, G.; Jarvi, K.; Salonia, A.; Weidner, W.; Zini, A. Varicocele and male factor infertility treatment: A new meta-analysis and review of the role of varicocele repair. Eur. Urol. 2011, 60, 796–808. [Google Scholar] [CrossRef]
- Esteves, S.C.; Agarwal, A. Afterword to varicocele and male infertility: Current concepts and future perspectives. Asian J. Androl. 2016, 18, 319–322. [Google Scholar] [CrossRef]
- Hosseinifar, H.; Sabbaghian, M.; Nasrabadi, D.; Modarresi, T.; Dizaj, A.V.; Gourabi, H.; Gilani, M.A. Study of the effect of varicocelectomy on sperm proteins expression in patients with varicocele and poor sperm quality by using two-dimensional gel electrophoresis. J. Assist. Reprod. Genet. 2014, 31, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Hosseinifar, H.; Gourabi, H.; Salekdeh, G.H.; Alikhani, M.; Mirshahvaladi, S.; Sabbaghian, M.; Modarresi, T.; Gilani, M.A. Study of sperm protein profile in men with and without varicocele using two-dimensional gel electrophoresis. Urology 2013, 81, 293–300. [Google Scholar] [CrossRef]
- Camargo, M.; Intasqui Lopes, P.; Del Giudice, P.T.; Carvalho, V.M.; Cardozo, K.H.; Andreoni, C.; Fraietta, R.; Bertolla, R.P. Unbiased label-free quantitative proteomic profiling and enriched proteomic pathways in seminal plasma of adult men before and after varicocelectomy. Hum. Reprod. (Oxford, England) 2013, 28, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Sharma, R.; Samanta, L.; Durairajanayagam, D.; Sabanegh, E. Proteomic signatures of infertile men with clinical varicocele and their validation studies reveal mitochondrial dysfunction leading to infertility. Asian J. Androl. 2016, 18, 282–291. [Google Scholar] [CrossRef]
- Baker, M.A.; Nixon, B.; Naumovski, N.; Aitken, R.J. Proteomic insights into the maturation and capacitation of mammalian spermatozoa. Syst. Biol. Reprod. Med. 2012, 58, 211–217. [Google Scholar] [CrossRef]
- Yu, H.; Diao, H.; Wang, C.; Lin, Y.; Yu, F.; Lu, H.; Xu, W.; Li, Z.; Shi, H.; Zhao, S.; et al. Acetylproteomic analysis reveals functional implications of lysine acetylation in human spermatozoa (sperm). Mol. Cell. Proteom. MCP 2015, 14, 1009–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krejci, J.; Stixova, L.; Pagacova, E.; Legartova, S.; Kozubek, S.; Lochmanova, G.; Zdrahal, Z.; Sehnalova, P.; Dabravolski, S.; Hejatko, J.; et al. Post-Translational Modifications of Histones in Human Sperm. J. Cell. Biochem. 2015, 116, 2195–2209. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Jiang, M.; Zhou, T.; Guo, Y.; Cui, Y.; Guo, X.; Sha, J. Insights into the lysine acetylproteome of human sperm. J. Proteom. 2014, 109, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Pang, A.; Rennert, O. Protein acetylation and spermatogenesis. Reprod. Syst. Sex. Disord. Curr. Res. 2013, (Suppl 1), 5. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Sharma, R.; Durairajanayagam, D.; Cui, Z.; Ayaz, A.; Gupta, S.; Willard, B.; Gopalan, B.; Sabanegh, E. Differential proteomic profiling of spermatozoal proteins of infertile men with unilateral or bilateral varicocele. Urology 2015, 85, 580–588. [Google Scholar] [CrossRef]
- Samanta, L.; Swain, N.; Ayaz, A.; Venugopal, V.; Agarwal, A. Post-Translational Modifications in sperm Proteome: The Chemistry of Proteome diversifications in the Pathophysiology of male factor infertility. Biochim. Biophys. Acta 2016, 1860, 1450–1465. [Google Scholar] [CrossRef]
