Inhibition of RNA Helicase Activity Prevents Coxsackievirus B3-Induced Myocarditis in Human iPS Cardiomyocytes

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Screen Effective Antiviral Agent
2.2. E2CI Inhibit CVB3 Replication in HELA CELLS
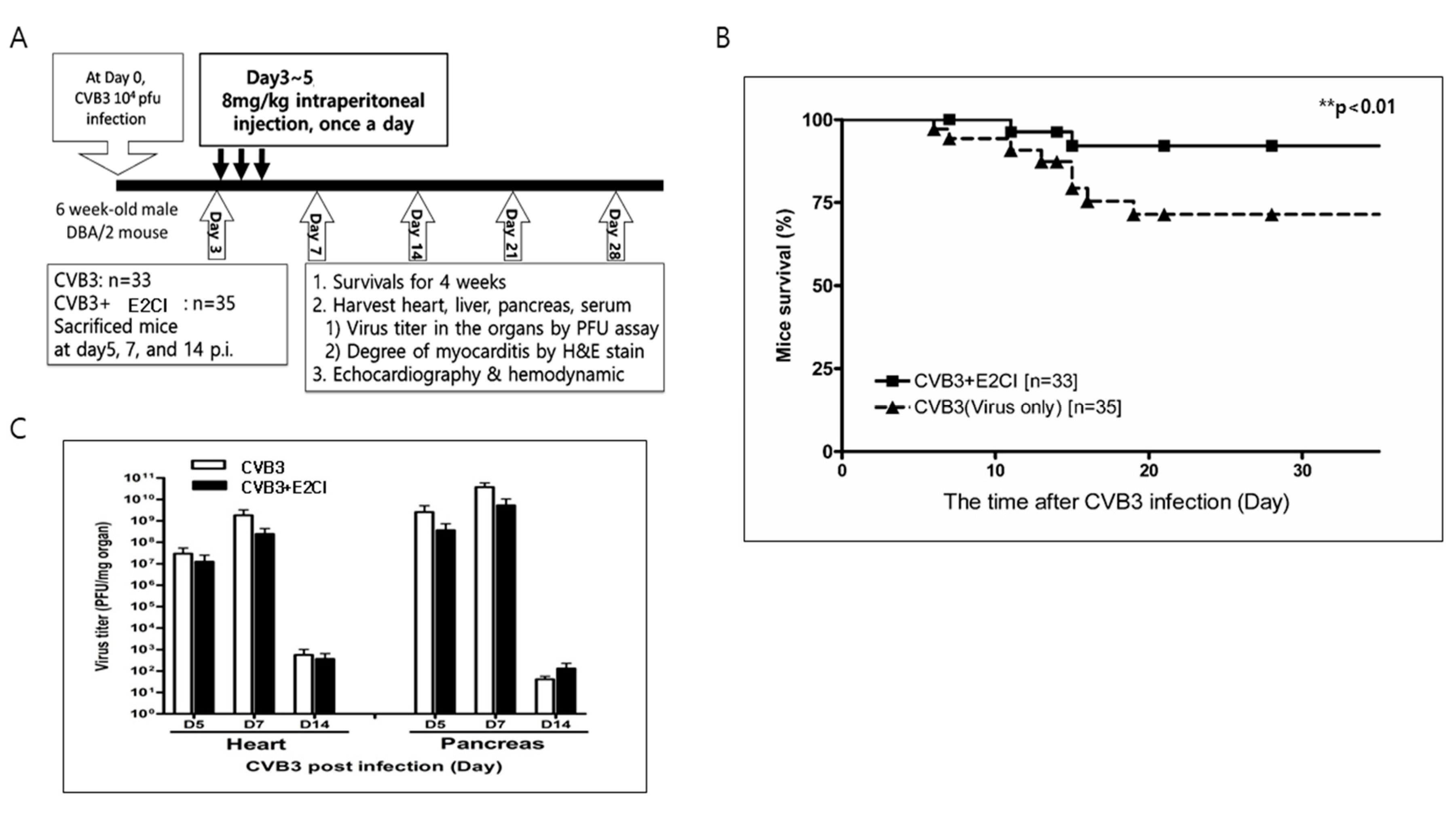
2.3. E2CI Decreases Mouse Mortality in a Murine Viral Myocarditis Model
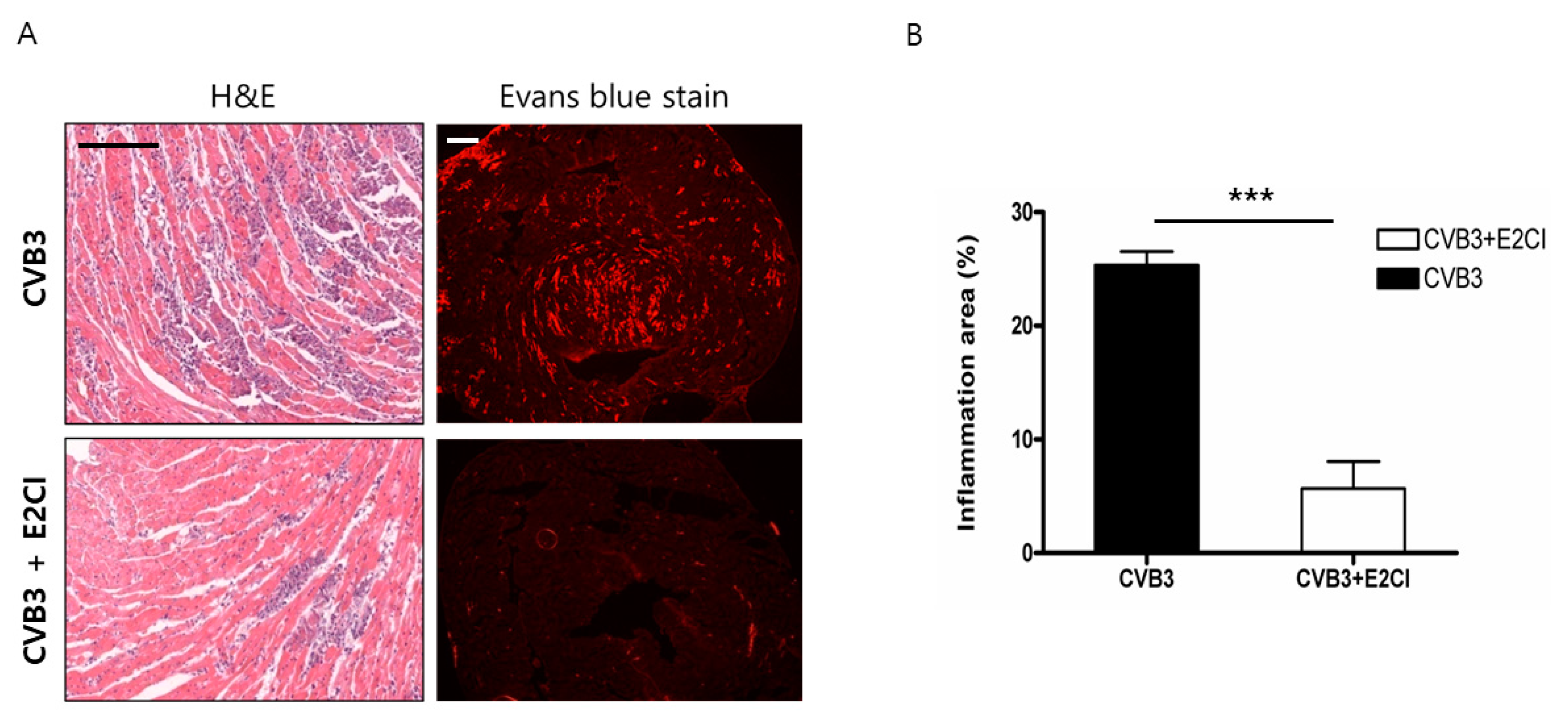
2.4. Decrease Cardiomyocyte Damage and Heart Inflammation
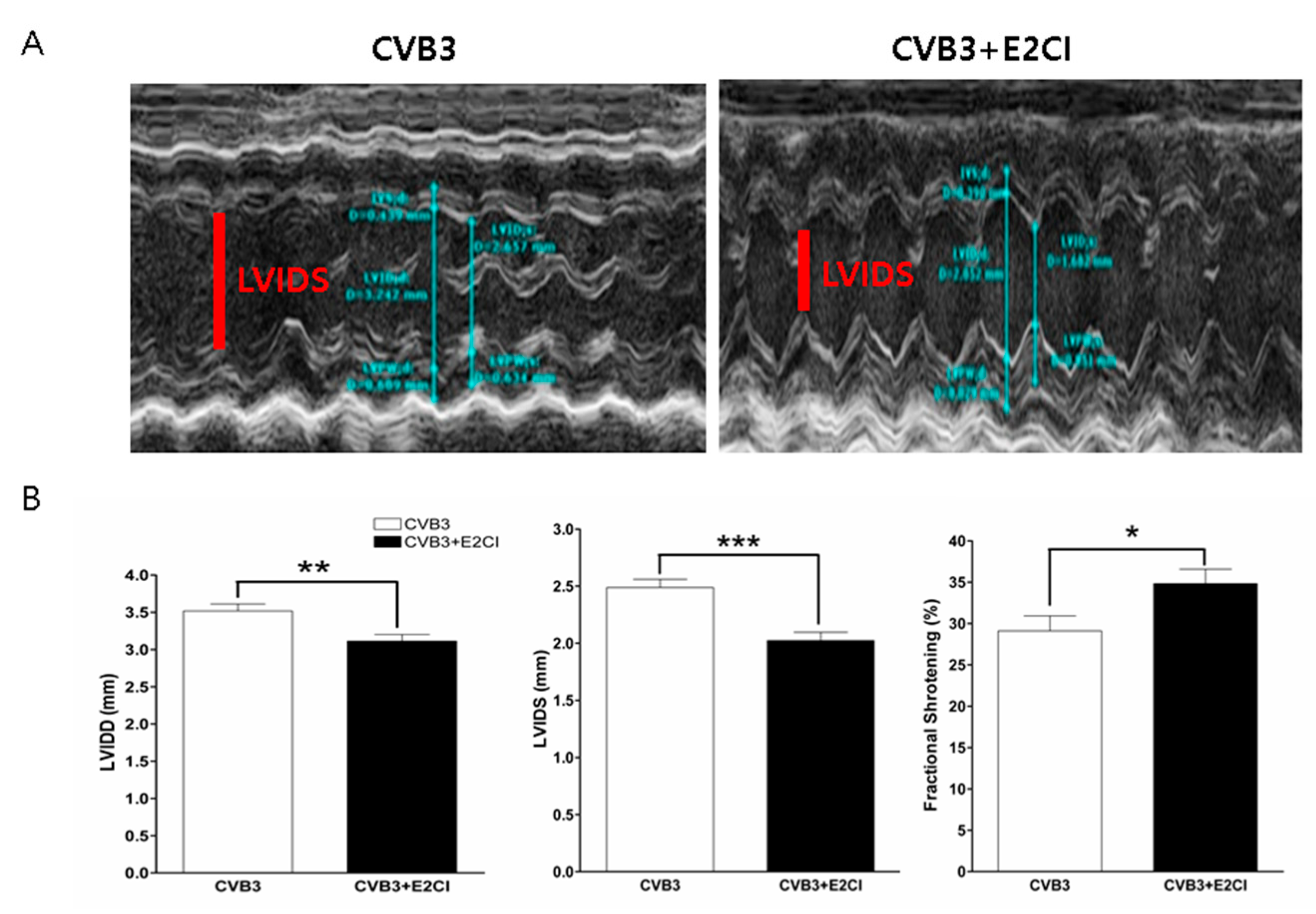
2.5. E2CI Prevent LV Dysfunction and Progression of Cardiomyopathy in CVB3 Infection
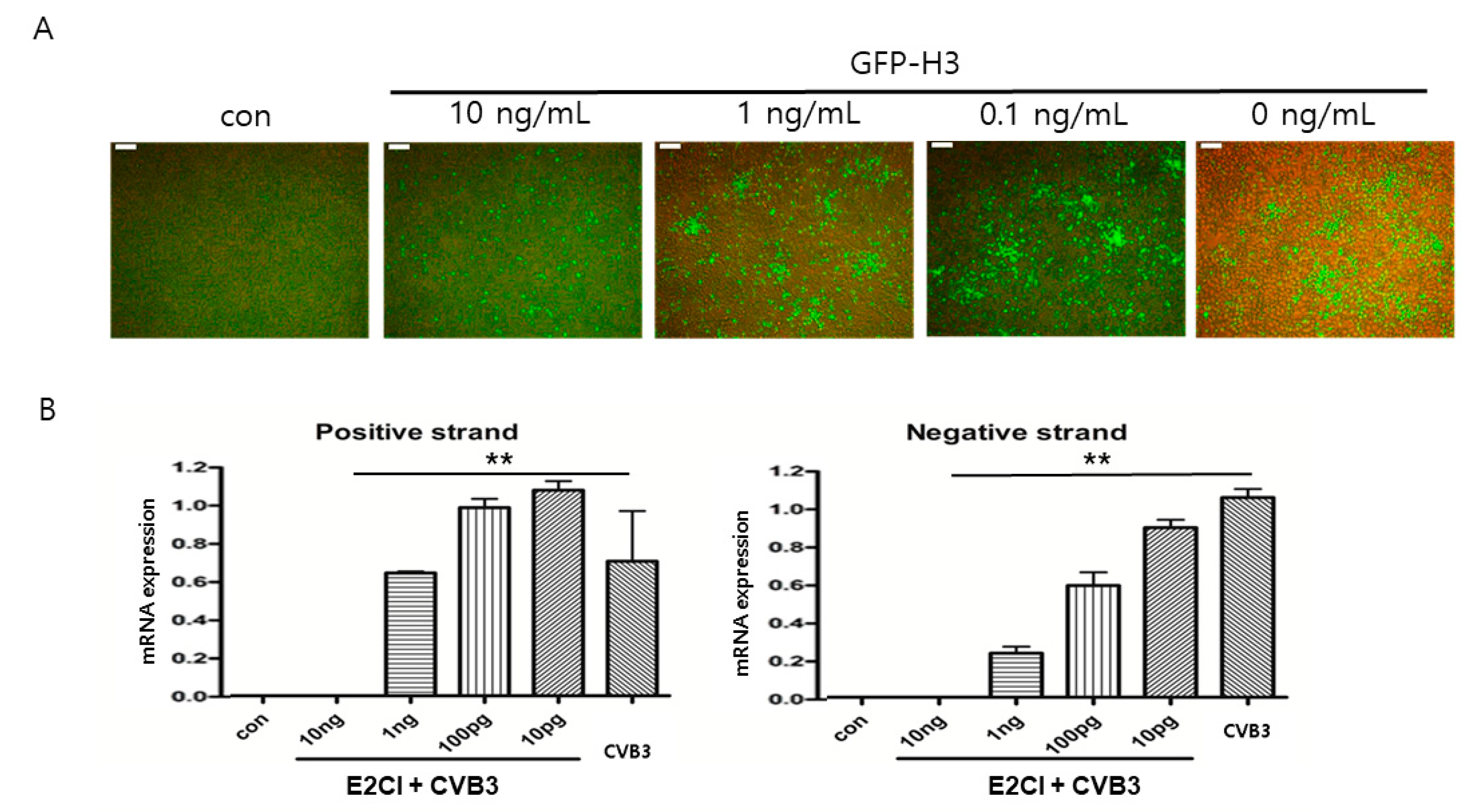
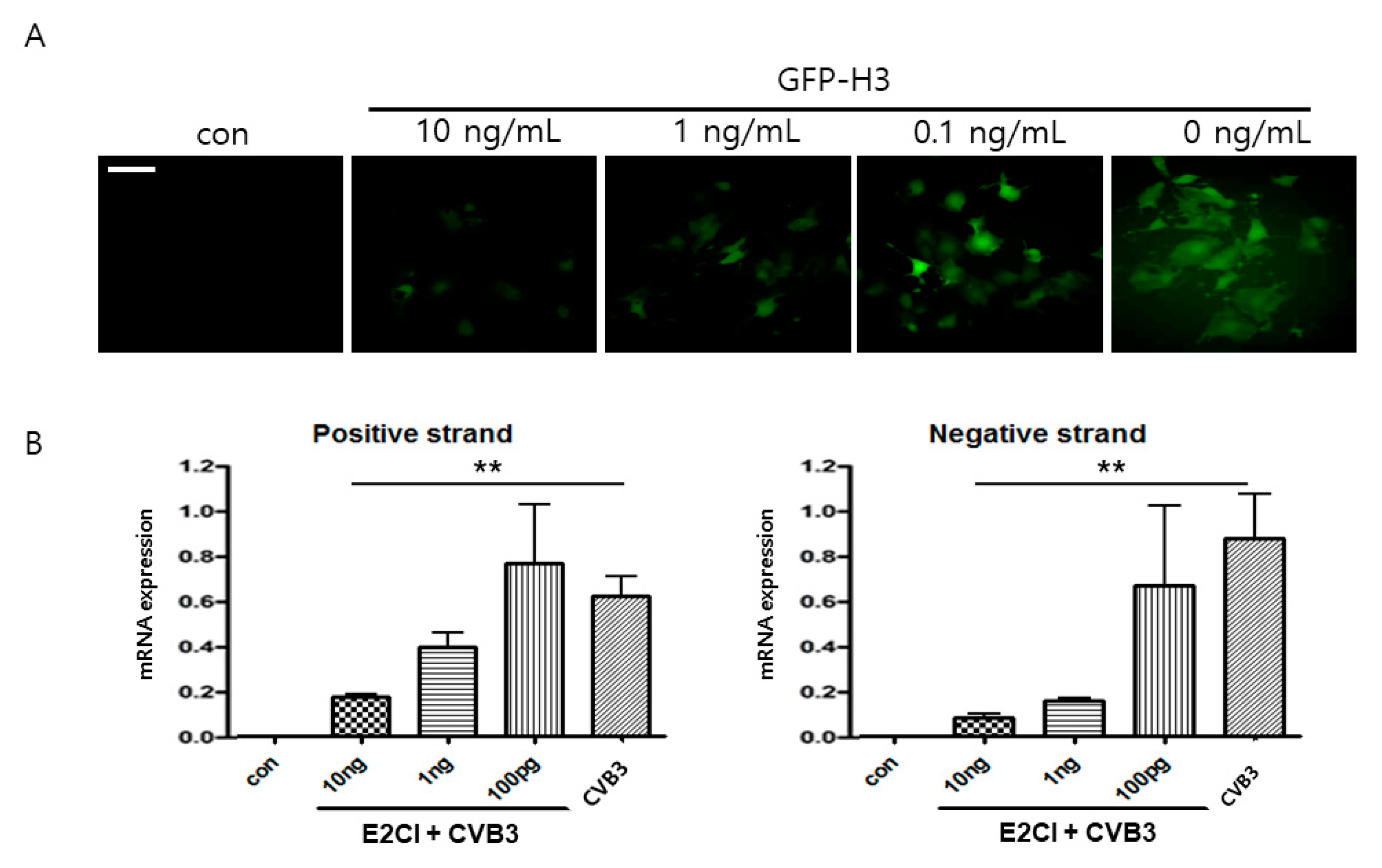
2.6. E2CI Inhibits CVB3 Replication in iPS Human Cardiomyocyte
3. Discussion
4. Materials and Methods
4.1. Virus and Cell Lines
4.2. Screening Antiviral Effect Chemical
4.3. Viral Myocarditis Murine Model
4.4. Human iPS Cardiomyocytes Cell Culture
4.5. Viral Genome Amplification
4.6. Histopathology and Organ Virus Titers
4.7. Mouse Echocardiography
