Functional Lipids in Autoimmune Inflammatory Diseases


1
Department of Health Sciences, Università degli Studi di Milano, 20142 Milan, Italy
2
Department of Pharmaceutical Sciences, Università degli Studi di Milano, 20133 Milan, Italy
3
State Key Laboratory of Virology, Wuhan Institute of Virology, Chinese Academy of Sciences, Wuhan 430071, China
4
Istituto di Scienze e Tecnologie Chimiche “Giulio Natta”, Consiglio Nazionale delle Ricerche, 20131 Milan, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(9), 3074; https://doi.org/10.3390/ijms21093074
Submission received: 17 March 2020
/
Revised: 22 April 2020
/
Accepted: 24 April 2020
/
Published: 27 April 2020
(This article belongs to the Special Issue Applications and Biotechnological Synthesis of Functional Lipids)
Abstract
:Lipids are apolar small molecules known not only as components of cell membranes but also, in recent literature, as modulators of different biological functions. Herein, we focused on the bioactive lipids that can influence the immune responses and inflammatory processes regulating vascular hyperreactivity, pain, leukocyte trafficking, and clearance. In the case of excessive pro-inflammatory lipid activity, these lipids also contribute to the transition from acute to chronic inflammation. Based on their biochemical function, these lipids can be divided into different families, including eicosanoids, specialized pro-resolving mediators, lysoglycerophospholipids, sphingolipids, and endocannabinoids. These bioactive lipids are involved in all phases of the inflammatory process and the pathophysiology of different chronic autoimmune diseases such as rheumatoid arthritis, multiple sclerosis, type-1 diabetes, and systemic lupus erythematosus.
1. Introduction: Lipids and Inflammation
Inflammation is an immune response that occurs following infections, cellular insults, or tissue insults and that can be spontaneously exhausted after the elimination of the damage. It includes an extensive network of cellular and molecular processes, in which a multitude of preformed or newly synthesized mediators are arranged to obtain specific responses. However, protracted and uncontrolled immune responses can lead to chronic inflammation, irreparable tissue damages, and chronic diseases. Uncontrolled immune responses also occur in many common autoimmune diseases, including type 1 diabetes, multiple sclerosis (MS), psoriasis, inflammatory bowel disease (IBD), and Grave’s disease.
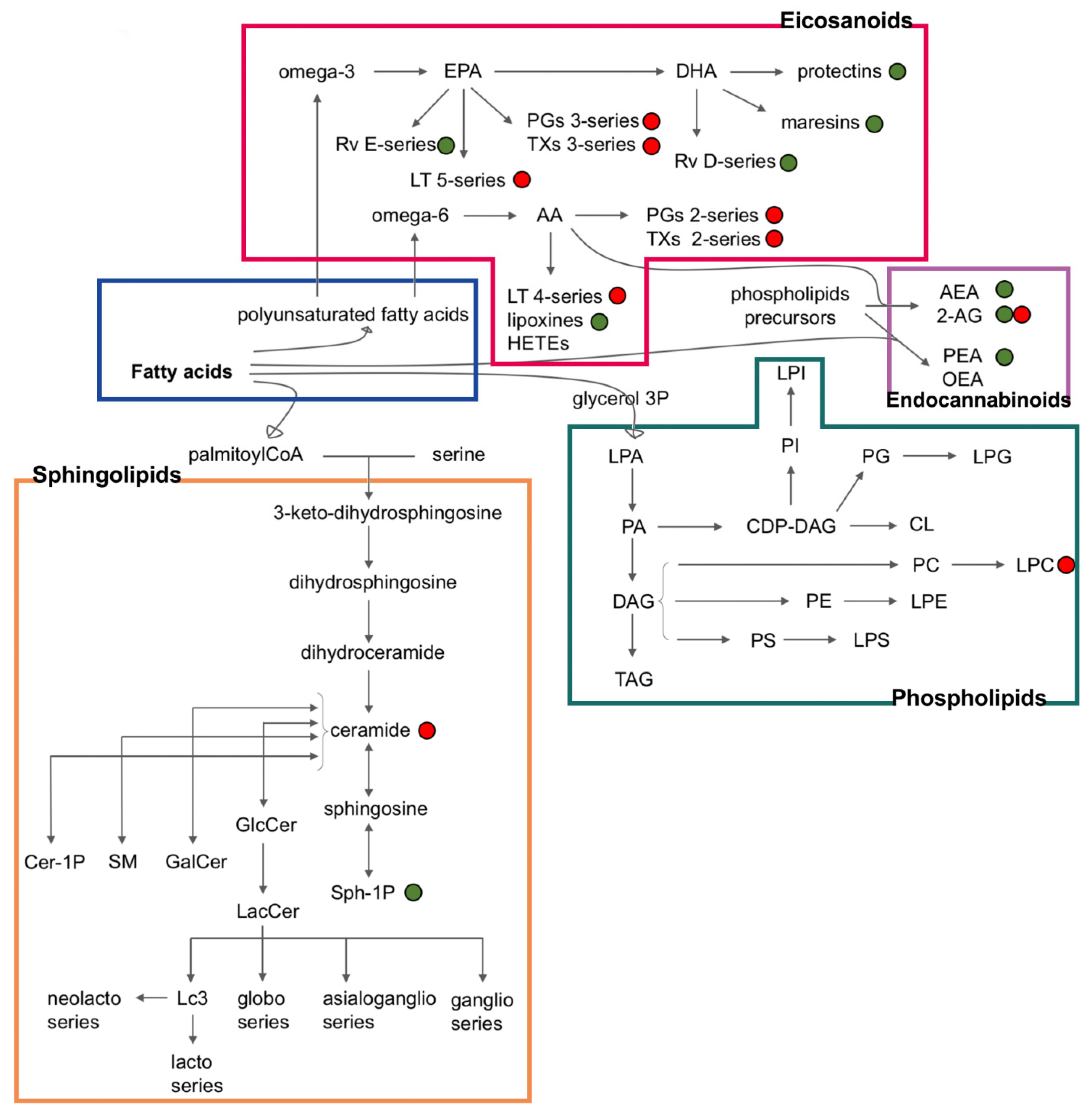
Endogenous bioactive lipids play a pivotal role in inflammatory processes and in triggering, coordinating, and confining immunity by regulating hypervascular reactivity, pain, leukocyte trafficking, and clearance [1,2,3,4]. Moreover, the accrual of inflammatory lipids can contribute to the transition from acute to chronic inflammation (Figure 1) [1].
Bioactive lipids can be divided into different families depending on their structure or biochemical function: eicosanoids, specialized pro-resolving mediators, lysoglycerophospholipids, sphingolipids, and endocannabinoids [3].
2. Lipids Involved in Inflammatory Responses
2.1. Eicosanoids
This well-known family of bioactive lipids includes a wide range of derivatives of fatty acids with a 20-carbon chain, such as arachidonic acid (AA), eicosapentaenoic acid (EPA), and docosahexaenoic acid (DHA). Eicosanoids are prevalently produced from AA, which can be released from membrane phospholipids primarily by phospholipase A2 and secondarily by phospholipase C. Three different enzymes drive the biosynthesis of eicosanoids: (1) cyclooxygenases 1 and 2 (COX-1/2) produce the class of prostanoids that comprises prostaglandins (PGs), prostacyclins, and thromboxanes (TXs); (2) lipoxygenases (5/12/15-LOX) generate leukotrienes (LTs), hydroxy-eicosatetraenoids, and lipoxins; and (3) P450 epoxygenase generates hydroxy-eicosatetraenoic acids (HETEs) and epoxy-eicosatrienoids [38,39]. Eicosanoids are divided into omega-6 and omega-3 families depending on the position of unsaturations of their precursor. The pro-inflammatory omega-6 family is derived from AA, whereas the anti-inflammatory omega-3 family is derived from EPA and DHA.
Eicosanoids are well-recognized for initiating inflammation and for controlling vascular tone, platelet aggregation, pain perception, ovulation, and embryo implantation. In general, the omega-6 eicosanoids are pro-inflammatory, whereas the omega-3 eicosanoids do not promote the inflammatory events and can also be anti-inflammatory and pro-resolving [40,41].
In particular, five mechanisms are related to inflammation induced by PGs: (1) boosting the release of pro-inflammatory cytokines [41,42]; (2) enhancing the innate immunity [43]; (3) activating the T-helper cells, TH1-related TH17 [44]; (4) contributing to leukocyte recruitment [41]; and (5) increasing the expression of pro-inflammatory genes such as NF-κB [45].
The leading roles of LTs in acute inflammation are to induce edema and to maintain an ongoing inflammatory status by acting as chemoattractants for neutrophils, macrophages, eosinophils, and TH17 lymphocytes. TXs and prostacyclins have prevalent functions of vasoconstriction and vasodilatation, respectively [1,3,46]. Epoxyeicosatrienoic acids (EETs) are synthesized from AA by cytochrome P450 epoxygenases. They modulate vasorelaxation, anti-inflammation—by the suppression of NF-kB activation—and fibrinolysis. They can be converted into the less active dihydroxyeicosatrienoic acids (DHETs) by soluble epoxide hydrolase [47].
2.2. Sphingolipids
Sphingolipids are amino alcohols synthesized de novo from the condensation of serine and acyl-CoA. They are involved in a multitude of pathophysiological functions [48], such as regulation of apoptosis [49,50], proliferation [51], differentiation [52], autophagy [53,54], invasiveness [55,56], modification of signaling cascade [57,58], and mediation of inflammatory responses by cytokines [59]. In particular, ceramide and sphingosine promote apoptosis via different pathways, which involves the catalytic activity of Bcl-2, protein kinase C, protein phosphatases 1–2, and proteases. In contrast with ceramide, which is predominantly pro-apoptotic [60], sphingosine-1-phosphate (S1P) is mainly an anti-apoptotic messenger [61] that can modulate mitogenesis, cell migration, cytoskeletal rearrangement, and angiogenesis [54,62]. The phosphate forms of sphingolipids are notably related to inflammation, where S1P acts on either COX-2 or NF-kB and ceramide-1P acts on phospholipases A2 [63,64].
2.3. Endocannabinoids
Endocannabinoids are a group of molecules that can bind and activate cannabinoid receptors (CB1 and CB2) in the same way as the tetrahydrocannabinol (THC), the main psychoactive component of Cannabis sativa. These lipid mediators share a cannabimimetic action but have different chemical structures, which have chemical functionalities such as amides, esters, or ethers of long-chain polyunsaturated fatty acids. The most studied molecules of this class are anandamide (AEA), 2-arachidonoilglycerol (2-AG), 2-AG-ether, O-arachidonoylethanolamine, arachidonoyldopamine, and palmitoylethanolamide (PEA). These are known as potent immunoregulatory, endocrine, and inflammatory modulators [65,66]. In particular, AEA and PEA have anti-inflammatory properties [67,68], whereas 2-AG has both pro- and anti-inflammatory properties [69,70,71]. Thus, some dysfunctions in tissue homeostasis and chronic inflammatory status were related to changes in the concentrations, metabolism, and receptors of endocannabinoids [72].
