Inhibitory Effective Perturbations of Cilobradine (DK-AH269), A Blocker of HCN Channels, on the Amplitude and Gating of Both Hyperpolarization-Activated Cation and Delayed-Rectifier Potassium Currents


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
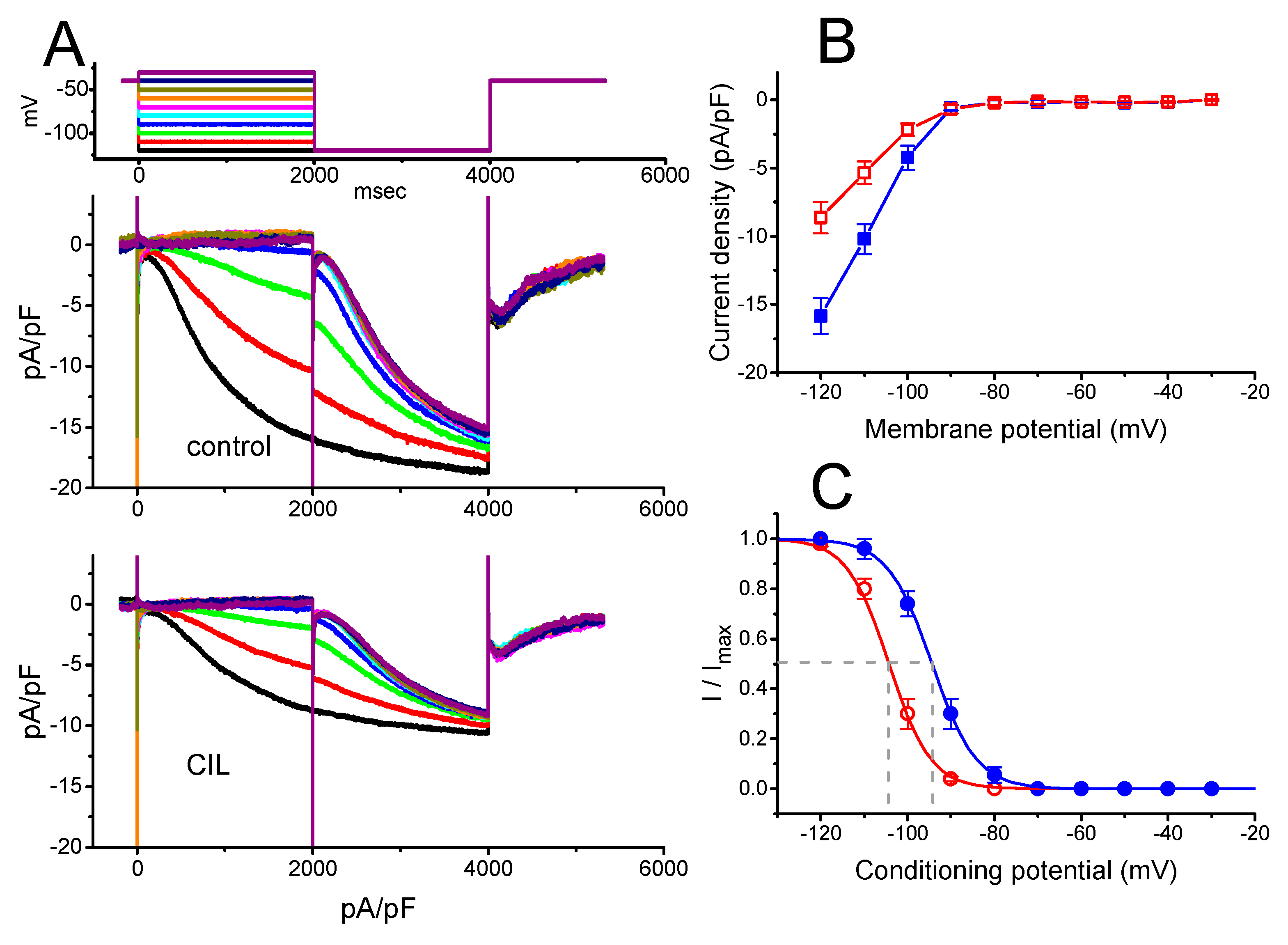
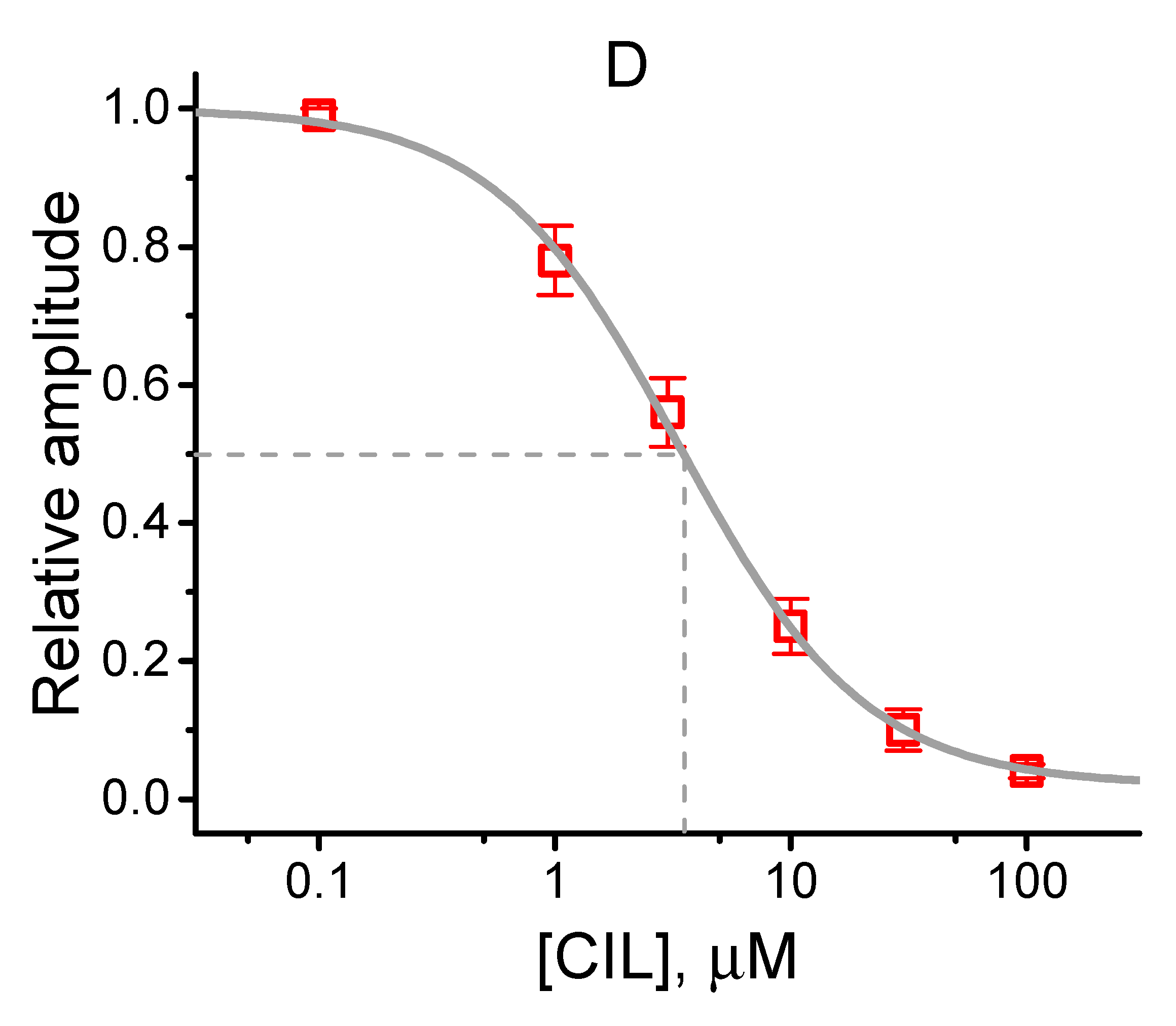
2.1. Effects of CIL on Hyperpolarization-Activated Cation Current (Ih) Recorded from Pituitary Tumor (GH3) Cells
2.2. Effects of CIL on the Impedance Magnitude (IM) as a Function of Frequency (i.e., IM Plot)
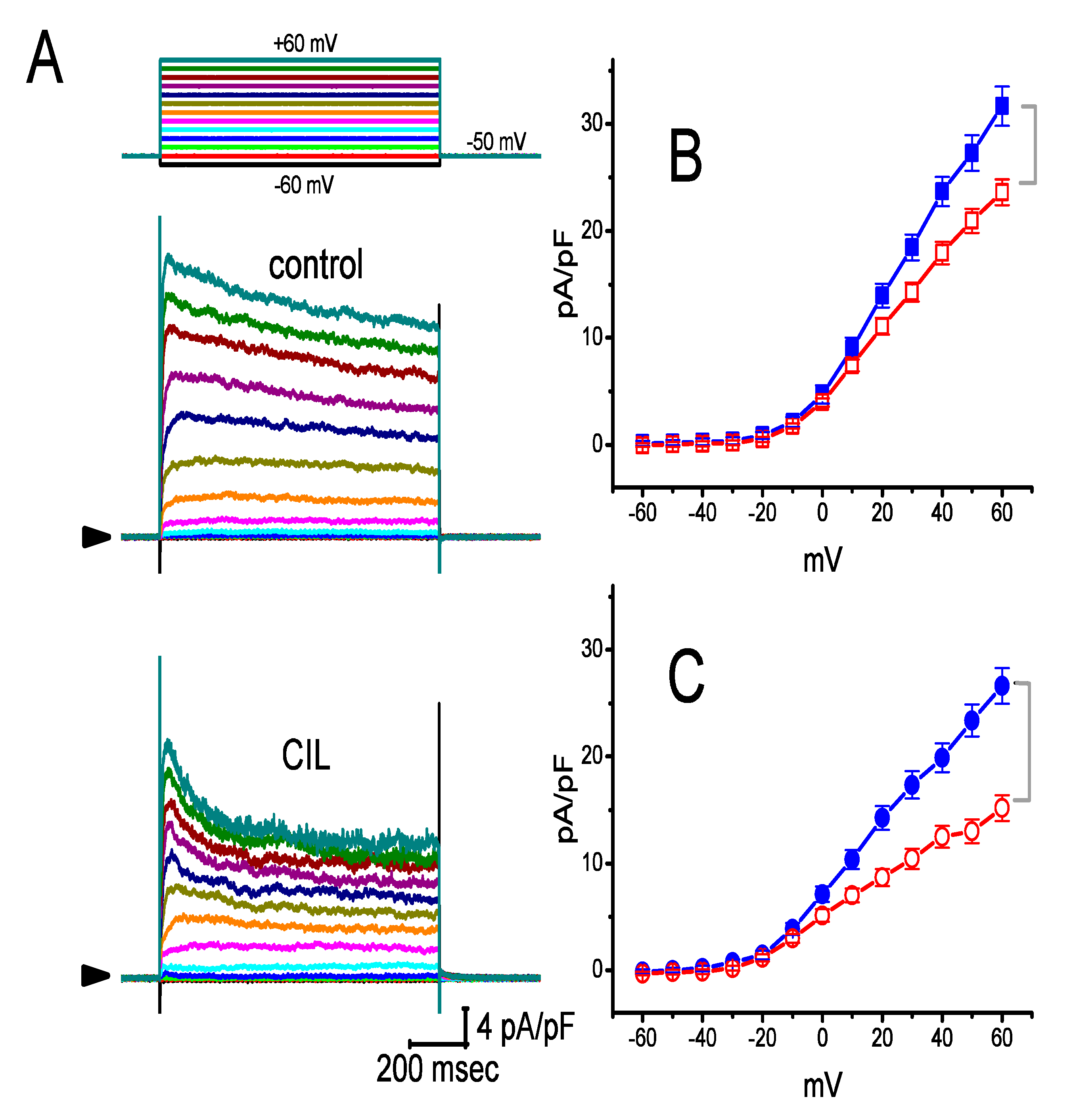
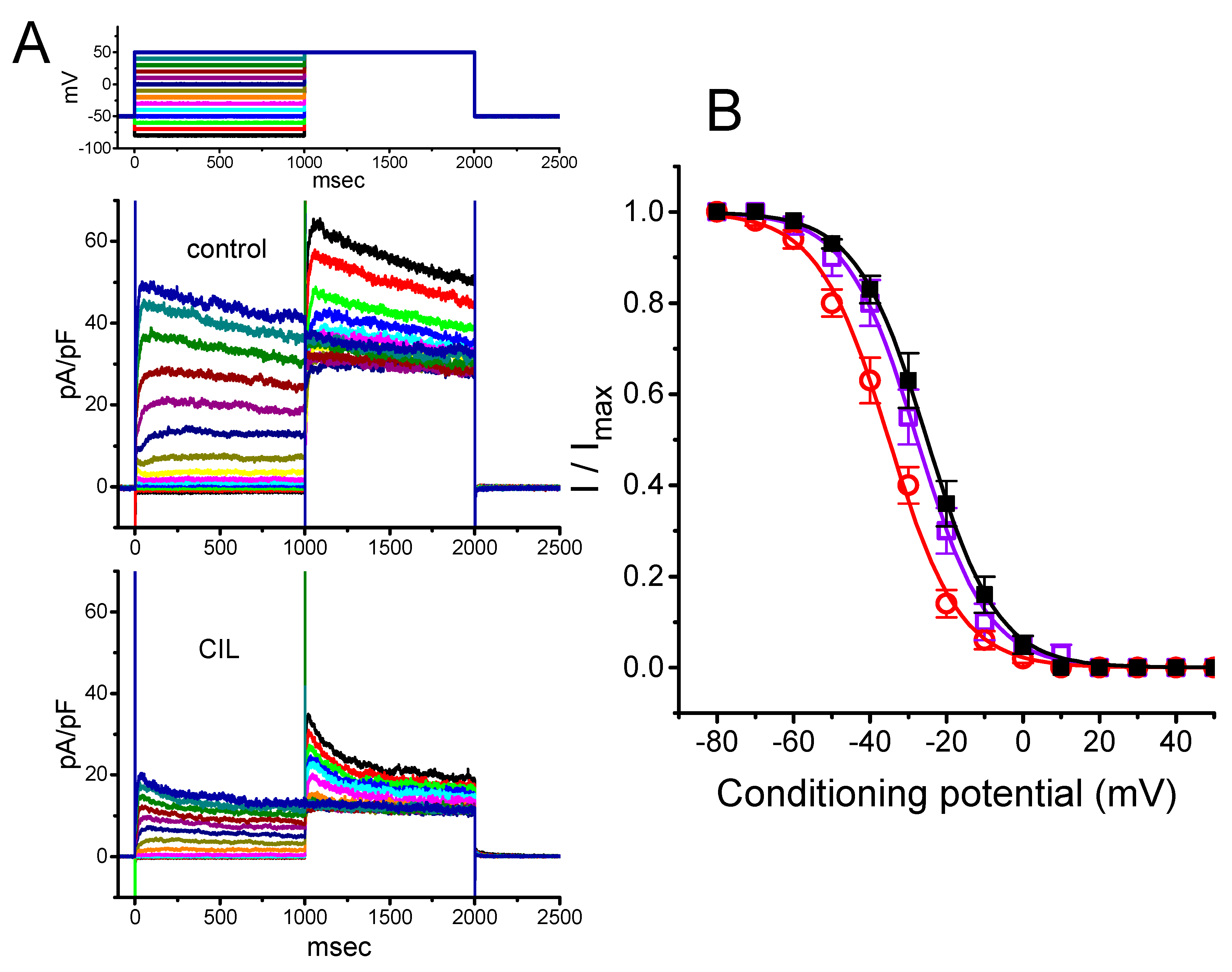
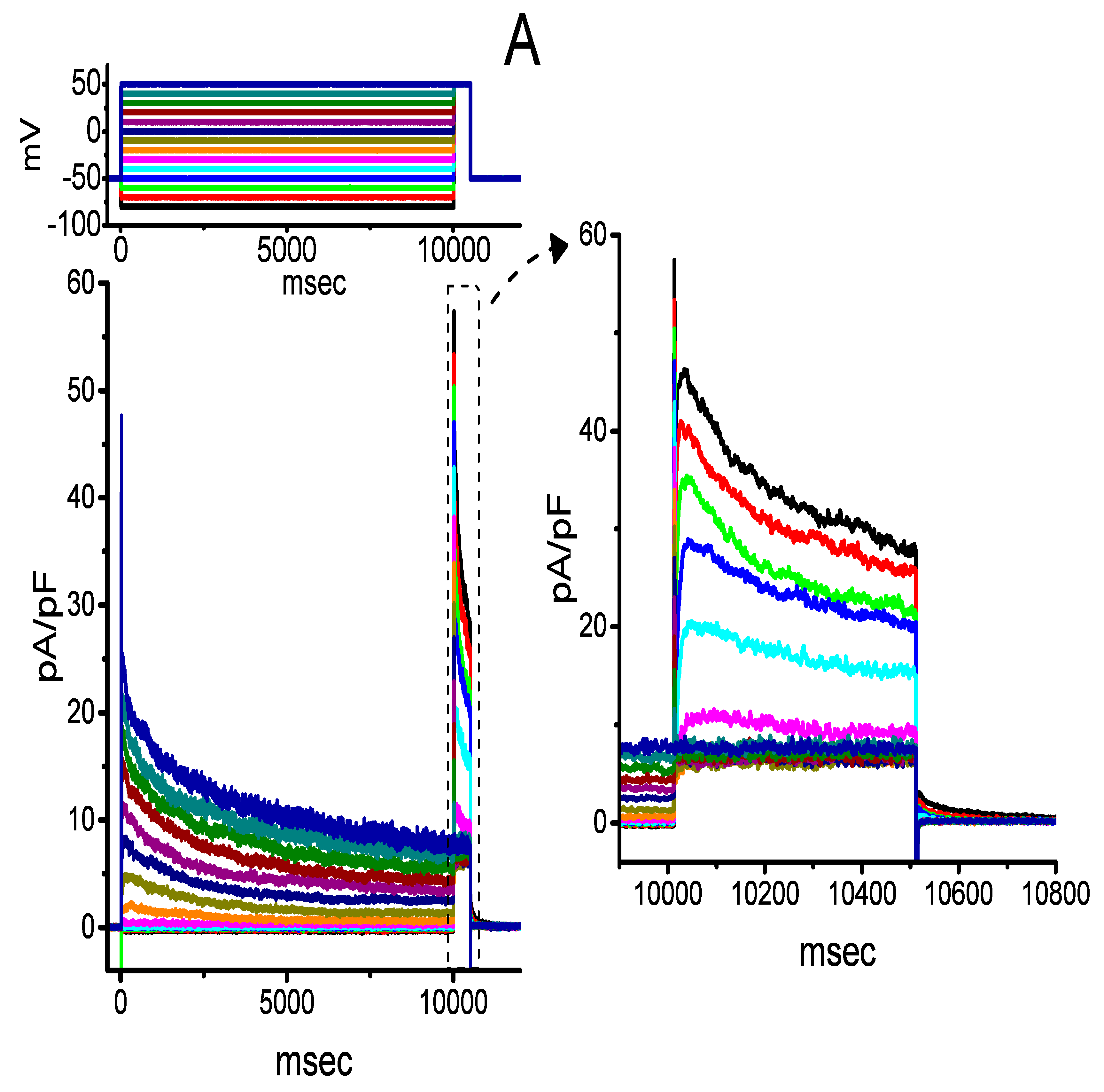
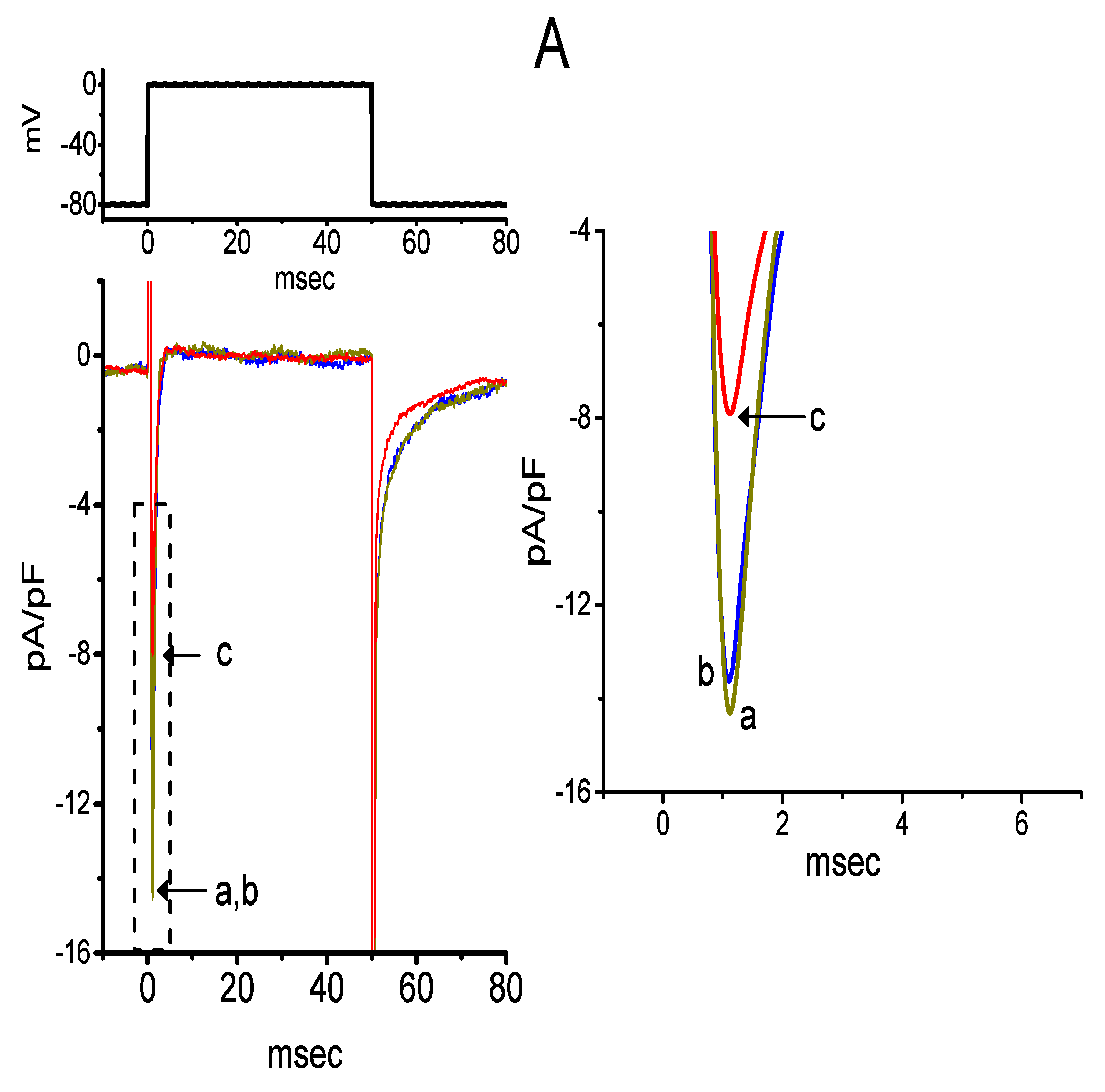
2.3. Averaged Current Density Versus Voltage Relationship of IK(DR) Produced by the Presence of CIL
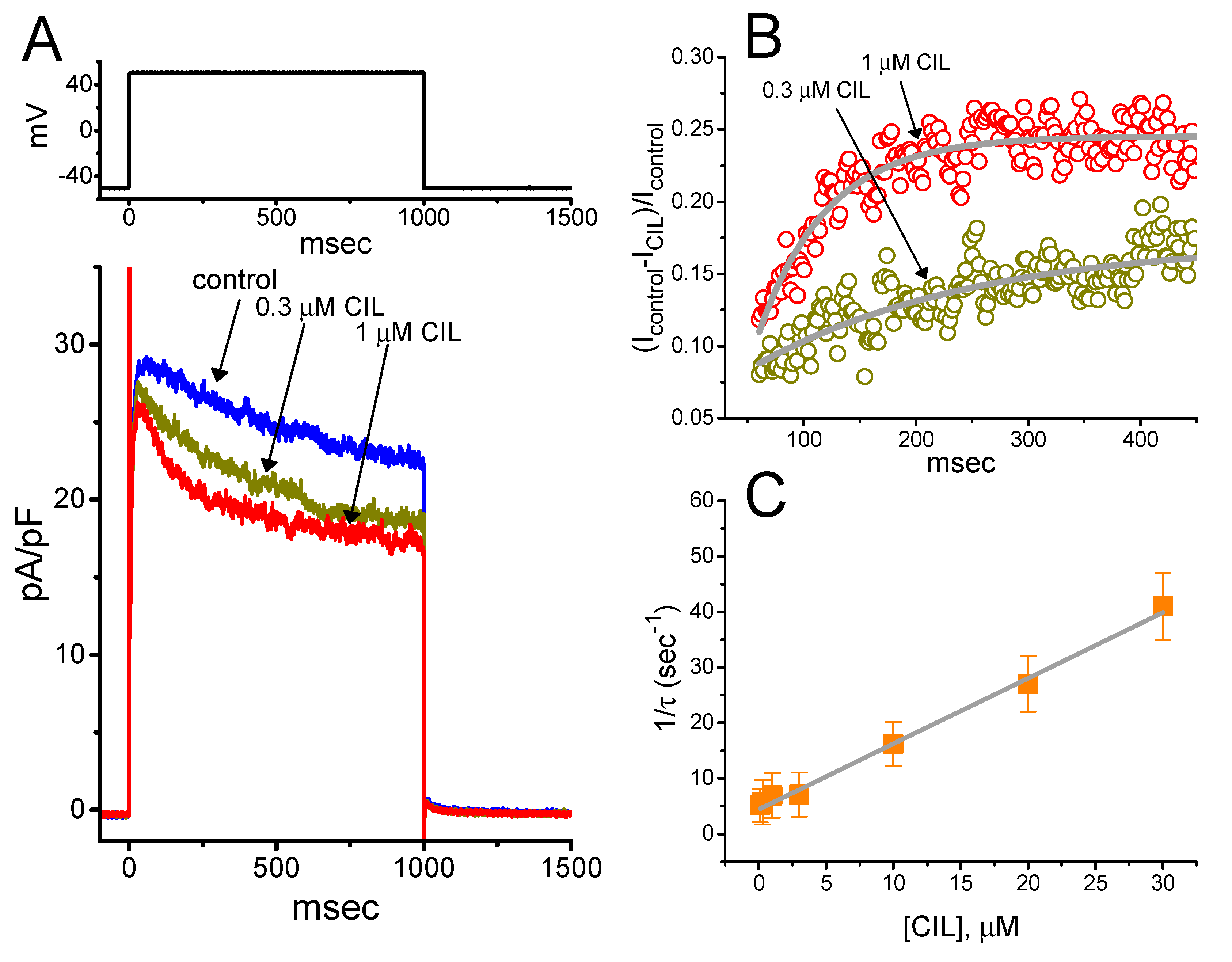
2.4. Kinetic Evaluation of the CIL-Inhibited IK(DR) Observed in GH3 Cells
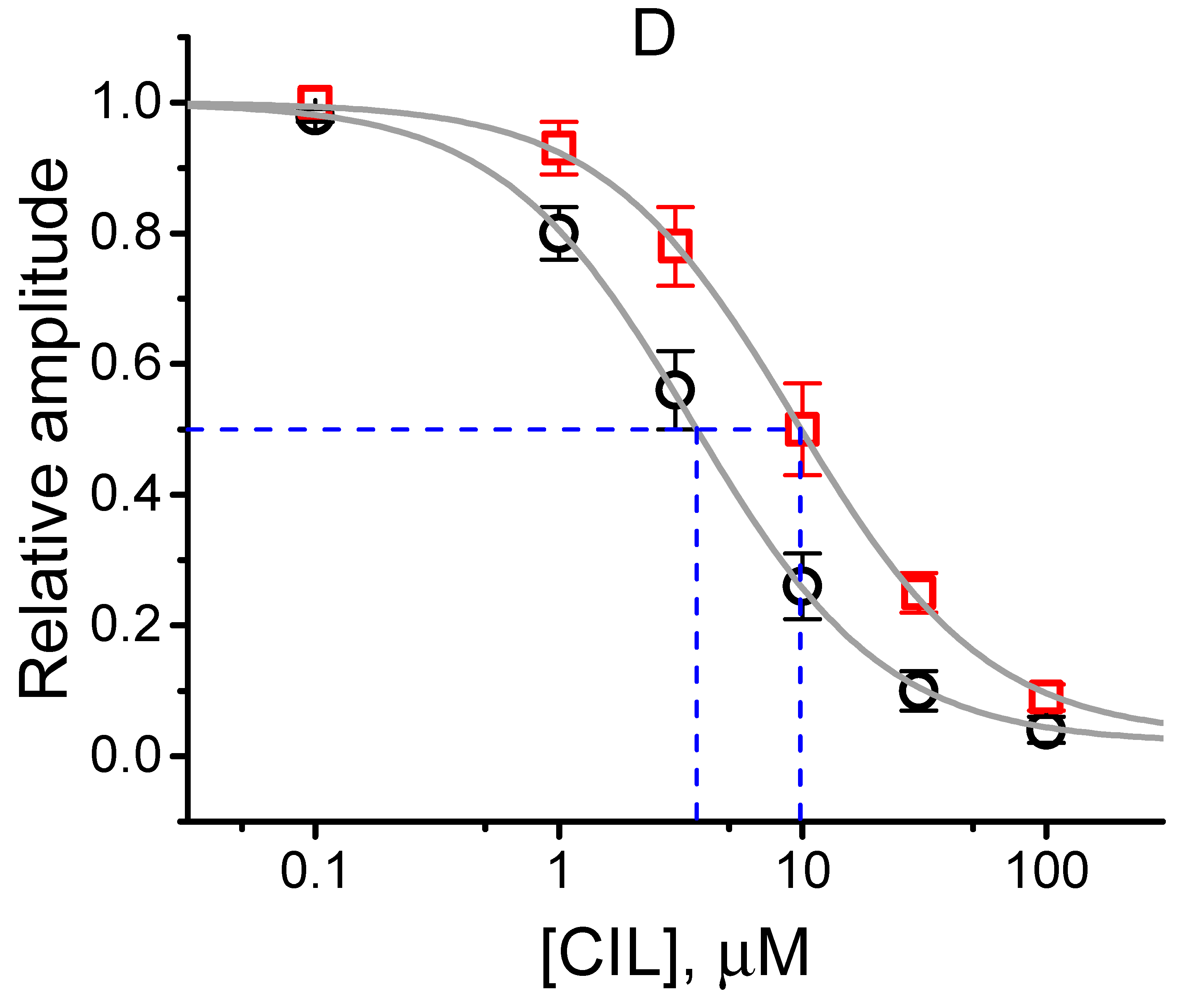
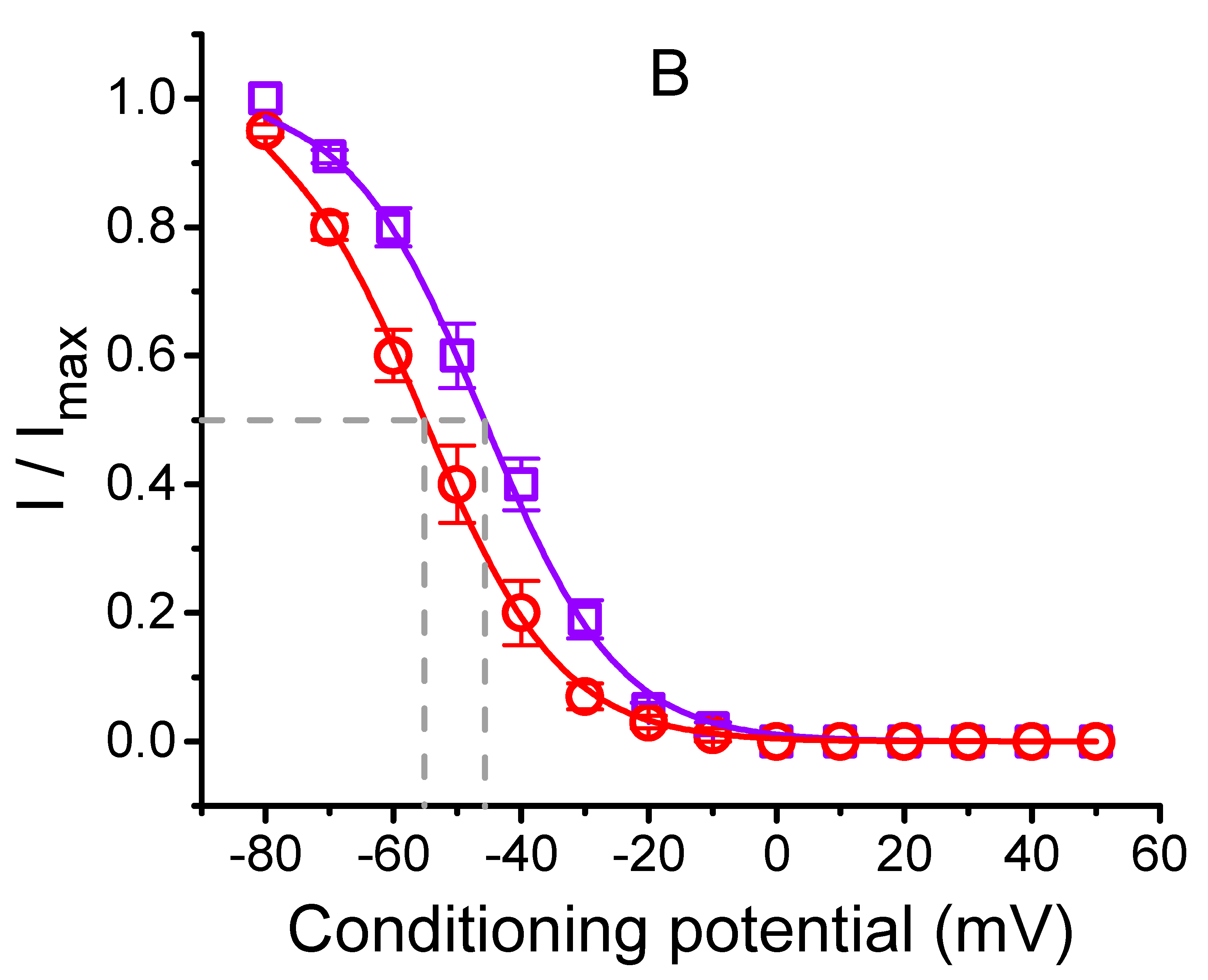
2.5. Steady-State Inactivation Curve of IK(DR) Obtained in the Presence of CIL
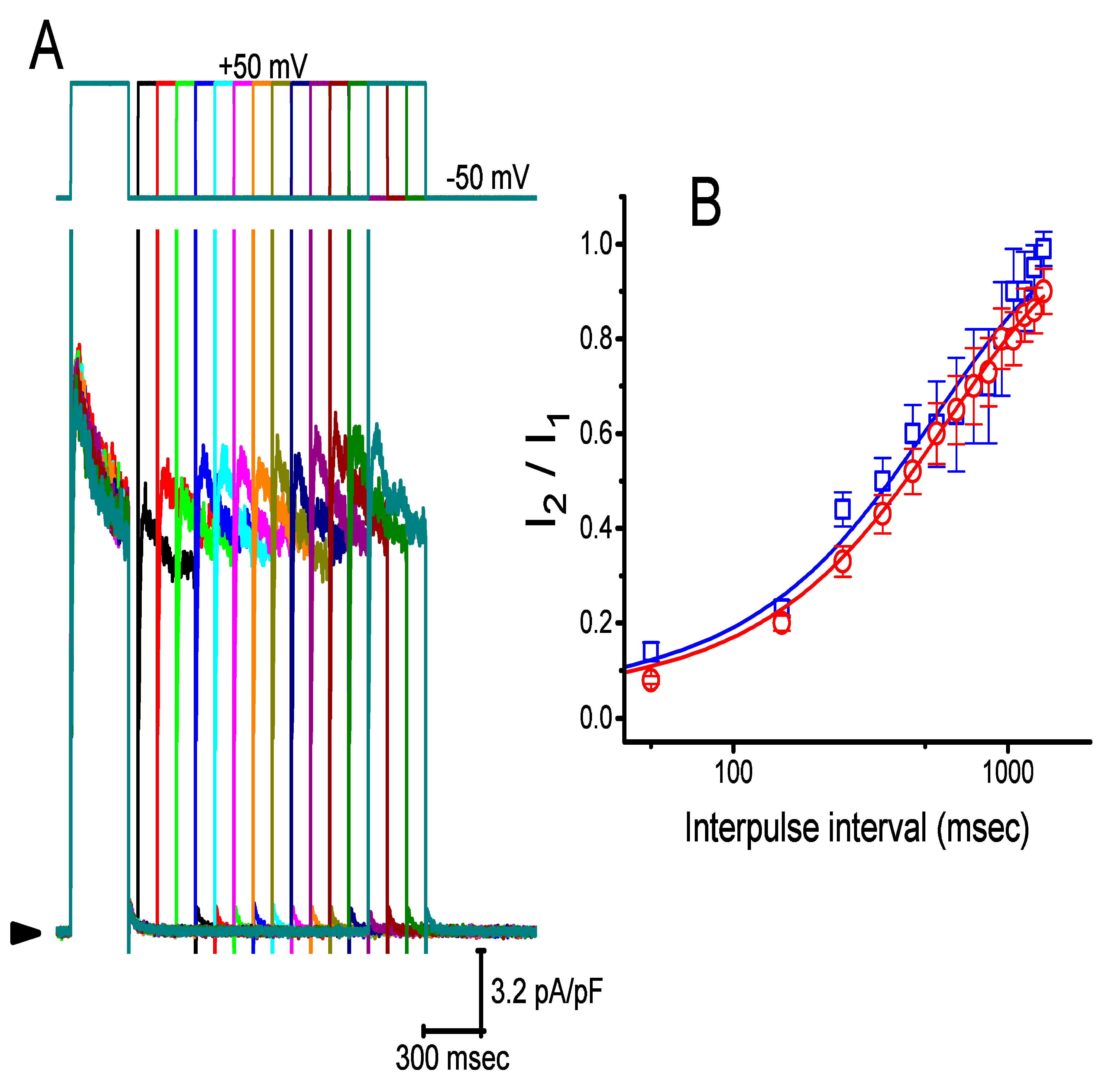
2.6. Effect of CIL on Recovery of IK(DR) from Inactivation in GH3 Cells
2.7. Inhibitory Effect of CIL on the Steady-State Inactivation Curve of IK(DR) Evoked by 10-s Conditioning Pulses
2.8. Failure of CIL to Suppress Voltage-Gated Na+ Current (INa) Evoked by Brief Depolarization
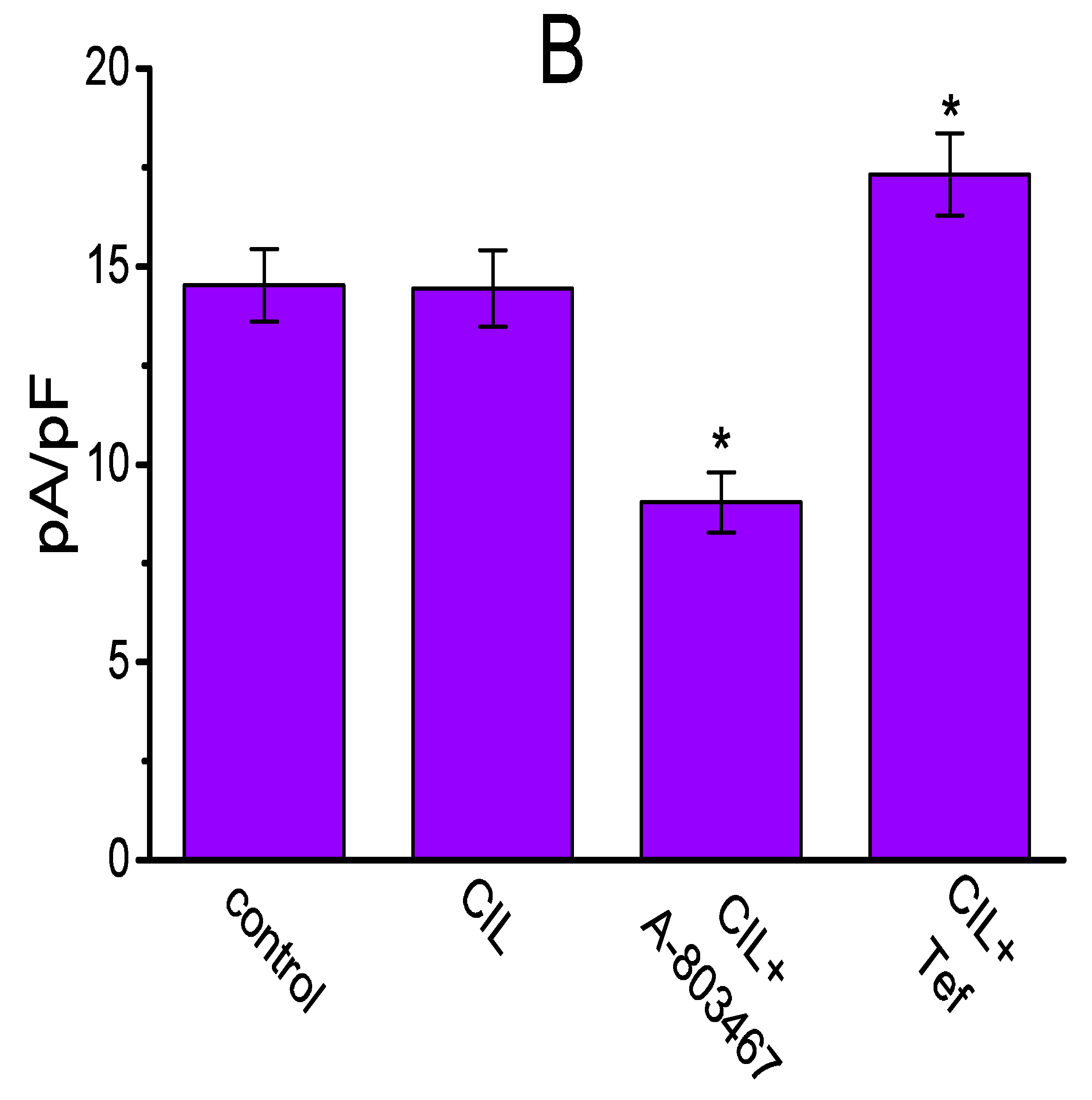
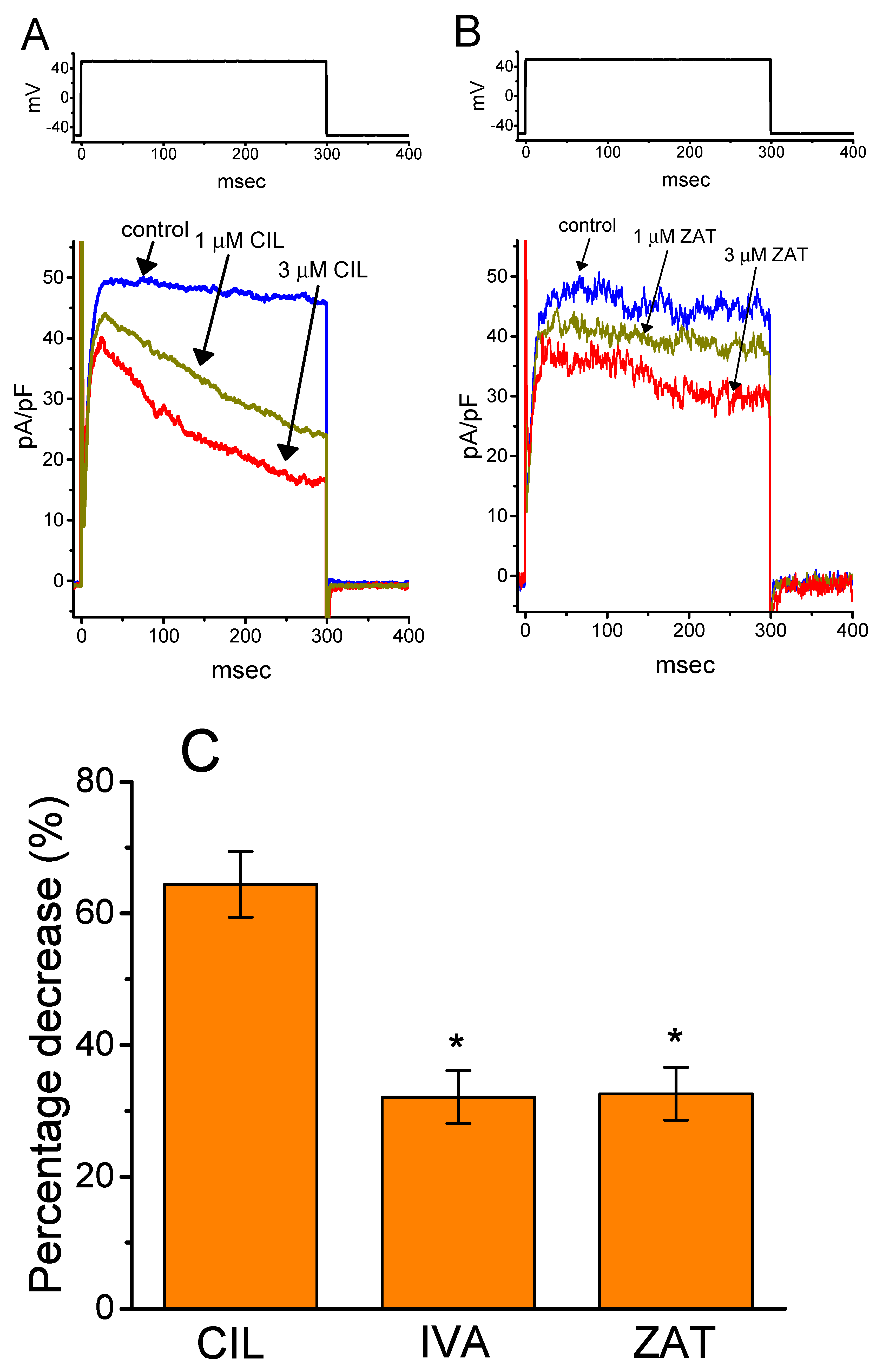
2.9. Effect of CIL, Ivabradine, or Zatebradine on IK(DR) Recorded from Heart-Derived H9c2 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals, Drugs, and Solutions
4.2. Cell Preparations
4.3. Electrophysiological Measurements
4.4. Data Recordings
4.5. Data Analyses
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CIL | Cilobradine, DK-AH269, or |
| fR | resonance frequency |
