Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review


1
Department of Food Science and Engineering, School of Chemistry and Chemical Engineering, Harbin Institute of Technology, Harbin 150090, China
2
Institute of Microbiology, Heilongjiang Academy of Sciences, Harbin 150010, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(3), 995; https://doi.org/10.3390/ijms21030995
Submission received: 21 December 2019
/
Revised: 21 January 2020
/
Accepted: 27 January 2020
/
Published: 3 February 2020
(This article belongs to the Section Molecular Microbiology)
Abstract
:Gamma-aminobutyric acid (GABA) is widely distributed in nature and considered a potent bioactive compound with numerous and important physiological functions, such as anti-hypertensive and antidepressant activities. There is an ever-growing demand for GABA production in recent years. Lactic acid bacteria (LAB) are one of the most important GABA producers because of their food-grade nature and potential of producing GABA-rich functional foods directly. In this paper, the GABA-producing LAB species, the biosynthesis pathway of GABA by LAB, and the research progress of glutamate decarboxylase (GAD), the key enzyme of GABA biosynthesis, were reviewed. Furthermore, GABA production enhancement strategies are reviewed, from optimization of culture conditions and genetic engineering to physiology-oriented engineering approaches and co-culture methods. The advances in both the molecular mechanisms of GABA biosynthesis and the technologies of synthetic biology and genetic engineering will promote GABA production of LAB to meet people’s demand for GABA. The aim of the review is to provide an insight of microbial engineering for improved production of GABA by LAB in the future.
1. Introduction
Gamma-aminobutyric acid (GABA) is a four-carbon free amino acid that is produced from l-glutamic acid by glutamate decarboxylase (GAD) and is widely present in microorganisms, plants, and animals [1]. It is well known that GABA is considered a bioactive component with multiple physiological functions [2]. GABA acts as a major inhibitory neurotransmitter that sends chemical messages in the mammalian central nervous system [3]. Moreover, GABA also plays an important role in behavior, cognition, and the body’s response to stress. GABA as a supplement is involved in improving sleeplessness and depression [4,5], enhancing immunity [5], relieving anxiety and menopausal syndrome [6], regulating blood pressure [7], fighting obesity [8], and improving visual cortical function [9].
At present, food containing GABA cannot meet people’s needs because of the low content of GABA. Microorganisms are an important source of GABA. Up to now, it has been confirmed that many types of microorganisms can synthesize GABA, including yeast, fungi, and bacteria [10,11] (Supplementary Table S1). Microorganisms grow faster than plants, do not need much space for cultivation, and are eco-friendly for consumers. Furthermore, it is easy to control production of microorganisms.
Lactic acid bacteria (LAB) are widely used in the foods industry, particularly in manufacturing fermented foods for many centuries [12]. Because of the generally recognized as safe (GRAS) status of LAB and their high application potential in the fermentation industry, GABA-producing LAB have received extensive attention in recent years. A large number of GABA-producing LAB were isolated from fermented food and used in the manufacturing of naturally health-oriented foods enriched with GABA. In view of the food-grade nature of LAB and their potential as a functional food, this paper will focus on the GABA-producing LAB.
In the present review, we will describe GABA-producing LAB species and the biosynthetic pathway of GABA in LAB, with particular emphasis on the diversity of the GAD system of LAB. Furthermore, GABA production enhancement strategies are discussed, from optimization of culture conditions and genetic engineering to physiology-oriented engineering approaches and co-culture methods.
2. GABA-Producing LAB Species
LAB are important GABA-producing microorganisms because of their various production characteristics and probiotic effects. A number of LAB strains with GABA-producing capability have been isolated from traditional fermented foods such as cheese, kimchi, paocai, yoghurt, and fermented soy beans and so on (Supplementary Table S2).
The genus Lactobacillus has abundant GABA-producing species, including Lactobacillus brevis [13,14,15,16,17,18,19], Lactobacillus buchneri [20,21], Lactobacillus delbrueckii subsp. bulgaricus [17,22], Lactobacillus fermentum [23,24], Lactobacillus helveticus [25], Lactobacillus paracasei [17,26], Lactobacillus plantarum [17,26,27], and so on. Furthermore, some Streptococcus thermophilus and Lactococcus lactis strains display GABA production abilities, which are the best candidates for production of GABA-rich milk products [17,19,26,28]. In recent years, it has been found that some species from the genera Enterococcus, Leuconostoc, Pediococcus, Propionibacterium, and Weissella are capable of producing GABA (Supplementary Table S2).
l-glutamate-rich fermented foods are important isolation sources of GABA-producing LAB. Cheese contains a large number of caseins that could produce plenty of l-glutamate. Lb. buchneri, Lb. brevis, Lb. paracasei, Lb. plantarum, Lb. delbrueckii subsp. bulgaricus, and Lc. lactis strains isolated from traditional or commercial cheeses have been shown to produce GABA [17,19,20,26]. Generally, an acid environment is beneficial for the growth of GABA-producing LAB, such as in Korean kimchi and Chinese paocai.
The GABA production capacity of different species is greatly varied. A large number of studies have shown that Lb. brevis can produce a high yield of GABA compared with other LAB species (Supplementary Table S2). Up to now, the highest yield of GABA by Lb. brevis is 205 g/L [18]. At the same time, various strains of one species have obvious differences in GABA productivity (Supplementary Table S2). Different strains of Lb. brevis produce GABA in the range from 15.0 mg/L to 205 g/L [17,18]. Similarly, different Lb. plantarum strains isolated from various cheeses have diverse GABA productivity [17,26,27].
3. Biosynthesis of GABA in LAB
3.1. Biosynthetic Pathway of GABA in LAB
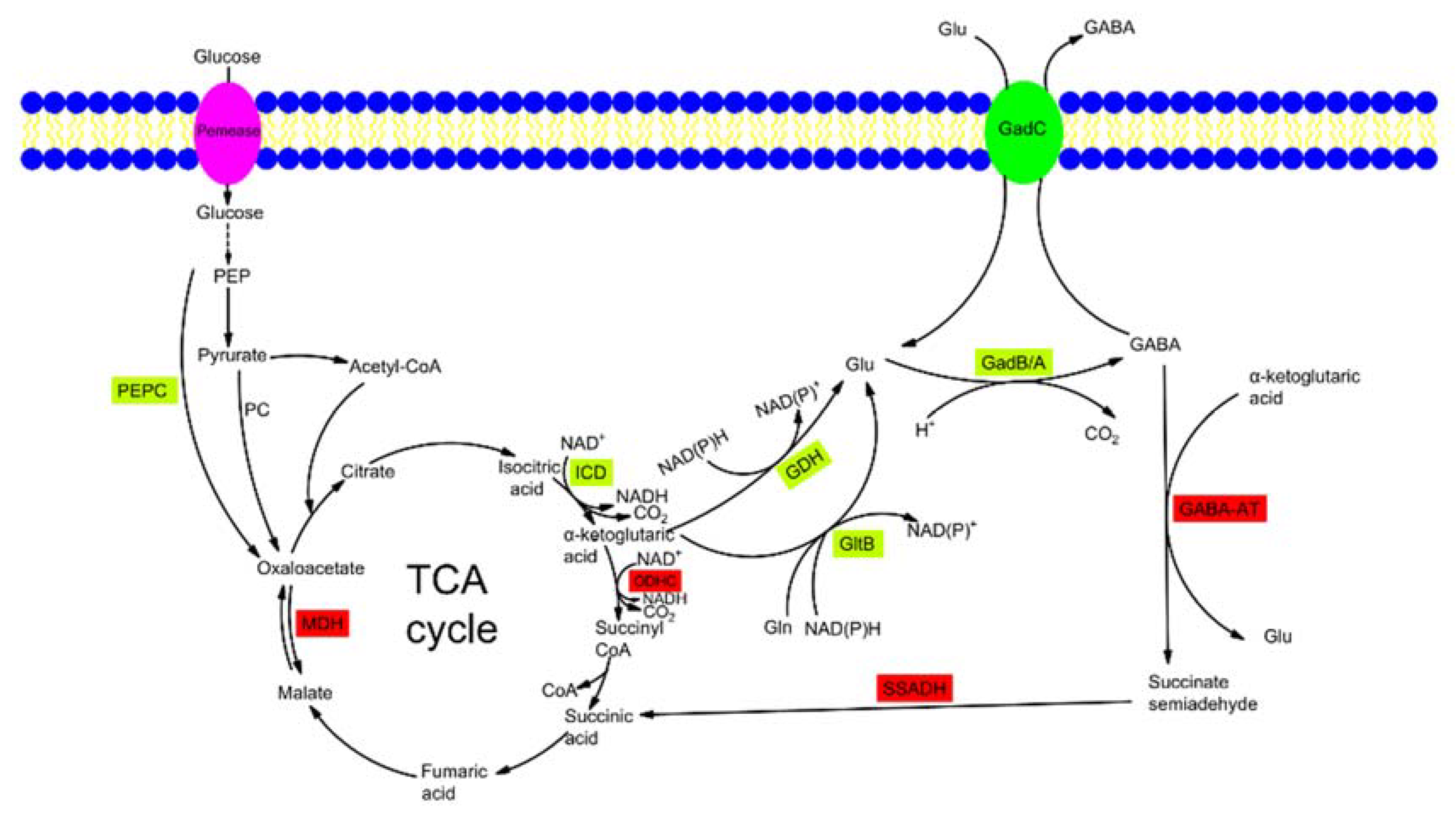
The biosynthesis of GABA by microorganisms is performed by the glutamic acid decarboxylase (GAD, EC 4.1.1.15) system, which is composed of the GAD enzyme (encoded by gadA or gadB) and glutamate/GABA antiporter GadC [29,30,31,32,33]. The biosynthetic pathway of GABA by microbes is shown in Figure 1.
l-glutamate is transported into a cell through GadC. The decarboxylation of l-glutamate is catalyzed by GAD with cofactor pyridoxal-5′-phosphate (PLP), and leads to the formation of GABA and release of CO2 as byproduct (Figure 1). Finally, the decarboxylated product GABA is exported to the extracellular matrix by GadC [29]. As the precursor of l-glutamate, a-ketoglutarate is synthesized from glucose via the glycolysis pathway and part of the tricarboxylic acid (TCA) cycle, and then it is converted into l-glutamate by l-glutamate dehydrogenase (GDH, EC 1.4.1.4). The 2-oxoglutarate dehydrogenase complex (ODHC) is the key enzyme of the TCA cycle and acts at the branching point of metabolic flux between l-glutamate synthesis and energy supply. Moreover, it competes with GDH for the substrate a-ketoglutarate.
In some bacteria, such as Escherichia coli and Listeria monocytogenes, the molecular mechanisms of GABA degradation have been clarified [34,35]. GABA is degraded to succinic semialdehyde (SSA) by the major GABA-degradative enzyme GABA aminotransferase (GABA-AT, EC 2.6.1.19), and then it later converts to succinic acid at catalysis of succinate semialdehyde dehydrogenase (SSADH, EC 1.2.1.16) for entry into the TCA cycle.