2.4. Lysoglycerophospholipids
These lipids are asymmetrically distributed in the plasma membrane. In particular, phosphatidylethanolamine, phosphatidylserine, and phosphatidylinositol are mainly located in the inner membrane leaflet, whereas phosphatidylcholine is located in the outer membrane leaflet. Lysoglycerophospholipids contain glycerol backbones linked to two long fatty acid chains and a polar head comprising a phosphate modified with ethanolamine, choline, inositol, or serine [73]. The most active forms of lysoglycerophospholipids derive from the hydrolytic removal of one of the fatty acids from the membrane phospholipids. This hydrolysis process affords lysophosphatidylcholines (LPC), lysophosphatidylinositols (LPI), lysophosphatidylethanolamines (LPE), lysophosphatidylserines (LPS), and lysophosphatidic acid (LPA). Lysophospholipids act as signal molecules in the inflammatory cascade and in the essential processes of cellular and tissue life, such as plasma membrane shaping, cell growth, and cell death [33]. LPC and LPA modulate the immune response by controlling the distribution, trafficking, and activation of immune cells [33,74]. Therefore, they have been linked with different inflammatory diseases such as diabetes, obesity [75], atherosclerosis, cancer [76], and rheumatoid arthritis (RA) [77].
3. The Connection between Lipids and Some Related Inflammatory Diseases
Uncontrolled chronic inflammation occurs in autoimmune inflammatory diseases (AIDs) and is induced by the overreaction of the immune system to the organs or tissues of the body. The lipid-mediated inflammation and pathogenesis of AIDs have been investigated, and the results are shown in Table 1 and summarized as follows.
3.1. Rheumatoid Arthritis
RA is a systemic AID of the joints that is characterized by excessive synovial and joint inflammation, which contribute to the deterioration of bone and cartilage. Eicosanoids are implicated in the development of synovitis and the disintegration of the joints in inflammatory arthritis. The pathogenesis of rheumatoid and psoriatic arthritis was proposed to arise from an imbalanced regulation of pro- and anti-inflammatory eicosanoids [5,78]. In particular, the anti-inflammatory EPA-derived eicosanoids, including 11-HEPE, 12-HEPE, and 15-HEPE, were up-regulated. This increase was suggested to occur to equalize the inflammation induced by AA-derived eicosanoids [5]. On the contrary, another study demonstrated a decrease in some pro-resolving lipid mediators in the circulating plasma [28]. 15-(S)-HETE and 13-HODE stimulated the expression of placenta growth factor, which plays an essential role in RA. The up-regulation of placenta growth factor was related to COX-2 activity as well as PGE2 levels and membrane-bound receptors expression in eicosanoids (EP1, EP2, EP3, and EP4) [17,79]. In a murine model affected by RA, the levels of the different molecules (e.g., PGE2, PGD2, PGF2α, and TXB2) related to COX and LOX pathways [6,78] were significantly higher than the control. In contrast, the levels of metabolites 5-HpETE and LTD4 were lower than the control [6]. In addition, the early phase of RA was characterized by the synthesis of PGD2 within the joint, with the peak of expression being reached in the later stages. Moreover, serum PGD2 levels increased throughout the arthritic process, thus taking part in the anti-inflammation activity [80]. In RA, oxidative stress is accompanied by inflammation, and these were monitored by the increase in the plasma levels of 8-iso-prostaglandin F2α and 15-keto-dihydro-PGF(2α) [81,82].
The pathophysiology of RA comprises synovial inflammation, hyperplasia, and cartilage degradation; moreover, it could be linked to the endocannabinoid system. The activation of CB2 can alleviate the disease by inhibiting not only the development of autoantibodies, pro-inflammatory cytokines, and matrix metalloproteinases but also bone degradation, T-cell-mediated immune response, and fibroblast-like proliferation. Moreover, in the synovial fluid of patients with RA, the levels of AEA and 2-AG were found to increase and that of PEA was found to decrease, suggesting the prominent functional role of this pathway [32,66].
With reference to inflammatory lysophospholipids, the hydrolysis of circulating LPC contributed to the accumulation of the local production of LPA, which in turn amplified inflammation in synergy with TNF-α [33,34].
Ceramide and the activity of acid sphingomyelinase, which catalyzes the hydrolysis of sphingomyelin to ceramide, have been implicated in inflammatory arthritis. The inhibition of the activity of acid sphingomyelinase by genetic or pharmacological tools, such as amitriptyline, reduced the disease manifestations and levels of pro-inflammatory cytokines in a murine model [83]. A characteristic feature of RA is the disease fluctuation over the day, demonstrated by the changing symptoms and circulating markers of the inflammatory process. This joint inflammation oscillation was surprisingly linked to ceramide synthesis [84]. Furthermore, the sphingosine kinase-1 activity (SPHK1, which catalyzes the phosphorylation of sphingosine into S1P) increased at the sites of acute inflammation in both the murine model and patients. An increase in the production of S1P was hypothesized to cause the persistence of T-helper cells in the inflammation sites, contributing to the abnormal immune responses [35,36]. The expression of receptor 3 of S1P was associated with the development of autoimmunity as well as the increased downstream signaling and the production of cytokine IL-6 [85]. In contrast, the down-regulation of receptor 1 of S1P contributed to the regression of RA [86]. Elsewhere, it was postulated that acting on the mTOR pathway, S1P signaling could have a dual role: under physiological conditions, it maintains continuous bone turnover but leads to the pathogenesis of bone deformities during inflammation [87].
3.2. Type 1 Diabetes
Type 1 diabetes is characterized by the autoimmune-mediated destruction of pancreatic β-cells. The plasma lipid profile of patients with type 1 diabetes and murine models was characterized by the increase in the long-chain polyunsaturated triglycerides and the decreases in the long-chain lysophospholipids and cholesterol esters [88]. The corroborated hypothesis was the increase in the remodeling of both circulating lipoprotein and pro-inflammatory status in type 1 diabetes [88,89,90,91,92]. Notably, the levels of AA and AA-derived eicosanoids—such as thromboxane A2 (TXA2), leukotriene B4 (LTB4), PGD2, PGE2, 11-HETE, 12-HETE, 15-HETE, and 12-oxo-ETE—also decreased in a non-obese diabetic mouse, highlighting a reduced state of systemic inflammation [37].
The endocannabinoid system was shown to play a relevant role in the maintenance of effective immune responses in the gut, which is perpetually exposed to pathogenic insults [93]. Mononuclear phagocytes contributed to maintaining the equilibrium between inflammation and tolerance, which is a consequential state of hyporesponsiveness. In addition, AEA might protect against the development of autoimmune diabetes by modulating the immune-suppressive functions and the number of monocytes and macrophages [93].
When focusing on sphingolipidomics, an elevation in the levels of S1P was revealed in a cohort of patients with type 1 diabetes and in vivo models [7,8]. Other studies indicated that the pathogenesis of type 1 diabetes might be related to S1P pathways because it was involved in the disorders of T-cells migration and activation [94,95]. Moreover, in insulin-secreting cells, the low expression of endogenous S1P lyase contributed to the vulnerability of the cells to the toxicity brought by pro-inflammatory cytokines [96]. The role of S1P in inflammation and pathogenesis is not easy to understand. Autoimmune diabetes and especially its complications were associated with a dysmetabolism in the sphingolipids [7,9]. In particular, decreased plasma levels of very long-chain ceramide were associated with a significantly lower frequency of developing diabetes-related nephropathy and macroalbuminuria. Moreover, the reduction of sphingolipids containing fatty acid C24:1 was demonstrated and associated with a reduction in cardio- and neuro-protection [7,9].
3.3. Systemic Lupus Erythematosus
Systemic lupus erythematosus (SLE) is a disease of unknown etiology. SLE is more frequent in women than in men, and its symptomatology includes non-erosive and non-deforming arthritis, cutaneous rash, vasculitis, and other systemic manifestations [97,98]. Changes in the lipid composition in patients with SLE might be attributable to oxidation damages, mTORC1-dependent mitochondrial dysfunction [99], and cell death. In the pathophysiology of SLE, cell death concurred with the accrual in the production of autoantigens and autoantibodies [100]. Some alterations such as variations in the levels of plasmalogens and fatty acids could be associated with the changes in the lipid composition of the membrane of lymphocytes and with the deregulation of the immune system, including the abnormal recognition of autoantigens and enhanced production of antibodies. The uncontrolled production of reactive oxygen species (ROS) by neutrophils appeared at the onset and in the progression of SLE, thus causing increased oxidation in membrane lipids and the formation of other products such as isoprostanes [101,102]. To exemplify, the reaction of ROS with membrane lipids could lead to the formation of toxic lipidic hydroperoxides and their degradation products, which can further react with proteins and modify their structure and function [102]. The overproduction of ROS and lipid peroxidation products were related to the inflammatory status and therefore considered as a factor that, indeed, characterizes the disease [103].
Another piece of evidence is the high levels of glycosphingolipids in T lymphocyte membranes [104]. The disease did not modify the total content of circulating sphingolipids. However, it demonstrated an altered length of the FAs incorporated in SM and ceramide, with an increase of long-chain FAs and a decrease of very long-chain FAs [15]. Moreover, the plasma and serum levels of ceramide C24:1 increased owing to SLE renal complications. Therefore, ceramide C24:1 could be considered as a potential biomarker of lupus nephritis [10]. In particular, the levels of circulating ceramides and hexosylceramides were increased and sphingoid bases were decreased in SLE. These levels were associated with disease activity, and accordingly, they were normalized after immunosuppressive treatment [11]. Although the alteration in S1P expression was observed in different studies, its effects need to be further elucidated [10,11,105,106,107]. Sphingolipids profile modification could also occur in the vascular complication related to SLE, namely atherosclerosis [12].
Endocannabinoid modulation was demonstrated by higher 2-AG levels in patients with SLE compared with healthy subjects. The 2-AG increment was associated with disease regression and supported the hypothesis of its protective action against the pathogenesis of SLE [13]. However, another cohort of patients did not exhibit this biochemical alteration; therefore, 2-AG levels could be associated with the disease manifestation [108].