| HCN channel | hyperpolarization-activated cyclic nucleotide-gated channel |
| IC50 | the concentration for 50% inhibition |
| Ih | hyperpolarization-activated cation current |
| IK(DR) | delayed-rectifier K+ current |
| IM | impedance magnitude |
| INa | voltage-gated Na+ current |
| IVA | Ivabradine |
| KD | dissociation constant |
| KV channel | voltage-gated K+ channel |
| Tef | Tefluthrin |
| τ | time constant |
| TEA | tetraethylammonium chloride |
| Z | Impedance |
| ZAT | Zatebradine |
References
- Schmitz-Spanke, S.; Granetzny, A.; Stoffels, B.; Pomblum, V.J.; Gams, E.; Schipke, J.D. Effects of a bradycardic agent on postischemic cardiac recovery in rabbits. J. Physiol. Pharmacol. 2003, 55, 705–712. [Google Scholar]
- Stieber, J.; Wieland, K.; Stöckl, G.; Ludwig, A.; Hofmann, F. Bradycardic and proarrhythmic properties of sinus node inhibitors. Mol. Pharmacol. 2006, 69, 1328–1337. [Google Scholar] [CrossRef] [Green Version]
- Boldt, A.; Gergs, U.; Pönicke, K.; Simm, A.; Silber, R.E.; Neumann, J. Inotropic effects of ivabradine in the mammalian heart. Pharmacology 2010, 86, 249–258. [Google Scholar] [CrossRef]
- El-Kholy, W.; MacDonald, P.E.; Fox, J.M.; Bhattacharjee, A.; Xue, T.; Gao, X.; Zhang, Y.; Stieber, J.; Li, R.A.; Tsushima, R.G.; et al. Hyperpolarization-activated cyclic neucleotide-gated channels in pancreatic beta-cells. Mol. Endocrinol. 2007, 21, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Qu, J.; Hardy, A.; Zhang, N.; Diao, J.; Strijbos, P.J.; Tsushima, R.; Robinson, R.B.; Gaisano, H.Y.; et al. Functional characterization of HCN channels in rat pancreatic β cells. J. Endocrinol. 2009, 203, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, N.; Gyulkhandanyan, A.V.; Xu, E.; Gaisano, H.Y.; Wheeler, M.B.; Wang, Q. Presence of functional hyperpolarisation-activated cyclic nucleotide-gated channels in clonal alpha cell lines and rat islet alpha cells. Diabetologia 2008, 51, 2290–2298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irisawa, H.; Brown, H.F.; Giles, W. Cardiac pacemaking in the sinoatrial node. Physiol. Rev. 1993, 73, 197–227. [Google Scholar] [CrossRef]