The GABA-AT encoding gene gadT has been identified in the some species of LAB, such as Lb. fermentum, Lactobacillus frumenti, Lactobacillus gastricus, Lactobacillus gorilla, Lactobacillus mucosae, Lactobacillus oris, Lb. plantarum [36], Lactobacillus pontis, Lactobacillus reuteri [37], Lactobacillus similis, Lactobacillus vaginalis, Leuconostoc citreum [38], I subsp. gasicomitatum [39], Leuconostoc kimchii [40], Leuconostoc mesenteroides subsp. mesenteroides [41], Oenococcus oeni [41], and so on. Recently, it has been found that GABA production of Lb. plantarum is obviously increased in the fermentation with MSG [27]. At the same time, the expression of gene gadT was down-regulated by a factor of 0.26 times, indicating that the activity of GABA-AT was inhibited. The results show that GABA-AT activity has a direct relationship with the GABA production.
3.2. GAD System of LAB
The GAD system is present in various LAB species, including genera Enterococcus, Lactobacillus, Lactococcus, Pediococcus, Propionibacterium, and Streptococcus (Figure 2). Its genetic organization shows a high variability. Although some LAB species have GABA-producing ability, such as Lactobacillus casei, Lb. paracasei, Leuconostoc mesenteroides, and Weissella hellenica, there is little information about the related genes of GABA production in the genomes of these species [17,42,43,44].
Most GAD systems of LAB species locate in the chromosome; however, that of Lactobacillus sakei WiKim0074 is from its plasmid. A glutamate-tRNA ligase gene gltX is found upstream of the gadB-gadC operon of some species. Furthermore, glutamate synthases encoding genes gltB and gltC are located near the gadB-gadC gene in Lc. lactis [45,46]. It was speculated that these glutamate metabolism related genes could facilitate GABA production.
For different strains of the same species, genetic organization of the GAD system also presents variability. For example, the gadB-gadC genes are flanked by transposases elements in S. thermophilus. In S. thermophilus strains ACA-DC 2, B59671, and TH1435, S-ribosylhomocysteine lyase and ribonuclease Y locate, respectively, upstream and downstream of the gadB-gadC operon [47,48,49]. However, for strains ND03 and APC151, DUF1310 family protein and ABC transporter ATP-binding protein locate downstream of gadB-gadC [50,51]. Furthermore, the genetic organization of gadB-gadC in strain KLDS 3.1003 is very special, with an additional ca. 7 kb polynucleotide stretch including transposase, NUDIX domain-containing protein, and XRE family transcriptional regulator elements between S-ribosylhomocysteine lyase and GadB [52].
GAD is the key enzyme for the bioconversion of GABA and localizes in the cytoplasm. The strains with the GAD gene are able to synthesize GABA. Generally, most GABA-producing LAB species have a GAD-encoding gene (Figure 2 and Figure 3). Interestingly, Enterococcus avium 352 has three GADs, and most Lb. brevis strains contain two distinct GAD encoding genes [1,30,53,54].
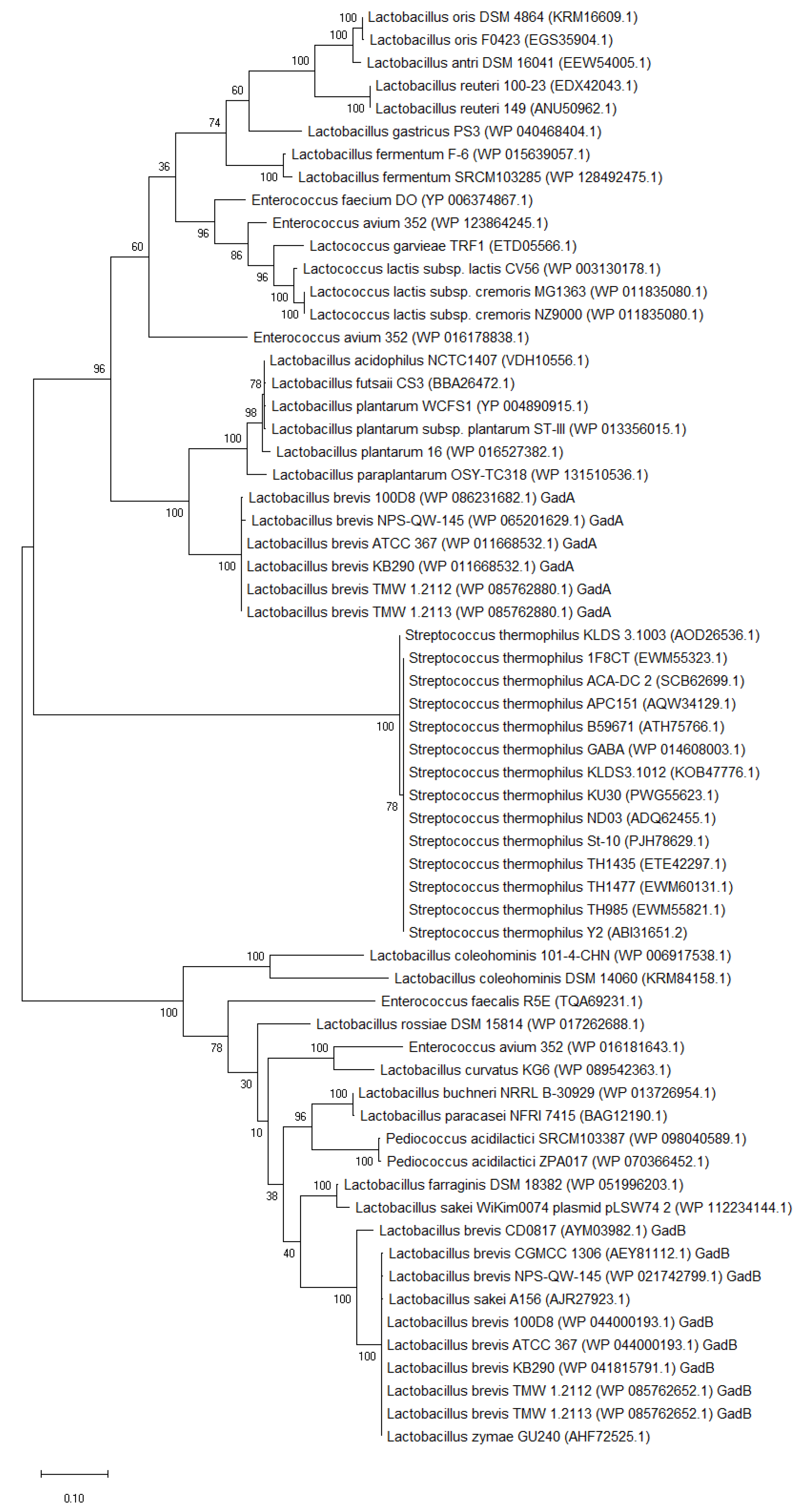
The phylogenetic tree analysis shows that the amino acid sequences of GADs from different LAB species are highly conserved in the same species (Figure 3). These GADs belong to the pyridoxal-5′-phosphate (PLP)-dependent decarboxylase super-family, and they possess a highly conserved lysine residue (Lys278 or Lys279), which is essential for the binding of the PLP, as well as the active site residues (Thr214 and Asp245; or Thr215 and Asp246) that promote decarboxylation.
It is worth noting that the names of two GAD-encoding genes in Lb. brevis are different in different studies. In this review, the classification of GADs from Lb. brevis was based on previous studies [30]. One gad gene (gadB) is located in an operon with gadC. However, the other gad gene (gadA) has a far genetic distance with those from other LAB and is located separately from the other gad genes [1,16,30,54]. GadA and GadB have only about 48% identity in amino acid sequences; however, the PLP-binding domain where the active site residues of both GADs are highly conserved. GAD activity was mainly contributed by GadB in Lb. brevis in the previous study [30].
Recently, it was found that a high GABA-producing Lb. brevis CD0817 contained only a gadB-gadC operon and was absent of gadA gene [55]. What is more, there are obvious differences between GadB of CD0817 and those in the other GABA-producing Lb. brevis strains, with only 91% of amino acid sequence identity. The results further demonstrated that GadB contributed to the main GAD activity [55]. gadA does not have a close relationship with the accumulation of GABA. It was speculated that this distinctive GAD system contributed to the high GABA productivity of Lb. brevis CD0817.
GADs have been isolated from several LAB species, and their biochemical properties have been characterized (Table 1). Although the characteristics of GADs from different LAB species and strains are very different, most LAB GADs exhibit optimal activity at pH 4.0–5.0 (Table 1). Moreover, GADs significantly lose activity at near-neutral pH (pH 7.0).
Some studies suggested that the C-terminal region of GAD is involved in the pH dependence of catalysis [66,71]. The C-terminally truncated mutant of Lb. plantarum GAD showed a pronounced catalytic activity in a broad pH range of 4.0–8.0 [66]. Compared with the wild type, a GAD mutant with a 14C-terminal residue truncation from Lb. brevis CGMCC 1306 exhibited an extended enzymatic activity toward near-neutral pH [71]. Therefore, the C-terminal GAD could as an effective target region for improving the pH dependence of GAD activity. The optimal temperatures of GADs are varied in different species of LAB, ranging from 30 to 60 °C (Table 1). S. thermophillus Y2 presented an increased GAD activity at 34–37 °C, but the GAD activity decreased when temperature continued to increase from 37 to 46 °C [28].
The crystal structure of GAD from Lb. brevis CGMCC 1306 has been successfully solved [72]. It is different from the GAD of E. coli, which contains six repetitive subunits. It consists of two repetitive subunits composed of a conserved lysine residue, Lys279, that binds to coenzyme PLP. Furthermore, some key residues of GAD (Ser126, Ser127, Cys168, Ile211, Ser276, His278, and Ser321) have been determined, which play essential roles in anchoring the PLP cofactor inside the active site and supporting GAD catalytic reactivity. The flexible loop (Tyr308-Glu312), which is positioned near the substrate-binding site, is involved in the catalytic reaction, and the conserved residue Tyr308 plays a critical role in decarboxylation of l-Glu.
GAD structural information will help us to understand its catalytic mechanism and the structure-function relationship. Moreover, it offers a new direction for improving GAD activity and stability by means of rational or semirational methods. Compared with the wild-type GAD of Lb. brevis CGMCC 1306, the mutant T215A, which the mutagenesis site is around the putative substrate pocket, showed a 1.6-fold improvement in catalytic efficiency [72].
The gadC gene has been identified to encode a GABA antiporter, which is involved in the GABA export and glutamate import [73]. Generally, LAB species contain a GABA antiporter, but Lb. fermentum possesses a GAD that is not accompanied by a GABA antiporter; Lb. reuteri has two GABA antiporters (Figure 2).