3.4. Inflammatory Bowel Diseases
The term IBD is comprehensive of disabling, chronic inflammatory processes directed against intestinal mucosae, such as ulcerative colitis and Crohn’s disease (CD). IBDs are characterized by mucosa cell necrosis or the ulceration and infiltration of neutrophils into lesions. The disruption of epithelial barrier function observed in patients with IBD has been traditionally attributed to cytokines, but some studies have shed light on the role of PGE2 in paracellular regulation and mucosa impairment [16,109]. These showed that PGE2 binds receptors EP1–EP4 and then acts, via a Ca2+-mediated pathway, on myosin light chain kinase. The activity of the kinase included changing the transmembrane distribution of the tight junction proteins and the peri-junctional actin rings [16]. An increase in PGE2 mucosa levels was also demonstrated in Rodríguez-Lagunas et al.’s studies [16]. Moreover, the epithelial cells displayed the deregulation of the balance of the local levels of eicosanoids and endocannabinoids. An increase in the levels of HxA3, a pro-inflammatory and neutrophil chemoattractant eicosanoid, as well as a concomitant decrease in the anti-inflammatory endocannabinoids, were shown [18]. A common complication of CD is fibrosis, which includes the excessive deposition of extracellular matrix and the obstruction of the gut lumen. This complication was related to a differential DNA methylation that causes a decrease in the expression of mRNA encoding for prostacyclin synthase and an increase in that for prostaglandin D2 [19]. Plasma levels in IBD in the murine model showed impairment in the profile of eicosanoids, which promote inflammation and may induce carcinogenesis. In particular, in a murine model (IL-10(−/−)), EETs and DHETs underwent a decrease compared with the wild type [20].
In the gastrointestinal tract, endocannabinoids have been proposed to control the muscular propulsion, but they can also control several pathological functions. Studies have shown [21,110,111] that in animal models as well as in patients with ulcerative colitis, colon inflammation is accompanied by an increment of anandamide but not 2-AG. These findings were registered in mouse colon and biopsy samples from patients, confirming a possible role of the protective action of anandamide and its possible therapeutic employment [21]. Another study revealed a change in the gene expression in the macrophages and lymphocytes of non-classical endocannabinoid receptors including GPR55 and MGL. These components played an essential role in the regulation of the immune response toward intestinal and systemic inflammation [110]. After inflammatory stimuli, cannabinoid receptors could be modulated in their intestinal localization. In particular, CB2 increased in colonic epithelial cells, and CB1 increased in enteric neurons and the endothelium [111].
3.5. Multiple Sclerosis
MS is an autoimmune disease of the central nervous system that causes progressive neurologic disability. It is characterized by chronic inflammation that damages myelin axons and the myelin sheath [25].
A targeted lipidomic study performed on the spinal cords of experimental autoimmune encephalomyelitis (EAE) highlighted a metabolic switch, which determined an increase of the PGE2 pathway and a decrease of the PGD2, PGI2, and 5-LO pathways. Eicosanoid levels in the spleen and plasma were also measured; however, significant fluctuations were not found. These changes, coupled with an elevation in the expression of PGE2 receptors (EP1, EP2, and EP4), were correlated with clinical symptoms [112,113]. Moreover, a decrease in the serum concentration of neuroprotective lysophospholipids in patients with MS was shown. This decrease was attributed both to an impaired spleen homing of T-cells and to the demoting remyelination. However, LPA might also trigger pro-inflammatory cellular responses depending on the binding receptor and the source [22]. Nevertheless, whether LPA can be used as a biomarker of MS remains unclear. Two studies investigated LPA levels in a cohort of patients (n = 20); although an elevation of LPA serum levels in the patients compared with healthy controls was demonstrated, the possible role of LPA in the disease progression was not elucidated [23,24].
The membrane lipid abnormalities in lymphocytes and monocytes were attributable to impaired membrane fluidity along with the disease progression [114]. The results from this study showed that the membrane lipids of patients with MS and control subjects have no significant differences. However, a correlation was demonstrated between membrane fluidity—measured by lipid composition (phospholipids, fatty acids, and cholesterol)—and disease progression, estimated by the functional system score [114].
During MS, S1P levels were elevated in the central nervous system cell lineages. The S1P level in the EAE mice model affected with autoimmune EAE spinal cord was approximately twice that in the wild type, consistent with astrogliosis. Notably, the S1P levels decreased in S1P1 conditional null mutants [26].
3.6. Graves’ Disease
Graves’ disease is an autoimmune thyroid disease that represents one of the most common causes of hyperthyroidism. In addition to the signs and symptoms of hyperthyroidism, Graves’ disease is accompanied by a typical orbitopathy called Graves’ orbitopathy (GO). In particular, the orbital tissue of patients with GO showed elevated levels of S1P compared with the control samples. S1P was hypothesized to act as a chemoattractant for T-cells during disease progression [27]. The initial binding of T-cells activated the orbital fibroblasts via CD40, which caused an augmentation of S1P levels [27]. In addition, the role of S1P on adipogenesis and fibrosis [29,30] and in pro-inflammatory responses has been demonstrated. More specifically, IL-1β enhanced the expression of S1P receptors and sphingosine kinase in GO orbital fibroblasts; this in turn increased the expression of other pro-inflammatory mediators, including ICAM-1, COX-2, and IL-6 proteins [31].
4. Anti-Inflammatory Lipids and their Therapeutic Potential
Some recent works have focused on the functions of the precursors of eicosanoids: omega-3 polyunsaturated fatty acids (ω-3 PUFAs), EPA, and DHA. However, these precursors were proposed as potential candidates for the prevention or even treatment of some AIDs, such as type 1 diabetes, RA, SLE, and MS. Many of the beneficial effects of ω-3 PUFAs could be assigned to their anti-inflammatory properties coupled with the regulation of mTOR activity [115,116,117]. In particular, the anti-inflammatory properties of marine-derived ω-3 PUFAs could be related to a change in the fatty acid composition of the cell membranes [118] along with a decrease in the levels of eicosanoids, cytokines, and adhesion molecules. Another evidence of the anti-inflammatory properties of ω-3 PUFAs was the increasing levels of pro-resolving mediators [118,119]. In clinical trials, pro-resolving mediators both reduced inflammation—for example, decreased LTB4 [120]—and stabilized advanced atherosclerotic plaques in patients with RA [121]. Moreover, some symptomatic benefits could be obtained by combining paracetamol with fish oil (rich in PUFAs). This combination provided superior suppression of inflammatory PGE2 synthesis [122]. Nevertheless, note that information on ω-3 PUFAs supplementation based on clinical trials might be difficult to interpret owing to the differences in dose, duration, and drug interactions [119]. In a murine model of type 1 diabetes, dietary treatment with ω-3 PUFAs reduced the incidence of autoimmunity in pancreatic islets, modulated the differentiation of Th- and T-regulatory cells, and decreased the levels of pro-inflammatory mediators such as IFN-γ, IL-17, IL-6, and TNF-α [123]. In a murine model of colitis, EPA and DHA supplementation caused a significant increase in the levels of some anti-inflammatory eicosanoids. However, this change was not sufficient to alleviate colitis [124]. A study suggested that micronized PEA may be considered in relapsing–remitting MS to reduce the cutaneous adverse effects related to the subcutaneous administration of interferon (IFN)-β1 [125]. Lipoxins, resolvins, and protectins co-administered with aspirin might be useful in various rheumatological conditions [97]. To exemplify, the combination of ω-3 with non-steroidal anti-inflammatory drugs generated bioactive lipids that could be used downstream by leukocytes to counteract inflammation propagation [126]. Some results suggested that the administration of PPAR ligands such as 15d-PGJ2 may be a novel therapeutic strategy for MS because PPAR ligands efficiently reduced the severity of the inflammation by reducing both the secretion of encephalitogenic T-cells and cytokines and the consequent demyelination [127]. Moreover, EETs could be considered as a novel therapeutic agent for rheumatic disorders because they promoted tissue regeneration along with the attenuation of bone loss and osteoclast activity [128].
The stabilized cyclic phosphatic acid 2-carba-cPA (obtained by replacing one of the cyclic phosphate oxygen molecules with a methylene group at the sn-2 position) as well as its precursor, cyclic phosphatic acid, protected the oligodendrocytes by reducing mitochondrial apoptosis. This suggested that the modulation of LPA could be relevant for the treatment of demyelinating conditions [129]. Endocannabinoids can also display anti-inflammatory effects in different AIDs; however, their pharmacological potential is still debated. The exogenous supplementation of either COX-2 inhibitors [130], cannabinoid receptor agonists [65,66], or endocannabinoid degradation inhibitors [65,130,131,132] increased the levels of anti-rheumatic N-acylethanolamines. In the same way, in some diseases, the modulation of S1P pathway could be accomplished indirectly and not using exogenous S1P. S1P seemed to have a central role in interfering with an uncontrolled immune system by altering lymphocyte recruitment and the cellular adhesion processes. Moreover, blocking the S1P receptor function demonstrated efficacy in the treatment of MS. Fingolimod, the first non-selective S1PR agonist, and siponimod (BAF312), a selective S1PR1 agonist, have been approved to reduce inflammation in the treatment of relapsing–remitting and secondary progressive MS. Other S1PR modulators such as ponesimod (ACT128800), ozanimod (RPC1063), ceralifimod (ONO-4641), GSK2018682, and MT-1303 have been promising to reduce inflammatory demyelination and are also under clinical trials for MS treatment [133,134,135,136,137,138,139,140,141,142].
5. Concluding Remarks and Possible Roles of Biotechnology in the Prevention and Therapy of Autoimmune Inflammatory Diseases (AIDs)
Numerous new studies have proved the role of different functional lipids in the treatment of several AIDs, which are increasingly prevalent health problems among the global population. Nowadays, the prevalence of diagnosed autoimmune conditions is 7%, 6%, and 5% in the U.S., E.U. (average of a group of countries), and China, respectively [143]. Thus, further studies are urgently needed to shed light on the mechanisms and therapies of these diseases so as to counteract them.
It has been recognized that ω-3 PUFAs have anti-inflammatory properties and their presence in nutrition contributes to the prevention of many inflammatory diseases, independently from the mechanism of action. Nevertheless, for the development of drugs based on ω-3 PUFA derivatives for the cure of a given inflammatory disease, in-depth knowledge of the mechanism of action of the disease itself is required. Therefore, the administration or inclusion of ω-3 PUFAs in human diet appears as the most natural way to reduce AIDs insurgence. In particular, the availability of ω-3 PUFAs in human diet could dramatically change their benefits.
Although out of the scope of this review, emphasizing that biotechnology, in particular metabolic engineering, is increasingly adopted for the production of ω-3 PUFAs from different microbial strains is important. Analogously, plants have also been genetically engineered to produce high amounts of DHA and EPA. These achievements would undoubtedly allow a broader diffusion of these fatty acids, thus helping to improve the general health conditions. Human health will particularly benefit if these molecules are included as an essential part of diet [144,145,146].
An ideal tool to test the role of different types of functional lipids might be microfluidics tools combined with biotechnological techniques [147,148]. Such tools have allowed human cells and histological tissues to be cultivated and treated under strictly controlled conditions. These approaches, even if they are only at the initial stages of development, have already been proven as a strategic system to provide new insights into several areas of medicine, especially therapeutic devices with the role of organoids. From a pharmacological point of view, biotechnology could positively contribute to the production of the intermediates involved in the regulation of inflammatory diseases. This review provides an overview of AIDs and aims to encourage scientists to develop engineered microorganisms for the large-scale production of selected functional lipids or their derivatives.
Funding
This research received no external funding.