- Biel, M.; Wahl-Schott, C.; Michalakis, S.; Zong, X. Hyperpolarization-activated cation channels: From genes to function. Physiol. Rev. 2009, 89, 847–885. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.C.; Wang, Y.J.; Wu, P.Y.; Wu, S.N. Tramadol-induced block of hyperpolarization-activated cation current in rat pituitary lactotrophs. Naunyn Schmiedebergs Arch. Pharmacol. 2009, 379, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Stojilkovic, S.S.; Tabak, J.; Bertram, R. Ion channels and signaling in the pituitary gland. Endoc. Rev. 2010, 31, 845–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretschmannova, K.; Kucka, M.; Gonzalez-Iglesias, A.E.; Stojilkovic, S.S. The expression and role of hyperpolarization-activated and cyclic nucleotide-gated channels in endocrine anterior pituitary cells. Mol. Endocrinol. 2012, 26, 153–164. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Chen, F.; Li, B.; Hu, Z. Neurophysiology of HCN channels: From cellular functions to multiple regulations. Prog. Neurobiol. 2014, 112, 1–23. [Google Scholar] [CrossRef]
- Spinelli, V.; Sartiani, L.; Mugelli, A.; Romanelli, M.N.; Cerbai, E. Hyperpolarization-activated cyclic-nucleotide-gated channels: Pathophysiological developmental and pharmacological insights into their function in cellular excitability. Can. J. Physiol. Pharmacol. 2018, 96, 977–984. [Google Scholar] [CrossRef]