- Walker, W.H.; Easton, E.; Moreci, R.S.; Toocheck, C.; Anamthathmakula, P.; Jeyasuria, P. Restoration of Spermatogenesis and Male Fertility Using an Androgen Receptor Transgene. PLoS ONE 2015, 10, e0120783. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, L.; Smith, L.B. Androgen receptor roles in spermatogenesis and infertility. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Stanton, P.G.; Sluka, P.; Foo, C.F.; Stephens, A.N.; Smith, A.I.; McLachlan, R.I.; O’Donnell, L. Proteomic changes in rat spermatogenesis in response to in vivo androgen manipulation; impact on meiotic cells. PLoS ONE 2012, 7, e41718. [Google Scholar] [CrossRef] [Green Version]
- Solakidi, S.; Psarra, A.M.; Nikolaropoulos, S.; Sekeris, C.E. Estrogen receptors alpha and beta (ERalpha and ERbeta) and androgen receptor (AR) in human sperm: Localization of ERbeta and AR in mitochondria of the midpiece. Hum. Reprod. (Oxford, England) 2005, 20, 3481–3487. [Google Scholar] [CrossRef] [Green Version]
- Aquila, S.; Middea, E.; Catalano, S.; Marsico, S.; Lanzino, M.; Casaburi, I.; Barone, I.; Bruno, R.; Zupo, S.; Ando, S. Human sperm express a functional androgen receptor: Effects on PI3K/AKT pathway. Hum. Reprod. (Oxford, England) 2007, 22, 2594–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkaram, A.; McCullough, A. Varicocele and its effect on testosterone: Implications for the adolescent. Transl. Androl. Urol. 2014, 3, 413–417. [Google Scholar]
- Gioeli, D.; Paschal, B.M. Post-translational modification of the androgen receptor. Mol. Cell. Endocrinol. 2012, 352, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Coffey, K.; Robson, C.N. Regulation of the androgen receptor by post-translational modifications. J. Endocrinol. 2012, 215, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, T.; Sontag, E.; Chen, P.; Levine, A. Transcriptional control of human p53-regulated genes. Nat. Rev. Mol. Cell Biol. 2008, 9, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, S.; Gentile, T.; Cuomo, F.; De Filippo, S.; Aprea, G.E.; Guida, J. Quantitative evaluation of p53 as a new indicator of DNA damage in human spermatozoa. J. Hum. Reprod. Sci. 2014, 7, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, S.; Gentile, T.; Gentile, M.; Morelli, A.; Donnarumma, F.; Cuomo, F.; De Filippo, S.; Montano, L. p53 Protein Evaluation on Spermatozoa DNA in Fertile and Infertile Males. J. Hum. Reprod. Sci. 2019, 12, 114–121. [Google Scholar] [CrossRef]
- Chang, F.W.; Sun, G.H.; Cheng, Y.Y.; Chen, I.C.; Chien, H.H.; Wu, G.J. Effects of varicocele upon the expression of apoptosis-related proteins. Andrologia 2010, 42, 225–230. [Google Scholar] [CrossRef]
- Huang, Y.; Jin, Y.; Yan, C.H.; Yu, Y.; Bai, J.; Chen, F.; Zhao, Y.Z.; Fu, S.B. Involvement of Annexin A2 in p53 induced apoptosis in lung cancer. Mol. Cell. Biochem. 2008, 309, 117–123. [Google Scholar] [CrossRef]
- Munuce, M.J.; Marini, P.E.; Teijeiro, J.M. Expression profile and distribution of Annexin A1, A2 and A5 in human semen. Andrologia 2019, 51, e13224. [Google Scholar] [CrossRef]
