Composing the Early Embryonic Microenvironment: Physiology and Regulation of Oviductal Secretions

 , ,
, ,
Abstract
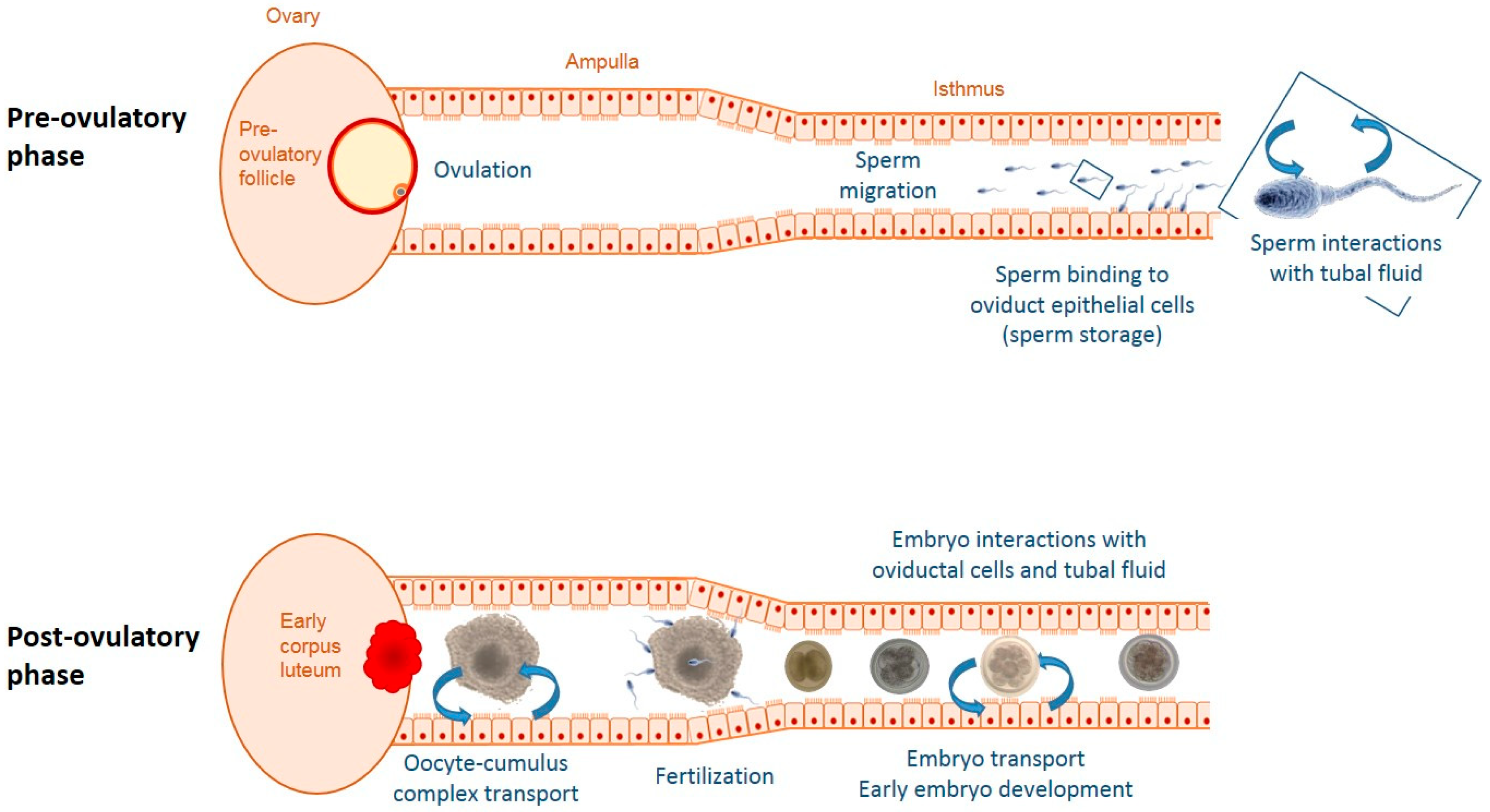
:1. Introduction
2. Origin and Renewal of the Tubal Fluid
2.1. Potential Participation of the Follicular Fluid in the Composition of the Tubal Fluid
2.2. Production Rate and Renewal of Oviductal Secretions
3. Molecular Components of the Tubal Fluid
3.1. Small Metabolites and Carbohydrates
3.2. Lipids
3.3. Proteins
3.4. Hormones
4. Factors and Processes Regulating Oviductal Secretions
4.1. Ovulation
4.2. Insemination and Gametes
4.3. Pregnancy and Embryo
4.4. Sex Steroid Hormones
4.5. Treatments of Estrus Synchronization and Superovulation
4.6. Metabolism and Energy Balance
4.7. Heat Stress
5. Concluding Remarks and Perspectives
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AGTR2 | Angiotensin II receptor type 2 |
| COC | Cumulus–oocyte complex |
| DAPI | 4′,6-diamidino-2-phénylindole |
| DEGs | Differentially expressed genes |
| eCG | Equine chorionic gonadotropin |
| EP2,4 | Prostaglandin E2 receptor 2,4 |
| ESR1 | Estrogen receptor 1 |
| EVs | Extracellular vesicles |
| FF | Follicular fluid |
| FLT1 | Vascular endothelial growth factor receptor 1 |
| FSH | Follicle stimulating hormone |
| hCG | human chorionic gonadotropin |
| HSP | Heat shock protein |
| IVF | In vitro fertilization |
| LHCG | Luteinizing hormone chorionic gonadotropin |
| OEC | Oviductal epithelial cell |
| OF | Oviductal fluid |
| OVGP1 | Oviduct-specific glycoprotein 1 |
| PG | Prostaglandins |
| PGR | Progesterone receptor |
| SOF | Synthetic oviductal fluid |
| TICAM2 | Toll-like receptor adaptor molecule 2 |
| VEGF | Vascular endothelial growth factor |
References
- Rizos, D.; Clemente, M.; Bermejo-Alvarez, P.; de La Fuente, J.; Lonergan, P.; Gutierrez-Adan, A. Consequences of in vitro culture conditions on embryo development and quality. Reprod. Domest. Anim. 2008, 43, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Rizos, D.; Ward, F.; Duffy, P.; Boland, M.P.; Lonergan, P. Consequences of bovine oocyte maturation, fertilization or early embryo development in vitro versus in vivo: Implications for blastocyst yield and blastocyst quality. Mol. Reprod. Dev. 2002, 61, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Sudano, M.J.; Santos, V.G.; Tata, A.; Ferreira, C.R.; Paschoal, D.M.; Machado, R.; Buratini, J.; Eberlin, M.N.; Landim-Alvarenga, F.D. Phosphatidylcholine and sphingomyelin profiles vary in Bos taurus indicus and Bos taurus taurus in vitro- and in vivo-produced blastocysts. Biol. Reprod. 2012, 87, 130. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, P. State-of-the-art embryo technologies in cattle. Soc. Reprod. Fertil. Suppl. 2007, 64, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Maillo, V.; Lopera-Vasquez, R.; Hamdi, M.; Gutierrez-Adan, A.; Lonergan, P.; Rizos, D. Maternal-embryo interaction in the bovine oviduct: Evidence from in vivo and in vitro studies. Theriogenology 2016, 86, 443–450. [Google Scholar] [CrossRef]
- Maillo, V.; Sanchez-Calabuig, M.J.; Lopera-Vasquez, R.; Hamdi, M.; Gutierrez-Adan, A.; Lonergan, P.; Rizos, D. Oviductal response to gametes and early embryos in mammals. Reproduction 2016, 152, R127–R141. [Google Scholar] [CrossRef] [Green Version]
- Alminana, C.; Bauersachs, S. Extracellular Vesicles in the Oviduct: Progress, Challenges and Implications for the Reproductive Success. Bioengineering 2019, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Leese, H.J.; Tay, J.I.; Reischl, J.; Downing, S.J. Formation of Fallopian tubal fluid: Role of a neglected epithelium. Reproduction 2001, 121, 339–346. [Google Scholar] [CrossRef]
- Brussow, K.P.; Ratky, J.; Rodriguez-Martinez, H. Fertilization and early embryonic development in the porcine fallopian tube. Reprod. Domest. Anim. 2008, 43, 245–251. [Google Scholar] [CrossRef]
- Brussow, K.-P.; Ratky, J.; Schneider, F.; Torner, H.; Kanitz, W.; Solti, L. Effects of follicular fluid on the transport of porcine oocytes into the oviduct at ovulation. Reprod. Domest. Anim. 1999, 34, 423–429. [Google Scholar] [CrossRef]