- Chang, W.T.; Gao, Z.H.; Lo, Y.C.; Wu, S.N. Evidence for effective inhibitory actions on hyperpolarization-activated cation current caused by Ganoderma triterpenoids, the main active constituents of Ganoderma spores. Molecules 2019, 24, 4256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, H.T.; Liu, Y.C.; Liu, P.Y.; Wu, S.N. Concerted suppression of Ih and activation of IK(M) by ivabradine, an HCN-channel inhibitor, in pituitary cells and hippocampal neurons. Brain Res. Bullet. 2019, 149, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Tanguay, J.; Callahan, K.M.; D’Avanzo, N. Characterization of drug binding within the HCN1 channel pore. Sci. Rep. 2019, 9, 465. [Google Scholar] [CrossRef]
- Van Bogaert, P.P.; Pittoors, F. Use-dependent blockade of cardiac pacemaker current (If) by cilobradine and zatebradine. Eur. J. Pharmacol. 2003, 478, 161–171. [Google Scholar] [CrossRef]
- Bucchi, A.; Barbuti, A.; Baruscotti, M.; DiFrancesco, D. Heart rate reduction via selective ‘funny’ channel blockers. Curr. Opin. Pharmacol. 2007, 7, 208–213. [Google Scholar] [CrossRef]
- Novella Romanelli, M.; Sartiani, L.; Masi, A.; Mannaioni, G.; Manetti, D.; Mugelli, A.; Cerbai, E. HCN channels modulators: The need for selectivity. Curr. Top. Med. Chem. 2016, 16, 1764–1791. [Google Scholar] [CrossRef]
- DiFrancesco, J.C.; DiFranscesco, D. Dysfunctional HCN ion channels in neurological diseases. Front. Cell. Neurosci. 2015, 6, 174. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Lin, M.W.; Lin, A.A.; Peng, H.; Wu, S.N. Evidence for state-dependent block of DPI 201-106, a synthetic inhibitor of Na+ channel inactivation, on delayed-rectifier K+ current in pituitary tumor (GH3) cells. J. Physiol. Pharmacol. 2008, 59, 409–423. [Google Scholar]
- Fletcher, P.A.; Sherman, A.; Stojilkovic, S.S. Common and diverse elements of ion channels and receptors underlying electrical activity in endocrine pituitary cells. Mol. Cell. Endocrinol. 2018, 463, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.L.; Chang, W.T.; Chan, C.H.; Wu, S.N. Evidence for effective multiple K+-current inhibitions by tolvaptan, a non-peptide antagonist of vasopressin V2 receptor. Front. Pharmacol. 2019, 10, 76. [Google Scholar] [CrossRef]
