Dihydrochalcone Derivatives from Populus balsamifera L. Buds for the Treatment of Psoriasis


 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Antiproliferative Potential
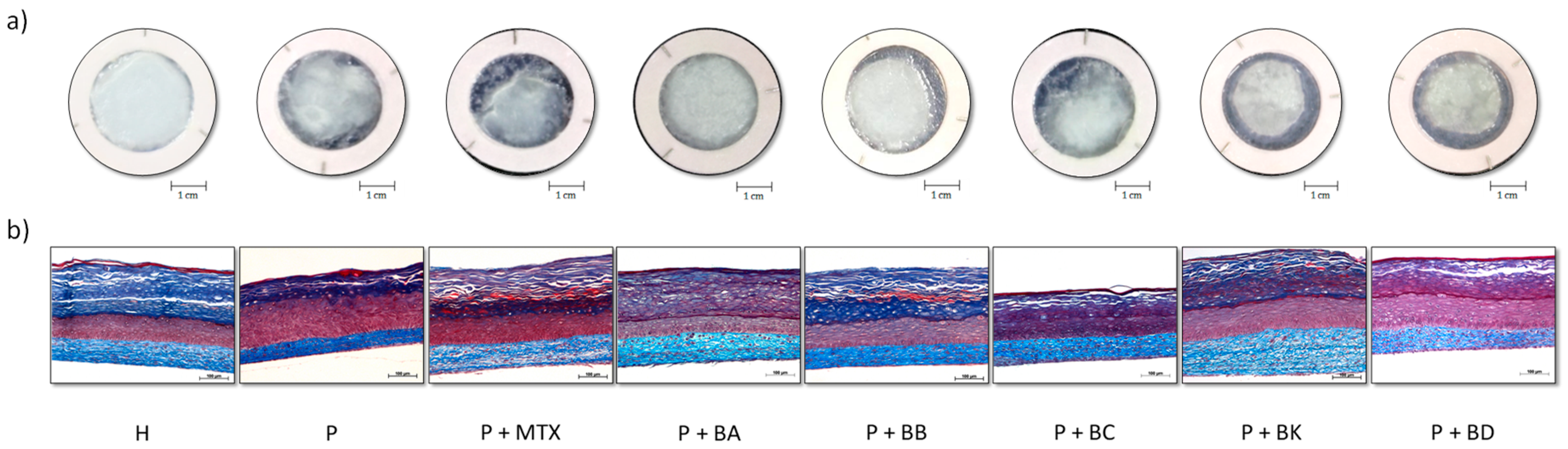
2.2. Macroscopic and Histological Analyses
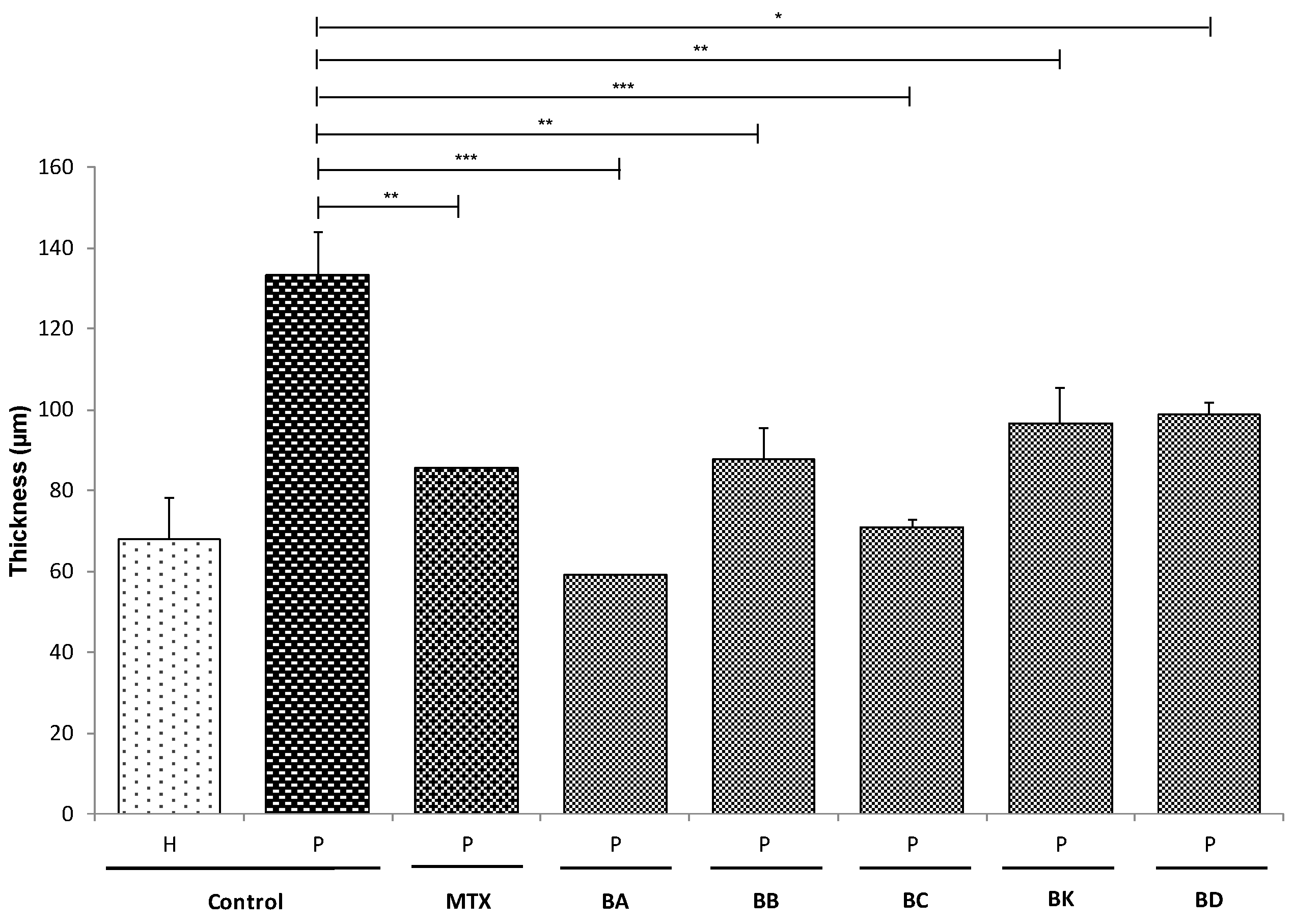
2.3. Living Epidermis Thickness
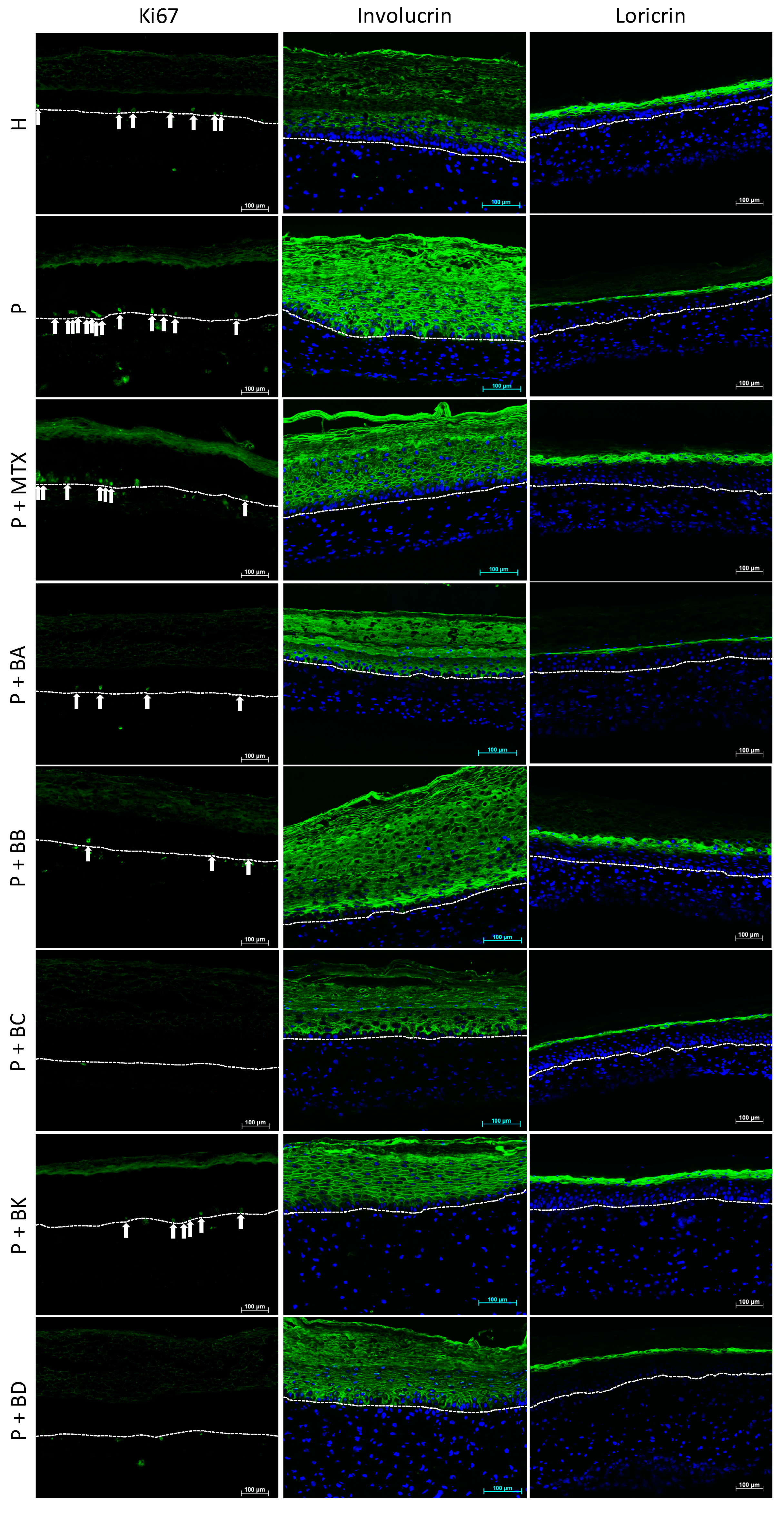
2.4. Immunofluorescence Staining
2.5. Anti-Inflammatory and Antioxidant Properties
3. Discussion
3.1. Histological Analysis: The New Compounds Regulated the Differentiation of Psoriatic Keratinocytes
3.2. Improvement of the Epithelial Phenotype
3.2.1. Prevention of Hyperproliferation of Psoriatic Keratinocytes
3.2.2. Improvement of Barrier Protein Expression
3.3. Diminution of Macrophage-Induced Inflammation and Potential Antioxidant Effect
4. Materials and Methods
4.1. Donors
4.2. Ethical Considerations
4.3. Compound Preparation
4.4. Sulforhodamine B Assay
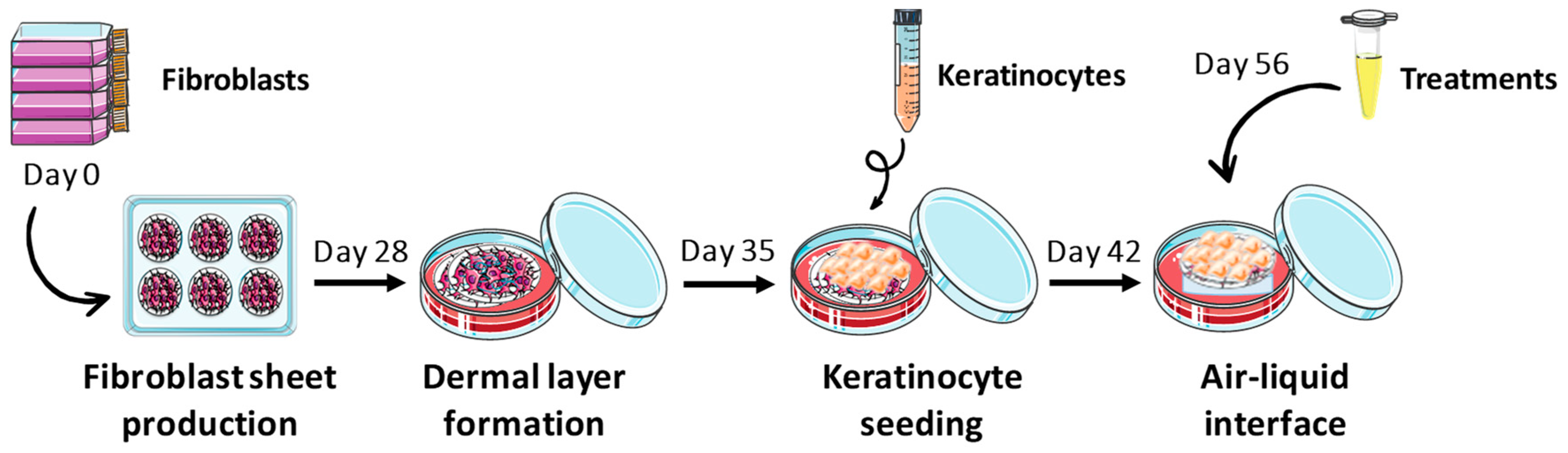
4.5. Tissue-Engineered Skin Production
4.6. Histology and Immunofluorescence Staining
4.7. Measurement of Anti-Inflammatory Activity by Nitrite Quantification
4.8. Evaluation of Antioxidant Activity Using Cell-Based Assay
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kalinin, A.; Marekov, L.N.; Steinert, P.M. Assembly of the epidermal cornified cell envelope. J. Cell Sci. 2001, 114, 3069–3070. [Google Scholar]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex: Cornified envelope precursors, S100 proteins and the ‘fused genes’ family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef]
- Raychaudhuri, S.; Farber, E. The prevalence of psoriasis in the world. J. Eur. Acad. Dermatol. Venereol. 2001, 15, 16–17. [Google Scholar] [CrossRef]
