A microRNA Expression Profile as Non-Invasive Biomarker in a Large Arrhythmogenic Cardiomyopathy Cohort

 , , , , and
, , , , and
Abstract
:1. Introduction
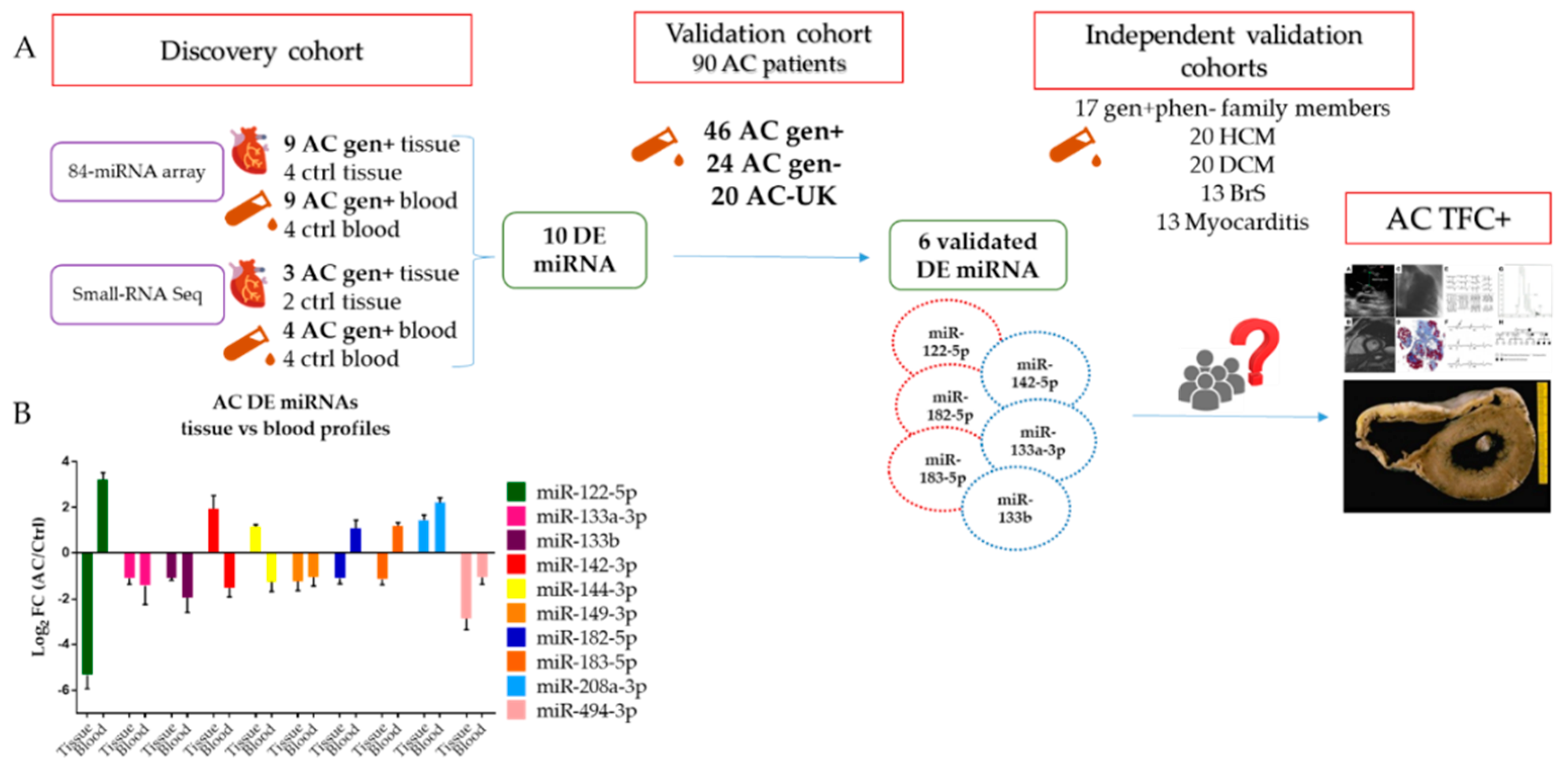
2. Results
2.1. 84 miRNA Cardiac-Related Array Screening on Myocardial Tissue of AC Patients