- Hansen, C.; Srikandakumar, A.; Downey, B.R. Presence of follicular fluid in the porcine oviduct and its contribution to the acrosome reaction. Mol. Reprod. Dev. 1991, 30, 148–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolle, S.; Dubielzig, S.; Reese, S.; Wehrend, A.; Konig, P.; Kummer, W. Ciliary transport, gamete interaction, and effects of the early embryo in the oviduct: Ex vivo analyses using a new digital videomicroscopic system in the cow. Biol. Reprod. 2009, 81, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabler, C.; Odau, S.; Muller, K.; Schon, J.; Bondzio, A.; Einspanier, R. Exploring Cumulus-Oocyte-Complex-Oviductal Cell Interactions: Gene Profiling Inthe Bovine Oviduct. J. Physiol. Pharmacol. 2008, 59, 29–42. [Google Scholar] [PubMed]
- da Silveira, J.C.; Andrade, G.M.; Del Collado, M.; Sampaio, R.V.; Sangalli, J.R.; Silva, L.A.; Pinaffi, F.V.L.; Jardim, I.B.; Cesar, M.C.; Nogueira, M.F.G.; et al. Supplementation with small-extracellular vesicles from ovarian follicular fluid during in vitro production modulates bovine embryo development. PLoS ONE 2017, 12, e0179451. [Google Scholar] [CrossRef] [PubMed]
- Roberts, G.P.; Parker, J.M.; Symonds, H.W. Proteins in the luminal fluid from the bovine oviduct. J. Reprod. Fertil. 1975, 45, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Engle, C.C.; Witherspoon, D.M.; Foley, C.W. Technique for continuous collection of equine oviduct secretions. Am. J. Vet. Res. 1970, 31, 1889–1896. [Google Scholar]
- Leese, H.J.; Hugentobler, S.A.; Gray, S.M.; Morris, D.G.; Sturmey, R.G.; Whitear, S.L.; Sreenan, J.M. Female reproductive tract fluids: Composition, mechanism of formation and potential role in the developmental origins of health and disease. Reprod. Fertil. Dev. 2008, 20, 1–8. [Google Scholar] [CrossRef]
- Hugentobler, S.A.; Sreenan, J.M.; Humpherson, P.G.; Leese, H.J.; Diskin, M.G.; Morris, D.G. Effects of changes in the concentration of systemic progesterone on ions, amino acids and energy substrates in cattle oviduct and uterine fluid and blood. Reprod. Fertil. Dev. 2010, 22, 684–694. [Google Scholar] [CrossRef]
- Ferraz, M.; Rho, H.S.; Hemerich, D.; Henning, H.H.W.; van Tol, H.T.A.; Holker, M.; Besenfelder, U.; Mokry, M.; Vos, P.; Stout, T.A.E.; et al. An oviduct-on-a-chip provides an enhanced in vitro environment for zygote genome reprogramming. Nat. Commun. 2018, 9, 4934. [Google Scholar] [CrossRef] [Green Version]
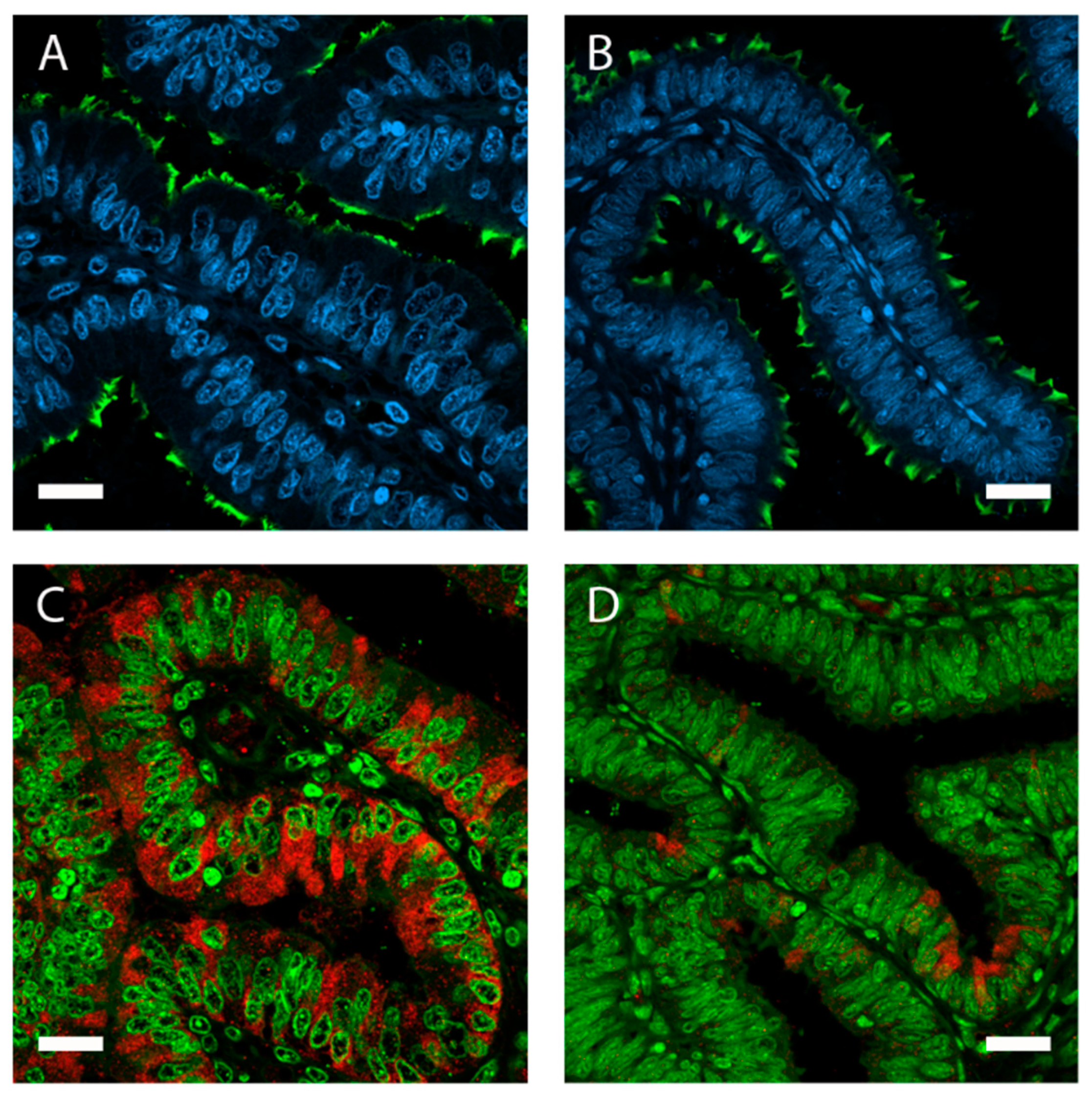
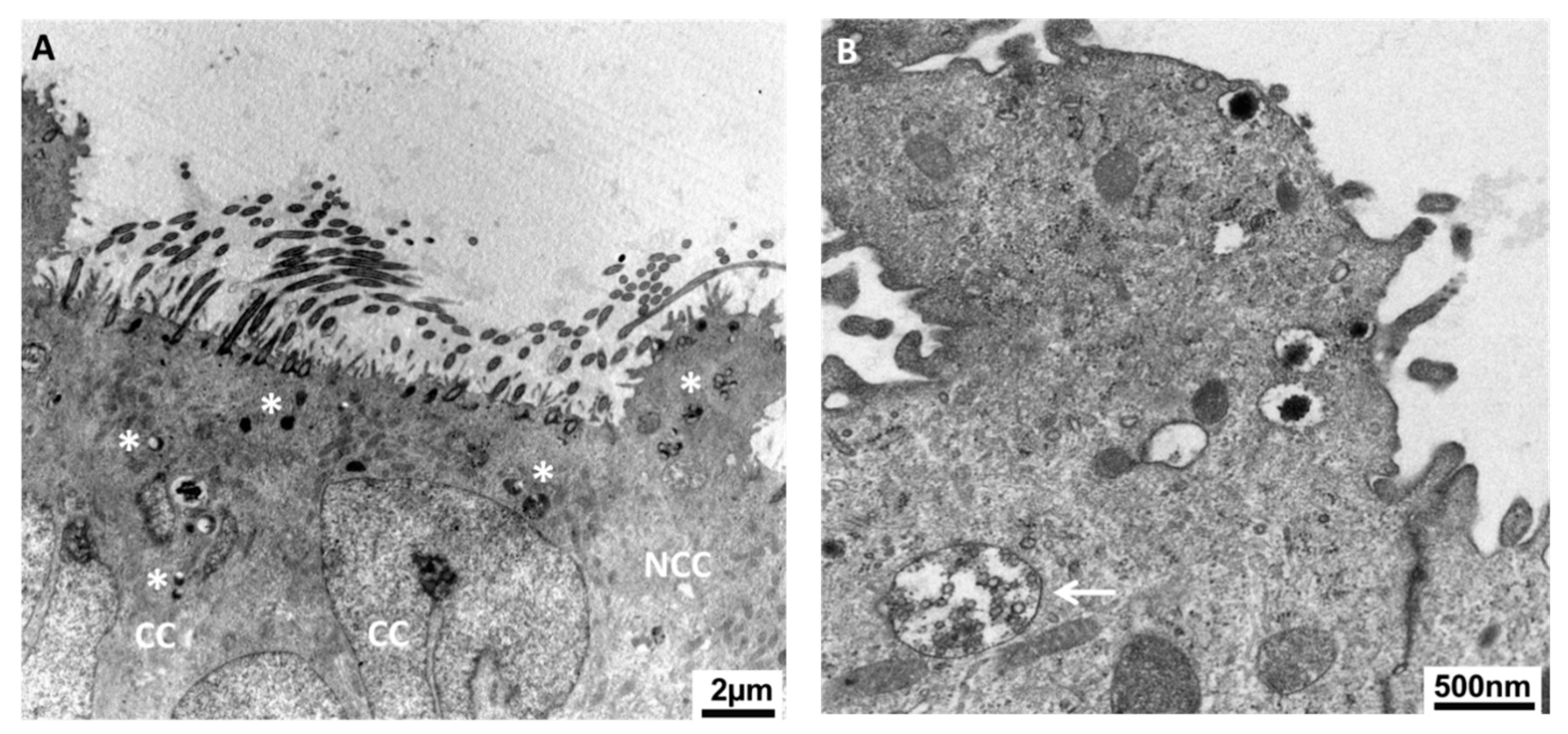
- Ito, S.; Kobayashi, Y.; Yamamoto, Y.; Kimura, K.; Okuda, K. Remodeling of bovine oviductal epithelium by mitosis of secretory cells. Cell Tissue Res. 2016, 366, 403–410. [Google Scholar] [CrossRef]
- Steinhauer, N.; Boos, A.; Gunzel-Apel, A.R. Morphological changes and proliferative activity in the oviductal epithelium during hormonally defined stages of the oestrous cycle in the bitch. Reprod. Domest. Anim. 2004, 39, 110–119. [Google Scholar] [CrossRef] [PubMed]
- George, S.H.; Milea, A.; Shaw, P.A. Proliferation in the normal FTE is a hallmark of the follicular phase, not BRCA mutation status. Clin. Cancer Res. 2012, 18, 6199–6207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Syed, S.M.; Tanwar, P.S. In vivo genetic cell lineage tracing reveals that oviductal secretory cells self-renew and give rise to ciliated cells. Development 2017, 144, 3031–3041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugentobler, S.A.; Diskin, M.G.; Leese, H.J.; Humpherson, P.G.; Watson, T.; Sreenan, J.M.; Morris, D.G. Amino acids in oviduct and uterine fluid and blood plasma during the estrous cycle in the bovine. Mol. Reprod. Dev. 2007, 74, 445–454. [Google Scholar] [CrossRef]
- Menezo, Y.; Guerin, P.; Elder, K. The oviduct: A neglected organ due for re-assessment in IVF. Reprod. Biomed. Online 2015, 30, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Lamy, J.; Gatien, J.; Dubuisson, F.; Nadal-Desbarats, L.; Salvetti, P.; Mermillod, P.; Saint-Dizier, M. Metabolomic profiling of bovine oviductal fluid across the oestrous cycle using proton nuclear magnetic resonance spectroscopy. Reprod. Fertil. Dev. 2018, 30, 1021–1028. [Google Scholar] [CrossRef]
- Li, S.; Winuthayanon, W. Oviduct: Roles in fertilization and early embryo development. J. Endocrinol. 2017, 232, R1–R26. [Google Scholar] [CrossRef] [Green Version]
- Hugentobler, S.A.; Humpherson, P.G.; Leese, H.J.; Sreenan, J.M.; Morris, D.G. Energy substrates in bovine oviduct and uterine fluid and blood plasma during the oestrous cycle. Mol. Reprod. Dev. 2008, 75, 496–503. [Google Scholar] [CrossRef]
- Tervit, H.R.; Whittingham, D.G.; Rowson, L.E. Successful culture in vitro of sheep and cattle ova. J. Reprod. Fertil. 1972, 30, 493–497. [Google Scholar] [CrossRef]