- Camisa, C. Pathogenesis of Psoriasis. In Handbook of Psoriasis; Blackwell Publishing: Oxford, UK, 2004; pp. 45–60. [Google Scholar]
- Kerkhof, P.C.M.v.d. Textbook of Psoriasis; Blackwell: Malden, MA, USA, 2003. [Google Scholar]
- Eroschenko, V.P. DiFiore’s Atlas of Histology with Functional Correlations, 11th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008. [Google Scholar]
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef]
- Gaspari, A.A. Innate and adaptive immunity and the pathophysiology of psoriasis. J. Am. Acad. Dermatol. 2006, 54 (Suppl. S2), S67–S80. [Google Scholar] [CrossRef]
- Lowes, M.A.; Bowcock, A.M.; Krueger, J.G. Pathogenesis and therapy of psoriasis. Nature 2007, 445, 866–873. [Google Scholar] [CrossRef]
- Balato, A.; Balato, N.; Megna, M.; Schiattarella, M.; Lembo, S.; Ayala, F. Pathogenesis of Psoriasis: The Role of Pro-Inflammatory Cytokines Produced by Keratinocytes. In Psoriasis; Soung, J., Ed.; IntechOpen: London, UK, 2012. [Google Scholar]
- Hwang, S.M.; Ahn, S.K.; Menon, G.K.; Choi, E.H.; Lee, S.H. Basis of occlusive therapy in psoriasis: Correcting defects in permeability barrier and calcium gradient. Int. J. Dermatol. 2001, 40, 223–231. [Google Scholar] [CrossRef]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef]
- Steven, A.C.; Bisher, M.E.; Roop, D.R.; Steinert, P.M. Biosynthetic pathways of filaggrin and loricrin—Two major proteins expressed by terminally differentiated epidermal keratinocytes. J. Struct. Biol. 1990, 104, 150–162. [Google Scholar] [CrossRef]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H.I. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Ishida-Yamamoto, A.; Takahashi, H.; Iizuka, H. Immunoelectron microscopy links molecules and morphology in the studies of keratinization. Eur. J. Dermatol. 2000, 10, 429–435. [Google Scholar]