2.2. Small RNA-Seq on Myocardial Tissue of AC Patients
2.3. 84 miRNA Cardiac-Related Array Screening in the Blood of AC Patients
2.4. Small RNA-Seq on Blood of AC Patients
2.5. Shared miRNA between AC- Tissue and AC-Blood Profiles
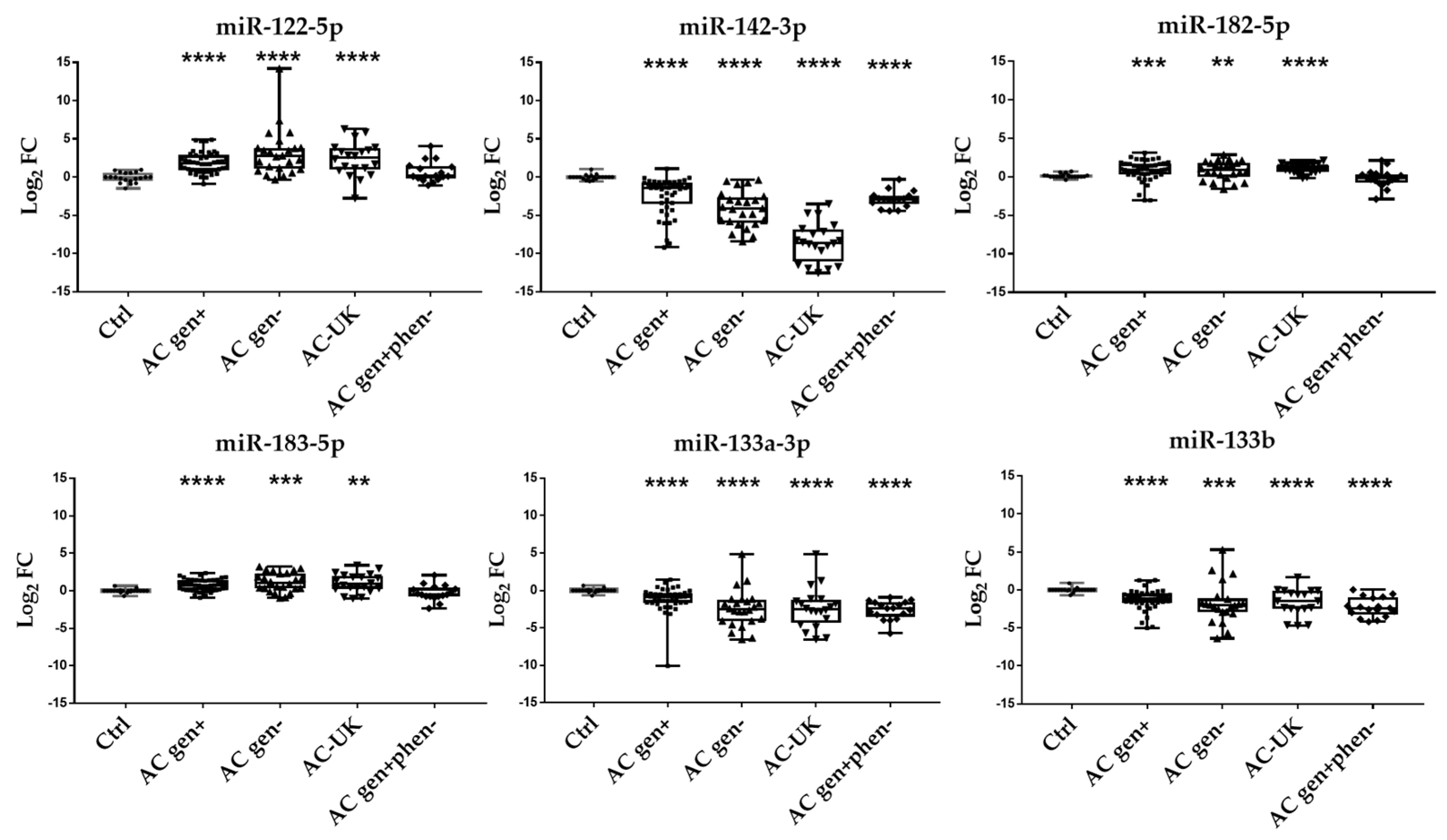
2.6. Validation of DE miRNA in a Larger AC Gen+ Cohort
2.7. DE miRNA in Independent Validation Cohorts
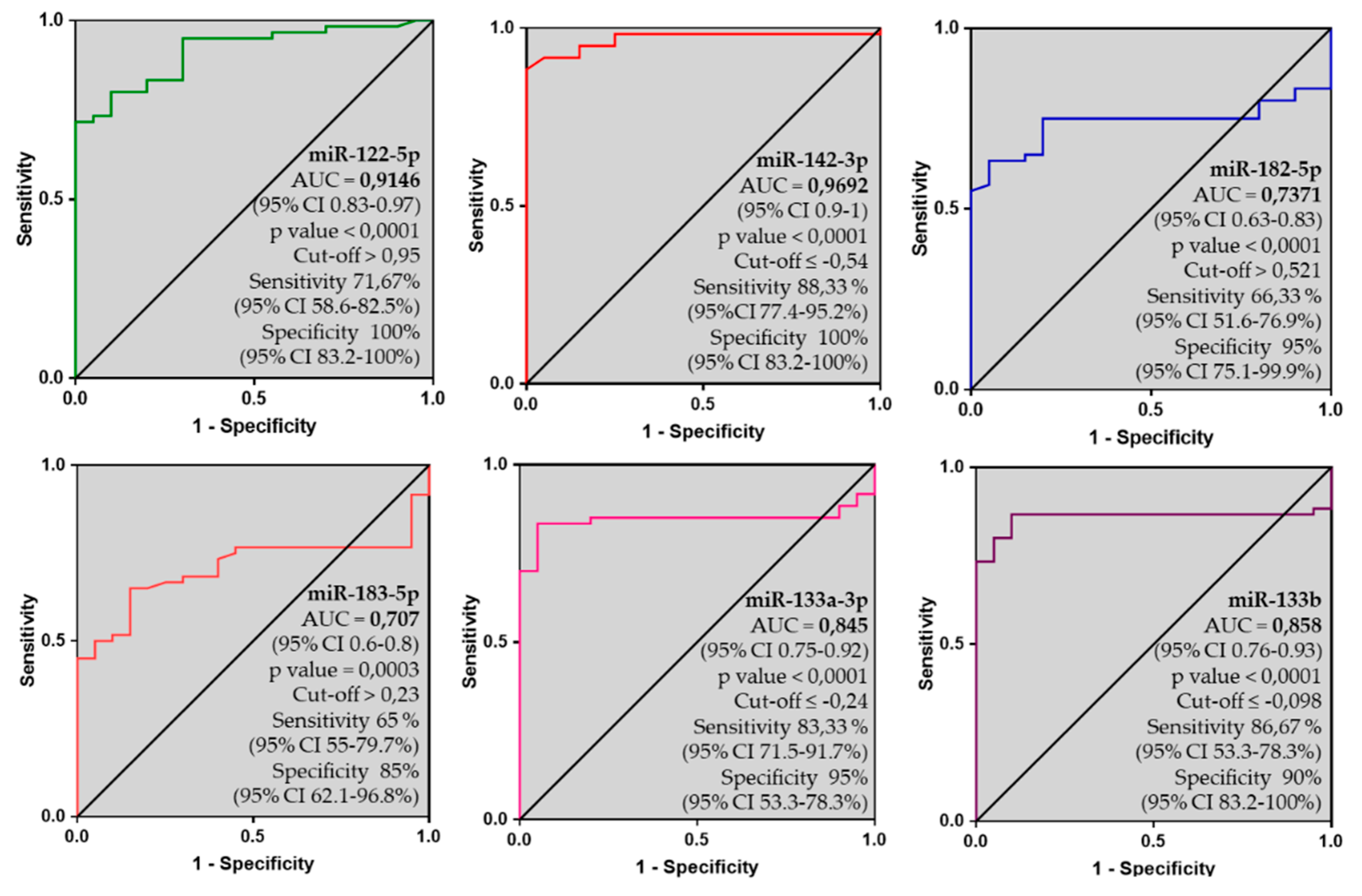
2.8. 6-miRNA Panel as Potential Biomarker of AC
2.9. In Silico Prediction Analysis
3. Discussion
3.1. Previous Studies on miRNA in AC
3.2. Comparison between Tissue and Blood miRNA Profiles
3.3. Discriminatory Power: A New Biomarker for AC?
4. Materials and Methods
4.1. Study Cohort
4.2. miRNA Isolation and Quantification
4.3. miRNA Screening
4.4. Quantitative PCR Validation
4.5. In Silico Target Prediction
4.6. Statistical Analysis
5. Conclusions
6. Limitations
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thiene, G.; Nava, A.; Corrado, D.; Rossi, L.; Pennelli, N. Right ventricular cardiomyopathy and sudden death in young people. N. Engl. J. Med. 1988, 318, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Basso, C.; Thiene, G.; Corrado, D.; Angelini, A.; Nava, A.; Valente, M. Arrhythmogenic right ventricular cardiomyopathy. Dysplasia, dystrophy, or myocarditis? Circulation 1996, 94, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Marcus, F.I.; McKenna, W.J.; Sherrill, D.; Basso, C.; Bauce, B.; Bluemke, D.A.; Calkins, H.; Corrado, D.; Cox, M.G.; Daubert, J.P.; et al. Diagnosis of arrhythmogenic right ventricular cardiomyopathy/dysplasia: Proposed modification of the task force criteria. Circulation 2010, 121, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Basso, C.; Corrado, D.; Marcus, F.I.; Nava, A.; Thiene, G. Arrhythmogenic right ventricular cardiomyopathy. Lancet 2009, 373, 1289–1300. [Google Scholar] [CrossRef]
- Corrado, D.; Basso, C.; Pavei, A.; Michieli, P.; Schiavon, M.; Thiene, G. Trends in sudden cardiovascular death in young competitive athletes after implementation of a preparticipation screening program. JAMA 2006, 296, 1593–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilichou, K.; Thiene, G.; Bauce, B.; Rigato, I.; Lazzarini, E.; Migliore, F.; Perazzolo Marra, M.; Rizzo, S.; Zorzi, A.; Daliento, L.; et al. Arrhythmogenic cardiomyopathy. Orphanet J. Rare Dis. 2016, 11, 33. [Google Scholar] [CrossRef]