- Jordaens, L.; Van Hoeck, V.; De Bie, J.; Berth, M.; Marei, W.F.A.; Desmet, K.L.J.; Bols, P.E.J.; Leroy, J. Non-esterified fatty acids in early luteal bovine oviduct fluid mirror plasma concentrations: An ex vivo approach. Reprod. Biol. 2017, 17, 281–284. [Google Scholar] [CrossRef]
- Banliat, C.; Tomas, D.; Teixeira-Gomes, A.P.; Uzbekova, S.; Guyonnet, B.; Labas, V.; Saint-Dizier, M. Stage-dependent changes in oviductal phospholipid profiles throughout the estrous cycle in cattle. Theriogenology 2019, 135, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Belaz, K.R.; Tata, A.; Franca, M.R.; Santos da Silva, M.I.; Vendramini, P.H.; Fernandes, A.M.; D’Alexandri, F.L.; Eberlin, M.N.; Binelli, M. Phospholipid Profile and Distribution in the Receptive Oviduct and Uterus During Early Diestrus in Cattle. Biol. Reprod. 2016, 95, 127. [Google Scholar] [CrossRef] [PubMed]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal lipid composition and the role of ether lipids and phosphoinositides in exosome biology. J. Lipid Res. 2019, 60, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banliat, C.; Dubuisson, F.; Corbin, E.; Beurois, J.; Tomas, D.; Le Bourhis, D.; Salvetti, P.; Labas, V.; Mermillod, P.; Saint-Dizier, M. Intraoviductal concentrations of steroid hormones during in vitro culture changed phospholipid profiles and cryotolerance of bovine embryos. Mol. Reprod. Dev. 2019, 86, 661–672. [Google Scholar] [CrossRef]
- Georgiou, A.S.; Snijders, A.P.; Sostaric, E.; Aflatoonian, R.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; Martinez, E.A.; Wright, P.C.; Fazeli, A. Modulation of the oviductal environment by gametes. J. Proteome Res. 2007, 6, 4656–4666. [Google Scholar] [CrossRef]
- Georgiou, A.S.; Sostaric, E.; Wong, C.H.; Snijders, A.P.; Wright, P.C.; Moore, H.D.; Fazeli, A. Gametes alter the oviductal secretory proteome. Mol. Cell. Proteom. 2005, 4, 1785–1796. [Google Scholar] [CrossRef] [Green Version]
- Reynaud, K.; Saint-Dizier, M.; Tahir, M.Z.; Havard, T.; Harichaux, G.; Labas, V.; Thoumire, S.; Fontbonne, A.; Grimard, B.; Chastant-Maillard, S. Progesterone plays a critical role in canine oocyte maturation and fertilization. Biol. Reprod. 2015, 93, 87. [Google Scholar] [CrossRef] [Green Version]
- Soleilhavoup, C.; Riou, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Kohnke, P.; Reynaud, K.; de Graaf, S.P.; Gerard, N.; Druart, X. Proteomes of the female genital tract during the oestrous cycle. Mol. Cell. Proteom. 2016, 15, 93–108. [Google Scholar] [CrossRef] [Green Version]
- Lamy, J.; Labas, V.; Harichaux, G.; Tsikis, G.; Mermillod, P.; Saint-Dizier, M. Regulation of the bovine oviductal fluid proteome. Reproduction 2016, 152, 629–644. [Google Scholar] [CrossRef] [Green Version]
- Papp, S.M.; Frohlich, T.; Radefeld, K.; Havlicek, V.; Kosters, M.; Yu, H.; Mayrhofer, C.; Brem, G.; Arnold, G.J.; Besenfelder, U. A novel approach to study the bovine oviductal fluid proteome using transvaginal endoscopy. Theriogenology 2019, 132, 53–61. [Google Scholar] [CrossRef]
- Pillai, V.V.; Weber, D.M.; Phinney, B.S.; Selvaraj, V. Profiling of proteins secreted in the bovine oviduct reveals diverse functions of this luminal microenvironment. PLoS ONE 2017, 12, e0188105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, K.; Nelis, H.; Van Steendam, K.; Govaere, J.; Roels, K.; Ververs, C.; Leemans, B.; Wydooghe, E.; Deforce, D.; Van Soom, A. Proteome of equine oviducal fluid: Effects of ovulation and pregnancy. Reprod. Fertil. Dev. 2016, 29, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Hackenbroch, L.; Meyer, F.R.L.; Reiser, J.; Razzazi-Fazeli, E.; Nobauer, K.; Besenfelder, U.; Vogl, C.; Brem, G.; Mayrhofer, C. Identification of Rabbit Oviductal Fluid Proteins Involved in Pre-Fertilization Processes by Quantitative Proteomics. Proteomics 2019, 19, e1800319. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, Y.; Chang, C.; Wu, S.; Gao, J.; Zhang, Y.; Chen, Y.; Zhong, F.; Deng, G. Human fallopian tube proteome shows high coverage of mesenchymal stem cells associated proteins. Biosci. Rep. 2016, 36, e00297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferraz, M.; Carothers, A.; Dahal, R.; Noonan, M.J.; Songsasen, N. Oviductal extracellular vesicles interact with the spermatozoon’s head and mid-piece and improves its motility and fertilizing ability in the domestic cat. Sci. Rep. 2019, 9, 9484. [Google Scholar] [CrossRef] [PubMed]