- Ekanayake-Mudiyanselage, S.; Jensen, J.-M.; Proksch, E.; Aschauer, H.; Schmook, F.P.; Meingassner, J.G. Expression of Epidermal Keratins and the Cornified Envelope Protein Involucrin is Influenced by Permeability Barrier Disruption. J. Investig. Dermatol. 1998, 111, 517–523. [Google Scholar] [CrossRef] [Green Version]
- Ishida-Yamamoto, A.; Iizuka, H. Differences in Involucrin Immunolabeling within Cornified Cell Envelopes in Normal and Psoriatic Epidermis. J. Investig. Dermatol. 1995, 104, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-Q.; Man, X.-Y.; Li, W.; Zhou, J.; Landeck, L.; Cai, S.-Q.; Zheng, M. Regulation of Involucrin in Psoriatic Epidermal Keratinocytes: The Roles of ERK1/2 and GSK-3β. Cell Biochem. Biophys. 2013, 66, 523–528. [Google Scholar] [CrossRef]
- García-Pérez, M.-E.; Jean, J.; Pouliot, R. Antipsoriatic Drug Development: Challenges and New Emerging Therapies. Recent Pat. Inflamm. Allergy Drug Discov. 2012, 6, 3–21. [Google Scholar] [CrossRef]
- Kormeili, T.; Lowe, N.J.; Yamauchi, P.S. Psoriasis: Immunopathogenesis and evolving immunomodulators and systemic therapies; U.S. experiences. Br. J. Dermatol. 2004, 151, 3–15. [Google Scholar] [CrossRef]
- Raskin, I.; Ribnicky, D.M.; Komarnytsky, S.; Ilic, N.; Poulev, A.; Borisjuk, N.; Brinker, A.; Moreno, D.A.; Ripoll, C.; Yakoby, N.; et al. Plants and human health in the twenty-first century. Trends Biotechnol. 2002, 20, 522–531. [Google Scholar] [CrossRef]
- Harvey, A.L. Natural products in drug discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef]
- Bonesi, M.; Loizzo, M.R.; Menichini, F.; Tundis, R. Chapter 23—Flavonoids in Treating Psoriasis. In Immunity and Inflammation in Health and Disease; Chatterjee, S., Jungraithmayr, W., Bagchi, D., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 281–294. [Google Scholar]