- Ambros, V. microRNAs: Tiny regulators with great potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. Elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Garzon, R.; Calin, G.A.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Med. 2009, 60, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Dong, C.; Ji, C. MicroRNA and drug resistance. Cancer Gene Ther. 2010, 17, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Creemers, E.E.; Tijsen, A.J.; Pinto, Y.M. Circulating microRNAs: Novel biomarkers and extracellular communicators in cardiovascular disease? Circ. Res. 2012, 110, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Corsten, M.F.; Dennert, R.; Jochems, S.; Kuznetsova, T.; Devaux, Y.; Hofstra, L.; Wagner, D.R.; Staessen, J.A.; Heymans, S.; Schroen, B. Circulating MicroRNA-208b and MicroRNA-499 reflect myocardial damage in cardiovascular disease. Circ. Cardiovasc. Genet. 2010, 3, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.L.; Lu, X.L.; Yan, C.Y.; Wan, Q.L.; Cheng, G.C.; Li, Y.M. Circulating miR-122-5p as a potential novel biomarker for diagnosis of acute myocardial infarction. Int. J. Clin. Exp. Pathol. 2015, 8, 16014–16019. [Google Scholar] [PubMed]
- van Rooij, E.; Sutherland, L.B.; Liu, N.; Williams, A.H.; McAnally, J.; Gerard, R.D.; Richardson, J.A.; Olson, E.N. A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure. Proc. Natl. Acad. Sci. USA 2006, 103, 18255–18260. [Google Scholar] [CrossRef] [Green Version]
- Kuster, D.W.; Mulders, J.; Ten Cate, F.J.; Michels, M.; Dos Remedios, C.G.; da Costa Martins, P.A.; van der Velden, J.; Oudejans, C.B. MicroRNA transcriptome profiling in cardiac tissue of hypertrophic cardiomyopathy patients with MYBPC3 mutations. J. Mol. Cell. Cardiol. 2013, 65, 59–66. [Google Scholar] [CrossRef]
- Fang, L.; Ellims, A.H.; Moore, X.L.; White, D.A.; Taylor, A.J.; Chin-Dusting, J.; Dart, A.M. Circulating microRNAs as biomarkers for diffuse myocardial fibrosis in patients with hypertrophic cardiomyopathy. J. Transl. Med. 2015, 13, 314. [Google Scholar] [CrossRef]