- Lamy, J.; Nogues, P.; Combes-Soia, L.; Tsikis, G.; Labas, V.; Mermillod, P.; Druart, X.; Saint-Dizier, M. Identification by proteomics of oviductal sperm-interacting proteins. Reproduction 2018, 155, 457–466. [Google Scholar] [CrossRef]
- Ghersevich, S.; Massa, E.; Zumoffen, C. Oviductal secretion and gamete interaction. Reproduction 2015, 149, R1–R14. [Google Scholar] [CrossRef]
- Goncalves, J.S.; Vannucchi, C.I.; Braga, F.C.; Paula-Lopes, F.F.; Milazzotto, M.P.; Assumpcao, M.E.; Visintin, J.A. Oestrogen and progesterone receptor gene expression in canine oocytes and cumulus cells throughout the oestrous cycle. Reprod. Domest. Anim. 2009, 44, 239–242. [Google Scholar] [CrossRef]
- Banliat, C.; Tsikis, G.; Teixeira-Gomes, A.P.; Labas, V.; Com, E.; Pineau, C.; Mermillod, P.; Guyonnet, B.; Saint-Dizier, M. A proteomic approach to decipher embryo-maternal interactions in the oviduct. In Proceedings of the 35th Annual Meeting of the European Embryo Transfer Association (AETE), Murcia, Spain, 13–14 September 2019; p. 721. [Google Scholar]
- Alminana, C.; Corbin, E.; Tsikis, G.; Alcantara-Neto, A.S.; Labas, V.; Reynaud, K.; Galio, L.; Uzbekov, R.; Garanina, A.S.; Druart, X.; et al. Oviduct extracellular vesicles protein content and their role during oviduct-embryo cross-talk. Reproduction 2017, 154, 153–168. [Google Scholar] [CrossRef]
- Hunter, R.H. Components of oviduct physiology in eutherian mammals. Biol. Rev. Camb. Philos. Soc. 2012, 287, 244–255. [Google Scholar] [CrossRef]
- Richardson, L.L.; Oliphant, G. Steroid concentrations in rabbit oviducal fluid during oestrus and pseudopregnancy. J. Reprod. Fertil. 1981, 62, 427–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballester, L.; Romero-Aguirregomezcorta, J.; Soriano-Ubeda, C.; Matas, C.; Romar, R.; Coy, P. Timing of oviductal fluid collection, steroid concentrations, and sperm preservation method affect porcine in vitro fertilization efficiency. Fertil. Steril. 2014, 102, 1762–1768. [Google Scholar] [CrossRef] [PubMed]
- Nelis, H.; Vanden Bussche, J.; Wojciechowicz, B.; Franczak, A.; Vanhaecke, L.; Leemans, B.; Cornillie, P.; Peelman, L.; Van Soom, A.; Smits, K. Steroids in the equine oviduct: Synthesis, local concentrations and receptor expression. Reprod. Fertil. Dev. 2015, 28, 1390–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamy, J.; Liere, P.; Pianos, A.; Aprahamian, F.; Mermillod, P.; Saint-Dizier, M. Steroid hormones in bovine oviductal fluid during the estrous cycle. Theriogenology 2016, 86, 1409–1420. [Google Scholar] [CrossRef]
- Pinto-Bravo, P.; Galvao, A.; Rebordao, M.R.; Amaral, A.; Ramilo, D.; Silva, E.; Szostek-Mioduchowska, A.; Alexandre-Pires, G.; Roberto da Costa, R.; Skarzynski, D.J.; et al. Ovarian steroids, oxytocin, and tumor necrosis factor modulate equine oviduct function. Domest. Anim. Endocrinol. 2017, 61, 84–99. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Wakamiya, K.; Kohka, M.; Yamamoto, Y.; Okuda, K. Summer heat stress affects prostaglandin synthesis in the bovine oviduct. Reproduction 2013, 146, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Martinez, H.; Petroni, A.; Einarsson, S.; Kindahl, H. Concentrations of prostaglandin F2 alpha in the pig oviductal fluid. Prostaglandins 1983, 25, 413–424. [Google Scholar] [CrossRef]
- Malysz-Cymborska, I.; Ziecik, A.J.; Waclawik, A.; Andronowska, A. Effect of hCG and eCG treatments on prostaglandins synthesis in the porcine oviduct. Reprod. Domest. Anim. 2013, 48, 1034–1042. [Google Scholar] [CrossRef]
- Ezzati, M.; Djahanbakhch, O.; Arian, S.; Carr, B.R. Tubal transport of gametes and embryos: A review of physiology and pathophysiology. J. Assist. Reprod. Genet. 2014, 31, 1337–1347. [Google Scholar] [CrossRef] [Green Version]
- Lei, Z.M.; Toth, P.; Rao, C.V.; Pridham, D. Novel coexpression of human chorionic gonadotropin (hCG)/human luteinizing hormone receptors and their ligand hCG in human fallopian tubes. J. Clin. Endocrinol. Metab. 1993, 77, 863–872. [Google Scholar] [CrossRef]
- Zhang, M.; Shi, H.; Segaloff, D.L.; Van Voorhis, B.J. Expression and localization of luteinizing hormone receptor in the female mouse reproductive tract. Biol. Reprod. 2001, 64, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Magid, M.S.; Kramer, E.E.; Chen, Y.T. Follicle-stimulating hormone receptor is expressed in human ovarian surface epithelium and fallopian tube. Am. J. Pathol. 1996, 148, 47–53. [Google Scholar] [PubMed]