The GadR is a positive transcriptional regulator that can activate the gadB gene and control the GABA conversion in Lc. lactis and Lb. brevis [74,75]. In Lc. lactis, GadR expression is chloride and glutamate independent [74]. The study showed that GadR has a close relationship with the GABA production, and hyper expression of GadR could increase the GABA productivity [75]. At the same time, GadR could improve the acid resistance of the strain [75]. Some potential transcriptional regulators of GABA production were found in several LAB species such as Lb. buchneri, Lactobacillus coleohominis, Lactobacillus curvatus, Lactobacillus farraginis, Lb. sakei, Lactococcus lactis subsp. cremoris, Lactococcus lactis subsp. lactis, and Pediococcus acidilactici (Figure 2). Generally, gadR locates immediately upstream of the operon gadB-gadC. It is different from the high identities of GadB of different LAB species; these GadRs present extremely low identities in amino acids sequences (Figure 2).
In a word, the GAD system of LAB has strain specificity. A great number of researchers indicated that strains with identical GAD systems showed different GABA productivities. Therefore, GABA production is a complicated process.
4. Improvement of the Production of GABA of LAB
A number of LAB species have shown strain-specific capability in the synthesis of GABA. A number of studies have demonstrated that culture conditions play an important role in GABA production. Therefore, developing higher-yield strains (e.g., by strain selection and breeding, mutagenesis, or genetic manipulation) and optimizing fermentation conditions are two approaches for improving GABA yield. At the same time, fermented food with GABA-producing LAB is a good choice because GABA extraction is eliminated.
4.1. Optimization of Culture Conditions
The GABA production capacity of strains is significantly affected by culture conditions. A large number of studies have been performed to improve GABA yield via optimizing fermentation conditions, such as optimizing the initial pH value of culture medium, fermentation temperature, fermentation time, l-glutamic acid concentration, PLP, media additives, carbon source, nitrogen source, and so on [14,76,77,78].
The pH value is a key factor for GABA biosynthesis by LAB; it not only influences the growth of bacteria, but also affects the GAD activity [15,21,28,79,80]. Some studies have shown that the initial pH of fermentation medium affected GABA synthesis [15,79]. Lb. brevis NCL912 could produce the maximum GABA level at initial pH 5.0 [15]. Compared with initial pH 4.0 and pH 6.0, GABA production of Lb. paracasei NFRI 7415 was significantly enhanced, reaching 210 mM at initial pH 5.0 [80]. The optimal initial pH is also 5.0 for GABA synthesis of Lb. buchneri [21]. However, Lb. brevis GABA100 achieved the highest GABA yield at initial pH 3.5 during the fermentation of black raspberry juice [79]. Therefore, the optimal conditions of fermenting microorganisms vary according to the different properties of GADs, with optimal pH ranging from pH 3.5–5.0.
Maintaining low pH (about 5) is necessary for effective GABA production [28,80]. During the fermentation of LAB, the pH value of the culture continuously decreases. GABA production of S. thermophilus Y2 was significantly increased by means of adjusting the pH of culture medium to pH 4.5 every 12 h by adding NaOH or HCl after free incubation for 24 h [28]. The results have demonstrated that keeping an optimal pH during GABA production is an effective way to enhance GABA yield.
Culture temperature is an important factor for GABA production and growth of bacteria. The fermentation of Lb. brevis strain GABA 100 at 30 °C showed higher production of GABA in black raspberry juice than those at 25 and 37 °C [79]. The optimal temperature is 37 °C for GABA production of Lb. brevis CRL 1942 [77].
GAD activity is the key factor to determine the GABA yield of a strain. It is not only affected by pH and temperature but also by l-glutamic acid and PLP. As the substrate of GAD, l-glutamic acid is an indispensable compound in the medium for the synthesis of GABA by LAB, as LAB cannot synthesize enough l-glutamic acid for GABA production. Generally, monosodium glutamate (MSG) is used in GABA production because it can produce l-glutamic acid by hydrolysis.
Increasing MSG is aimed to stimulate the production of GABA by GAD via the GABA shunt way. At the same time, some researchers have demonstrated that excessive MSG could inhibit cell growth and decrease GABA production [27,28,77]. The optimal concentrations of MSG are different for various microorganisms in GABA production. The GABA production of S. thermophillus Y2 has no obvious change at the range from 10 to 20 g/L MSG [28]. The optimal MSG content is 270 mM for GABA production of Lb. brevis CRL 1942 [77]. MSG significantly induced GABA production in Lb. plantarum CGMCC 1.2437T during fermentation [27]. At conditions containing 100 mM L-MSG, the GABA yield was 721.35 mM, which was 7.7 times more than that without supplement MSG. Transcriptomic analysis revealed that MSG increased the expression of key enzymes of carbohydrate metabolism, fatty acid synthesis, and amino acid metabolism [27].
PLP can increase GAD activity by acting as a cofactor for the enzyme GAD. The effect of PLP varies with the time of the addition of PLP. It was found that PLP could greatly promote GABA production of Lb. paracasei at 10 or 100 μM concentrations in the initial culture medium [80]. Yang et al. (2008) investigated the effect of PLP on GABA production. A 0.02 mM PLP sample was added in medium at different culture times respectively [28]. The addition of PLP could enhance GABA synthesis. Especially, when PLP was added at 48 h of fermentation, the GABA yield was much higher than that of PLP addition at 0 and 24 h of fermentation. It was speculated that PLP could have easily been influenced by some metabolites and lost its function as coenzyme of GAD during the fermentation. Therefore, it may be more efficient to enhance GABA yield by the addition of PLP at 48 h.
4.2. Genetic Engineering
Genetic engineering is an important strategy to improve GABA bioconversion and enhance the yield of GABA through the directed modulation of metabolic pathways. The direct approach is to overexpress the key enzyme GAD. Additional GAD-encoding genes were heterologously or homologously overexpressed in model strains E. coli, Corynebacterium glutamicum, and LAB such as Lb. sakei, Lb. plantarum, Bifidobacterium longum, and so on [58,60,81,82,83,84]. The efficiency of GABA producers was obviously improved. A recombinant C. glutamicum was constructed by the co-expression of two GAD genes, gadB1 and gadB2, from Lb. brevis Lb85 [85]. Compared with the gadB1 or gadB2 single-expressing strains, GABA production by the co-expressing strain increased more than two-fold, and up to 18.66 ± 2.11 g/L after 84 h of fermentation.
In addition to overexpression of the GAD gene, the other genes in the pathway of GABA synthesis are overproduced in order to increase GABA productivity of strain, such as the glutamate: GABA antiporter gene gadC and the regulator gene gadR. The GABA yield of the engineered E. coli increased 2.5-fold in the early culture period via the co-overexpression of gad and gadC [82]. C. glutamicum strain ATCC 13032 was genetically engineered to synthesize GABA by the introduction of the gadB2, gadC, and gadR complex from Lb. brevis Lb85, and the GABA yield was up to 2.15 g/L with 72 h [86].
Furthermore, enhanced GABA biosynthesis efficiency and GABA productivity have also been achieved by inactivation of competing pathways of the GABA production. GABA aminotransferase gene gabT redirects GABA into the TCA cycle and results in GABA degradation. When the genes gadB and gadC were co-overexpressed in the gabT mutant strain, a final GABA concentration increased from 5.09 to 5.46 g/L [87].
GABA production needs sufficient precursor (l-glutamate), but GABA-producing LAB cannot naturally synthesize high concentrations of this compound; therefore, the fermentation process for GABA production by LAB must supply exogenous l-glutamate. Some overproducing l-glutamate recombinant strains have been developed to provide l-glutamate [88,89]. The recombinant E. coli producing 0.92 g/L of GABA directly from 10 g/L of glucose was constructed by the introduction of a synthetic protein scaffold consisting of three enzymes (isocitrate dehydrogenase, glutamate synthase, and GAD) of the GABA pathway [88]. Moreover, the novel GABA production system has been engineered for GABA production based on glucose medium by the co-localization of GABA transporter GadC and GABA shunt enzymes (i.e., succinate semialdehyde dehydrogenase and GABA-AT) [89].
Additionally, C. glutamicum is used to produce GABA using endogenous l-glutamate because of its strong amino acid producing ability [90]. A recombinant C. glutamicum harboring E. coli-derived GAD GadB yielded 8 g/L of GABA without the addition of l-glutamate after 96 h at 30 °C [91]. The serine/threonine protein kinase G gene pknG controls the activity of 2-oxoglutarate dehydrogenase (ODH) in the TCA branch point leading to glutamate synthesis [92]. GadB was expressed in the C. glutamicum strain GAD ∆pknG containing a deletion of the pknG gene in order to enhance GABA production by increasing the intracellular concentration of its precursor l-glutamate. The C. glutamicum strain GAD ∆pknG is an efficient strain in the de novo biosynthesis of GABA using its own accumulated l-glutamate, its GABA yield is 2.29-fold higher than that of strain GAD [92].
It was also found that GABA production was significantly increased by improvement of l-glutamate supplement via deletion of the 2-oxoglutarate decarboxylase subunit gene odhA or pyruvate carboxylase gene pyc [93]. C. glutamicum recombinants of overexpression of the phosphoenolpyruvate carboxylase gene ppc and/or deletion of the malate dehydrogenase gene mdh were also constructed to increase the l-glutamate supplement by efficient oxaloacetate supply [94].
Generally, the optimal pH of GAD activity is about 4.0–5.0, which is not good for the growth of GABA-producing strains. To expand the active pH range of the GAD GadB1 from Lb. brevis Lb85, three excellent mutants—GadB1E312S, GadB1T17I/D294G/Q346H, and GadB1T17I/D294G/E312S/Q346H—were obtained by directed evolution and site-specific mutagenesis. The GAD of these mutants showed higher activity and catalytic efficiency at a near-neutral pH (6.0) [95].
In order to resolve the discrepancy of optimal pH between GAD activity and cell growth, recombinant C. glutamicum strains were constructed by expressing E. coli GAD mutants with an expanded pH range activity; it provided a balanced condition for cell growth and GABA production. The recombinant strain could efficiently synthesize GABA in a culture pH around 5–7, and it produced 5.89 ± 0.35 g/L GABA in GP1 medium at pH 7.0 under the PH36 promoter, which was 17-fold higher than that of C. glutamicum expressing wild-type E. coli GAD in the same conditions [96].
In a word, genetic engineering is a useful strategy to improve GAD activity.
4.3. Physiology-Oriented Engineering Strategy
LAB are usually faced with various environmental stresses during the fermentation process and industry application, including acid, cold, heat, drying, oxidative stress, and so forth [97,98]. In response to these challenges, LAB strains need not only excellent metabolic capabilities but also strong physiological robustness and environmental fitness [99,100,101]. Therefore, physiology-oriented engineering has become an important way to increase work efficiency of industrially useful strains by improving their physiological performances [101].