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yun, S.H.; Lee, W.G.; Kim, Y.C.; Ju, E.S.; Lim, B.K.; Choi, J.O.; Kim, D.K.; Jeon, E.S. Antiviral activity of coxsackievirus B3 3C protease inhibitor in experimental murine myocarditis. J. Infect. Dis. 2012, 205, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Baboonian, C.; Davies, M.J.; Booth, J.C.; McKenna, W.J. Coxsackie B viruses and human heart disease. Curr. Top. Microbiol. Immunol. 1997, 223, 31–52. [Google Scholar] [PubMed]
- Herskowitz, A.; Beisel, K.W.; Wolfgram, L.J.; Rose, N.R. Coxsackievirus B3 murine myocarditis: Wide pathologic spectrum in genetically defined inbred strains. Hum. Pathol. 1985, 16, 671–673. [Google Scholar] [CrossRef]
- Feldman, A.M.; McNamara, D. Myocarditis. N. Engl. J. Med. 2000, 343, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, K.U.; Badorff, C. The immune system in viral myocarditis: Maintaining the balance. Circ. Res. 1999, 85, 559–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowlton, K.U.; Jeon, E.S.; Berkley, N.; Wessely, R.; Huber, S. A mutation in the puff region of VP2 attenuates the myocarditic phenotype of an infectious cDNA of the Woodruff variant of coxsackievirus B3. J. Virol. 1996, 70, 7811–7818. [Google Scholar] [CrossRef] [Green Version]