Acknowledgments
MDC was supported by the Ph.D. program in Molecular and Translational Medicine by the Università degli Studi di Milano, Milan, Italy. We are also thankful for the financial support from the Chinese Academy of Sciences President’s International Fellowship Initiative (CAS-PIFI; No. 2020VBA0011).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AIDs | autoimmune inflammatory diseases |
| COX | cyclooxygenases |
| LOX | lipoxygenases |
| AA | arachidonic acid |
| EPA | eicosapentaenoic acid |
| DHA | docosahexaenoic acid |
| GL | glycerophospholipids |
| LTs | leukotrienes |
| PGs | prostaglandins |
| TXs | thromboxanes |
| Rv | resolvins |
| AEA | anandamide |
| 2-AG | 2-arachidonoilglycerol |
| PEA | palmitoylethanolamide |
| OEA | O-arachidonoylethanolamine |
| LPA | lysophoshatidic acid |
| PA | phosphatidic acid |
| DAG | diacyglicerols |
| TAG | triacylglicerols |
| CDP-DAG | cytidine ciphosphate diacylglycerol |
| CL | cardiolipins |
| PI | phosphatidyl inositoles |
| LPI | lysophosphatidyl inositoles |
| PG | phosphatidyl glicerols |
| LPG | lysophosphatidyl glicerols |
| PE | phosphatidyl ethanolamines |
| LPE | lysophosphatidyl ethanolamines |
| PC | phosphatidyl cholines |
| LPC | lysophosphatidyl cholines |
| PS | phosphatidyl serines |
| LPS | lysophosphatidyl serines |
| GlcCer | glucosylceramide |
| LacCer | lactosylceramide |
| S1P | sphingosine 1-phosphate |
| GalCer | galactosylceramide |
| SM | sphingomyelins |
| Cer-1P | ceramide 1-phsosphate |
| EETs | Epoxyeicosatrienoic acids |
| DHETs | dihydroxyeicosatrienoic acids |
| HETEs | hydroxyeicosatetraenoic acids |
| CB | cannabinoid receptor |
| RA | rheumatoid arthritis |
| SLE | systemic lupus erythematosus |
| ROS | reactive oxygen species |
| IBD | inflammatory bowel disease |
| CD MS | Crohn’s disease multiple sclerosis |
| GO | called Graves’ orbitopathy |
| ω-3 PUFAs | omega-3 polyunsaturated fatty acids |
References
- Gilroy, D.W.; Bishop-Bailey, D. Lipid mediators in immune regulation and resolution. Br. J. Pharmacol. 2019, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Tselepis, A.D.; Chapman, M.J. Inflammation, bioactive lipids and atherosclerosis: Potential roles of a lipoprotein-associated phospholipase A2, platelet activating factor-acetylhydrolase. Atheroscler. Suppl. 2002, 3, 57–68. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Leuti, A.; Maccarrone, M. Bioactive lipids and chronic inflammation: Managing the fire within. Front. Immunol. 2018, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Leishman, E.; Kunkler, P.E.; Hurley, J.H.; Miller, S.; Bradshaw, H.B. Bioactive Lipids in Cancer, Inflammation and Related Diseases; Springer Nature: Cham, Switzerland, 2019; ISBN 9783030216368. [Google Scholar]
- Coras, R.; Kavanaugh, A.; Boyd, T.; Huynh, Q.; Pedersen, B.; Armando, A.M.; Dahlberg-Wright, S.; Marsal, S.; Jain, M.; Paravar, T.; et al. Pro- and anti-inflammatory eicosanoids in psoriatic arthritis. Metabolomics 2019, 15, 1–9. [Google Scholar] [CrossRef]
- Wang, N.; Dai, R.; Wang, W.; Peng, Y.; Zhao, X.; Bi, K. Simultaneous profiling of eicosanoid metabolome in plasma by UPLC–MS/MS method: Application to identify potential makers for rheumatoid arthritis. Talanta 2016, 161, 157–164. [Google Scholar] [CrossRef]
- Fox, T.E.; Bewley, M.C.; Unrath, K.A.; Pedersen, M.M.; Anderson, R.E.; Jung, D.Y.; Jefferson, L.S.; Kim, J.K.; Bronson, S.K.; Flanagan, J.M.; et al. Circulating sphingolipid biomarkers in models of type 1 diabetes. J. Lipid Res. 2011, 52, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Klein, R.L.; Hammad, S.M.; Baker, N.L.; Hunt, K.J.; Al Gadban, M.M.; Cleary, P.A.; Virella, G.; Lopes-Virella, M.F. Decreased plasma levels of select very long chain ceramide species Are associated with the development of nephropathy in type 1 diabetes. Metabolism 2014, 63, 1287–1295. [Google Scholar] [CrossRef] [Green Version]
- Bozic, J.; Markotic, A.; Cikes-Culic, V.; Novak, A.; Borovac, J.A.; Vucemilovic, H.; Trgo, G.; Ticinovic Kurir, T. Ganglioside GM3 content in skeletal muscles is increased in type 2 but decreased in type 1 diabetes rat models: Implications of glycosphingolipid metabolism in pathophysiology of diabetes. J. Diabetes 2018, 10, 130–139. [Google Scholar] [CrossRef]
- Patyna, S.; Büttner, S.; Eckes, T.; Obermüller, N.; Bartel, C.; Braner, A.; Trautmann, S.; Thomas, D.; Geiger, H.; Pfeilschifter, J.; et al. Blood ceramides as novel markers for renal impairment in systemic lupus erythematosus. Prostaglandins Other Lipid Mediat. 2019, 144, 106348. [Google Scholar] [CrossRef]
- Checa, A.; Idborg, H.; Zandian, A.; Sar, D.G.; Surowiec, I.; Trygg, J.; Svenungsson, E.; Jakobsson, P.J.; Nilsson, P.; Gunnarsson, I.; et al. Dysregulations in circulating sphingolipids associate with disease activity indices in female patients with systemic lupus erythematosus: A cross-sectional study. Lupus 2017, 26, 1023–1033. [Google Scholar] [CrossRef]
- Al Gadban, M.M.; Alwan, M.M.; Smith, K.J.; Hammad, S.M. Accelerated vascular disease in systemic lupus erythematosus: Role of macrophage. Clin. Immunol. 2015, 157, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Navarini, L.; Bisogno, T.; Mozetic, P.; Piscitelli, F.; Margiotta, D.P.E.; Basta, F.; Afeltra, A.; Maccarrone, M. Endocannabinoid system in systemic lupus erythematosus: First evidence for a deranged 2-arachidonoylglycerol metabolism. Int. J. Biochem. Cell Biol. 2018, 99, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Zhou, J.; Yang, S.; Li, H.; Wang, C.; Fang, X.; Fan, Y.; Zhang, J.; Han, X.; Wen, C. Oxidative stress leads to reduction of plasmalogen serving as a novel biomarker for systemic lupus erythematosus. Free Radic. Biol. Med. 2016, 101, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Hu, C.; Zhao, Y.; He, L.; Zhou, J.; Li, H.; Du, Y.; Wang, Y.; Wen, C.; Han, X.; et al. Shotgun lipidomics revealed altered profiles of serum lipids in systemic lupus erythematosus closely associated with disease activity. Biomolecules 2018, 8, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Lagunas, M.J.; Martín-Venegas, R.; Moreno, J.J.; Ferrer, R. PGE2 promotes Ca2+-mediated epithelial barrier disruption through EP1 and EP4 receptors in Caco-2 cell monolayers. Am. J. Physiol. Cell Physiol. 2010, 299, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.Y.; Yang, R.S.; Lin, T.H.; Tang, C.H.; Chiu, Y.C.; Liou, H.C.; Fu, W.M. Enhancement of PLGF production by 15-(S)-HETE via PI3K-Akt, NF-κB and COX-2 pathways in rheumatoid arthritis synovial fibroblast. Eur. J. Pharmacol. 2013, 714, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Szabady, R.L.; Louissaint, C.; Lubben, A.; Xie, B.; Reeksting, S.; Tuohy, C.; Demma, Z.; Foley, S.E.; Faherty, C.S.; Llanos-Chea, A.; et al. Intestinal P-glycoprotein exports endocannabinoids to prevent inflammation and maintain homeostasis. J. Clin. Investig. 2018, 128, 4044–4056. [Google Scholar] [CrossRef]
- Sadler, T.; Bhasin, J.M.; Xu, Y.; Barnholz-Sloan, J.; Chen, Y.; Ting, A.H.; Stylianou, E. Genome-wide analysis of DNA methylation and gene expression defines molecular characteristics of Crohn’s disease-associated fibrosis. Clin. Epigenet. 2016, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yang, A.L.; Liao, J.; Li, H.; Dong, H.; Chung, Y.T.; Bai, H.; Matkowskyj, K.A.; Hammock, B.D.; Yang, G.Y. Soluble epoxide hydrolase gene deficiency or inhibition attenuates chronic active inflammatory bowel disease in IL-10(2/2) mice. Dig. Dis. Sci. 2012, 57, 2580–2591. [Google Scholar] [CrossRef]
- D’Argenio, G.; Valenti, M.; Scaglione, G.; Cosenza, V.; Sorrentini, I.; Di Marzo, V. Di Up-regulation of anandamide levels as an endogenous mechanism and a pharmacological strategy to limit colon inflammation. FASEB J. 2006, 20, 568–570. [Google Scholar] [CrossRef]
- Schmitz, K.; Brunkhorst, R.; de Bruin, N.; Mayer, C.A.; Häussler, A.; Ferreiros, N.; Schiffmann, S.; Parnham, M.J.; Tunaru, S.; Chun, J.; et al. Dysregulation of lysophosphatidic acids in multiple sclerosis and autoimmune encephalomyelitis. Acta Neuropathol. Commun. 2017, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Ju, W.; Wu, X.; Zhan, X. Elevated lysophosphatidic acid levels in the serum and cerebrospinal fluid in patients with multiple sclerosis: Therapeutic response and clinical implication. Neurol. Res. 2018, 40, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Balood, M.; Zahednasab, H.; Siroos, B.; Mesbah-Namin, S.A.; Torbati, S.; Harirchian, M.H. Elevated serum levels of lysophosphatidic acid in patients with multiple sclerosis. Hum. Immunol. 2014, 75, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Kihara, Y. Systematic Understanding of Bioactive Lipids in Neuro-Immune Interactions: Lessons from an Animal Model of Multiple Sclerosis. In Bioactive Lipids in Cancer, Inflammation and Related Diseases; Springer Nature: Cham, Switzerland, 2019; ISBN 9783030216368. [Google Scholar]
- Choi, J.W.; Gardell, S.E.; Herr, D.R.; Rivera, R.; Lee, C.W.; Noguchi, K.; Teo, S.T.; Yung, Y.C.; Lu, M.; Kennedy, G.; et al. FTY720 (fingolimod) efficacy in an animal model of multiple sclerosis requires astrocyte sphingosine 1-phosphate receptor 1 (S1P1) modulation. Proc. Natl. Acad. Sci. USA 2011, 108, 751–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plöhn, S.; Edelmann, B.; Japtok, L.; He, X.; Hose, M.; Hansen, W.; Schuchman, E.H.; Eckstein, A.; Berchner-Pfannschmidt, U. CD40 enhances sphingolipids in orbital fibroblasts: Potential role of sphingosine-1-phosphate in inflammatory T-cell migration in Graves’ orbitopathy. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5391–5397. [Google Scholar] [CrossRef] [PubMed]