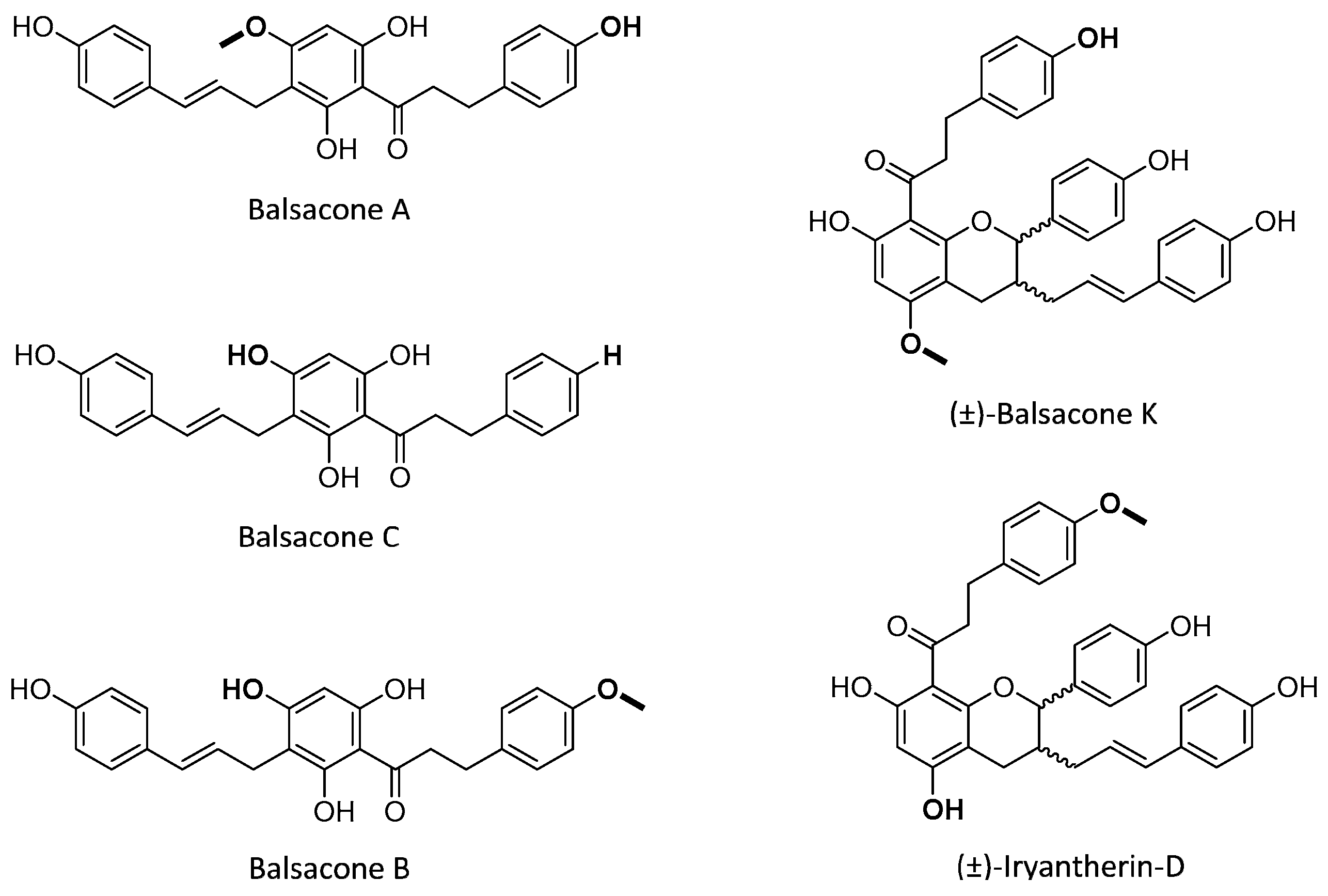
- Lavoie, S.; Legault, J.; Simard, F.; Chiasson, E.; Pichette, A. New antibacterial dihydrochalcone derivatives from buds of Populus balsamifera. Tetrahedron Lett. 2013, 54, 1631–1633. [Google Scholar] [CrossRef]
- Simard, F.; Legault, J.; Lavoie, S.; Pichette, A. Balsacones D-I, dihydrocinnamoyl flavans from Populus balsamifera buds. Phytochemistry 2014, 100, 141–149. [Google Scholar] [CrossRef]
- Simard, F.; Gauthier, C.; Chiasson, E.; Lavoie, S.; Mshvildadze, V.; Legault, J.; Pichette, A. Antibacterial Balsacones J–M, Hydroxycinnamoylated Dihydrochalcones from Populus balsamifera Buds. J. Nat. Prod. 2015, 78, 1147–1153. [Google Scholar] [CrossRef]
- Uprety, Y.; Asselin, H.; Dhakal, A.; Julien, N. Traditional use of medicinal plants in the boreal forest of Canada: Review and perspectives. J. Ethnobiol. Ethnomed. 2012, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marles, R.J. Aboriginal Plant Use in Canada’s Northwest Boreal Forest; UBC Press: Vancouver, BC, Canada, 2000. [Google Scholar]
- Simard, F.; Gauthier, C.; Legault, J.; Lavoie, S.; Mshvildadze, V.; Pichette, A. Structure elucidation of anti-methicillin resistant Staphylococcus aureus (MRSA) flavonoids from balsam poplar buds. Bioorg. Med. Chem. 2016, 24, 4188–4198. [Google Scholar] [CrossRef] [PubMed]
- Côté, H.; Pichette, A.; Simard, F.; Ouellette, M.-E.; Ripoll, L.; Mihoub, M.; Grimard, D.; Legault, J. Balsacone C, a New Antibiotic Targeting Bacterial Cell Membranes, Inhibits Clinical Isolates of Methicillin-Resistant Staphylococcus aureus (MRSA) Without Inducing Resistance. Front. Microbiol. 2019, 10, 2341. [Google Scholar] [CrossRef] [Green Version]