- Badorff, C.; Lee, G.H.; Lamphear, B.J.; Martone, M.E.; Campbell, K.P.; Rhoads, R.E.; Knowlton, K.U. Enteroviral protease 2A cleaves dystrophin: Evidence of cytoskeletal disruption in an acquired cardiomyopathy. Nat. Med. 1999, 5, 320–326. [Google Scholar] [CrossRef]
- Esfandiarei, M.; Luo, H.; Yanagawa, B.; Suarez, A.; Dabiri, D.; Zhang, J.; McManus, B.M. Protein kinase B/Akt regulates coxsackievirus B3 replication through a mechanism which is not caspase dependent. J. Virol. 2004, 78, 4289–4298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racaniello, V.R. Chapter24, The Picornaviridae; The Viruses and Their Replication. Fields Virology, 5th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 1, p. 829. [Google Scholar]
- Lim, B.K.; Choe, S.C.; Shin, J.O.; Ho, S.H.; Kim, J.M.; Yu, S.S.; Kim, S.; Jeon, E.S. Local expression of interleukin-1 receptor antagonist by plasmid DNA improves mortality and decreases myocardial inflammation in experimental coxsackieviral myocarditis. Circulation 2002, 105, 1278–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, B.K.; Choi, J.H.; Nam, J.H.; Gil, C.O.; Shin, J.O.; Yun, S.H.; Kim, D.K.; Jeon, E.S. Virus receptor trap neutralizes coxsackievirus in experimental murine viral myocarditis. Cardiovasc. Res. 2006, 71, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.K.; Yun, S.H.; Ju, E.S.; Kim, B.K.; Lee, Y.J.; Yoo, D.K.; Kim, Y.C.; Jeon, E.S. Soluble coxsackievirus B3 3C protease inhibitor prevents cardiomyopathy in an experimental chronic myocarditis murine model. Virus Res. 2015, 199, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.M.; Lim, B.K.; Ho, S.H.; Yun, S.H.; Shin, J.O.; Park, E.M.; Kim, D.K.; Kim, S.; Jeon, E.S. TNFR-Fc fusion protein expressed by in vivo electroporation improves survival rates and myocardial injury in coxsackievirus induced murine myocarditis. Biochem. Biophys. Res. Commun. 2006, 344, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.J. From birth to death: The complex lives of eukaryotic mRNAs. Science 2005, 309, 1514–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanner, N.K.; Linder, P. DExD/H box RNA helicases: From generic motors to specific dissociation functions. Mol. Cell 2001, 8, 251–262. [Google Scholar] [CrossRef]