- Arnardottir, H.H.; Dalli, J.; Norling, L.V.; Colas, R.A.; Perretti, M.; Serhan, C.N. Resolvin D3 Is Dysregulated in Arthritis and Reduces Arthritic Inflammation. J. Immunol. 2016, 197, 2362–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, J.; Chae, M.K.; Lee, J.H.; Lee, E.J.; Yoon, J.S. Sphingosine-1-phosphate mediates fibrosis in orbital fibroblasts in graves’ orbitopathy. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2544–2553. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.E.; Lee, J.H.; Chae, M.K.; Lee, E.J.; Yoon, J.S. The role of sphingosine-1-phosphate in adipogenesis of graves’ orbitopathy. Investig. Ophthalmol. Vis. Sci. 2016, 57, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.; Chae, M.K.; Han, S.A.; Lee, E.J.; Lee, J.H.; Yoon, J.S. Sphingosine-1-phosphate is involved in inflammatory reactions in patients with Graves’ orbitopathy. Inflamm. Res. 2017, 66, 535–545. [Google Scholar] [CrossRef]
- Richardson, D.; Pearson, R.G.; Kurian, N.; Latif, M.L.; Garle, M.J.; Barrett, D.A.; Kendall, D.A.; Scammell, B.E.; Reeve, A.J.; Chapman, V. Characterisation of the cannabinoid receptor system in synovial tissue and fluid in patients with osteoarthritis and rheumatoid arthritis. Arthritis Res. Ther. 2008, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sevastou, I.; Kaffe, E.; Mouratis, M.A.; Aidinis, V. Lysoglycerophospholipids in chronic inflammatory disorders: The PLA 2/LPC and ATX/LPA axes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2013, 1831, 42–60. [Google Scholar] [CrossRef] [PubMed]
- Nikitopoulou, I.; Oikonomou, N.; Karouzakis, E.; Sevastou, I.; Nikolaidou-Katsaridou, N.; Zhao, Z.; Mersinias, V.; Armaka, M.; Xu, Y.; Masu, M.; et al. Autotaxin expression from synovial fibroblasts is essential for the pathogenesis of modeled arthritis. J. Exp. Med. 2012, 209, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaigirdar, S.A.; Benson, R.A.; Elmesmari, A.; Kurowska-Stolarska, M.S.; McInnes, I.B.; Garside, P.; MacLeod, M.K.L. Sphingosine-1-phosphate promotes the persistence of activated CD4 T cells in inflamed sites. Front. Immunol. 2017, 8, 1627. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, M.F.; Garcia-Carbonell, R.; Whisenant, K.D.; Guma, M. Fibroblast-like synoviocyte metabolism in the pathogenesis of rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 1–12. [Google Scholar] [CrossRef]
- Fahrmann, J.; Grapov, D.; Yang, J.; Hammock, B.; Fiehn, O.; Bell, G.I.; Hara, M. Systemic alterations in the metabolome of diabetic NOD mice delineate increased oxidative stress accompanied by reduced inflammation and hypertriglyceremia. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E978–E989. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Smith, W.L.; DeWitt, D.L.; Garavito, R.M. Cyclooxygenases: Structural, Cellular, and Molecular Biology. Annu. Rev. Biochem. 2000, 69, 145–182. [Google Scholar] [CrossRef] [Green Version]
- Basil, M.C.; Levy, B.D. Specialized pro-resolving mediators: Endogenous regulators of infection and inflammation. Nat. Rev. Immunol. 2016, 16, 51–67. [Google Scholar] [CrossRef]
- Aoki, T.; Narumiya, S. Prostaglandins and chronic inflammation. Trends Pharmacol. Sci. 2012, 33, 304–311. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Segi-Nishida, E.; Miyachi, Y.; Narumiya, S. Prostacyclin-IP signaling and prostaglandin E2-EP2/EP4 signaling both mediate joint inflammation in mouse collagen-induced arthritis. J. Exp. Med. 2006, 203, 325–335. [Google Scholar] [CrossRef]
- Hirata, T.; Narumiya, S. Prostanoids as Regulators of Innate and Adaptive Immunity. Adv. Immunol. 2012, 116, 143–174. [Google Scholar] [PubMed]
- Yao, C.; Sakata, D.; Esaki, Y.; Li, Y.; Matsuoka, T.; Kuroiwa, K.; Sugimoto, Y.; Narumiya, S. Prostaglandin E2-EP4 signaling promotes immune inflammation through TH1 cell differentiation and TH17 cell expansion. Nat. Med. 2009, 15, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, S.J.; Collier, J.J. The gene encoding cyclooxygenase-2 is regulated by IL-1β and prostaglandins in 832/13 rat insulinoma cells. Cell. Immunol. 2011, 271, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Tie, C.; Wang, Z.; Zhang, J.L. Highly sensitive and specific derivatization strategy to profile and quantitate eicosanoids by UPLC-MS/MS. Anal. Chim. Acta 2017, 950, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A.; Fang, X.; Snyder, G.D.; Weintraub, N.L. Epoxyeicosatrienoic acids (EETs): Metabolism and biochemical function. Prog. Lipid Res. 2004, 43, 55–90. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Siskind, L.J.; Mullen, T.D.; Rosales, K.R.; Clarke, C.J.; Hernandez-Corbacho, M.J.; Edinger, A.L.; Obeid, L.M. The BCL-2 protein BAK is required for long-chain ceramide generation during apoptosis. J. Biol. Chem. 2010, 285, 11818–11826. [Google Scholar] [CrossRef] [Green Version]
- Mullen, T.D.; Jenkins, R.W.; Clarke, C.J.; Bielawski, J.; Hannun, Y.A.; Obeid, L.M. Ceramide synthase-dependent ceramide generation and programmed cell death: Involvement of salvage pathway in regulating postmitochondrial events. J. Biol. Chem. 2011, 286, 15929–15942. [Google Scholar] [CrossRef] [Green Version]
- Kohno, M.; Momoi, M.; Oo, M.L.; Paik, J.-H.; Lee, Y.-M.; Venkataraman, K.; Ai, Y.; Ristimaki, A.P.; Fyrst, H.; Sano, H.; et al. Intracellular Role for Sphingosine Kinase 1 in Intestinal Adenoma Cell Proliferation. Mol. Cell. Biol. 2006, 26, 7211–7223. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Spassieva, S.D.; Bieberich, E. Ceramide and S1P signaling in embryonic stem cell differentiation. Methods Mol. Biol. 2018, 1697, 153–171. [Google Scholar]
- Mishra, S.K.; Gao, Y.G.; Deng, Y.; Chalfant, C.E.; Hinchcliffe, E.H.; Brown, R.E. CPTP: A sphingolipid transfer protein that regulates autophagy and inflammasome activation. Autophagy 2018, 14, 862–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, M.; Kitatani, K.; Kondo, T.; Hashimoto-Nishimura, M.; Asano, S.; Hayashi, A.; Mitsutake, S.; Igarashi, Y.; Umehara, H.; Takeya, H.; et al. Regulation of autophagy and its associated cell death by “sphingolipid rheostat”: Reciprocal role of ceramide and sphingosine 1-phosphate in the mammalian target of rapamycin pathway. J. Biol. Chem. 2012, 287, 39898–39910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Skočaj, M.; Kreft, M.E.; Resnik, N.; Veranič, P.; Franceschi, P.; Sepčić, K.; Guella, G. Comparative lipidomic study of urothelial cancer models: Association with urothelial cancer cell invasiveness. Mol. Biosyst. 2016, 12, 3266–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckham, T.H.; Lu, P.; Cheng, J.C.; Zhao, D.; Turner, L.S.; Zhang, X.; Hoffman, S.; Armeson, K.E.; Liu, A.; Marrison, T.; et al. Acid ceramidase-mediated production of sphingosine 1-phosphate promotes prostate cancer invasion through upregulation of cathepsin B. Int. J. Cancer 2012, 131, 2034–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carreira, A.C.; Ventura, A.E.; Varela, A.R.P.; Silva, L.C. Tackling the biophysical properties of sphingolipids to decipher their biological roles. Biol. Chem. 2015, 396, 597–609. [Google Scholar] [CrossRef]
- Trajkovic, K. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008, 319, 1244–1247. [Google Scholar] [CrossRef]
- Blaho, V.A.; Galvani, S.; Engelbrecht, E.; Liu, C.; Swendeman, S.L.; Kono, M.; Proia, R.L.; Steinman, L.; Han, M.H.; Hla, T. HDL-bound sphingosine-1-phosphate restrains lymphopoiesis and neuroinflammation. Nature 2015, 523, 342–346. [Google Scholar] [CrossRef] [Green Version]
- Patwardhan, G.A.; Beverly, L.J.; Siskind, L.J. Sphingolipids and mitochondrial apoptosis. J. Bioenerg. Biomembr. 2016, 48, 153–168. [Google Scholar] [CrossRef]
- Maceyka, M.; Payne, S.G.; Milstien, S.; Spiegel, S. Sphingosine kinase, sphingosine-1-phosphate, and apoptosis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2002, 1585, 193–201. [Google Scholar] [CrossRef]
- Taha, T.A.; Mullen, T.D.; Obeid, L.M. A house divided: Ceramide, sphingosine, and sphingosine-1-phosphate in programmed cell death. Biochim. Biophys. Acta Biomembr. 2006, 1758, 2027–2036. [Google Scholar] [CrossRef] [Green Version]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. Adv. Exp. Med. Biol. 2010, 688, 1–23. [Google Scholar] [PubMed] [Green Version]
- Dei Cas, M.; Ghidoni, R. Cancer Prevention and Therapy with Polyphenols: Sphingolipid-Mediated Mechanisms. Nutrients 2018, 10, 940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhouayek, M.; Muccioli, G.G. The endocannabinoid system in inflammatory bowel diseases: From pathophysiology to therapeutic opportunity. Trends Mol. Med. 2012, 18, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Gui, H.; Tong, Q.; Qu, W.; Mao, C.M.; Dai, S.M. The endocannabinoid system and its therapeutic implications in rheumatoid arthritis. Int. Immunopharmacol. 2015, 26, 86–91. [Google Scholar] [CrossRef]
- Cencioni, M.T.; Chiurchiù, V.; Catanzaro, G.; Borsellino, G.; Bernardi, G.; Battistini, L.; Maccarrone, M. Anandamide suppresses proliferation and cytokine release from primary human T-lymphocytes mainly via CB2 receptors. PLoS ONE 2010, 5, e8688. [Google Scholar] [CrossRef]
- Ribeiro, A.; Pontis, S.; Mengatto, L.; Armirotti, A.; Chiurchiù, V.; Capurro, V.; Fiasella, A.; Nuzzi, A.; Romeo, E.; Moreno-Sanz, G.; et al. A Potent Systemically Active N-Acylethanolamine Acid Amidase Inhibitor that Suppresses Inflammation and Human Macrophage Activation. ACS Chem. Biol. 2015, 10, 1838–1846. [Google Scholar] [CrossRef] [Green Version]