- Roncarati, R.; Viviani Anselmi, C.; Losi, M.A.; Papa, L.; Cavarretta, E.; Da Costa Martins, P.; Contaldi, C.; Saccani Jotti, G.; Franzone, A.; Galastri, L.; et al. Circulating miR-29a, among other up-regulated microRNAs, is the only biomarker for both hypertrophy and fibrosis in patients with hypertrophic cardiomyopathy. J. Am. Coll. Cardiol. 2014, 63, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Chen, F.; Tong, J.; Li, Y.; Cai, J.; Wang, Y.; Li, P.; Hao, Y.; Tian, W.; Lv, Y.; et al. Circulating microRNAs as novel biomarkers for dilated cardiomyopathy. Cardiol. J. 2017, 24, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Schotterl, S.; Backes, D.; Brunner, E.; Hahn, J.K.; Ionesi, E.; Aidery, P.; Sticht, C.; Labeit, S.; Kandolf, R.; et al. Inhibition of miR-208b improves cardiac function in titin-based dilated cardiomyopathy. Int. J. Cardiol. 2017, 230, 634–641. [Google Scholar] [CrossRef]
- Rubis, P.; Toton-Zuranska, J.; Wisniowska-Smialek, S.; Holcman, K.; Kolton-Wroz, M.; Wolkow, P.; Wypasek, E.; Natorska, J.; Rudnicka-Sosin, L.; Pawlak, A.; et al. Relations between circulating microRNAs (miR-21, miR-26, miR-29, miR-30 and miR-133a), extracellular matrix fibrosis and serum markers of fibrosis in dilated cardiomyopathy. Int. J. Cardiol. 2017, 231, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, S.; Dong, T.; Yang, J.; Xie, Y.; Wu, Y.; Kang, K.; Hu, S.; Gou, D.; Wei, Y. Profiling of differentially expressed microRNAs in arrhythmogenic right ventricular cardiomyopathy. Sci. Rep. 2016, 6, 28101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommariva, E.; D’Alessandra, Y.; Farina, F.M.; Casella, M.; Cattaneo, F.; Catto, V.; Chiesa, M.; Stadiotti, I.; Brambilla, S.; Dello Russo, A.; et al. MiR-320a as a Potential Novel Circulating Biomarker of Arrhythmogenic CardioMyopathy. Sci. Rep. 2017, 7, 4802. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Hsiao, Y.W.; Chang, S.L.; Lin, Y.J.; Lo, L.W.; Chung, F.P.; Chiang, S.J.; Hu, Y.F.; Tuan, T.C.; Chao, T.F.; et al. Circulating microRNAs in arrhythmogenic right ventricular cardiomyopathy with ventricular arrhythmia. Europace 2018, 20, f37–f45. [Google Scholar] [CrossRef]
- Rainer, J.; Meraviglia, V.; Blankenburg, H.; Piubelli, C.; Pramstaller, P.P.; Paolin, A.; Cogliati, E.; Pompilio, G.; Sommariva, E.; Domingues, F.S.; et al. The arrhythmogenic cardiomyopathy-specific coding and non-coding transcriptome in human cardiac stromal cells. BMC Genom. 2018, 19, 491. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- Gurha, P.; Chen, X.; Lombardi, R.; Willerson, J.T.; Marian, A.J. Knockdown of Plakophilin 2 Downregulates miR-184 Through CpG Hypermethylation and suppression of the E2F1 pathway and leads to enhanced adipogenesis In Vitro. Circ. Res. 2016, 119, 731–750. [Google Scholar] [CrossRef] [Green Version]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Chen, S.N.; Gurha, P.; Lombardi, R.; Ruggiero, A.; Willerson, J.T.; Marian, A.J. The hippo pathway is activated and is a causal mechanism for adipogenesis in arrhythmogenic cardiomyopathy. Circ. Res. 2014, 114, 454–468. [Google Scholar] [CrossRef]
- Calore, M.; Lorenzon, A.; Vitiello, L.; Poloni, G.; Khan, M.A.F.; Beffagna, G.; Dazzo, E.; Sacchetto, C.; Polishchuk, R.; Sabatelli, P.; et al. A novel murine model for arrhythmogenic cardiomyopathy points to a pathogenic role of Wnt signaling and miRNA dysregulation. Cardiovasc. Res. 2018, 115, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef] [PubMed]
- Beffagna, G.; Occhi, G.; Nava, A.; Vitiello, L.; Ditadi, A.; Basso, C.; Bauce, B.; Carraro, G.; Thiene, G.; Towbin, J.A.; et al. Regulatory mutations in transforming growth factor-beta3 gene cause arrhythmogenic right ventricular cardiomyopathy type 1. Cardiovasc. Res. 2005, 65, 366–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.K.; Zhu, J.Q.; Zhang, J.T.; Li, Q.; Li, Y.; He, J.; Qin, Y.W.; Jing, Q. Circulating microRNA: A novel potential biomarker for early diagnosis of acute myocardial infarction in humans. Eur. Heart J. 2010, 31, 659–666. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, S.; Marzolf, B.; Troisch, P.; Brightman, A.; Hu, Z.; Hood, L.E.; Galas, D.J. Circulating microRNAs, potential biomarkers for drug-induced liver injury. Proc. Natl. Acad. Sci. USA 2009, 106, 4402–4407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, G.; Kirschner, M.B.; van Zandwijk, N. Circulating microRNAs: Association with disease and potential use as biomarkers. Crit. Rev. Oncol. Hematol. 2011, 80, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Oikawa, K.; Takanashi, M.; Kudo, M.; Ohyashiki, J.; Ohyashiki, K.; Kuroda, M. Down-regulation of miR-92 in human plasma is a novel marker for acute leukemia patients. PLoS ONE 2009, 4, e5532. [Google Scholar] [CrossRef]
- Elliott, P.M.; Anastasakis, A.; Borger, M.A.; Borggrefe, M.; Cecchi, F.; Charron, P.; Hagege, A.A.; Lafont, A.; Limongelli, G.; et al.; Authors/Task Force members 2014 esc guidelines on diagnosis and management of hypertrophic cardiomyopathy: The task force for the diagnosis and management of hypertrophic cardiomyopathy of the european society of cardiology (esc). Eur. Heart J. 2014, 35, 2733–2779. [Google Scholar]