- Jean, J.; Lapointe, M.; Soucy, J.; Pouliot, R. Development of an in vitro psoriatic skin model by tissue engineering. J. Dermatol. Sci. 2009, 53, 19–25. [Google Scholar] [CrossRef]
- Jean, J.; Soucy, J.; Pouliot, R. Effects of Retinoic Acid on Keratinocyte Proliferation and Differentiation in a Psoriatic Skin Model. Tissue Eng. Part A 2011, 17, 1859–1868. [Google Scholar] [CrossRef]
- Jean, J.; Leroy, M.; Duque-Fernandez, A.; Bernard, G.; Soucy, J.; Pouliot, R. Characterization of a psoriatic skin model produced with involved or uninvolved cells. J. Tissue Eng. Regen. Med. 2015, 9, 789–798. [Google Scholar] [CrossRef]
- Grenier, A.; Gendreau, I.; Pouliot, R. Optimization of the Self-Assembly Method for the Production of Psoriatic Skin Substitutes. In Cell Culture; Mehanna, R.A., Ed.; IntechOpen: London, UK, 2018. [Google Scholar]
- Carrascosa, J.M.; de la Cueva, P.; Ara, M.; Puig, L.; Bordas, X.; Carretero, G.; Ferrándiz, L.; Sánchez-Carazo, J.L.; Daudén, E.; López-Estebaranz, K.L.; et al. Methotrexate in Moderate to Severe Psoriasis: Review of the Literature and Expert Recommendations. Actas Dermo Sifiliogr. (Engl. Ed.) 2016, 107, 194–206. [Google Scholar] [CrossRef]
- Huang, T.-H.; Lin, C.-F.; Alalaiwe, A.; Yang, S.-C.; Fang, J.-Y. Apoptotic or Antiproliferative Activity of Natural Products against Keratinocytes for the Treatment of Psoriasis. Int. J. Mol. Sci. 2019, 20, 2558. [Google Scholar] [CrossRef] [Green Version]
- Ando, M.; Kawashima, T.; Kobayashi, H.; Ohkawara, A. Immunohistological detection of proliferating cells in normal and psoriatic epidermis using Ki-67 monoclonal antibody. J. Dermatol. Sci. 1990, 1, 441–446. [Google Scholar] [CrossRef]