- Maestroni, G.J.M. The endogenous cannabinoid 2-arachidonoyl glycerol as in vivo chemoattractant for dendritic cells and adjuvant for Th1 response to a soluble protein. FASEB J. 2004, 18, 1914–1916. [Google Scholar] [CrossRef] [Green Version]
- Gallily, R.; Breuer, A.; Mechoulam, R. 2-Arachidonylglycerol, an endogenous cannabinoid, inhibits tumor necrosis factor-α production in murine macrophages, and in mice. Eur. J. Pharmacol. 2000, 406, 5–7. [Google Scholar] [CrossRef]
- Chang, Y.H.; Lee, S.T.; Lin, W.W. Effects of cannabinoids on LPS-stimulated inflammatory mediator release from macrophages: Involvement of eicosanoids. J. Cell. Biochem. 2001, 81, 715–723. [Google Scholar] [CrossRef]
- Barrie, N.; Manolios, N. The endocannabinoid system in pain and inflammation: Its relevance to rheumatic disease. Eur. J. Rheumatol. 2017, 4, 210–218. [Google Scholar] [CrossRef]
- Ridgway, N.D.; McLeod, R.S. Biochemistry of Lipids, Lipoproteins and Membranes: Sixth Edition; Elsevier: Amsterdam, Netherlands, 2015; ISBN 9780444634382. [Google Scholar]
- Knowlden, S.; Georas, S.N. The Autotaxin–LPA Axis Emerges as a Novel Regulator of Lymphocyte Homing and Inflammation. J. Immunol. 2014, 192, 851–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimerl, S.; Fischer, M.; Baessler, A.; Liebisch, G.; Sigruener, A.; Wallner, S.; Schmitz, G. Alterations of plasma lysophosphatidylcholine species in obesity and weight loss. PLoS ONE 2014, 9, e111348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piñeiro, R.; Falasca, M. Lysophosphatidylinositol signalling: New wine from an old bottle. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2012, 1821, 694–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, B.; Schiller, J.; Wagner, U.; Häntzschel, H.; Arnold, K. The phosphatidylcholine/lysophosphatidylcholine ratio in human plasma is an indicator of the severity of rheumatoid arthritis: Investigations by 31P NMR and MALDI-TOF MS. Clin. Biochem. 2005, 38, 925–933. [Google Scholar] [CrossRef]
- Korotkova, M.; Jakobsson, P.J. Persisting eicosanoid pathways in rheumatic diseases. Nat. Rev. Rheumatol. 2014, 10, 229–241. [Google Scholar] [CrossRef]
- Hoxha, M. A systematic review on the role of eicosanoid pathways in rheumatoid arthritis. Adv. Med. Sci. 2018, 63, 22–29. [Google Scholar] [CrossRef]
- Maicas, N.; Ibáñez, L.; Alcaraz, M.J.; Úbeda, A.; Ferrándiz, M.L. Prostaglandin D 2 regulates joint inflammation and destruction in murine collagen-induced arthritis. Arthritis Rheum. 2012, 64, 130–140. [Google Scholar] [CrossRef]
- Basu, S.; Whiteman, M.; Mattey, D.L.; Halliwell, B. Raised levels of F2-isoprostanes and prostaglandin F2α in different rheumatic diseases. Ann. Rheum. Dis. 2001, 60, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Łuczaj, W.; Gindzienska-Sieskiewicz, E.; Jarocka-Karpowicz, I.; Andrisic, L.; Sierakowski, S.; Zarkovic, N.; Waeg, G.; Skrzydlewska, E. The onset of lipid peroxidation in rheumatoid arthritis: Consequences and monitoring. Free Radic. Res. 2016, 50, 304–313. [Google Scholar] [CrossRef]
- Beckmann, N.; Becker, K.A.; Walter, S.; Becker, J.U.; Kramer, M.; Hessler, G.; Weber, S.; Göthert, J.R.; Fassbender, K.; Gulbins, E.; et al. Regulation of Arthritis Severity by the Acid Sphingomyelinase. Cell. Physiol. Biochem. 2017, 43, 1460–1471. [Google Scholar] [CrossRef] [Green Version]
- Poolman, T.M.; Gibbs, J.; Walker, A.L.; Dickson, S.; Farrell, L.; Hensman, J.; Kendall, A.C.; Maidstone, R.; Warwood, S.; Loudon, A.; et al. Rheumatoid arthritis reprograms circadian output pathways. Arthritis Res. Ther. 2019, 21, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Kohno, M.; Nagahara, H.; Murakami, K.; Sagawa, T.; Kasahara, A.; Kaneshita, S.; Kida, T.; Fujioka, K.; Wada, M.; et al. Upregulation of sphingosine-1-phosphate receptor 3 on fibroblast-like synoviocytes is associated with the development of collageninduced arthritis via increased interleukin-6 production. PLoS ONE 2019, 14, 1–15. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, K.H.; Jin, S.; Kim, J.; Yoo, I.; Pack, S.P.; Ha, U.H.; Park, T.W.; Choi, S.A.; Yuk, S.H.; et al. Decreased expression of sphingosine-1-phosphate receptor 1 in the blood leukocyte of rheumatoid arthritis patients. Immune Netw. 2018, 18, 4–9. [Google Scholar] [CrossRef]
- Xiao, L.; Zhou, Y.; Friis, T.; Beagley, K.; Xiao, Y. S1P-S1PR1 Signaling: The “Sphinx” in Osteoimmunology. Front. Immunol. 2019, 10, 1–18. [Google Scholar] [CrossRef]
- Balderas, C.; Rupérez, F.J.; Ibañez, E.; Señorans, J.; Guerrero-Fernández, J.; Casado, I.G.; Gracia-Bouthelier, R.; García, A.; Barbas, C. Plasma and urine metabolic fingerprinting of type 1 diabetic children. Electrophoresis 2013, 34, 2882–2890. [Google Scholar] [CrossRef]
- Murfitt, S.A.; Zaccone, P.; Wang, X.; Acharjee, A.; Sawyer, Y.; Koulman, A.; Roberts, L.D.; Cooke, A.; Griffin, J.L. Metabolomics and Lipidomics Study of Mouse Models of Type 1 Diabetes Highlights Divergent Metabolism in Purine and Tryptophan Metabolism Prior to Disease Onset. J. Proteome Res. 2018, 17, 946–960. [Google Scholar] [CrossRef] [Green Version]
- La Torre, D.; Seppänen-Laakso, T.; Larsson, H.E.; Hyötyläinen, T.; Ivarsson, S.A.; Lernmark, Å.; Orešič, M. Decreased cord-blood phospholipids in young age-at-onset type 1 diabetes. Diabetes 2013, 62, 3951. [Google Scholar] [CrossRef] [Green Version]
- Fievet, C.; Ziegler, O.; Parra, H.J.; Mejean, L.; Fruchart, J.C.; Drouin, P. Depletion in choline containing phospholipids of LpB particles in adequately controlled type I insulin-dependent diabetes mellitus. Diabetes Metab. 1990, 16, 64–69. [Google Scholar]
- Denimal, D.; Pais de Barros, J.P.; Petit, J.M.; Bouillet, B.; Vergès, B.; Duvillard, L. Significant abnormalities of the HDL phosphosphingolipidome in type 1 diabetes despite normal HDL cholesterol concentration. Atherosclerosis 2015, 241, 752–760. [Google Scholar] [CrossRef]
- Acharya, N.; Penukonda, S.; Shcheglova, T.; Hagymasi, A.T.; Basu, S.; Srivastava, P.K. Endocannabinoid system acts as a regulator of immune homeostasis in the gut. Proc. Natl. Acad. Sci. USA 2017, 114, 5005–5010. [Google Scholar] [CrossRef] [Green Version]
- Lemos, J.P.; Smaniotto, S.; Messias, C.V.; Moreira, O.C.; Cotta-de-almeida, V.; Dardenne, M.; Savino, W.; Mendes-da-cruz, D.A. Sphingosine-1-phosphate receptor 1 is involved in non-obese diabetic mouse thymocyte migration disorders. Int. J. Mol. Sci. 2018, 19, 1446. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Bolick, D.T.; Lukashev, D.; Lappas, C.; Sitkovsky, M.; Lynch, K.R.; Hedrick, C.C. Sphingosine-1-phosphate reduces CD4 + T-cell activation in type 1 diabetes through regulation of hypoxia-inducible factor short isoform I.1 and CD69. Diabetes 2008, 57, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Hahn, C.; Tyka, K.; Saba, J.D.; Lenzen, S.; Gurgul-Convey, E. Overexpression of sphingosine-1-phosphate lyase protects insulin-secreting cells against cytokine toxicity. J. Biol. Chem. 2017, 292, 20292–20304. [Google Scholar] [CrossRef] [Green Version]
- Das, U.N. Current and emerging strategies for the treatment and management of systemic lupus erythematosus based on molecular signatures of acute and chronic inflammation. J. Inflamm. Res. 2010, 3, 143–170. [Google Scholar] [CrossRef] [Green Version]
- Gualtierotti, R.; Guarnaccia, L.; Beretta, M.; Navone, S.E.; Campanella, R.; Riboni, L.; Rampini, P.; Marfia, G. Modulation of Neuroinflammation in the Central Nervous System: Role of Chemokines and Sphingolipids. Adv. Ther. 2017, 34, 396–420. [Google Scholar] [CrossRef]
- Oaks, Z.; Winans, T.; Caza, T.; Fernandez, D.; Liu, Y.; Landas, S.K.; Banki, K.; Perl, A. Mitochondrial Dysfunction in the Liver and Antiphospholipid Antibody Production Precede Disease Onset and Respond to Rapamycin in Lupus-Prone Mice. Arthritis Rheumatol. 2016, 68, 2728–2739. [Google Scholar] [CrossRef]
- Mistry, P.; Kaplan, M.J. Cell death in the pathogenesis of systemic lupus erythematosus and lupus nephritis. Clin. Immunol. 2017, 185, 59–73. [Google Scholar] [CrossRef]
- Jovanović, V.; Abdul Aziz, N.; Lim, Y.T.; Ng Ai Poh, A.; Jin Hui Chan, S.; Ho Xin Pei, E.; Lew, F.C.; Shui, G.; Jenner, A.M.; Bowen, L.; et al. Lipid Anti-Lipid Antibody Responses Correlate with Disease Activity in Systemic Lupus Erythematosus. PLoS ONE 2013, 8, 55639. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.B.; Pereira, A.M.; Melo, T.; Paiva, A.; Domingues, M.R. Lipidomics in autoimmune diseases with main focus on systemic lupus erythematosus. J. Pharm. Biomed. Anal. 2019, 174, 386–395. [Google Scholar] [CrossRef]
- Elloumi, N.; Ben Mansour, R.; Marzouk, S.; Mseddi, M.; Fakhfakh, R.; Gargouri, B.; Masmoudi, H.; Lassoued, S. Differential reactive oxygen species production of neutrophils and their oxidative damage in patients with active and inactive systemic lupus erythematosus. Immunol. Lett. 2017, 184, 1–6. [Google Scholar] [CrossRef]
- McDonald, G.; Deepak, S.; Miguel, L.; Hall, C.J.; Isenberg, D.A.; Magee, A.I.; Butters, T.; Jury, E.C. Normalizing glycosphingolipids restores function in CD4+ T cells from lupus patients. J. Clin. Investig. 2014, 124, 712–724. [Google Scholar] [CrossRef] [Green Version]
- Watson, L.; Tullus, K.; Marks, S.D.; Holt, R.C.L.; Pilkington, C.; Beresford, M.W. Increased serum concentration of sphingosine-1-phosphate in juvenile-onset systemic lupus erythematosus. J. Clin. Immunol. 2012, 32, 1019–1025. [Google Scholar] [CrossRef]
- Sun, X.J.; Wang, C.; Zhang, L.X.; Yu, F.; Chen, M.; Zhao, M.H. Sphingosine-1-phosphate and its receptors in anti-neutrophil cytoplasmic antibody-associated vasculitis. Nephrol. Dial. Transplant. 2017, 32, 1313–1322. [Google Scholar] [CrossRef] [Green Version]