- Nazmi, A.R.; Ozorowski, G.; Pejic, M.; Whitelegge, J.P.; Gerke, V.; Luecke, H. N-terminal acetylation of annexin A2 is required for S100A10 binding. Biol. Chem. 2012, 393, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, T.R.; Samanta, L.; Agarwal, A.; Pushparaj, P.N.; Panner Selvam, M.K.; Sharma, R. Proteomic Signatures Reveal Differences in Stress Response, Antioxidant Defense and Proteasomal Activity in Fertile Men with High Seminal ROS Levels. Int. J. Mol. Sci. 2019, 20, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanta, L.; Agarwal, A.; Swain, N.; Sharma, R.; Gopalan, B.; Esteves, S.C.; Durairajanayagam, D.; Sabanegh, E. Proteomic Signatures of Sperm Mitochondria in Varicocele: Clinical Use as Biomarkers of Varicocele Associated Infertility. J. Urol. 2018, 200, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Brown, K.; Hirschey, M.D.; Verdin, E.; Chen, D. Calorie restriction reduces oxidative stress by SIRT3-mediated SOD2 activation. Cell Metab. 2010, 12, 662–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, R.; Coleman, M.C.; Pennington, J.D.; Ozden, O.; Park, S.-H.; Jiang, H.; Kim, H.-S.; Flynn, C.R.; Hill, S.; McDonald, W.H. Sirt3-mediated deacetylation of evolutionarily conserved lysine 122 regulates MnSOD activity in response to stress. Mol. Cell 2010, 40, 893–904. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, J.; Lin, Y.; Lei, Q.; Guan, K.L.; Zhao, S.; Xiong, Y. Tumour suppressor SIRT3 deacetylates and activates manganese superoxide dismutase to scavenge ROS. EMBO Rep. 2011, 12, 534–541. [Google Scholar] [CrossRef]
- O’Flaherty, C. Peroxiredoxins: Hidden players in the antioxidant defence of human spermatozoa. Basic Clin. Androl. 2014, 24, 4. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.; Sharma, R.; Agarwal, A. Proteomic analysis of mature and immature ejaculated spermatozoa from fertile men. Asian J. Androl. 2016, 18, 735–746. [Google Scholar]
- Parmigiani, R.B.; Xu, W.S.; Venta-Perez, G.; Erdjument-Bromage, H.; Yaneva, M.; Tempst, P.; Marks, P.A. HDAC6 is a specific deacetylase of peroxiredoxins and is involved in redox regulation. Proc. Natl. Acad. Sci. USA 2008, 105, 9633–9638. [Google Scholar] [CrossRef] [Green Version]
- Swain, N.; Samanta, L.; Agarwal, A.; Kumar, S.; Dixit, A.; Gopalan, B.; Durairajanayagam, D.; Sharma, R.; Pushparaj, P.N.; Baskaran, S. Aberrant Upregulation of Compensatory Redox Molecular Machines May Contribute to Sperm Dysfunction in Infertile Men with Unilateral Varicocele: A Proteomic Insight. Antioxid. Redox Signal. 2019, 32, 504–521. [Google Scholar] [CrossRef]
- Nowicka-Bauer, K.; Lepczynski, A.; Ozgo, M.; Kamieniczna, M.; Fraczek, M.; Stanski, L.; Olszewska, M.; Malcher, A.; Skrzypczak, W.; Kurpisz, M.K. Sperm mitochondrial dysfunction and oxidative stress as possible reasons for isolated asthenozoospermia. J. Physiol. Pharmacol. An Off. J. Pol. Physiol. Soc. 2018, 69. [Google Scholar] [CrossRef]
- Cimen, H.; Han, M.J.; Yang, Y.; Tong, Q.; Koc, H.; Koc, E.C. Regulation of succinate dehydrogenase activity by SIRT3 in mammalian mitochondria. Biochemistry 2010, 49, 304–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finley, L.W.; Haas, W.; Desquiret-Dumas, V.; Wallace, D.C.; Procaccio, V.; Gygi, S.P.; Haigis, M.C. Succinate dehydrogenase is a direct target of sirtuin 3 deacetylase activity. PLoS ONE 2011, 6, e23295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Fang, C.; Zong, N.C.; Liem, D.A.; Cadeiras, M.; Scruggs, S.B.; Yu, H.; Kim, A.K.; Yang, P.; Deng, M.; et al. Regulation of acetylation restores proteolytic function of diseased myocardium in mouse and human. Mol. Cell. Proteom. MCP 2013, 12, 3793–3802. [Google Scholar] [CrossRef] [Green Version]
- Kerns, K.; Morales, P.; Sutovsky, P. Regulation of sperm capacitation by the 26S proteasome: An emerging new paradigm in spermatology. Biol. Reprod. 2016, 94, 1–9. [Google Scholar] [CrossRef]