- So, E.C.; Liu, P.Y.; Lee, C.C.; Wu, S.N. High effectiveness in actions of cardilzomib on delayed-rectifier K+ current and on spontaneous action potentials. Front. Pharmacol. 2019, 10, 1163. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.H.; Volk, T.; Ehmke, H. Heterogeneity of Kv2.1 mRNA expression and delayed rectifier current in single isolated myocytes from rat left ventricle. Circ. Res. 2001, 88, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.T.; Lo, Y.C.; Gao, Z.H.; Wu, S.N. Evidence for the capability of roxadustat (FG-4592), an oral HIF prolyl-hydroxylase inhibitor, to perturb membrane ionic currents: An unidentified yet important action. Int. J. Mol. Sci. 2019, 20, 6027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.N.; Chern, J.H.; Shen, S.; Chen, H.H.; Hsu, Y.T.; Lee, C.C.; Chan, M.H.; Lai, M.C.; Shie, F.S. Stimulatory actions of a novel thiourea derivative on large-conductance, calcium-activated potassium channels. J. Cell. Physiol. 2017, 232, 3409–3421. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.C.; Ruan, J.S.; Wu, S.N. Evidence of decreased activity in intermediate-conductance calcium-activated potassium channels during retinoic acid-induced differentiation in motor neuron-like NSC-34 cells. Cell. Physiol. Biochem. 2018, 48, 2374–2388. [Google Scholar] [CrossRef] [PubMed]
- Gutfreund, Y.; Yarom, Y.; Segev, I. Subthreshold oscillations and resonant frequency in guinea-pig cortical neurons: Physiology and modeling. J. Physiol. 1995, 483.3, 621–640. [Google Scholar] [CrossRef]
- Buskila, Y.; Kékesi, O.; Bellot-Saez, A.; Seah, W.; Berg, T.; Trpceski, M.; Yerbury, J.J.; Ooi, L. Dynamic interplay between H-current and M-current controls motoneuron hyperexcitability in amyotrophic lateral sclerosis. Cell. Death Dis. 2019, 10, 310. [Google Scholar] [CrossRef]