The decreasing pH of the environment is a key type of stress for cell growth during GABA production. The acid-resistance system of LAB includes a proton pump, neutralization processes, DNA and protein damage repair, and so forth [97]. As the common acid resistance pathway, FoF1-ATPase regulates the cytoplasmic pH by means of expelling intracellular protons and generating the proton motive force [102]. GABA biosynthesis is archived through the decarboxylation of glutamate in the cytoplasm, of which this process needs to consume intracellular protons. Therefore, GAD system can maintain the intracellular pH value of bacteria and plays an important role in the acid resistance of cell [103]. In order to shift the influx of protons toward the GAD system and increase GABA production, a FoF1-ATPase-deficient and GAD-overexpressed Lb. brevis strain NRA6 was constructed [101]. Compared with those of the wild-type strain, the GABA conversion rate and yield of NRA6 obviously increased and was up to 98.42% and 43.65 g/L [101].
During the fermentation process, LAB will produce H2O2, which has potentially toxic effects for cells and inhibits cell growth [98]. In order to improve the antioxidative property of strains, a heme-dependent catalase (CAT) encoding gene katE of Lb. brevis CGMCC1306 was overexpressed. The recombinant strain Lb. brevis CAT showed a marked increased survival (823-fold) in an oxidizing environment [104]. Furthermore, the GABA production ability of the engineered Lb. brevis CAT was obviously enhanced and reached 66.4 g/L.
The results offer references to enhance the GABA production level by means of improving the technological properties of strains with physiology-oriented engineering strategies.
4.4. The Co-Culture Engineering
At present, the use of more than one microorganism in the fermentation industry is popular because some substances produced by co-culture strains could improve each other’s growth [105,106]. For this reason, co-fermentation with different strains is a very important and promising way for the high productivity of GABA. Some reports have described the production of GABA by co-fermentation.
As common co-culturing strains, S. thermophilus IFO13957 and Lb. delbrueckii subsp. bulgaricus IAM1120 were co-fermented and significantly raised GABA levels in the medium up to 15 mM, while when two strains were cultured separately, S. thermophilus IFO13957 and Lb. bulgaricus IAM1120 produced GABA around 0.25 and 0.15 mM, respectively [107].
A large number of Lb. brevis strains with high GABA yields were identified. Generally, Lb. brevis is isolated from fermented vegetables and do not have proteolytic activity because of the absence of extracellular proteinase-encoding genes. In order to resolve the limited application of Lb. brevis in fermented milk, a co-culture approach was adopted [108,109]. Conventional dairy starters were used in the co-fermentation because of their high proteolytic activity. They can break down milk protein into peptides and offer sufficient nutrition for Lb. brevis. A GABA-producing Lb. brevis 877G strain was used to ferment milk with the protease producer Lb. sakei 795, resulting in a significant increase in the GABA production (22.51 mM) [108].
One study demonstrated the dairy starter S. thermophilus YI-B1 could promote the viability and GABA biosynthesis of Lb. brevis NPS-QW-145 in milk [109]. The maximum concentration of GABA was 314 mg/kg containing 2 g/L of MSG supplement. During the fermentation, S. thermophilus provided sufficient amino acids or peptides and released formic acid, folic acid, and fatty acids, which could increase the growth of the Lactobacillus strains. However, Lb. bulgaricus had no obvious effect on the GABA production of Lb. brevis in milk [109]. It was speculated that Lb. bulgaricus and Lb. brevis belonged to the same Lactobacillus genus and were competitive. It was noted that not all tested strains of S. thermophilus could improve the GABA yield of Lb. brevis 145. This result indicated that the promoting of effect depends on the strain specificity.
Lb. brevis GABA 100 isolated from Korean kimchi displayed high GABA-producing ability [79]. A co-cultivation of Lb. brevis GABA 100 and Bifidobacterium bifidum BGN4 increased the GABA production in the fermentation of Gastrodia elata [110]. It was suggested the enhancement of GABA was achieved by the combination, further decreased the culture pH, and provided a suitable pH environment compared with the single culture of Lb. brevis GABA 100.
A combination of Le. mesenteroides SM and Lb. plantarum K154 was used for the production of a water dropwort (Oenanthe javanica DC)-based functional fermented food rich in GABA [111]. Le. mesenteroides is a dominant bacterium at the start of kimchi fermentation, is capable of producing biopolymers and CO2, and provides a thick broth and an enhanced acidic growth environment for the GABA-producing strain Lb. plantarum K154.
TRSF flour (turmeric and roasted soybean flour) was fermented by the two microorganism strains Bacillus subtilis HA and Lb. plantarum K154. It was found that, in the final stage of fermentation, GABA accumulation from co-fermentation was higher, up to 1.78% more than that of the single-fermented system [112]. B. subtilis HA and Lb. plantarum EJ2014 were used for the two-step fermentation of Cucurbita moschata, which is a pumpkin species rich in fiber, minerals, and carotenoids [113]. B. subtilis HA starter was inoculated in pumpkin paste with 5% MSG (w/w), firstly, and cultured for 1 d. Subsequently, Lb. plantarum EJ2014 was added and incubated for 7 d. The co-fermented pumpkin contained 1.47% GABA. B. subtilis could synthesize PLP and boost the GAD activity of Lb. plantarum. At the same time, B. subtilis has strong proteinase activity and offers protein hydrolysate for the growth of Lb. plantarum.
The co-cultures of LAB and fungi for GABA production had been reported [114]. Co-cultivation of Lb. plantarum K154 and fungus Ceriporia lacerate efficiently produced GABA (15.53 mg/mL) and other functional ingredients, such as peptides and polysaccharides. C. lacerate produced plenty of exopolysaccharides, protease, cellulose, and α-amylase, which provide rich nutrients for the growth of Lb. plantarum. Recently, a two-step fermentation process for GABA production was developed by Candida rugosa 8YB and Lactobacillus futsaii CS3, the former produces l-glutamic acid, and the latter utilizes l-glutamic acid to produce GABA [115]. Therefore, co-culturing with different strains seems to be a promising way to generate GABA.
5. Conclusions
Abundant GABA-producing LAB resources have great potential for developing naturally fermented functional food products. The elucidation of the GABA biosynthetic pathway and its regulation mechanism, and the understanding of biochemical properties of the key enzyme GAD in GABA synthesis, lay the theory to improve GABA production at genetic and metabolic levels. Although some studies have partly clarified molecular mechanisms of GABA biosynthesis, the comprehensive mechanism of GABA biosynthesis, especially the regulatory mechanism, needs further study. A large number of GABA-producing LAB genomes have been published, and complete genome information will deepen the comprehension of GABA metabolic activities of strains at the molecular level. GABA synthesis in LAB has been associated with acid resistance of strains, and it has obvious effects on cell physiology. The knowledge of GABA-producing cell physiology offers references to enhance the GABA production level by means of improving cellular technological properties with physiology-oriented engineering strategies. A number of studies suggested that genetic engineering is an effective approach to promote GABA bioconversion through directed modulation of the GABA metabolic pathway, such as the overexpression of GAD, GABA antiporter GadC and regulator, the inactivation of competitive pathway of GABA synthesis, and the improvement of l-glutamate supply. At the same time, co-culture engineering is also an emerging approach with significant advantages for the production of GABA, especially those that effectively utilize raw materials as the precursors for GABA synthesis and produce a variety of beneficial products. The continued advances in both synthetic biology and genetic engineering will promote GABA production of LAB to meet people’s demand for GABA.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/1422-0067/21/3/995/s1.
Author Contributions
Y.C. developed the ideas presented in this manuscript, collected the literature, and wrote the manuscript. X.Q. contributed with discussion and evaluation of the manuscript. K.M. and S.N. collected corresponding data and drew Figure 1, Figure 2 and Figure 3, Table 1, and Supplementary Tables S1 and S2. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant numbers 31471712 and 31371827.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| CAT | Catalase |
| GABA | Gamma-aminobutyric acid |
| GABA-AT | GABA aminotransferase |
| GAD | Glutamate decarboxylase |
| GDH | l-glutamate dehydrogenase |
| GRAS | generally recognized as safe |
| LAB | lactic acid bacteria |
| MSG | monosodium glutamate |
| ODHC | 2-oxoglutarate dehydrogenase complex |
| PLP | pyridoxal-5′-phosphate |
| SSA | succinic semialdehyde |
| SSADH | succinate semialdehyde dehydrogenase |
| TCA | tricarboxylic acid |
References