- Pinto, Y.M.; Elliott, P.M.; Arbustini, E.; Adler, Y.; Anastasakis, A.; Bohm, M.; Duboc, D.; Gimeno, J.; de Groote, P.; Imazio, M.; et al. Proposal for a revised definition of dilated cardiomyopathy, hypokinetic non-dilated cardiomyopathy, and its implications for clinical practice: A position statement of the esc working group on myocardial and pericardial diseases. Eur. Heart J. 2016, 37, 1850–1858. [Google Scholar] [CrossRef] [Green Version]
- Priori, S.G.; Blomstrom-Lundqvist, C.; Mazzanti, A.; Blom, N.; Borggrefe, M.; Camm, J.; Elliott, P.M.; Fitzsimons, D.; Hatala, R.; Hindricks, G.; et al. 2015 esc guidelines for the management of patients with ventricular arrhythmias and the prevention of sudden cardiac death: The task force for the management of patients with ventricular arrhythmias and the prevention of sudden cardiac death of the european society of cardiology (esc). Endorsed by: Association for european paediatric and congenital cardiology (aepc). Eur. Heart J. 2015, 36, 2793–2867. [Google Scholar]
- Caforio, A.L.; Pankuweit, S.; Arbustini, E.; Basso, C.; Gimeno-Blanes, J.; Felix, S.B.; Fu, M.; Helio, T.; Heymans, S.; Jahns, R.; et al. Current state of knowledge on aetiology, diagnosis, management, and therapy of myocarditis: A position statement of the european society of cardiology working group on myocardial and pericardial diseases. Eur. Heart J. 2013, 34, 2636–2648, 2648a–2648d. [Google Scholar] [CrossRef]
- Hoo, Z.H.; Candlish, J.; Teare, D. What is an roc curve? Emerg. Med. J. EMJ 2017, 34, 357–359. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Specimen | AC TFC Criteria | Gene | cDNA | Protein | dbSNP ID | MAF gnomAD | ACMG Variant Classification |
|---|---|---|---|---|---|---|---|---|
| AC 1 | RV myocardial tissue | 2M | PKP2 | c.1643delG | p.Gly548ValfsX15 | rs794729137 | 0.00002 | Pathogenic |
| AC 2 | RV myocardial tissue | 3M | PKP2 | c.2013delC | p.Lys672ArgfsX12 | rs764817683 | 0.0000081 | Pathogenic |
| AC 3 | RV myocardial tissue | 1M, 2m | PKP2 | chr12:32,782,127-133,275,309 heterozygous deletion | Pathogenic | |||
| AC 4 | RV myocardial tissue | 2M | DSP | c.2956C > T | p.Gln986X | NA | NA | Pathogenic |
| AC 5 | RV myocardial tissue | 4M | DSP | c.5644G > T | p.Glu1882X | NA | NA | Pathogenic |
| AC 6 | RV myocardial tissue | 3M, 1m | DSP | c.5511dupT | p.Arg1838SerfsX19 | NA | NA | Pathogenic |