- Gawronska, B.; Paukku, T.; Huhtaniemi, I.; Wasowicz, G.; Ziecik, A.J. Oestrogen-dependent expression of LH/hCG receptors in pig Fallopian tube and their role in relaxation of the oviduct. J. Reprod. Fertil. 1999, 115, 293–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malysz-Cymborska, I.; Andronowska, A. Downregulation of LH and FSH receptors after hCG and eCG treatments in the porcine oviduct. Domest. Anim. Endocrinol. 2016, 57, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, Y.; Forde, N.; Blum, H.; Graf, A.; Piégu, B.; Mermillod, P.; Wolf, E.; Lonergan, P.; Saint-Dizier, M. Relative effects of location relative to the corpus luteum and lactation on the transcriptome of the bovine oviduct epithelium. BMC Genom. 2019, 20, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alminana, C.; Caballero, I.; Heath, P.R.; Maleki-Dizaji, S.; Parrilla, I.; Cuello, C.; Gil, M.A.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; et al. The battle of the sexes starts in the oviduct: Modulation of oviductal transcriptome by X and Y-bearing spermatozoa. BMC Genom. 2014, 15, 293. [Google Scholar] [CrossRef] [Green Version]
- Sostaric, E.; Dieleman, S.J.; van de Lest, C.H.; Colenbrander, B.; Vos, P.L.; Garcia-Gil, N.; Gadella, B.M. Sperm binding properties and secretory activity of the bovine oviduct immediately before and after ovulation. Mol. Reprod. Dev. 2008, 75, 60–74. [Google Scholar] [CrossRef]
- Marey, M.A.; Liu, J.; Kowsar, R.; Haneda, S.; Matsui, M.; Sasaki, M.; Takashi, S.; Hayakawa, H.; Wijayagunawardane, M.P.; Hussein, F.M.; et al. Bovine oviduct epithelial cells downregulate phagocytosis of sperm by neutrophils: Prostaglandin E2 as a major physiological regulator. Reproduction 2014, 147, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Marey, M.A.; Yousef, M.S.; Kowsar, R.; Hambruch, N.; Shimizu, T.; Pfarrer, C.; Miyamoto, A. Local immune system in oviduct physiology and pathophysiology: Attack or tolerance? Domest. Anim. Endocrinol. 2016, 56, S204–S211. [Google Scholar] [CrossRef]
- Yousef, M.S.; Marey, M.A.; Hambruch, N.; Hayakawa, H.; Shimizu, T.; Hussien, H.A.; Abdel-Razek, A.K.; Pfarrer, C.; Miyamoto, A. Sperm Binding to Oviduct Epithelial Cells Enhances TGFB1 and IL10 Expressions in Epithelial Cells as Well as Neutrophils In Vitro: Prostaglandin E2 As a Main Regulator of Anti-Inflammatory Response in the Bovine Oviduct. PLoS ONE 2016, 11, e0162309. [Google Scholar] [CrossRef]
- Wetscher, F.; Havlicek, V.; Huber, T.; Muller, M.; Brem, G.; Besenfelder, U. Effect of morphological properties of transferred embryonic stages on tubal migration Implications for in vivo culture in the bovine oviduct. Theriogenology 2005, 64, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Betteridge, K.J.; Eaglesome, M.D.; Flood, P.F. Embryo transport through the mare’s oviduct depends upon cleavage and is independent of the ipsilateral corpus luteum. J. Reprod. Fertil. Suppl. 1979, 27, 387–394. [Google Scholar]
- Weber, J.A.; Freeman, D.A.; Vanderwall, D.K.; Woods, G.L. Prostaglandin-E2 Hastens Oviductal Transport of Equine Embryos. Biol. Reprod. 1991, 45, 544–546. [Google Scholar] [CrossRef] [PubMed]
- Rizos, D.; Ramirez, M.A.; Pintado, B.; Lonergan, P.; Gutierrez-Adan, A. Culture of bovine embryos in intermediate host oviducts with emphasis on the isolated mouse oviduct. Theriogenology 2010, 73, 777–785. [Google Scholar] [CrossRef]
- Gad, A.; Hoelker, M.; Besenfelder, U.; Havlicek, V.; Cinar, U.; Rings, F.; Held, E.; Dufort, I.; Sirard, M.A.; Schellander, K.; et al. Molecular mechanisms and pathways involved in bovine embryonic genome activation and their regulation by alternative in vivo and in vitro culture conditions. Biol. Reprod. 2012, 87, 100. [Google Scholar] [CrossRef]
- Enright, B.P.; Lonergan, P.; Dinnyes, A.; Fair, T.; Ward, F.A.; Yang, X.; Boland, M.P. Culture of in vitro produced bovine zygotes in vitro vs. in vivo: Implications for early embryo development and quality. Theriogenology 2000, 54, 659–673. [Google Scholar] [CrossRef]
- Lee, K.F.; Yao, Y.Q.; Kwok, K.L.; Xu, J.S.; Yeung, W.S. Early developing embryos affect the gene expression patterns in the mouse oviduct. Biochem. Biophys. Res. Commun. 2002, 292, 564–570. [Google Scholar] [CrossRef]