- Jankowsky, E.; Gross, C.H.; Shuman, S.; Pyle, A.M. Active disruption of an RNA-protein interaction by a DExH/D RNA helicase. Science 2001, 291, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Ulvila, J.; Hultmark, D.; Ramet, M. RNA silencing in the antiviral innate immune defence—Role of DEAD-box RNA helicases. Scand. J. Immunol. 2010, 71, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Ranji, A.; Boris-Lawrie, K. RNA helicases: Emerging roles in viral replication and the host innate response. RNA Biol. 2010, 7, 775–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yedavalli, V.S.; Zhang, N.; Cai, H.; Zhang, P.; Starost, M.F.; Hosmane, R.S.; Jeang, K.T. Ring expanded nucleoside analogues inhibit RNA helicase and intracellular human immunodeficiency virus type 1 replication. J. Med. Chem. 2008, 51, 5043–5051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinkert, S.; Westermann, D.; Wang, X.; Klingel, K.; Dorner, A.; Savvatis, K.; Grossl, T.; Krohn, S.; Tschope, C.; Zeichhardt, H.; et al. Prevention of cardiac dysfunction in acute coxsackievirus B3 cardiomyopathy by inducible expression of a soluble coxsackievirus-adenovirus receptor. Circulation 2009, 120, 2358–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, S.-H.; Shin, H.-H.; Ju, E.-S.; Lee, Y.-J.; Lim, B.-K.; Jeon, E.-S. Inhibition of RNA Helicase Activity Prevents Coxsackievirus B3-Induced Myocarditis in Human iPS Cardiomyocytes. Int. J. Mol. Sci. 2020, 21, 3041. https://doi.org/10.3390/ijms21093041
Yun S-H, Shin H-H, Ju E-S, Lee Y-J, Lim B-K, Jeon E-S. Inhibition of RNA Helicase Activity Prevents Coxsackievirus B3-Induced Myocarditis in Human iPS Cardiomyocytes. International Journal of Molecular Sciences. 2020; 21(9):3041. https://doi.org/10.3390/ijms21093041
Chicago/Turabian StyleYun, Soo-Hyeon, Ha-Hyeon Shin, Eun-Seon Ju, You-Jung Lee, Byung-Kwan Lim, and Eun-Seok Jeon. 2020. "Inhibition of RNA Helicase Activity Prevents Coxsackievirus B3-Induced Myocarditis in Human iPS Cardiomyocytes" International Journal of Molecular Sciences 21, no. 9: 3041. https://doi.org/10.3390/ijms21093041