- Wu, Q.L.; Shah, N.P. High γ-aminobutyric acid production from lactic acid bacteria: Emphasis on Lactobacillus brevis as a functional dairy starter. Crit. Rev. Food Sci. Nutr. 2017, 57, 3661–3672. [Google Scholar] [CrossRef] [Green Version]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Food 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Wu, J.Y.; Matsuda, T.; Roberts, E. Purification and characterization of glutamate decarboxylase from mouse brain. J. Biol. Chem. 1973, 248, 3029–3034. [Google Scholar]
- Okada, T.; Sugishita, T.; Murakami, T.; Murai, H.; Saikusa, T.; Horino, T.; Onoda, A.; Kajmoto, O.; Takahashi, R.; Takahashi, T. Effect of the defatted rice germ enriched with GABA for sleeplessness depression.; autonomic disorder by oral administration. J. Jpn. Soc. Food Sci. Technol. 2000, 47, 596–603. [Google Scholar] [CrossRef] [Green Version]
- Abdou, A.M.; Higashiguchi, S.; Horie, K.; Kim, M.; Hatta, H.; Yokogoshi, H. Relaxation and immunity enhancement effects of γ-aminobutyric acid (GABA) administration in humans. Biofactors 2006, 26, 201–208. [Google Scholar] [CrossRef]
- Wong, C.G.; Bottiglieri, T.; Snead, O.C. GABA, gammahydroxybutyric acid, and neurological disease. Ann. Neurol. 2003, 54, 3–12. [Google Scholar] [CrossRef]
- Hayakawa, K.; Kimura, M.; Kasaha, K.; Matsumoto, K.; Sansawa, H.; Yamori, Y. Effect of a γ-aminobutyric acidenriched dairy product on the blood pressure of spontaneously hypertensive and normotensive Wistar-Kyoto rats. Brit. J. Nutr. 2004, 92, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.H.; Moon, Y.J.; Soh, J.R.; Cha, Y.S. Effect of water extract of germinated brown rice on adiposity and obesity indices in mice fed a high fat diet. J. Food Sci. Nutr. 2005, 10, 251–256. [Google Scholar] [CrossRef]
- Leventhal, A.G.; Wang, Y.; Pu, M.; Zhou, Y.; Ma, Y. GABA and its agonists improved visual cortical function in senescent monkeys. Science 2003, 300, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Uda, I.; Tagami, K.; Furuya, Y.; Endo, Y.; Fujimoto, K. The production of a new tempeh like fermented soybean containing a high level of γ-aminobutyric acid by anaerobic incubation with Rhizopus. Biosci. Biotechnol. Biochem. 2003, 67, 1018–1023. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lee, C.; Pan, T. Improvement of monacolin K, gamma-aminobutyric acid and citrinin production ratio as a function of environmental conditions of Monascus purpureus NTU 601. J. Ind. Microbiol. Biot. 2003, 30, 669–676. [Google Scholar] [CrossRef]
- Liu, W.J.; Pang, H.L.; Zhang, H.P.; Cai, Y.M. Biodiversity of lactic acid bacteria. In Lactic Acid Bacteria-Fundamentals and Practice; Zhang, H.P., Cai, Y.M., Eds.; Springer Publishing: New York, NY, USA, 2014; pp. 103–203. [Google Scholar]
- Seo, M.J.; Nam, Y.D.; Lee, S.Y.; Park, S.L.; Yi, S.H.; Lim, S.I. Expression and characterization of a glutamate decarboxylase from Lactobacillus brevis 877G producing gamma-aminobutyric acid. Biosci. Biotechnol. Biochem. 2013, 77, 853–856. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qiu, T.; Gao, D.; Cao, Y. Medium optimization for production of gamma-aminobutyric acid by Lactobacillus brevis NCL912. Amino Acids 2010, 38, 1439–1945. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qiu, T.; Huang, G.; Cao, Y. Production of gamma-aminobutyric acid by Lactobacillus brevis NCL912 using fed-batch fermentation. Microb. Cell Fact. 2010, 9, 85. [Google Scholar] [CrossRef] [Green Version]
- Park, K.B.; Oh, S.H. Cloning, sequencing and expression of a novel glutamate decarboxylase gene from a newly isolated lactic acid bacterium, Lactobacillus brevis OPK-3. Bioresource Technol. 2007, 98, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Siragusa, S.; de Angelis, M.; di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of γ-aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microb. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Liu, X.; Fu, J.; Wang, S.; Chen, Y.; Chang, K.; Li, H. Substrate sustained release-based high efficacy biosynthesis of GABA by Lactobacillus brevis NCL912. Micro. Cell Fact. 2018, 17, 80. [Google Scholar] [CrossRef] [Green Version]
- Diana, M.; Tres, A.; Quílez, J.; Llombart, M.; Rafecas, M. Spanish cheese screening and selection of lactic acid bacteria with high gamma-aminobutyric acid production. LWT-Food Sci. Technol. 2014, 56, 351–355. [Google Scholar] [CrossRef]
- Park, K.B.; Oh, S.H. Isolation and characterization of Lactobacillus buchneri strains with high gamma-aminobutyric acid producing capacity from naturally aged cheese. Food Sci. Biotechnol. 2006, 15, 86–90. [Google Scholar]
- Cho, Y.R.; Chang, J.Y.; Chang, H.C. Production of γ-aminobutyric acid (GABA) by Lactobacillus buchneri isolated from Kimchi and its neuroprotective effect on neuronal cells. J. Microbiol. Biotechnol. 2007, 17, 104–109. [Google Scholar]
- Gangaraju, D.S.; Murty, V.R.; Prapulla, S.G. Probiotic-mediated biotransformation of monosodium glutamate to γ-aminobutyric acid: Differential production in complex and minimal media and kinetic modelling. Ann. Microbiol. 2014, 64, 229–237. [Google Scholar] [CrossRef]
- Woraharn, S.; Lailerd, N.; Sivamaruthi, B.S.; Wangcharoen, W.; Sirisattha, S.; Peerajan, S.; Chaiyasut, C. Evaluation of factors that influence the L-glutamic and γ-aminobutyric acid production during Hericium erinaceus fermentation by lactic acid bacteria. CyTA-J. Food 2016, 14, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Li, D.; Qin, H. Molecular cloning, expression, and immobilization of glutamate decarboxylase from Lactobacillus fermentum YS2. Electron J. Biotechnol. 2017, 27, 8–13. [Google Scholar] [CrossRef]
- Sun, T.S.; Zhao, S.P.; Wang, H.K.; Cai, C.K.; Chen, Y.F.; Zhang, H.P. ACE-inhibitory activity and gamma-aminobutyric acid content of fermented skim milk by Lactobacillus helveticus isolated from Xinjiang koumiss in China. Eur. Food Res. Technol. 2009, 228, 607–612. [Google Scholar] [CrossRef]
- Franciosi, E.; Carafa, I.; Nardin, T.; Schiavon, S.; Poznanski, E.; Cavazza, A.; Larcher, R.; Tuohy, K.M. Biodiversity and γ-aminobutyric acid production by lactic acid bacteria isolated from traditional alpine raw cow’s milk cheeses. BioMed Res. Int. 2015, 2015, 625740. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, K.; Jiang, Y.; Feng, X.; Li, L.; Dang, F.; Zhang, W.; Man, C. Transcriptomic response to GABA-producing Lactobacillus plantarum CGMCC 1.2437T induced by L-MSG. PLoS ONE 2018, 13, e0199021. [Google Scholar] [CrossRef]
- Yang, S.Y.; Lu, F.X.; Lu, Z.X.; Bie, X.M.; Jiao, Y.; Sun, L.J.; Yu, B. Production of γ-aminobutyric acid by Streptococcus salivarius subsp. thermophilus Y2 under submerged fermentation. Amino Acids 2008, 34, 473–478. [Google Scholar]
- Small, P.L.; Waterman, S.R. Acid stress, anaerobiosis and gadCB: Lessons from Lactococcus lactis and Escherichia coli. Trends Microbiol. 1998, 6, 214–216. [Google Scholar] [CrossRef]
- Lyu, C.J.; Zhao, W.R.; Peng, C.L.; Hu, S.; Fang, H.; Hua, Y.J.; Yao, S.J.; Huang, J.; Mei, L.H. Exploring the contributions of two glutamate decarboxylase isozymes in Lactobacillus brevis to acid resistance and γ-aminobutyric acid production. Microb. Cell Fact. 2018, 17, 1–14. [Google Scholar] [CrossRef]
- Wu, Q.; Tun, H.M.; Law, Y.S.; Khafipour, E.; Shah, N.P. Common distribution of gad operon in Lactobacillus brevis and its GadA contributes to efficient GABA synthesis toward cytosolic near-neutral pH. Front. Microbiol. 2017, 8, 206. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.L.; Shah, N.P. Restoration of GABA production machinery in Lactobacillus brevis by accessible carbohydrates, anaerobiosis and early acidification. Food Microbiol. 2018, 69, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA production and structure of gadB/gadC genes in Lactobacillus and Bifidobacterium strains from human microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Dover, S.; Halpern, Y.S. Control of the pathway of γ-aminobutyrate breakdown in Escherichia coli K-12. J. Bacteriol. 1972, 110, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feehily, C.; O’Byrne, C.P.; Karatzas, K.A.G. Functional γ-aminobutyrate shunt in Listeria monocytogenes: Role in acid tolerance and succinate biosynthesis. Appl. Environ. Microb. 2013, 79, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleerebezem, M.; Boekhorst, J.; van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, H.; Toh, H.; Fukuda, S.; Horikawa, H.; Oshima, K.; Suzuki, T.; Murakami, M.; Hisamatsu, S.; Kato, Y.; Takizawa, T.; et al. Comparative genome analysis of Lactobacillus reuteri and Lactobacillus fermentum reveal a genomic island for Reuterin and Cobalamin production. DNA Res. 2008, 15, 151–161. [Google Scholar] [CrossRef]
- Kim, J.F.; Jeong, H.; Lee, J.S.; Choi, S.H.; Ha, M.; Hur, C.G.; Kim, J.S.; Lee, S.; Park, H.S.; Park, Y.H.; et al. Complete genome sequence of Leuconostoc citreum KM20. J. Bacteriol. 2008, 190, 3093–3094. [Google Scholar] [CrossRef] [Green Version]
- Johansson, P.; Paulin, L.; Sade, E.; Salovuori, N.; Alatalo, E.R.; Bjorkroth, K.J.; Auvinen, P. Genome sequence of a food spoilage lactic acid bacterium.; Leuconostoc gasicomitatum LMG 18811T, in Association with specific spoilage reactions. Appl. Environ. Microbiol. 2011, 77, 4344–4351. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.M.; Cho, Y.J.; Kim, B.K.; Roe, J.H.; Kang, S.O.; Nahm, B.H.; Jeong, G.; Han, H.U.; Chun, J. Complete genome sequence analysis of Leuconostoc kimchii IMSNU 11154. J. Bacteriol. 2010, 192, 3844–3845. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [Green Version]
- Nejati, F.; Rizzello, C.D.; Cagno, R.; Sheikh-Zeinoddin, M.; Diviccaro, A.; Minervini, F.; Gobetti, M. Manufacture of a functional fermented milk enriched of angiotensin-I converting enzyme (ACE)-inhibitory peptides and gamma amino butyric acid (gaba). LWT-Food Sci. Technol. 2013, 51, 183–189. [Google Scholar] [CrossRef]