- Rush, A.M.; Cummins, T.R. Painful research: Identification of a small-molecule inhibitor that selectively targets Nav1.8 sodium channels. Mol. Interv. 2007, 7, 192–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, M.C.; Tseng, R.C.; Huang, C.W.; Wu, S.N. The novel direct modulatory effects of perampanel, an antagonist of AMPA receptors, on voltage-gated sodium and M-type potassium currents. Biomolecules 2019, 9, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bois, P.; Guinamard, R.; Chemaly, A.E.; Faivre, J.F.; Bescond, J. Molecular regulation and pharmacology of pacemaker channels. Curr. Pharm. Des. 2007, 13, 2338–2349. [Google Scholar] [CrossRef] [PubMed]
- Kranig, S.A.; Duhme, N.; Waldeck, C.; Draguhn, A.; Reichinnek, S.; Both, M. Different functions of hyperpolarization-activated cation channels for hippocampal sharp waves and ripples in vitro. Neuroscience 2013, 228, 325–333. [Google Scholar] [CrossRef]
- Maccarone, R.; Izzizzari, G.; Gargini, C.; Cervetto, L.; Bisti, S. The impact of organic inhibitors of the hyperpolarization activated current (Ih) on the electroretinogram (ERG) of rodents. Arch. Ital. Biol. 2004, 142, 95–103. [Google Scholar]
- Hellbach, A.; Tiozzo, S.; Ohn, J.; Liebling, M.; De Tomaso, A.W. Characterization of the HCN and cardiac function in a colonial ascidian. J. Exp. Zool. A Ecol. Genet. Physiol. 2011, 315, 476–486. [Google Scholar] [CrossRef]
- Vélez de Mendizábal, N.; Staab, A.; Schäfer, H.G.; Trommeshauser, D.; Döge, C.; Klüglich, M.; Roberts, J.; Trocóniz, I.F. Joint population pharmacokinetic/pharmacodynamic model for the heart rate effects at rest and at the end of exercise for cilobradine. Pharm. Res. 2013, 30, 1110–1122. [Google Scholar] [CrossRef]