- Sirsjö, A.; Karlsson, M.; Gidöf, A.; Rollman, O.; Törmä, H. Increased expression of inducible nitric oxide synthase in psoriatic skin and cytokine-stimulated cultured keratinocytes. Br. J. Dermatol. 1996, 134, 643–648. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, A.D.; Weller, R.; Copeland, P.; Benjamin, N.; Ralston, S.H.; Grabowski, P.; Herriot, R. Detection of nitric oxide and nitric oxide synthases in psoriasis. Arch. Dermatol. Res. 1998, 290, 3–8. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, M.-E.; Allaeys, I.; Rusu, D.; Pouliot, R.; Janezic, T.S.; Poubelle, P.E. Picea mariana polyphenolic extract inhibits phlogogenic mediators produced by TNF-α-activated psoriatic keratinocytes: Impact on NF-κB pathway. J. Ethnopharmacol. 2014, 151, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Tekin, N.S.; Ilter, N.; Sancak, B.; Ozden, M.G.; Gurer, M.A. Nitric oxide levels in patients with psoriasis treated with methotrexate. Mediat. Inflamm. 2006, 2006, 16043. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Fontánez, N.; Soler, D.C.; McCormick, T.S. Current knowledge on psoriasis and autoimmune diseases. Psoriasis (Auckl. N.Z.) 2016, 6, 7–32. [Google Scholar]
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Huang, T. Oxidative stress in psoriasis and potential therapeutic use of antioxidants. Free Radic. Res. 2016, 50, 585–595. [Google Scholar] [CrossRef]
- Bickers, D.R.; Athar, M. Oxidative Stress in the Pathogenesis of Skin Disease. J. Investig. Dermatol. 2006, 126, 2565–2575. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Mrowietz, U.; Rostami-Yazdi, M. Oxidative stress in the pathogenesis of psoriasis. Free Radic. Biol. Med. 2009, 47, 891–905. [Google Scholar] [CrossRef]
- Lei, Y.; Wang, K.; Deng, L.; Chen, Y.; Nice, E.C.; Huang, C. Redox Regulation of Inflammation: Old Elements, a New Story. Med. Res. Rev. 2015, 35, 306–340. [Google Scholar] [CrossRef]