- Alminana, C.; Heath, P.R.; Wilkinson, S.; Sanchez-Osorio, J.; Cuello, C.; Parrilla, I.; Gil, M.A.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; et al. Early developing pig embryos mediate their own environment in the maternal tract. PLoS ONE 2012, 7, e33625. [Google Scholar] [CrossRef] [Green Version]
- Hamdi, M.; Sanchez Calabuig, M.J.; Rodriguez-Alonso, B.; Bages-Arnal, S.; Roussi, K.; Sturmey, R.G.S.; Gutierrez-Adan, A.; Lonergan, P.; Rizos, D. Gene expression and metabolic response of bovine oviduct epithelial cells to the early embryo. Reproduction 2019, 158, 85–94. [Google Scholar] [CrossRef]
- Schmaltz-Panneau, B.; Cordova, A.; Dhorne-Pollet, S.; Hennequet-Antier, C.; Uzbekova, S.; Martinot, E.; Doret, S.; Martin, P.; Mermillod, P.; Locatelli, Y. Early bovine embryos regulate oviduct epithelial cell gene expression during in vitro co-culture. Anim. Reprod. Sci. 2014, 149, 103–116. [Google Scholar] [CrossRef]
- Maillo, V.; Gaora, P.O.; Forde, N.; Besenfelder, U.; Havlicek, V.; Burns, G.W.; Spencer, T.E.; Gutierrez-Adan, A.; Lonergan, P.; Rizos, D. Oviduct-Embryo Interactions in Cattle: Two-Way Traffic or a One-Way Street? Biol. Reprod. 2015, 92, 144. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Alonso, B.; Hamdi, M.; Sanchez, J.M.; Maillo, V.; Gutierrez-Adan, A.; Lonergan, P.; Rizos, D. An approach to study the local embryo effect on gene expression in the bovine oviduct epithelium in vivo. Reprod. Domest. Anim. 2019, 15, 1516–1523. [Google Scholar] [CrossRef] [PubMed]
- Smits, K.; De Coninck, D.I.; Van Nieuwerburgh, F.; Govaere, J.; Van Poucke, M.; Peelman, L.; Deforce, D.; Van Soom, A. The Equine Embryo Influences Immune-Related Gene Expression in the Oviduct. Biol. Reprod. 2016, 94, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijayagunawardane, M.P.B.; Miyamoto, A.; Cerbito, W.A.; Acosta, T.J.; Takagi, M.; Sato, K. Local distributions of oviductal estradiol, progesterone, prostaglandins, oxytocin and endothelin-1 in the cyclic cow. Theriogenology 1998, 49, 607–618. [Google Scholar] [CrossRef]
- Binelli, M.; Gonella-Diaza, A.M.; Mesquita, F.S.; Membrive, C.M.B. Sex Steroid-Mediated Control of Oviductal Function in Cattle. Biology 2018, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aviles, M.; Gutierrez-Adan, A.; Coy, P. Oviductal secretions: Will they be key factors for the future ARTs? Mol. Hum. Reprod. 2010, 16, 896–906. [Google Scholar] [CrossRef]
- Saint-Dizier, M.; Marnier, C.; Tahir, M.Z.; Grimard, B.; Thoumire, S.; Chastant-Maillard, S.; Reynaud, K. OVGP1 is expressed in the canine oviduct at the time and place of oocyte maturation and fertilization. Mol. Reprod. Dev. 2014, 81, 972–982. [Google Scholar] [CrossRef]
- Chen, S.; Palma-Vera, S.E.; Kempisty, B.; Rucinski, M.; Vernunft, A.; Schoen, J. In Vitro Mimicking of Estrous Cycle Stages: Dissecting the Impact of Estradiol and Progesterone on Oviduct Epithelium. Endocrinology 2018, 159, 3421–3432. [Google Scholar] [CrossRef]
- Ulbrich, S.E.; Kettler, A.; Einspanier, R. Expression and localization of estrogen receptor alpha, estrogen receptor beta and progesterone receptor in the bovine oviduct in vivo and in vitro. J. Steroid. Biochem. Mol. Biol. 2003, 84, 279–289. [Google Scholar] [CrossRef]
- Saint-Dizier, M.; Sandra, O.; Ployart, S.; Chebrout, M.; Constant, F. Expression of nuclear progesterone receptor and progesterone receptor membrane components 1 and 2 in the oviduct of cyclic and pregnant cows during the post-ovulation period. Reprod. Biol. Endocrinol. 2012, 10, 76. [Google Scholar] [CrossRef]
- Licht, P.; Gallo, A.B.; Aggarwal, B.B.; Farmer, S.W.; Castelino, J.B.; Papkoff, H. Biological and binding activities of equine pituitary gonadotrophins and pregnant mare serum gonadotrophin. J. Endocrinol. 1979, 83, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Juarez-Mosqueda, M.L.; Anzaldua Arce, S.R.; Palma Lara, I.; Garcia Dalman, C.; Cornejo Cortes, M.A.; Cordova Izquierdo, A.; Villasenor Gaona, H.; Trujillo Ortega, M.E. Oestrus synchronization treatment induces histomorphological changes on the uterine tube epithelium of the gilt. Anat. Histol. Embryol. 2015, 44, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Fontes, P.K.; Castilho, A.C.; Razza, E.M.; Ereno, R.L.; Satrapa, R.A.; Barros, C.M. Prostaglandin receptors (EP2 and EP4) and angiotensin receptor (AGTR2) mRNA expression increases in the oviducts of Nelore cows submitted to ovarian superstimulation. Anim. Reprod. Sci. 2014, 151, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salzano, A.; De Canditiis, C.; Della Ragione, F.; Prandi, A.; Zullo, G.; Neglia, G.; Campanile, G.; Gasparrini, B. Evaluation of factors involved in the failure of ovum capture in superovulated buffaloes. Theriogenology 2018, 122, 102–108. [Google Scholar] [CrossRef]