- Proenza, C.; Angoli, D.; Agranovich, E.; Macri, V.; Accili, E.A. Pacemaker channels produce an instantaneous current. J. Biol. Chem. 2002, 277, 5101–5109. [Google Scholar] [CrossRef] [Green Version]
- Kuo, P.C.; Yang, C.J.; Lee, Y.C.; Chen, P.C.; Liu, Y.C.; Wu, S.N. The comprehensive electrophysiological study of curcuminoids on delayed-rectifier K+ currents in insulin-secreting cells. Eur. J. Pharmacol. 2018, 819, 233–241. [Google Scholar] [CrossRef]
- Lo, Y.C.; Yang, S.R.; Huang, M.H.; Liu, Y.C.; Wu, S.N. Characterization of chromanol 293B-induced block of the delayed-rectifier K+ current in heart-derived H9c2 cells. Life Sci. 2005, 76, 2275–2286. [Google Scholar] [CrossRef]
- Wang, Y.J.; Chen, B.S.; Lin, M.W.; Lin, A.A.; Peng, H.; Sung, R.J.; Wu, S.N. Time-dependent block of ultrarapid-delayed rectifier K+ currents by aconitine, a potent cardiodoxin, in heart-derived H9c2 myoblasts and in neonatal rat ventricular myocytes. Toxicol. Sci. 2008, 106, 454–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dini, L.; Del Lungo, M.; Resta, F.; Melchiorre, M.; Spinelli, V.; Di Cesare Mannelli, L.; Ghelardini, C.; Laurino, A.; Sartiani, L.; Coppini, R.; et al. Selective blockade of HCN1/HCN2 channels as a potential pharmacological strategy against pain. Front. Pharmacol. 2018, 9, 1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resta, F.; Micheli, L.; Laurino, A.; Spnelli, V.; Mello, T.; Sartiani, L.; Di Cesare Mannelli, L.; Cerbai, E.; Ghelardini, C.; Romanelli, M.N.; et al. Selective HCN1 block as a strategy to control oxaliplatin-induced neuropathy. Neuropharmacology 2018, 131, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, G.C.; Risner-Janiczek, J.R.; Jones, S.M.; Holt, J.R. HCN channels expressed in the inner ear are necessary for normal balance function. J. Neurosci. 