| AC 7 | RV myocardial tissue | 2M | DSG2 | c.797A > G | p.Asn266Ser | rs121913011 | 0.0000041 | Pathogenic |
| AC 8 | RV myocardial tissue | 2M | DSG2 | c.1672C > T | p.Gln558X | NA | NA | Pathogenic |
| AC 9 | RV myocardial tissue | 1M, 2m | DSG2 | c.2033G > C | p.Gly678Ala | rs372494397 | 0.00004 | VUS * |
| AC 1 | Whole blood | 2M | PKP2 | c.1643delG | p.Gly548ValfsX15 | rs794729137 | 0.00002 | Pathogenic |
| AC 2 | Whole blood | 3M | PKP2 | c.2013delC | p.Lys672ArgfsX12 | rs764817683 | 0.0000081 | Pathogenic |
| AC 10 | Whole blood | 3M, 1m | PKP2 | c.631C > T | p.Gln211X | NA | NA | Pathogenic |
| AC 11 | Whole blood | 3M | PKP2 | c.2013delC | p.Lys672ArgfsX12 | rs764817683 | 0.0000081 | Pathogenic |
| AC 12 | Whole blood | 2M, 2m | PKP2 | c.964_965delinsC | p.Gly322ProfsX30 | NA | NA | Likely pathogenic |
| AC 13 | Whole blood | 3M | DSG2 | c.797A > G | p.Asn266Ser | rs121913011 | 0.0000041 | Pathogenic |
| AC 14 | Whole blood | 3M | DSP | c.3889C > T | p.Gln1297X | NA | 0.0000041 | Pathogenic |
| AC 15 | Whole blood | 1M, 2m | DSG2/ DSC2 | chr18:31,065,974-31,549,007 heterozygous deletion | Pathogenic | |||
| AC 16 | Whole blood | 2M, 1m | DSP | c.337C > T | p.Gln113X | NA | NA | Pathogenic |
| DSG2 | c.797A > G | p.Asn266Ser | rs121913011 | 0.0000041 | Pathogenic | |||
| Validation and Independent Validation Cohorts | ||||||
|---|---|---|---|---|---|---|
| Log2FC (mean ±SEM) p value (Mann-Whitney test) p value* (Kruskal-Wallis adjusted FDR) | ||||||
| miR-122-5p | miR-142-3p | miR-182-5p | miR-183-5p | miR-133a-3p | miR-133b | |
| AC gen+ | 1.87 ± 0.20 | −2.4 ± 0.35 | 0.78 ± 0.19 | 0.71 ± 0.12 | −1.15 ± 0.27 | −1.29 ± 0.22 |
| p < 0.0001 | p < 0.0001 | p = 0.0002 | p < 0.0001 | p < 0.0001 | p < 0.0001 | |
| p* < 0.0001 | p* < 0.0001 | p* = 0.0045 | p* = 0.0161 | p* = 0.0081 | p* = 0.0007 | |
| AC gen− | 3.20 ± 0.64 | −4.23 ± 0.47 | 0.82 ± 0.24 | 1.16 ± 0.26 | −2.44 ± 0.51 | −1.75 ± 0.57 |
| p < 0.0001 | p < 0.0001 | p = 0.0096 | p = 0.0009 | p < 0.0001 | p = 0.0002 | |
| p* < 0.0001 | p* < 0.0001 | p* = 0.0170 | p* = 0.0029 | p* < 0.0001 | p* < 0.0001 | |
| AC-UK | 2.34 ± 0.49 | −8.54 ± 0.57 | 1.05 ± 0.13 | 1.04 ± 0.28 | −2.91 ± 0.59 | −1.63 ± 0.42 |
| p < 0.0001 | p < 0.0001 | p < 0.0001 | p = 0.001 | p < 0.0001 | p < 0.0001 | |
| p* < 0.0001 | p* < 0.0001 | p* = 0.0021 | p* = 0.0074 | p* < 0.0001 | p* = 0.0007 | |