- Carvalho, N.A.; Baruselli, P.S.; Zicarelli, L.; Madureira, E.H.; Visintin, J.A.; D’Occhio, M.J. Control of ovulation with a GnRH agonist after superstimulation of follicular growth in buffalo: Fertilization and embryo recovery. Theriogenology 2002, 58, 1641–1650. [Google Scholar] [CrossRef]
- Jordaens, L.; Van Hoeck, V.; Maillo, V.; Gutierrez-Adan, A.; Marei, W.F.A.; Vlaeminck, B.; Thys, S.; Sturmey, R.G.; Bols, P.E.J.; Leroy, J. Maternal metabolic stress may affect oviduct gatekeeper function. Reproduction 2017, 153, 759–773. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, C.C.L.; Rodriguez-Villamil, P.; Vasconcelos, F.R.; Nagano, C.S.; Rossetto, R.; Moura, A.; Rondina, D. Proteome of the periovulatory oviduct and uterus of goats as related to nutritional balance. Reprod. Domest. Anim. 2018, 53, 1085–1095. [Google Scholar] [CrossRef]
- Maillo, V.; Rizos, D.; Besenfelder, U.; Havlicek, V.; Kelly, A.K.; Garrett, M.; Lonergan, P. Influence of lactation on metabolic characteristics and embryo development in postpartum Holstein dairy cows. J. Dairy Sci. 2012, 95, 3865–3876. [Google Scholar] [CrossRef]
- Rizos, D.; Carter, F.; Besenfelder, U.; Havlicek, V.; Lonergan, P. Contribution of the female reproductive tract to low fertility in postpartum lactating dairy cows. J. Dairy Sci. 2010, 93, 1022–1029. [Google Scholar] [CrossRef]
- Fenwick, M.A.; Llewellyn, S.; Fitzpatrick, R.; Kenny, D.A.; Murphy, J.J.; Patton, J.; Wathes, D.C. Negative energy balance in dairy cows is associated with specific changes in IGF-binding protein expression in the oviduct. Reproduction 2008, 135, 63–75. [Google Scholar] [CrossRef] [Green Version]
- Gegenfurtner, K.; Frohlich, T.; Kosters, M.; Mermillod, P.; Locatelli, Y.; Fritz, S.; Salvetti, P.; Forde, N.N.; Lonergan, P.; Wolf, E.; et al. Influence of metabolic status and genetic merit for fertility on proteomic composition of bovine oviduct fluid. Biol. Reprod. 2019, 101, 893–905. [Google Scholar] [CrossRef]
- Desmet, K.L.; Van Hoeck, V.; Gagne, D.; Fournier, E.; Thakur, A.; O’Doherty, A.M.; Walsh, C.P.; Sirard, M.A.; Bols, P.E.; Leroy, J.L. Exposure of bovine oocytes and embryos to elevated non-esterified fatty acid concentrations: Integration of epigenetic and transcriptomic signatures in resultant blastocysts. BMC Genom. 2016, 17, 1004. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. Effects of heat stress on mammalian reproduction. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 3341–3350. [Google Scholar] [CrossRef] [PubMed]
- Bahat, A.; Eisenbach, M. Sperm thermotaxis. Mol. Cell. Endocrinol. 2006, 252, 115–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, R.H. Temperature gradients in female reproductive tissues. Reprod. Biomed. Online 2012, 24, 377–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapala, L.; Starzynski, R.R.; Trzeciak, P.Z.; Dabrowski, S.; Gajewska, M.; Jurka, P.; Smolarczyk, R.; Duszewska, A.M. Influence of elevated temperature on bovine oviduct epithelial cells (BOECs). PLoS ONE 2018, 13, e0198843. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor Studied | OF or OEC Component | Species | Main Result | Ref. |
|---|---|---|---|---|
| Ovulation | OF proteins | Cattle | Comparison between ipsilateral and contralateral oviducts identified up to 115 differentially abundant proteins across the estrous cycle | [39] |
| Ovulation | OF proteins | Horse | Seven proteins were differentially abundant between ipsilateral and contralateral oviducts in non-pregnant mares | [42] |
| Ovulation | OF steroid hormones | Cattle | Concentrations of OF progesterone and progesterone metabolites differed between ipsilateral and contralateral oviducts across the estrous cycle | [55] |
| Ovulation | OF steroid hormones | Horse | Concentrations of OF progesterone differed between ipsilateral and contralateral oviducts in the post-ovulatory period | [54] |
| Proximity of the corpus luteum | OEC gene expression | Cattle | Irrespective of the metabolic status of females, comparison between ipsilateral and contralateral oviducts identified 192 and 2583 DEGs in the ampulla and isthmus, respectively, on day 3 post-estrus | [66] |