- Zigo, M.; Manaskova-Postlerova, P.; Jonakova, V.; Kerns, K.; Sutovsky, P. Compartmentalization of the proteasome-interacting proteins during sperm capacitation. Sci. Rep. 2019, 9, 12583. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, D.; Gygi, S.P. An iterative statistical approach to the identification of protein phosphorylation motifs from large-scale data sets. Nat. Biotechnol. 2005, 23, 1391–1398. [Google Scholar] [CrossRef]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics (Oxford, England) 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Narushima, Y.; Kozuka-Hata, H.; Koyama-Nasu, R.; Tsumoto, K.; Inoue, J.; Akiyama, T.; Oyama, M. Integrative Network Analysis Combined with Quantitative Phosphoproteomics Reveals Transforming Growth Factor-beta Receptor type-2 (TGFBR2) as a Novel Regulator of Glioblastoma Stem Cell Properties. Mol. Cell. Proteom. MCP 2016, 15, 1017–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panner Selvam, M.K.; Agarwal, A.; Dias, T.R.; Martins, A.D.; Samanta, L. Presence of Round Cells Proteins do not Interfere with Identification of Human Sperm Proteins from Frozen Semen Samples by LC-MS/MS. Int. J. Mol. Sci. 2019, 20, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Protein Name | Accession Number | Abundance | NSAF Ratio (UV vs. BV) | |
|---|---|---|---|---|---|
| UV | BV | ||||
| ACTB | actin, cytoplasmic 1 | 4501885 | H | H | 0.55 |
| EEF1A1 | elongation factor 1-alpha 1 | 4503471 | H | H | 0.56 |
| FASN | fatty acid synthase | 41872631 | H | H | 0.59 |
| HSPA5 | 78 glucose-regulated protein precursor | 16507237 | H | H | 0.56 |
| HSP90AA1 | heat shock protein HSP 90-alpha isoform 1 | 153792590 | H | H | 0.32 |
| MYH9 | myosin-9 | 12667788 | H | H | 0.4 |
| PKM | pyruvate kinase isozymes M1/M2 isoform c | 332164775 | H | H | 0.63 |
| TUBA3C | tubulin alpha-3C/D chain | 17921993 | H | H | 0.48 |
| TUBB4B | tubulin beta-4B chain | 5174735 | H | H | 0.41 |
| ACAT1 | acetyl-CoA acetyltransferase, mitochondrial precursor (involved in fatty acid metabolism and ketone body formation not lysine-acetylation of proteins) | 4557237 | L | H | 0.19 |
| P4HB | protein disulfide-isomerase precursor | 20070125 | L | H | 0.31 |
| ALDOA | fructose-bisphosphate aldolase A isoform 2 | 342187211 | M | H | 0.59 |
| ANXA1 | annexin A1 | 4502101 | M | H | 0.46 |
| ANXA2 | annexin A2 isoform 1 | 50845388 | M | H | 0.21 |
| ANXA5 | annexin A5 | 4502107 | M | H | 0.22 |
| ANXA6 | annexin A6 isoform 1 | 71773329 | M | H | 0.18 |
| ATP5A1 | ATP synthase subunit alpha, mitochondrial precursor | 4757810 | M | H | 0.58 |
| ATP5B | ATP synthase subunit beta, mitochondrial precursor | 32189394 | M | H | 0.3 |
| PDIA3 | protein disulfide-isomerase A3 precursor | 21361657 | M | H | 0.26 |
| HK1 | hexokinase-1 isoform HKI-ta/tb | 15991831 | M | H | 0.5 |
| HSPA8 | heat shock cognate 71 protein isoform 1 | 5729877 | M | H | 0.37 |
| HSP90AB1 | heat shock protein HSP 90-beta | 20149594 | M | H | 0.42 |
| LCP1 | plastin-2 | 167614506 | M | H | 0.65 |
| HSP90B1 | endoplasmin precursor | 4507677 | M | H | 0.27 |
| RUVBL1 | ruvB-like 1 | 4506753 | M | H | 0.55 |
| HYOU1 | hypoxia up-regulated protein 1 precursor | 5453832 | M | H | 0.17 |
| CCT2 | T-complex protein 1 subunit beta isoform 1 | 5453603 | M | H | 0.55 |
| RUVBL2 | ruvB-like 2 | 5730023 | M | H | 0.37 |