- Ferretti, G.; Bacchetti, T.; Campanati, A.; Simonetti, O.; Liberati, G.; Offidani, A. Correlation between lipoprotein(a) and lipid peroxidation in psoriasis: Role of the enzyme paraoxonase-1. Br. J. Dermatol. 2012, 166, 204–207. [Google Scholar] [CrossRef] [PubMed]
- Gabr, S.A.; AAl-Ghadir, H. Role of cellular oxidative stress and cytochrome c in the pathogenesis of psoriasis. Arch. Dermatol. Res. 2012, 304, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Şikar Aktürk, A.; Özdoğan, H.K.; Bayramgürler, D.; Çekmen, M.B.; Bilen, N.; Kıran, R. Nitric oxide and malondialdehyde levels in plasma and tissue of psoriasis patients. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Kadam, D.P.; Suryakar, A.N.; Ankush, R.D.; Kadam, C.Y.; Deshpande, K.H. Role of Oxidative Stress in Various Stages of Psoriasis. Indian J. Clin. Biochem. 2010, 25, 388–392. [Google Scholar] [CrossRef] [Green Version]
- Lorthois, I.; Simard, M.; Morin, S.; Pouliot, R. Infiltration of T Cells into a Three-Dimensional Psoriatic Skin Model Mimics Pathological Key Features. Int. J. Mol. Sci. 2019, 20, 1670. [Google Scholar] [CrossRef] [Green Version]
- Germain, L.; Rouabhia, M.; Guignard, R.; Carrier, L.; Bouvard, V.; Auger, F.A. Improvement of human keratinocyte isolation and culture using thermolysin. Burns 1993, 19, 99–104. [Google Scholar] [CrossRef]
- Auger, F.A.; Valle, C.A.L.; Guignard, R.; Tremblay, N.; Noel, B.; Goulet, F.; Germain, L. Skin equivalent produced with human collagen. In Vitro Cell. Dev. Biol. Anim. 1995, 31, 432–439. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Legault, J.; Girard-Lalancette, K.; Grenon, C.; Dussault, C.; Pichette, A. Antioxidant Activity, Inhibition of Nitric Oxide Overproduction, and In Vitro Antiproliferative Effect of Maple Sap and Syrup from Acer saccharum. J. Med. Food 2010, 13, 460–468. [Google Scholar] [CrossRef]