- Demirbaş, F.; Ispirli, H.; Kurnaz, A.A.; Yilmaz, M.T. Antimicrobial and functional properties of lactic acid bacteria isolated from sourdoughs. LWT-Food Sci. Technol. 2017, 79, 361–366. [Google Scholar] [CrossRef]
- Barla, F.; Koyanagi, T.; Tokuda, N.; Matsui, H.; Katayama, T.; Kumagai, H.; Michihata, T.; Sasaki, T.; Tsuji, A.; Enomoto, T. The γ-aminobutyric acid-producing ability under low pH conditions of lactic acid bacteria isolated from traditional fermented foods of Ishikawa Prefecture.; Japan.; with a strong ability to produce ACE-inhibitory peptides. Biotechnol. Rep. 2016, 10, 105–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegmann, U.; O’Connell-Motherway, M.; Zomer, A.; Buist, G.; Shearman, C.; Canchaya, C.; Ventura, M.; Goesmann, A.; Gasson, M.J.; Kuipers, O.P.; et al. Complete genome sequence of the prototype lactic acid bacterium Lactococcus lactis subsp. cremoris MG1363. J. Bacteriol. 2007, 189, 3256–3270. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Lu, Y.; Teng, K.-L.; Chen, M.-L.; Zheng, H.-J.; Zhu, Y.-Q.; Zhong, J. Complete genome sequence of Lactococcus lactis subsp. lactis CV56, a probiotic strain isolated from the vaginas of healthy women. J. Bacteriol. 2011, 193, 2886–2887. [Google Scholar] [PubMed] [Green Version]
- Alexandraki, V.; Kazou, M.; Blom, J.; Pot, B.; Tsakalidou, E.; Papadimitriou, K. The complete genome sequence of the yogurt isolate Streptococcus thermophilus ACA-DC 2. Stand Genomic Sci. 2017, 12, 18. [Google Scholar] [CrossRef] [Green Version]
- Renye, J.A.; Needleman, D.S.; Somkuti, G.A.; Steinberg, D.H. Complete genome sequence of Streptococcus thermophilus strain B59671, which naturally produces the broad-spectrum bacteriocin thermophilin 110. Genome Announc. 2017, 5, e01213-17. [Google Scholar] [CrossRef] [Green Version]
- Treu, L.; Vendramin, V.; Bovo, B.; Campanaro, S.; Corich, V.; Giacomini, A. Whole-genome sequences of Streptococcus thermophilus strains TH1435 and TH1436, isolated from raw goat milk. Genome Announc. 2014, 2, e01129-13. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Chen, X.; Wang, J.; Zhao, W.; Shao, Y.; Wu, L.; Zhou, Z.; Sun, T.; Wang, L.; Meng, H.; et al. Complete genome sequence of Streptococcus thermophilus strain ND03. J. Bacteriol. 2011, 193, 793–794. [Google Scholar] [CrossRef] [Green Version]
- Linares, D.M.; Arboleya, S.; Ross, R.P.; Stanton, C. Complete genome sequence of the gamma-aminobutyric acid-producing strain Streptococcus thermophilus APC151. Genome Announc. 2017, 5, e00205-17. [Google Scholar] [CrossRef] [Green Version]
- Evivie, S.E.; Li, B.; Ding, X.; Meng, Y.; Yu, S.; Du, J.; Xu, M.; Li, W.; Jin, D.; Huo, G.; et al. Complete genome sequence of Streptococcus thermophilus KLDS 3.1003.; a strain with high antimicrobial potential against foodborne and vaginal pathogens. Front. Microbiol. 2017, 8, 1238. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Li, L.; Zhao, Q.; Wang, P.; Zuo, X. Complete genome sequence of bile-isolated Enterococcus avium strain 352. Gut Pathog. 2019, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Li, H.X.; Li, W.M.; Liu, X.H.; Cao, Y.S. GadA gene locus in Lactobacillus brevis NCL912 and its expression during fed-batch fermentation. FEMS Microbiol. Lett. 2013, 349, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Chang, K.; Ding, G.; Wu, H.; Chen, Y.; Jia, M.; Liu, X.; Wang, S.; Jin, Y.; Pan, H.; et al. Genomic insights into a robust gamma-aminobutyric acid-producer Lactobacillus brevis CD0817. AMB Express 2019, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Shim, J.M.; Yao, Z.; Kim, J.A.; Kim, H.J.; Kim, J.H. Characterization of a glutamate decarboxylase (GAD) from Enterococcus avium M5 isolated from Jeotgal, a Korean fermented seafood. J. Microbiol. Biotechnol. 2017, 27, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Zhang, J.; Ma, S.; Wang, L.; Wang, D.; Zhang, J.; Gao, Q. Purification and characterization of glutamate decarboxylase from Enterococcus raffinosus TCCC11660. J. Ind. Microbiol. Biot. 2017, 44, 817–824. [Google Scholar] [CrossRef]
- Park, J.Y.; Jeong, S.J.; Kim, J.H. Characterization of a glutamate decarboxylase (GAD) gene from Lactobacillus zymae. Biotechnol. Lett. 2014, 36, 1791–1799. [Google Scholar] [CrossRef]
- Huang, J.; Mei, L.H.; Sheng, Q.; Yao, S.J.; Lin, D.Q. Purification and characterization of glutamate decarboxylase of Lactobacillus brevis CGMCC 1306 isolated from fresh milk. Chin. J. Chem. Eng. 2007, 15, 157–161. [Google Scholar] [CrossRef]
- Fan, E.; Huang, J.; Hu, S.; Mei, L.; Yu, K. Cloning, sequencing and expression of a glutamate decarboxylase gene from the GABA-producing strain Lactobacillus brevis CGMCC 1306. Ann. Microbiol. 2012, 62, 689–698. [Google Scholar] [CrossRef]
- Lim, H.S.; Seo, D.H.; Cha, I.T.; Lee, H.; Nam, Y.D.; Seo, M.J. Expression and characterization of glutamate decarboxylase from Lactobacillus brevis HYE1 isolated from kimchi. World J. Microbiol. Biotechnol. 2018, 34, 44. [Google Scholar] [CrossRef]
- Uenoa, Y.; Hayakawa, K.; Takahashi, S.; Oda, K. Purification and characterization of glutamate decarboxylase from Lactobacillus brevis IFO 12005. Biosci. Biotechnol. Biochem. 1997, 61, 1168–1171. [Google Scholar] [CrossRef] [PubMed]
- Hirage, K.; Ueno, Y.; Oda, K. Glutamate decarboxylase from Lactobacillus brevis: Activation by ammonium sulfate. Biosci. Biotechnol. Biochem. 2008, 72, 1299–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Song, T.; Kong, W.; Yoon, M. Characterization of γ-aminobutyric acid (GABA) produced by a lactic acid bacterium from button mushroom bed. J. Mushroom Sci. Prod. 2013, 11, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Komatsuzaki, N.; Nakamura, T.; Kimura, T.; Shima, J. Characterization of glutamate decarboxylase from a high gamma-aminobutyric acid (GABA)-producer, Lactobacillus paracasei. Biosci. Biotechnol. Biochem. 2008, 72, 278–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.M.; Kim, H.; Joo, Y.; Lee, S.J.; Lee, Y.J.; Lee, S.J.; Lee, D.W. Characterization of glutamate decarboxylase from Lactobacillus plantarum and its C-terminal function for the pH dependence of activity. J. Agric. Food Chem. 2014, 62, 12186–12193. [Google Scholar] [CrossRef] [PubMed]
- Sa, H.D.; Park, J.Y.; Jeong, S.J.; Lee, K.W.; Kim, J.H. Characterization of glutamate decarboxylase (GAD) from Lactobacillus sakei A156 isolated from jeot-gal. J. Microbiol. Biotechnol. 2015, 25, 696–703. [Google Scholar] [CrossRef]
- Yu, J.J.; Oh, S.H. γ-Aminobutyric acid production and glutamate decarboxylase activity of Lactobacillus sakei OPK2- 59 isolated from Kimchi. Korean J. Microbiol. 2011, 7, 104–109. [Google Scholar]
- Nomura, M.; Kimoto, H.; Someya, Y.; Furukawa, S.; Suzuki, I. Production of gamma-aminobutyric acid by cheese starters during cheese ripening. J. Dairy Sci. 1998, 81, 1486–1491. [Google Scholar] [CrossRef]
- Nomura, M.; Nakajima, I.; Fujita, Y.; Kobayashi, M.; Kimoto, H.; Suzuki, I.; Aso, H. Lactococcus lactis contains only one glutamate decarboxylase gene. Microbiology 1999, 145, 1375–1480. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Lin, L.; Hu, S.; Huang, J.; Mei, L. C-terminal truncation of glutamate decarboxylase from Lactobacillus brevis CGMCC 1306 extends its activity toward near-neutral pH. Enzym. Microb. Technol. 2012, 50, 263–269. [Google Scholar] [CrossRef]
- Huang, J.; Fang, H.; Gai, Z.-C.; Mei, J.-Q.; Li, J.-N.; Hu, S.; Lv, C.-J.; Zhao, W.-R.; Mei, L.-H. Lactobacillus brevis CGMCC 1306 glutamate decarboxylase: Crystal structure and functional analysis. Biochem. Biophys. Res. Commun. 2018, 503, 1703–1709. [Google Scholar] [PubMed]
- Ma, D.; Lu, P.; Yan, C.; Fan, C.; Yin, P.; Wang, J.; Shi, Y. Structure and mechanism of a glutamate-GABA antiporter. Nature 2012, 483, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.W.; Leenhouts, K.; Burghoorn, J.; Brands, J.R.; Venema, G.; Kok, J. A chloride-inducible acid resistance mechanism in Lactococcus lactis and its regulation. Mol. Microbiol. 1998, 27, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Ren, C.; Xu, Y. Deciphering the crucial roles of transcriptional regulator GadR on gamma-aminobutyric acid production and acid resistance in Lactobacillus brevis. Microb. Cell Fact. 2019, 18, 108. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cao, Y. Lactic acid bacterial cell factories for gamma-aminobutyric acid. Amino Acids 2010, 39, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Villegas, J.M.; Brown, L.; de Giori, G.S.; Hebert, E.M. Optimization of batch culture conditions for GABA production by Lactobacillus brevis CRL 1942.; isolated from quinoa sourdough. LWT-Food Sci. Technol. 2016, 67, 22–26. [Google Scholar] [CrossRef]
- Wu, C.H.; Hsueh, Y.H.; Kuo, J.M.; Liu, S.J. Characterization of a potential probiotic Lactobacillus brevis RK03 and efficient production of γ-aminobutyric acid in batch fermentation. Int. J. Mol. Sci. 2018, 19, 143. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Lee, M.Y.; Ji, G.E.; Lee, Y.S.; Hwang, K.T. Production of γ-aminobutyric acid in black raspberry juice during fermentation by Lactobacillus brevis GABA100. Int. J. Food Microbiol. 2009, 130, 12–16. [Google Scholar] [CrossRef]
- Komatsuzaki, N.; Shima, J.; Kawamoto, S.; Momose, H.; Kimura, T. Production of γ-aminobutyric acid (GABA) by Lactobacillus paracasei isolated from traditional fermented foods. Food Microbiol. 2005, 22, 497–504. [Google Scholar] [CrossRef]
- Kook, M.; Seo, M.; Cheigh, C.; Lee, S.; Pyun, Y.; Park, H. Enhancement of γ-amminobutyric acid production by Lactobacillus sakei B2–16 expressing glutamate decarboxylase from Lactobacillus plantarum ATCC 14917. J. Korean Soc. Appl. Biol. Chem. 2010, 53, 816–820. [Google Scholar] [CrossRef]