| PHGDH | D-3-phosphoglycerate dehydrogenase | 23308577 | M | H | 0.57 |
| CANX | calnexin precursor | 10716563 | VL | H | 0.08 |
| AKR1B1 | aldose reductase | 4502049 | VL | L | 0.12 |
| PSMD13 | 26S proteasome non-ATPase regulatory subunit 13 isoform 1 | 157502193 | VL | L | 0.05 |
| Gene Symbol | Protein Name | Accession Number | Abundance | NSAF Ratio (UV vs. BV) | |
|---|---|---|---|---|---|
| UV | BV | ||||
| ENO1 | alpha-enolase isoform 1 | 4503571 | H | H | 2.05 |
| MDH2 | malate dehydrogenase, mitochondrial precursor | 21735621 | H | H | 3.91 |
| ACADVL | very long-chain specific acyl-CoA dehydrogenase, mitochondrial isoform 2 precursor | 76496475 | H | L | 10.53 |
| PARK7 | protein DJ-1 | 31543380 | L | L | 3.78 |
| ACADM | medium-chain specific acyl-CoA dehydrogenase, mitochondrial isoform b precursor | 187960098 | M | L | 6.83 |
| ALDH9A1 | 4-trimethylaminobutyraldehyde dehydrogenase | 115387104 | M | L | 12.28 |
| CALM1 | calmodulin | 58218968 | M | L | 5.22 |
| cpt2 | carnitine O-palmitoyltransferase 2, mitochondrial precursor | 4503023 | M | L | 2.24 |
| HSPA9 | stress-70 protein, mitochondrial precursor | 24234688 | M | L | 2.29 |
| IPO5 | importin-5 | 24797086 | M | L | 4.17 |
| PSMB1 | proteasome subunit beta type-1 | 4506193 | M | L | 6.66 |
| SOD2 | superoxide dismutase [Mn], mitochondrial isoform A precursor | 67782305 | M | L | 10.51 |
| DNPEP | aspartyl aminopeptidase | 156416028 | M | L | 5.45 |
| HIST1H2AA | histone H2A type 1-A | 25092737 | M | L | 3.86 |
| HIST1H2BA | histone H2B type 1-A | 24586679 | M | L | 2.40 |
| DLD | dihydrolipoyl dehydrogenase, mitochondrial precursor | 91199540 | H | M | 10.28 |
| FLNB | filamin-B isoform 2 | 105990514 | H | M | 2.03 |
| GPI | glucose-6-phosphate isomerase isoform 2 | 18201905 | H | M | 8.39 |
| SERPINB6 | serpin B6 | 41152086 | H | M | 4.68 |
| LAP3 | cytosol aminopeptidase | 41393561 | H | M | 8.54 |
| DECR1 | 2,4-dienoyl-CoA reductase, mitochondrial precursor | 4503301 | M | M | 2.65 |
| DLAT | dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial precursor | 31711992 | M | M | 2.36 |
| DLST | dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex, mitochondrial isoform 1 precursor | 19923748 | M | M | 2.91 |
| ETFA | electron transfer flavoprotein subunit alpha, mitochondrial isoform a | 4503607 | M | M | 2.77 |
| GSR | glutathione reductase, mitochondrial isoform 1 precursor | 50301238 | M | M | 6.39 |
| PRDX1 | peroxiredoxin-1 | 320461711 | M | M | 2.04 |
| SDHA | succinate dehydrogenase [ubiquinone] flavoprotein subunit, mitochondrial | 156416003 | M | M | 2.35 |
| APMAP | adipocyte plasma membrane-associated protein | 24308201 | M | M | 2.45 |
| ASRGL1 | l-asparaginase | 145275200 | M | M | 1.99 |
| GOT2 | aspartate aminotransferase, mitochondrial precursor | 73486658 | H | VL | 31.16 |
| CLGN | calmegin precursor | 4758004 | L | VL | 3.77 |
| FTH1 | ferritin heavy chain | 56682959 | L | VL | 6.51 |
| HAGH | hydroxyacylglutathione hydrolase, mitochondrial isoform 1 precursor | 94538322 | L | VL | 8.18 |
| MDH1 | malate dehydrogenase, cytoplasmic isoform 1 | 312283701 | L | VL | 5.46 |
| MIF | macrophage migration inhibitory factor | 4505185 | L | VL | 6.98 |
| PPP3CA | serine/threonine-protein phosphatase 2B catalytic subunit alpha isoform isoform 2 | 194688147 | L | VL | 10.66 |
| RPL5 | 60S ribosomal protein L5 | 14591909 | L | VL | 8.34 |
| SUCLA2 | succinyl-CoA ligase [ADP-forming] subunit beta, mitochondrial precursor | 11321583 | L | VL | 46.32 |