- Snider, A.J. Sphingosine kinase and sphingosine-1-phosphate: Regulators in autoimmune and inflammatory disease. Int. J. Clin. Rheumtol. 2013, 8, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Rahaman, O.; Bhattacharya, R.; Liu, C.S.C.; Raychaudhuri, D.; Ghosh, A.R.; Bandopadhyay, P.; Pal, S.; Goswami, R.P.; Sircar, G.; Ghosh, P.; et al. Cutting Edge: Dysregulated Endocannabinoid-Rheostat for Plasmacytoid Dendritic Cell Activation in a Systemic Lupus Endophenotype. J. Immunol. 2019, 202, 1674–1679. [Google Scholar] [CrossRef] [Green Version]
- Onyiah, J.C.; Colgan, S.P. Cytokine responses and epithelial function in the intestinal mucosa. Cell. Mol. Life Sci. 2016, 73, 4203–4212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grill, M.; Hasenoehrl, C.; Kienzl, M.; Kargl, J.; Schicho, R. Cellular localization and regulation of receptors and enzymes of the endocannabinoid system in intestinal and systemic inflammation. Histochem. Cell Biol. 2019, 151, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, R.J.; Andrews, C.; Brice, D.P.; Durum, S.K.; McLean, M.H. Can we target endogenous anti-inflammatory responses as a therapeutic strategy for inflammatory bowel disease? Inflamm. Bowel Dis. 2018, 24, 2123–2134. [Google Scholar] [CrossRef] [PubMed]
- Kihara, Y.; Matsushita, T.; Kita, Y.; Uematsu, S.; Akira, S.; Kira, J.; Ishii, S. Targeted lipidomics reveals mPGES-1-PGE2. Proc. Natl. Acad. Sci. USA 2009, 2–7. [Google Scholar]
- Palumbo, S.; Bosetti, F. Alterations of brain eicosanoid synthetic pathway in multiple sclerosis and in animal models of demyelination: Role of cyclooxygenase-2. Prostaglandins Leukot. Essent. Fat. Acids 2013, 89, 273. [Google Scholar] [CrossRef]
- Hon, G.M.; Hassan, M.S.; Van Rensburg, S.J.; Abel, S.; Erasmus, R.T.; Matsha, T. Peripheral blood mononuclear cell membrane fluidity and disease outcome in patients with multiple sclerosis. Indian J. Hematol. Blood Transfus. 2012, 28, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Very long-chain n-3 fatty acids and human health: Fact, fiction and the future. Proc. Nutr. Soc. 2018, 77, 52–72. [Google Scholar] [CrossRef] [Green Version]
- Curado Borges, M.; de Miranda Moura dos Santos, F.; Weiss Telles, R.; Melo de Andrade, M.V.; Toulson Davisson Correia, M.I.; Lanna, C.C.D. Omega-3 fatty acids, inflammatory status and biochemical markers of patients with systemic lupus erythematosus: A pilot study. Rev. Bras. Reumatol. 2017, 57, 526–534. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Bi, X.; Wang, S.; Zhang, Z.; Li, F.; Zhao, A.Z. Therapeutic Potential of ω-3 Polyunsaturated Fatty Acids in Human Autoimmune Diseases. Front. Immunol. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Dawczynski, C.; Hackermeier, U.; Viehweger, M.; Stange, R.; Springer, M.; Jahreis, G. Incorporation of n-3 PUFA and -linolenic acid in blood lipids and red blood cell lipids together with their influence on disease activity in patients with chronic inflammatory arthritis - A randomized controlled human intervention trial. Lipids Health Dis. 2011, 10, 130. [Google Scholar] [CrossRef] [Green Version]
- Yates, C.M.; Calder, P.C.; Ed Rainger, G. Pharmacology and therapeutics of omega-3 polyunsaturated fatty acids in chronic inflammatory disease. Pharmacol. Ther. 2014, 141, 272–282. [Google Scholar] [CrossRef]
- Jiang, J.; Li, K.; Wang, F.; Yang, B.; Fu, Y.; Zheng, J.; Li, D. Effect of Marine-Derived n-3 Polyunsaturated Fatty Acids on Major Eicosanoids: A Systematic Review and Meta-Analysis from 18 Randomized Controlled Trials. PLoS ONE 2016, 11, e0147351. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Caughey, G.E.; James, M.J.; Proudman, S.M.; Cleland, L.G. Fish oil supplementation increases the cyclooxygenase inhibitory activity of paracetamol in rheumatoid arthritis patients. Complement. Ther. Med. 2010, 18, 171–174. [Google Scholar] [CrossRef]
- Bi, X.; Li, F.; Liu, S.; Jin, Y.; Zhang, X.; Yang, T.; Dai, Y.; Li, X.; Zhao, A.Z. Ω-3 Polyunsaturated Fatty Acids Ameliorate Type 1 Diabetes and Autoimmunity. J. Clin. Investig. 2017, 127, 1757–1771. [Google Scholar] [CrossRef] [Green Version]
- Bosco, N.; Brahmbhatt, V.; Oliveira, M.; Martin, F.P.; Lichti, P.; Raymond, F.; Mansourian, R.; Metairon, S.; Pace-Asciak, C.; Bastic Schmid, V.; et al. Effects of increase in fish oil intake on intestinal eicosanoids and inflammation in a mouse model of colitis. Lipids Health Dis. 2013, 12, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Orefice, N.S.; Alhouayek, M.; Carotenuto, A.; Montella, S.; Barbato, F.; Comelli, A.; Calignano, A.; Muccioli, G.G.; Orefice, G. Oral Palmitoylethanolamide Treatment Is Associated with Reduced Cutaneous Adverse Effects of Interferon-β1a and Circulating Proinflammatory Cytokines in Relapsing–Remitting Multiple Sclerosis. Neurotherapeutics 2016, 13, 428–438. [Google Scholar] [CrossRef]
- Serhan, C.N.; Clish, C.B.; Brannon, J.; Colgan, S.P.; Chiang, N.; Gronert, K. Novel functional sets of lipid-derived mediators with antiinflammatory actions generated from omega-3 fatty acids via cyclooxygenase 2-nonsteroidal antiinflammatory drugs and transcellular processing. J. Exp. Med. 2000, 192, 1197–1204. [Google Scholar] [CrossRef] [Green Version]
- Diab, A.; Deng, C.; Smith, J.D.; Hussain, R.Z.; Phanavanh, B.; Lovett-Racke, A.E.; Drew, P.D.; Racke, M.K. Peroxisome Proliferator-Activated Receptor-γ Agonist 15-Deoxy-Δ 12,14 12,14 -Prostaglandin J 2 Ameliorates Experimental Autoimmune Encephalomyelitis. J. Immunol. 2002, 168, 2508–2515. [Google Scholar] [CrossRef] [Green Version]
- Guan, H.; Zhao, L.; Cao, H.; Chen, A.; Xiao, J. Epoxyeicosanoids suppress osteoclastogenesis and prevent ovariectomy-induced bone loss. FASEB J. 2015, 29, 1092–1101. [Google Scholar] [CrossRef]
- Yamamoto, S.; Yamashina, K.; Ishikawa, M.; Gotoh, M.; Yagishita, S.; Iwasa, K.; Maruyama, K.; Murakami-Murofushi, K.; Yoshikawa, K. Protective and therapeutic role of 2-carba-cyclic phosphatidic acid in demyelinating disease. J. Neuroinflammation 2017, 14, 142. [Google Scholar] [CrossRef] [Green Version]
- Lowin, T.; Apitz, M.; Anders, S.; Straub, R.H. Anti-inflammatory effects of N-acylethanolamines in rheumatoid arthritis synovial cells are mediated by TRPV1 and TRPA1 in a COX-2 dependent manner. Arthritis Res. Ther. 2015, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Liang, P.; Liu, J.; Jiang, H.; Fan, X.; Chen, G.; Zhou, C. Elevation of arachidonoylethanolamide levels by activation of the endocannabinoid system protects against colitis and ameliorates remote organ lesions in mice. Exp. Ther. Med. 2017, 14, 5664–5670. [Google Scholar] [CrossRef]
- Marquéz, L.; Suárez, J.; Iglesias, M.; Bermudez-Silva, F.J.; de Fonseca, F.R.; Andreu, M. Ulcerative colitis induces changes on the expression of the endocannabinoid system in the human colonic tissue. PLoS ONE 2009, 4, e6893. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.A.; Barkhof, F.; Comi, G.; Hartung, H.P.; Khatri, B.O.; Montalban, X.; Pelletier, J.; Capra, R.; Gallo, P.; Izquierdo, G.; et al. Oral fingolimod or intramuscular interferon for relapsing multiple sclerosis. N. Engl. J. Med. 2010, 362, 402–415. [Google Scholar] [CrossRef]
- Baker, K.F.; Isaacs, J.D. Novel therapies for immune-mediated inflammatory diseases: What can we learn from their use in rheumatoid arthritis, spondyloarthritis, systemic lupus erythematosus, psoriasis, Crohn’s disease and ulcerative colitis? Ann. Rheum. Dis. 2018, 77, 175–187. [Google Scholar] [CrossRef]
- Sandborn, W.J.; Feagan, B.G.; Wolf, D.C.; D’Haens, G.; Vermeire, S.; Hanauer, S.B.; Ghosh, S.; Smith, H.; Cravets, M.; Frohna, P.A.; et al. Ozanimod induction and maintenance treatment for ulcerative colitis. N. Engl. J. Med. 2016, 374, 1574–1762. [Google Scholar] [CrossRef]
- Moon, H.; Chon, J.; Joo, J.; Kim, D.; In, J.; Lee, H.; Park, J.; Choi, J. FTY720 preserved islet β-cell mass by inhibiting apoptosis and increasing survival of β-cells in db/db mice. Diabetes. Metab. Res. Rev. 2013, 29, 19–24. [Google Scholar] [CrossRef]
- Baumruker, T.; Billich, A.; Brinkmann, V. FTY720, an immunomodulatory sphingolipid mimetic: Translation of a novel mechanism into clinical benefit in multiple sclerosis. Expert Opin. Investig. Drugs 2007, 16, 283–289. [Google Scholar] [CrossRef]
- Brown, B.A.; Kantesaria, P.P.; McDevitt, L.M. Fingolimod: A novel immunosuppressant for multiple sclerosis. Ann. Pharmacother. 2007, 41, 1660–1668. [Google Scholar] [CrossRef]
- Brinkmann, V.; Billich, A.; Baumruker, T.; Heining, P.; Schmouder, R.; Francis, G.; Aradhye, S.; Burtin, P. Fingolimod (FTY720): Discovery and development of an oral drug to treat multiple sclerosis. Nat. Rev. Drug Discov. 2010, 9, 883–897. [Google Scholar] [CrossRef]
- Maki, T.; Gottschalk, R.; Monaco, A.P. Prevention of autoimmune diabetes by FTY720 in nonobese diabetic mice. Transplantation 2002, 74, 1684–1686. [Google Scholar] [CrossRef]
- Maki, T.; Gottschalk, R.; Ogawa, N.; Monaco, A.P. Prevention and cure of autoimmune diabetes in nonobese diabetic mice by continuous administration of FTY720. Transplantation 2005, 79, 1051–1055. [Google Scholar] [CrossRef]
- Alperovich, G.; Rama, I.; Lloberas, N.; Franquesa, M.; Poveda, R.; Gomà, M.; Herrero-Fresneda, I.; Cruzado, J.M.; Bolaños, N.; Carrera, M.; et al. New immunosuppresor strategies in the treatment of murine lupus nephritis. Lupus 2007, 16, 18–24. [Google Scholar] [CrossRef]
- Elflein, J. Prevalence of Diagnosed Autoimmune Conditions in Selected Countries. 2019. Available online: https://www.statista.com/statistics/418328/diagnosed-autoimmune-conditions-prevalence-in-selected-countries/ (accessed on 9 December 2019).