- Le Vo, T.D.; Ko, J.; Park, S.J.; Lee, S.H.; Hong, S.H. Efficient gamma-aminobutyric acid bioconversion by employing synthetic complex between glutamate decarboxylase and glutamate/GABA antiporter in engineered Escherichia coli. J. Ind. Microbiol. Biotechnol. 2013, 40, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Park, K.B.; Ji, G.E.; Park, M.S.; Oh, S.H. Expression of rice glutamate decarboxylase in Bifidobacterium longum enhances gamma-aminobutyric acid production. Biotechnol. Lett. 2005, 27, 1681–1684. [Google Scholar] [CrossRef] [PubMed]
- Tajabadi, N.; Baradaran, A.; Ebrahimpour, A.; Rahim, R.A.; Bakar, F.A.; Manap, M.Y.A.; Mohammed, A.S.; Saari, N. 2015. Overexpression and optimization ofglutamate decarboxylase in Lactobacillus plantarum Taj-Apis362 for high gamma-aminobutyric acid production. Microbiol. Biotechnol. 2015, 8, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Jiang, J.; Li, Y.; Li, Y.; Xie, Y. Enhancement of γ-aminobutyric acid production in recombinant Corynebacterium glutamicum by co-expressing two glutamate decarboxylase genes from Lactobacillus brevis. J. Ind. Microbiol. Biotechnol. 2013, 40, 1285–1296. [Google Scholar] [CrossRef]
- Shi, F.; Li, Y. Synthesis of γ-aminobutyric acid by expressing Lactobacillus brevis-derived glutamate decarboxylase in the Corynebacterium glutamicum strain ATCC 13032. Biotechnol. Lett. 2011, 33, 2469–2674. [Google Scholar] [CrossRef]
- Le Vo, T.D.; Kim, T.W.; Hong, S.H. Effects of glutamate decarboxylase and gamma-aminobutyric acid (GABA) transporter on the bioconversion of GABA in engineered Escherichia coli. Bioprocess Biosyst. Eng. 2012, 35, 645–650. [Google Scholar] [CrossRef]
- Pham, V.D.; Lee, S.H.; Park, S.J.; Hong, S.H. Production of gamma-aminobutyric acid from glucose by introduction of synthetic scaffolds between isocitrate dehydrogenase.; glutamate synthase and glutamatede carboxylase in recombinant Escherichia coli. J. Biotechnol. 2015, 207, 52–57. [Google Scholar] [CrossRef]
- Pham, V.D.; Somasundaram, S.; Lee, S.H.; Park, S.J.; Hong, S.H. Engineering the intracellular metabolism of Escherichia coli to produce gamma-aminobutyric acid by co-localization of GABA shunt enzymes. Biotechnol. Lett. 2016, 38, 321–327. [Google Scholar] [CrossRef]
- Heider, S.A.E.; Wendisch, V.F. Engineering microbial cell factories: Metabolic engineering of Corynebacterium glutamicum with a focus on non-natural products. Biotechnol. J. 2015, 10, 1170–1184. [Google Scholar] [CrossRef]
- Takahashi, C.; Shirakawa, J.; Tsuchidate, T.; Okai, N.; Hatada, K.; Nakayama, H.; Tateno, T.; Ogino, C.; Kondo, A. Robust production of gamma-aminobutyric acid using recombinant Corynebacterium glutamicum expressing glutamate decarboxylase from Escherichia coli. Enzyme Microb. Technol. 2012, 51, 171–176. [Google Scholar] [CrossRef]
- Okai, N.; Takahashi, C.; Hatada, K.; Ogino, C.; Kondo, A. Disruption of pknG enhances production of gamma-aminobutyric acid by Corynebacterium glutamicum expressing glutamate decarboxylase. AMB Express 2014, 4, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Ni, Y.; Shi, F. Deletion of odhA or pyc improves production of gamma aminobutyric acid and its precursor L-glutamate in recombinant Corynebacterium glutamicum. Biotechnol. Lett. 2015, 37, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Zhang, M.; Li, Y.F. Overexpression of ppc or deletion of mdh for improving production of γ-aminobutyric acid in recombinant Corynebacterium glutamicum. World J. Microbiol. Biotechnol. 2017, 33, 122. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Xie, Y.; Jiang, J.; Wang, N.; Li, F.Y.; Wang, X. Directed evolution and mutagenesis of glutamate decarboxylase from Lactobacillus brevis Lb85 to broaden the range of its activity toward a near-neutral pH. Enzyme Microb. Technol. 2014, 61–62, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Yim, S.S.; Lee, S.H.; Kang, T.J.; Park, S.J.; Jeong, K.J. Enhanced production of gamma-aminobutyrate (GABA) in recombinant Corynebacterium glutamicum by expressing glutamate decarboxylase active in expanded pH range. Microb. Cell Fact. 2015, 14, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Cui, Y.H.; Qu, X.J. Mechanisms and improvement of acid resistance in lactic acid bacteria. Arch. Microbiol. 2018, 200, 195–201. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Li, Y. Engineering the antioxidative properties of lactic acid bacteria for improving its robustness. Curr. Opin. Biotechnol. 2013, 24, 142–147. [Google Scholar] [CrossRef]
- Zhu, L.J.; Zhu, Y.; Zhang, Y.P.; Li, Y. Engineering the robustness of industrial microbes through synthetic biology. Trends Microbiol. 2012, 20, 94–101. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, Y.P.; Li, Y. Understanding the industrial application potential of lactic acid bacteria through genomics. Appl. Microbiol. Biotechnol. 2009, 83, 597–610. [Google Scholar] [CrossRef]
- Lyu, C.; Zhao, W.; Hu, S.; Huang, J.; Lu, T.; Jin, Z.; Mei, L.; Yao, S. Physiology-oriented engineering strategy to improve gamma-aminobutyrate production in Lactobacillus brevis. J. Agric. Food Chem. 2017, 65, 858–866. [Google Scholar] [CrossRef]
- Kajfasz, J.K.; Quivey, R.G. Responses of lactic acid bacteria to acid stress. In Stress Responses of Lactic Acid Bacteria; Tsakalidou, E., Papadimitrious, K., Eds.; Springer Publishing: New York, NY, USA, 2011; pp. 23–53. [Google Scholar]
- Feehily, C.; Karatzas, K.A.G. Role of glutamate metabolism in bacterial responses towards acid and other stresses. J. Appl. Microbiol. 2013, 114, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Lyu, C.J.; Hu, S.; Huang, J.; Luo, M.Q.; Lu, T.; Mei, L.H.; Yao, S.J. Contribution of the activated catalase to oxdative stress resistance and γ-aminobutyric acid production in Lactobacillus brevis. Int. J. Food Microbiol. 2016, 238, 302–310. [Google Scholar] [CrossRef]
- Harcombe, W. Novel cooperation experimentally evolved between species. Evolution 2010, 64, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kerner, A.; Burns, M.A.; Lin, X.N. Microdroplet-enabled highly parallel co-cultivation of microbial communities. PLoS ONE 2011, 6, e17019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Hayakawa, K.; Ueno, H. Effect of co-culturing LAB on GABA production. J. Biol. Macromol. 2011, 11, 3–13. [Google Scholar]
- Seo, M.J.; Nam, Y.D.; Park, S.L.; Lee, S.Y.; Yi, S.H.; Lim, S.I. γ-aminobutyric acid production in skim milk co-fermented with Lactobacillus brevis 877G and Lactobacillus sakei 795. Food Sci. Biotechnol. 2013, 22, 751–755. [Google Scholar] [CrossRef]
- Wu, Q.; Law, Y.S.; Shah, N.P. Dairy Streptococcus thermophilus improves cell viability of Lactobacillus brevis NPS-QW-145 and its g-aminobutyric acid biosynthesis ability in milk. Sci. Rep. 2015, 2, 12885. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.A.; Park, M.S.; Kang, S.A.; Ji, G.E. Production of g-aminobutyric acid during fermentation of Gastrodia elata Bl. by co-culture of Lactobacillus brevis GABA 100 with Bifidobacterium bifidum BGN4. Food Sci. Biotechnol. 2014, 23, 459–466. [Google Scholar] [CrossRef]
- Kwon, S.Y.; Garcia, C.V.; Song, Y.C.; Lee, S.P. GABA-enriched water dropwort produced by co-fermentation with Leuconostoc mesenteroides SM and Lactobacillus plantarum K154. LWT-Food Sci. Technol. 2016, 73, 233–238. [Google Scholar] [CrossRef]
- Lim, J.S.; Garcia, C.; Lee, S.P. Optimized production of GABA and γ-PGA in a turmeric and roasted soybean mixture co-fermented by Bacillus subtilis and Lactobacillus plantarum. Food Sci. Technol. Res. 2016, 22, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Park, E.J.; Garcia, C.V.; Youn, S.J.; Park, C.D.; Lee, S.P. Fortification of γ-aminobutyric acid and bioactive compounds in Cucurbita moschata by novel two-step fermentation using Bacillus subtilis and Lactobacillus plantarum. LWT-Food Sci. Technol. 2019, 102, 22–29. [Google Scholar] [CrossRef]
- Lee, E.J.; Lee, S.P. Novel bioconversion of sodium glutamate to γ-amino butyric acid by co-culture of Lactobacillus plantarum K154 in Ceriporia lacerata culture broth. Food Sci. Biotechnol. 2014, 23, 1997–2005. [Google Scholar] [CrossRef]
- Sanchart, C.; Watthanasakphuban, N.; Boonseng, O.; Nguyen, T.H.; Haltrich, D.; Maneerat, S. Tuna condensate as a promising low-cost substrate for glutamic acid and GABA formation using Candida rugosa and Lactobacillus futsaii. Process Biochem. 2018, 70, 29–35. [Google Scholar] [CrossRef]
Figure 1.
The biosynthetic pathway of GABA by microbes. GABA-AT, GABA aminotransferase; GadB/GadA, glutamate decarboxylase; GadC, glutamate: γ-aminobutyrate antiporter; GDH, l-glutamate dehydrogenase; GltB, glutamate synthase; Icd, isocitrate dehydrogenase; MDH, malate dehydrogenase; ODHC, 2-oxoglutarate dehydrogenase complex; PC, pyruvate carboxylase; PEP, phosphoenolpyruvate; PEPC, phosphoenolpyruvate carboxylase; SSADH, succinate semialdehyde dehydrogenase; TCA cycle, tricarboxylic acid cycle. The expression of enzymes in green font will increase GABA production, and the expression of enzymes in red font will decrease GABA production.
Figure 1.
The biosynthetic pathway of GABA by microbes. GABA-AT, GABA aminotransferase; GadB/GadA, glutamate decarboxylase; GadC, glutamate: γ-aminobutyrate antiporter; GDH, l-glutamate dehydrogenase; GltB, glutamate synthase; Icd, isocitrate dehydrogenase; MDH, malate dehydrogenase; ODHC, 2-oxoglutarate dehydrogenase complex; PC, pyruvate carboxylase; PEP, phosphoenolpyruvate; PEPC, phosphoenolpyruvate carboxylase; SSADH, succinate semialdehyde dehydrogenase; TCA cycle, tricarboxylic acid cycle. The expression of enzymes in green font will increase GABA production, and the expression of enzymes in red font will decrease GABA production.

Figure 2.