| LACTB2 | beta-lactamase-like protein 2 | 7705793 | L | VL | 7.36 |
| C1QBP | complement component 1 Q subcomponent-binding protein, mitochondrial precursor | 4502491 | M | VL | 7.24 |
| CAPN1 | calpain-1 catalytic subunit | 311893363 | M | VL | 5.51 |
| FBP1 | fructose-1,6-bisphosphatase 1 | 189083692 | M | VL | 26.30 |
| SOD1 | superoxide dismutase [Cu-Zn] | 4507149 | M | VL | 5.68 |
| TKT | transketolase | 205277463 | M | VL | 11.17 |
| Gene Symbol | Protein Name | Accession Number | Abundance | Expression |
|---|---|---|---|---|
| GSS | glutathione synthetase | 4504169 | L | Unique in UV |
| PPP5C | serine/threonine-protein phosphatase 5 isoform 1 | 5453958 | L | Unique in UV |
| TALDO1 | transaldolase | 5803187 | L | Unique in UV |
| GNPDA1 | glucosamine-6-phosphate isomerase 1 | 13027378 | L | Unique in UV |
| CAT | catalase | 4557014 | M | Unique in UV |
| PNP | purine nucleoside phosphorylase | 157168362 | M | Unique in UV |
| SELENB P1 | selenium-binding protein 1 | 16306550 | M | Unique in UV |
| ACAA2 | 3-ketoacyl-CoA thiolase, mitochondrial | 167614485 | M | Unique in UV |
| ACTC1 | actin, alpha cardiac muscle 1 proprotein | 4885049 | L | Unique in BV |
| ATP5C1 | ATP synthase subunit gamma, mitochondrial isoform L (liver) precursor | 50345988 | L | Unique in BV |
| HMOX2 | heme oxygenase 2 | 8051608 | L | Unique in BV |
| PFN2 | profilin-2 isoform a | 16753215 | L | Unique in BV |
| PHB | prohibitin | 4505773 | L | Unique in BV |
| VDAC1 | voltage-dependent anion-selective channel protein 1 | 4507879 | L | Unique in BV |
| YWHAB | 14-3-3 protein beta/alpha | 21328448 | L | Unique in BV |
| YWHAG | 14-3-3 protein gamma | 21464101 | L | Unique in BV |
| DNAJA2 | dnaJ homolog subfamily A member 2 | 5031741 | L | Unique in BV |
| STOML2 | stomatin-like protein 2 | 7305503 | L | Unique in BV |
| CYCS | cytochrome c | 11128019 | L | Unique in BV |
| MTPN | myotrophin | 21956645 | L | Unique in BV |
| 7-Sep | septin-7 isoform 2 | 148352329 | VL | Unique in BV |
| TAGLN2 | transgelin-2 | 4507357 | VL | Unique in BV |
| Sperm Function | DEPs | Unilateral Varicocele | Bilateral Varicocele | ||
|---|---|---|---|---|---|
| Abundance | Expression | Abundance | Expression | ||
| Fertilization and acrosome reaction | HIST1H2B | Moderate | OE | Low | UE |
| Mitochondrial dysfunction and oxidative stress | SDHA | Moderate | OE | Moderate | UE |
| PRDX1 | Moderate | OE | Moderate | UE | |
| SOD1 | Moderate | OE | Very low | UE | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panner Selvam, M.K.; Samanta, L.; Agarwal, A. Functional Analysis of Differentially Expressed Acetylated Spermatozoal Proteins in Infertile Men with Unilateral and Bilateral Varicocele. Int. J. Mol. Sci. 2020, 21, 3155. https://doi.org/10.3390/ijms21093155
Panner Selvam MK, Samanta L, Agarwal A. Functional Analysis of Differentially Expressed Acetylated Spermatozoal Proteins in Infertile Men with Unilateral and Bilateral Varicocele. International Journal of Molecular Sciences. 2020; 21(9):3155. https://doi.org/10.3390/ijms21093155
Chicago/Turabian StylePanner Selvam, Manesh Kumar, Luna Samanta, and Ashok Agarwal. 2020. "Functional Analysis of Differentially Expressed Acetylated Spermatozoal Proteins in Infertile Men with Unilateral and Bilateral Varicocele" International Journal of Molecular Sciences 21, no. 9: 3155. https://doi.org/10.3390/ijms21093155