- Calder, P.C.; Bosco, N.; Bourdet-Sicard, R.; Capuron, L.; Delzenne, N.; Doré, J.; Franceschi, C.; Lehtinen, M.J.; Recker, T.; Salvioli, S.; et al. Health relevance of the modification of low grade inflammation in ageing (inflammageing) and the role of nutrition. Ageing Res. Rev. 2017, 40, 95–119. [Google Scholar] [CrossRef]
- Calder, P.C. Docosahexaenoic acid. Ann. Nutr. Metab. 2016, 69, 7–21. [Google Scholar] [CrossRef]
- Joffre, C.; Rey, C.; Layé, S. N-3 polyunsaturated fatty acids and the resolution of neuroinflammation. Front. Pharmacol. 2019, 10, 1022. [Google Scholar] [CrossRef] [Green Version]
- Apostolou, A.; Varone, A.; Kasendra, M.; Luc, R. Physiology and Pathophysiology of Human Gut: Intestine-on-chip. Patent WO 2019/060735 A1. 2019. Available online: https://patentimages.storage.googleapis.com/a4/8b/87/5fd99b5bb7f39c/WO2019060735A1.pdf (accessed on 28 March 2019).
- Marques, M.P.C.; Boyd, A.S.; Polizzi, K.; Szita, N. Microfluidic devices towards personalized health and wellbeing. J. Chem. Technol. Biotechnol. 2019, 94, 2412–2415. [Google Scholar] [CrossRef]
Figure 1.
Inflammatory lipids and their interconnections. The green points indicate lipids with anti-inflammatory properties, whereas the red points indicate lipids with pro-inflammatory properties [5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37]. FA: fatty acids and their derivatives; PUFA: polyunsaturated fatty acids containing both endocannabinoids and eicosanoids; AA: arachidonic acid; EPA: eicosapentaenoic acid; DHA: docosahexaenoic acid; GL: glycerophospholipids; LTs: leukotrienes; PGs: prostaglandins; TXs: thromboxanes; Rv: resolvins; AEA: anandamide; 2-AG: 2-arachidonoilglycerol; PEA: palmitoylethanolamide; OEA: O-arachidonoylethanolamine; LPA: lysophoshatidic acid; PA: phosphatidic acid; DAG: diacyglicerols; TAG: triacylglicerols; CDP-DAG: cytidine diphosphate diacylglycerol; CL: cardiolipins; PI: phosphatidyl inositols; LPI: lysophosphatidylinositols; PG: phosphatidyl glycerols; LPG: lysophosphatidylglycerols; PE: phosphatidylethanolamines; LPE: lysophosphatidylethanolamines; PC: phosphatidyl cholines; LPC: lysophosphatidylcholines; PS: phosphatidyl serines; LPS: lysophosphatidylserines; GlcCer: glucosylceramide; LacCer: lactosylceramide; S1P: sphingosine 1-phosphate; GalCer: galactosylceramide; SM: sphingomyelins; Cer-1P: ceramide 1-phosphate.
Figure 1.
Inflammatory lipids and their interconnections. The green points indicate lipids with anti-inflammatory properties, whereas the red points indicate lipids with pro-inflammatory properties [5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37]. FA: fatty acids and their derivatives; PUFA: polyunsaturated fatty acids containing both endocannabinoids and eicosanoids; AA: arachidonic acid; EPA: eicosapentaenoic acid; DHA: docosahexaenoic acid; GL: glycerophospholipids; LTs: leukotrienes; PGs: prostaglandins; TXs: thromboxanes; Rv: resolvins; AEA: anandamide; 2-AG: 2-arachidonoilglycerol; PEA: palmitoylethanolamide; OEA: O-arachidonoylethanolamine; LPA: lysophoshatidic acid; PA: phosphatidic acid; DAG: diacyglicerols; TAG: triacylglicerols; CDP-DAG: cytidine diphosphate diacylglycerol; CL: cardiolipins; PI: phosphatidyl inositols; LPI: lysophosphatidylinositols; PG: phosphatidyl glycerols; LPG: lysophosphatidylglycerols; PE: phosphatidylethanolamines; LPE: lysophosphatidylethanolamines; PC: phosphatidyl cholines; LPC: lysophosphatidylcholines; PS: phosphatidyl serines; LPS: lysophosphatidylserines; GlcCer: glucosylceramide; LacCer: lactosylceramide; S1P: sphingosine 1-phosphate; GalCer: galactosylceramide; SM: sphingomyelins; Cer-1P: ceramide 1-phosphate.


{kind=link}
Table 1.
Functional lipids and their corresponding role in autoimmune diseases.
| Autoimmune Diseases | Lipids | Roles | Changes 1 | References |
|---|---|---|---|---|
| Rheumatoid arthritis | 11-HEPE, 12-HEPE and 15-HEPE (EPA-derived) | anti-inflammatory | up | [5] |
| PGE2, 6,15-dk, dh, PGF1α, 12-HETE, 12LOX derived eicosanoids (AA-derived) | pro-inflammatory | up | [5] | |
| Resolving D1 and 17-HDoHE (DHA derived) | anti-inflammatory | down | [5] | |
| molecules from COX and LOX pathways | remodeling process of inflammation | up | [6,78] | |
| 5-HETE and LTD4 | remodeling process of inflammation | down | [6] | |
| PGE2 and 15-(S)-HETE | inflammatory stimulators | up | [17,79] | |
| PGD2 | anti-inflammatory | up | [80] | |
| 8-iso-PGF(2α) and 15-keto-dihydro-PGF(2α) | Oxidative stress and inflammation biomarkers | up | [81,82] | |
| Resolvin D3 | resolution of inflammation | down | [28] | |
| AEA and 2-AG | neovascularization, cartilage, and bone demolition | up | [32] | |
| PEA | anti-inflammatory | down | [32] | |
| LPA | pro-inflammatory | up | [33,34] | |
| S1P | abnormal immune responses | up | [35,36] | |
| Type 1 diabetes | thromboxane A2 (TXA2), leukotriene B4 (LTB4), PGD2, PGE2, 11-,12- and 15-HETE, and 12-oxo-ETE | impaired states of systemic inflammation | down | [37] |
| S1P | immunomodulation | up | [7,8] | |
| omega-9 esterified sphingolipids | cardio- and neuro-protection | down | [7,9] | |
| Systemic lupus erythematosus | Ceramide 24:1 | biomarker of lupus nephritis | up | [10] |
| Ceramides, hexosylceramides | associated with disease activity and vascular complication | up | [11,12] | |
| 2-AG | disease regression | up | [13] | |
| Phospholipids (PE) | increased oxidative stress | down | [14,15] | |
| Inflammatory bowel diseases | PGE2 | paracellular regulation | up | [16] |
| HxA3 | pro-inflammatory, activates migration | up | [18] | |
| Endocannabinoid agonist on CB2 | anti-inflammatory, inhibits migration | down | [18] | |
| prostaglandin D2 and prostacyclin | excessive deposition of extracellular matrix and obstruction of the gut lumen | down | [19] | |
| epoxyeicosatrienoic acids (EET) and dihydroxyeicosatrienoic acids (DHETs) | anti-inflammatory activity | down | [20] | |
| PGE2 and 5-HETE metabolites | pro-inflammatory | up | [20] | |
| Anandamide | Protective action | up | [21] | |
| Multiple sclerosis | Lysophospholipids | demoting remyelination | down | [22] |
| Lysophosphatidic acid (LPA) | Pro- and anti-inflammatory | up | [23,24] | |
| PGE2 | Pro-inflammatory | up | [25] | |
| LTC4, LTB4, LTD4 | anti-inflammatory | down | [25] | |
| S1P | astrogliosis | up | [26] | |
| Grave’s disease | S1P | chemoattractant for T cells, pro-inflammatory, fibrosis and adipogenesis | up | [27,29,30,31] |
1 This column reports if the lipids concentrations are incremented (up) or decremented (down) as a consequence of the biological response to the effects of the disease.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cas, M.D.; Roda, G.; Li, F.; Secundo, F. Functional Lipids in Autoimmune Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 3074. https://doi.org/10.3390/ijms21093074
AMA Style
Cas MD, Roda G, Li F, Secundo F. Functional Lipids in Autoimmune Inflammatory Diseases. International Journal of Molecular Sciences. 2020; 21(9):3074. https://doi.org/10.3390/ijms21093074
Chicago/Turabian StyleCas, Michele Dei, Gabriella Roda, Feng Li, and Francesco Secundo. 2020. "Functional Lipids in Autoimmune Inflammatory Diseases" International Journal of Molecular Sciences 21, no. 9: 3074. https://doi.org/10.3390/ijms21093074
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.