Genetic organization of the glutamate decarboxylase gene clusters among some lactic acid bacteria (LAB) strains. The number of amino acids within each encoded protein is shown above the corresponding protein. AAP, amino acid permease; Ace, sugar O-acetyltransferase; ACR, amino acid racemase; Ade, adenine deaminase; Adh, zinc-dependent alcohol dehydrogenase family protein; AFA, AAA family ATPase; Akr, aldo/keto reductase; Alt, PLP-dependent aminotransferase family protein; At, metN2, ABC transporter, ATP-binding protein; ATB, ABC transporter ATP-binding protein; ATL, aspartate—tRNA ligase; ATP, ABC transporter permease; Bca, branched-chain amino acid aminotransferase; CAL, carboxylate—amine ligase; CFD, C69 family dipeptidase; CrcB, fluoride efflux transporter; Ctp, cobalt ABC transporter, ATP-binding protein; CTR, Crp/Fnr family transcriptional regulator; DMG, DNA-3-methyladenine glycosylase; DUF, DUF1722 domain-containing protein; DUF1310, DUF1310 family protein; DUF1906, DUF1906 domain-containing protein; DUF4432, DUF4432 family protein; EAL, EAL domain-containing protein; EP, extracellular protein; FCA, formate C-acetyltransferase; FCoAT, formyl-CoA transferase; FtsX, FtsX-like permease family protein; GadB/GadA, glutamate decarboxylase; GadC/GadC1/GadC2, glutamate:gamma-aminobutyrate antiporter; GadR, transcriptional regulator; Gcl, glutamate-cysteine ligase; Gls, glutaminase; GltB, glutamate synthase large subunit; GltC, glutamate synthase subunit beta; GltX, glutamate-tRNA ligase; GuaD, guanine deaminase; Hig, HigA family addiction module antidote protein; Hp, hypothetical protein; HsdS, restriction endonuclease subunit S; HTHP, helix-turn-helix domain-containing protein; LacI, LacI family transcriptional regulator; LarB, nickel pincer cofactor biosynthesis protein; LarC, nickel pincer cofactor biosynthesis protein; LarE, ATP-dependent sacrificial sulfur transferase; LuxS, S-ribosylhomocysteine lyase; LysA, diaminopimelate decarboxylase; LysMP, LysM peptidoglycan-binding domain-containing protein; LysR, LysR family transcriptional regulator; LytF, gamma-d-glutamate-meso-diaminopimelate muropeptidase; MarR, MarR family transcriptional regulator; ME, ImmA/IrrE family metallo-endopeptidase; MFS, MFS transporter; Mic, mechanosensitive ion channel; Mp, membrane protein; MTT, class I SAM-dependent methyltransferase; MurI, glutamate racemase; Nat, N-acetyltransferase; NDLR, nickel-dependent lactate racemase; NDM, N-6 DNA methylase; NPCT, Na/Pi cotransporter family protein; Nudix, NUDIX domain-containing protein; Odr, oxidoreductase; Pck, phosphoenolpyruvate carboxykinase; Per, NCS2 family permease; Pmsk, plasmid maintenance system killer; Rb, ribokinase; Rc, ribonuclease HII; RE, restriction endonuclease; RFP, recombinase family protein; RibY, ribonuclease Y; Rp, 30S ribosomal protein S4; RplQ, ribosomal protein L17; RpoA, DNA-directed RNA polymerase, alpha subunit; SDR, SDR family oxidoreductase; SP, sulfate permease; Spx, Spx/MgsR family RNA polymerase-binding regulatory protein; SseA, sulfurtransferase; Tdk, thymidine kinase; Tn, transposase; TP, l,d-transpeptidase family protein; TR, transcriptional regulator; TSE, threonine/serine exporter family protein; YcxB, YcxB family protein.
Figure 2.
Genetic organization of the glutamate decarboxylase gene clusters among some lactic acid bacteria (LAB) strains. The number of amino acids within each encoded protein is shown above the corresponding protein. AAP, amino acid permease; Ace, sugar O-acetyltransferase; ACR, amino acid racemase; Ade, adenine deaminase; Adh, zinc-dependent alcohol dehydrogenase family protein; AFA, AAA family ATPase; Akr, aldo/keto reductase; Alt, PLP-dependent aminotransferase family protein; At, metN2, ABC transporter, ATP-binding protein; ATB, ABC transporter ATP-binding protein; ATL, aspartate—tRNA ligase; ATP, ABC transporter permease; Bca, branched-chain amino acid aminotransferase; CAL, carboxylate—amine ligase; CFD, C69 family dipeptidase; CrcB, fluoride efflux transporter; Ctp, cobalt ABC transporter, ATP-binding protein; CTR, Crp/Fnr family transcriptional regulator; DMG, DNA-3-methyladenine glycosylase; DUF, DUF1722 domain-containing protein; DUF1310, DUF1310 family protein; DUF1906, DUF1906 domain-containing protein; DUF4432, DUF4432 family protein; EAL, EAL domain-containing protein; EP, extracellular protein; FCA, formate C-acetyltransferase; FCoAT, formyl-CoA transferase; FtsX, FtsX-like permease family protein; GadB/GadA, glutamate decarboxylase; GadC/GadC1/GadC2, glutamate:gamma-aminobutyrate antiporter; GadR, transcriptional regulator; Gcl, glutamate-cysteine ligase; Gls, glutaminase; GltB, glutamate synthase large subunit; GltC, glutamate synthase subunit beta; GltX, glutamate-tRNA ligase; GuaD, guanine deaminase; Hig, HigA family addiction module antidote protein; Hp, hypothetical protein; HsdS, restriction endonuclease subunit S; HTHP, helix-turn-helix domain-containing protein; LacI, LacI family transcriptional regulator; LarB, nickel pincer cofactor biosynthesis protein; LarC, nickel pincer cofactor biosynthesis protein; LarE, ATP-dependent sacrificial sulfur transferase; LuxS, S-ribosylhomocysteine lyase; LysA, diaminopimelate decarboxylase; LysMP, LysM peptidoglycan-binding domain-containing protein; LysR, LysR family transcriptional regulator; LytF, gamma-d-glutamate-meso-diaminopimelate muropeptidase; MarR, MarR family transcriptional regulator; ME, ImmA/IrrE family metallo-endopeptidase; MFS, MFS transporter; Mic, mechanosensitive ion channel; Mp, membrane protein; MTT, class I SAM-dependent methyltransferase; MurI, glutamate racemase; Nat, N-acetyltransferase; NDLR, nickel-dependent lactate racemase; NDM, N-6 DNA methylase; NPCT, Na/Pi cotransporter family protein; Nudix, NUDIX domain-containing protein; Odr, oxidoreductase; Pck, phosphoenolpyruvate carboxykinase; Per, NCS2 family permease; Pmsk, plasmid maintenance system killer; Rb, ribokinase; Rc, ribonuclease HII; RE, restriction endonuclease; RFP, recombinase family protein; RibY, ribonuclease Y; Rp, 30S ribosomal protein S4; RplQ, ribosomal protein L17; RpoA, DNA-directed RNA polymerase, alpha subunit; SDR, SDR family oxidoreductase; SP, sulfate permease; Spx, Spx/MgsR family RNA polymerase-binding regulatory protein; SseA, sulfurtransferase; Tdk, thymidine kinase; Tn, transposase; TP, l,d-transpeptidase family protein; TR, transcriptional regulator; TSE, threonine/serine exporter family protein; YcxB, YcxB family protein.


Figure 3.
Phylogenetic tree (maximum-likelihood method) based on amino acid sequences of GAD from LAB. Figure was generated from MEGA (version X 10.1) after ClustalW alignment of GADs. The GenBank accession number of GAD is indicated in the braces.
Figure 3.
Phylogenetic tree (maximum-likelihood method) based on amino acid sequences of GAD from LAB. Figure was generated from MEGA (version X 10.1) after ClustalW alignment of GADs. The GenBank accession number of GAD is indicated in the braces.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The enzymatic properties of GAD from different LAB.
| Microbe | GAD Length (aa) | Optimal pH | Optimal Temperature (°C) | Predicted Molecular Weight (kDa) | Km (mM) | Vmax | Activators | Reference |
|---|---|---|---|---|---|---|---|---|
| Enterococcus avium M5 * | 467 | 4.5 | 55 | 54.4 | 3.26 ± 0.21 | 0.01 mM/min | Mn2+, Co2+, Ca2+, Zn2+ | [56] |
| Enterococcus raffinosus TCCC11660 | NR | 4.6 | 45 | 55.0 | 5.26 | 3.45 μM/min | Mo6+, Mg2+ | [57] |
| Lactobacillus fermentum * YS2 | 467 | 4.5 | 40 | 54.1 | NR | NR | Ca2+, Mg2+ | [24] |
| Lactobacillus zymae GU240 * | 479 | 4.5 | 41 | 53 | 1.7 | 0.01 mM/min | NR | [58] |
| Lb. brevis 877G * | 468 | 5.2 | 45 | 53 | 3.6 | 0.06 mM/min | Ca2+ | [13] |
| Lb. brevis CGMCC 1306 | 479 | 4.4 | 37 | ~62 | 8.22 | 6.59 U/mg | NR | [59] |
| Lb. brevis CGMCC 1306 * | 479 | 4.8 | 48 | 53.47 | 10.26 | 8.86 U/mg | NR | [60] |
| Lb. brevis HYE1 * | 469 | 4.0 | 55 | 54.32 | 4.99 | 0.224 mM/min | NR | [61] |
| Lb. brevis IFO 12005 | 480 | 4.2 | 30 | 54 | 9.3 | NR | NH4+ | [62,63] |
| Lb. brevis NPS-QW-145 GadA | 468 | 4.8 | 45 | 53.49 | 26.95 ± 2.437 | 9.16 μM/min | NR | [31] |
| Lb. brevis NPS-QW-145 GadB | 479 | 4.8 | 40 | 53.51 | 21.39 ± 1.142 | 32.56 μM/min | NR | [31] |
| Lb. hilgardii MYA-9 | NR | 4.6 | 37 | 60 | NR | NR | Na+, NH4+, Mg2+ | [64] |
| Lb. paracasei NFRI 7415 | 481 | 5.0 | 50 | 54.3 | 5.0 | 7.5 μM/min | Ca2+, NH4+ | [65] |
| Lb. plantarum ATCC 14917 * | 469 | 4.5 | 40 | 53 | 22.8 | 24.4 U/mg | NR | [66] |
| Lb. plantarum WCFS1 | 469 | 4.8 | 60 | 53.75 | 20.02 | 73.33 μM/min | NR | [31] |
| Lb. sakei A156 * | 479 | 5.0 | 55 | 54.4 | 0.045 | 0.011 mM/min | Ca2+, Mn2+, Co2+, Zn2+ | [67] |
| Lb. sakei OPK2-59 | 479 | 5.0 | 30 | 54.4 | NR | NR | Ca2+, Fe3+, Mg2+ | [68] |
| Lc. lactis 01-7 | 466 | 4.7 | NR | 53.93 | 0.51 | NR | NR | [69,70] |
| S. salivarius ssp. thermophilus Y2 | 459 | 4.0 | 40 | 47 | 2.3 | NR | Ba2+ | [28] |
* recombinant protein in E. coli. NR, not reported.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 995. https://doi.org/10.3390/ijms21030995
AMA Style
Cui Y, Miao K, Niyaphorn S, Qu X. Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review. International Journal of Molecular Sciences. 2020; 21(3):995. https://doi.org/10.3390/ijms21030995
Chicago/Turabian StyleCui, Yanhua, Kai Miao, Siripitakyotin Niyaphorn, and Xiaojun Qu. 2020. "Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review" International Journal of Molecular Sciences 21, no. 3: 995. https://doi.org/10.3390/ijms21030995
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.