| AC gen + phen− | 0.67 ± 0.32 | −2.88 ± 0.25 | −0.17 ± 0.28 | −0.27 ± 0.25 | −2.62 ± 0.30 | −2.27 ± 0.30 |
| p = 0.063 | p < 0.0001 | p = 0.269 | p = 0.060 | p < 0.0001 | p < 0.0001 | |
| p* = 0.1458 | p* = 0.0001 | p* = 0.7852 | p* = 0.4724 | p* < 0.0001 | p* < 0.0001 | |
| HCM | 1.98 ± 0.56 | −0.87 ± 0.07 | 0.72 ± 0.49 | 0.72 ± 0.43 | −0.58 ± 0.38 | −0.62 ± 0.34 |
| p = 0.002 | p = 0.073 | p = 0.049 | p = 0.026 | p = 0.217 | p = 0.465 | |
| p* = 0.0001 | p* = 0.2599 | p* = 0.0221 | p* = 0.0219 | p* = 0.1753 | p* = 0.1388 | |
| DCM | 1.45 ± 0.31 | −5.49 ± 0.66 | −0.83 ± 0.32 | −0.36 ± 0.30 | −2.90 ± 0.37 | −3.01 ± 0.32 |
| p < 0.0001 | p < 0.0001 | p = 0.085 | p = 0.792 | p < 0.0001 | p < 0.0001 | |
| p* = 0.0021 | p* < 0.0001 | p* = 0.2373 | p* = 0.8852 | p* < 0.0001 | p* < 0.0001 | |
| BrS | 2.84 ± 1.306 | −6.58 ± 0.59 | −1.27 ± 0.50 | −0.31 ± 0.40 | −4.38 ± 0.45 | −3.24 ± 0.51 |
| p = 0.057 | p < 0.0001 | p = 0.0087 | p = 0.2829 | p < 0.0001 | p < 0.0001 | |
| p* = 0.0037 | p* < 0.0001 | p* = 0.1689 | p* = 0.6971 | p* < 0.0001 | p* < 0.0001 | |
| Myocarditis | 4.57 ± 1.47 | −7.11 ± 0.48 | −1.38 ± 0.42 | −0.34 ± 0.39 | −1.61 ± 1.17 | −1.39 ± 1.21 |
| p = 0.0001 | p < 0.0001 | p = 0.0003 | p = 0.889 | p = 0.070 | p = 0.125 | |
| p* < 0.0001 | p* < 0.0001 | p* = 0.0748 | p* = 0.7245 | p* = 0.0013 | p* = 0.0042 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bueno Marinas, M.; Celeghin, R.; Cason, M.; Bariani, R.; Frigo, A.C.; Jager, J.; Syrris, P.; Elliott, P.M.; Bauce, B.; Thiene, G.; et al. A microRNA Expression Profile as Non-Invasive Biomarker in a Large Arrhythmogenic Cardiomyopathy Cohort. Int. J. Mol. Sci. 2020, 21, 1536. https://doi.org/10.3390/ijms21041536
Bueno Marinas M, Celeghin R, Cason M, Bariani R, Frigo AC, Jager J, Syrris P, Elliott PM, Bauce B, Thiene G, et al. A microRNA Expression Profile as Non-Invasive Biomarker in a Large Arrhythmogenic Cardiomyopathy Cohort. International Journal of Molecular Sciences. 2020; 21(4):1536. https://doi.org/10.3390/ijms21041536
Chicago/Turabian StyleBueno Marinas, Maria, Rudy Celeghin, Marco Cason, Riccardo Bariani, Anna Chiara Frigo, Joanna Jager, Petros Syrris, Perry M. Elliott, Barbara Bauce, Gaetano Thiene, and et al. 2020. "A microRNA Expression Profile as Non-Invasive Biomarker in a Large Arrhythmogenic Cardiomyopathy Cohort" International Journal of Molecular Sciences 21, no. 4: 1536. https://doi.org/10.3390/ijms21041536