- Girard-Lalancette, K.; Pichette, A.; Legault, J. Sensitive cell-based assay using DCFH oxidation for the determination of pro- and antioxidant properties of compounds and mixtures: Analysis of fruit and vegetable juices. Food Chem. 2009, 115, 720–726. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | IC50 (μM) |
|---|---|
| Balsacone A | 48 |
| Balsacone B | 76 |
| Balsacone C | 128 |
| (±)-Balsacone K | 45 |
| (±)-Iryantherin-D | 23 |
| Molecule | NO Inhibition IC50 (μM) 1 | Antioxidant IC50 (μM) 2 |
|---|---|---|
| Balsacone A | 17 ± 1 µM | 3.1 ± 0.5 µM |
| Balsacone B | 13.3 ± 0.8 µM | 1.05 ± 0.09 µM |
| Balsacone C | 29 ± 1 µM | 1.10 ± 0.08 µM |
| (±)-Balsacone K | 10.49 ± 0.07 µM | 1.8 ± 0.3 µM |
| (±)-Iryantherin-D | 26.9 ± 0.5 µM | 5.1 ± 0.8 µM |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bélanger, A.; Grenier, A.; Simard, F.; Gendreau, I.; Pichette, A.; Legault, J.; Pouliot, R. Dihydrochalcone Derivatives from Populus balsamifera L. Buds for the Treatment of Psoriasis. Int. J. Mol. Sci. 2020, 21, 256. https://doi.org/10.3390/ijms21010256
Bélanger A, Grenier A, Simard F, Gendreau I, Pichette A, Legault J, Pouliot R. Dihydrochalcone Derivatives from Populus balsamifera L. Buds for the Treatment of Psoriasis. International Journal of Molecular Sciences. 2020; 21(1):256. https://doi.org/10.3390/ijms21010256
Chicago/Turabian StyleBélanger, Audrey, Alexe Grenier, François Simard, Isabelle Gendreau, André Pichette, Jean Legault, and Roxane Pouliot. 2020. "Dihydrochalcone Derivatives from Populus balsamifera L. Buds for the Treatment of Psoriasis" International Journal of Molecular Sciences 21, no. 1: 256. https://doi.org/10.3390/ijms21010256