2011, 31, 16814–16825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zobeiri, M.; Chaudhary, R.; Blaich, A.; Rottmann, M.; Herrmann, S.; Meuth, P.; Bista, P.; Kanyshkova, T.; Lüttjohann, A.; Narayanan, V.; et al. The hyperpolarization-activated HCN channel is important for proper maintenance of oscillatory activity in the thalamocortical system. Cereb. Cortex 2019, 29, 2291–2304. [Google Scholar] [CrossRef] [Green Version]
- Chambers, A.R.; Pilati, N.; Balaram, P.; Large, C.H.; Kaczmarek, L.K.; Polley, D.B. Pharmacological modulation of Kv3.1 mitigates auditory midbrain temporal processing deficits following auditory nerve damage. Sci. Rep. 2017, 7, 17496. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.H.; Liu, P.Y.; Wu, S.N. Characterization of perturbing actions by verteporfin, a benzoporphyrin photosensitizer, on membrane ionic currents. Front. Chem. 2019, 7, 566. [Google Scholar] [CrossRef] [Green Version]













© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, T.-L.; Lu, T.-J.; Wu, S.-N. Inhibitory Effective Perturbations of Cilobradine (DK-AH269), A Blocker of HCN Channels, on the Amplitude and Gating of Both Hyperpolarization-Activated Cation and Delayed-Rectifier Potassium Currents. Int. J. Mol. Sci. 2020, 21, 2416. https://doi.org/10.3390/ijms21072416
Lu T-L, Lu T-J, Wu S-N. Inhibitory Effective Perturbations of Cilobradine (DK-AH269), A Blocker of HCN Channels, on the Amplitude and Gating of Both Hyperpolarization-Activated Cation and Delayed-Rectifier Potassium Currents. International Journal of Molecular Sciences. 2020; 21(7):2416. https://doi.org/10.3390/ijms21072416
Chicago/Turabian StyleLu, Te-Ling, Te-Jung Lu, and Sheng-Nan Wu. 2020. "Inhibitory Effective Perturbations of Cilobradine (DK-AH269), A Blocker of HCN Channels, on the Amplitude and Gating of Both Hyperpolarization-Activated Cation and Delayed-Rectifier Potassium Currents" International Journal of Molecular Sciences 21, no. 7: 2416. https://doi.org/10.3390/ijms21072416