| Proximity of the corpus luteum | OEC gene expression | Horse | Comparison between ipsilateral and contralateral oviducts indicated 164 DEGs in pregnant mares and 77 DEGs in cyclic mares | [84] |
| Ovulation and Insemination | OF proteins | Pig | Spermatozoa and oocyte–cumulus complexes altered the oviductal secretory proteome 24 h after ovulation and insemination | [35] |
| Insemination | OF proteins | Rabbit | Secreted OF proteins changed 4 and 8 h after insemination with region-specific alterations | [43] |
| Sex-sorted spermatozoa | OEC gene expression | Pig | Differentially expressed genes were identified in OECs in the presence of Y-chromosome bearing spermatozoa compared with X-chromosome bearing spermatozoa | [67] |
| Presence of embryos | OEC gene expression | Mouse | The expression of specific genes was upregulated in OECs in the presence of early embryos compared with non-fertilized oocytes | [78] |
| Presence of embryos | OEC gene expression | Pig | The expression of TICAM2 was upregulated in the oviduct epithelium by the presence of embryos | [79] |
| Presence of one and multiple embryos | OEC gene expression | Cattle | The presence of multiple embryos in the oviduct resulted in the detection of DEGs in the isthmus of beef heifers on day 3 post-estrus; no DEGs could be detected in the presence of a single eight-cell embryo | [82] |
| Presence of one embryo | OEC ciliary beating | Cattle | A local downregulation of particle transport speed was evidenced in the site of the embryo in oviduct sections ex vivo | [12] |
| Pregnancy | OF proteins | Horse | The presence of an embryo in the ipsilateral OF of pregnant mares induced regulation of 13 proteins compared with the contralateral side, and of 19 proteins compared with the ipsilateral side of non-pregnant mares. | [42] |
| Pregnancy | OEC gene expression | Horse | Comparison between ipsilateral pregnant and non-pregnant oviducts identified 253 upregulated genes and 108 downregulated genes in OECs | [84] |
| Sex steroid hormones | Phospholipids | Cattle | Different phospholipid profiles were evidenced in oviducts from females with contrasted progesterone and estradiol levels during early diestrus | [32] |
| Stage of cycle | OF steroid hormones, proteins, metabolites, and lipids | Cattle | Comparison between four stages of the estrous cycle identified differentially abundant OF sex steroid hormones, proteins, amino acids, energy substrates, and phospholipids in both sides relative to ovulation | [26,31,39,58] |
| Stage of cycle | OF steroid hormones | Horse | Comparison between pre-ovulatory and post-ovulatory oviducts identified differential OF levels of progesterone in the side of ovulation | [54] |
| Progesterone | OF proteins | Dog | Treatment with the progesterone receptor antagonist aglepristone induced changes in the abundance of 79 OF proteins at day 4 post-ovulation | [37] |
| Progesterone | OF ions and metabolites | Cattle | Systemic supplementation with progesterone induced changes in OF amino acids, sulfate, and sodium | [18] |
| Superovulation treatment | Prostaglandin synthesis | Pig | Stimulation with hCG/eCG before insemination affected prostaglandin synthesis pathway on day 3 post-estrus in gilts | [59] |
| Superovulation treatment | OEC gene expression | Cattle | Superovulation treatment with FSH and eCG changed the expression of prostaglandin receptors EP2 and EP4 in the ampulla and infundibulum and of AGTR2 in the isthmus | [94] |
| Superovulation treatment | OEC gene expression | Cattle (Buffalo) | Superovulation treatment with FSH decreased the expression of steroid hormone receptors PGR and ESR1, VEGF, and its receptor FLT1 | [95] |
| Estrus synchronization | OF proteins | Sheep | Proteins found differentially abundant between estrus and the luteal phase differed when comparing ewes in spontaneous cycles with those treated for estrus synchronization | [38] |
| Energy balance | OEC proteins | Goat | Comparison between four different diet groups identified seven differentially expressed proteins in ampullas of animals fed with 1.9 times live weight maintenance | [98] |
| Energy balance | OEC gene expression | Cattle | Negative energy balance was associated with changes in gene expression of IGFBP-2 and IGFBP-6 in the oviducts of lactating dairy cows | [101] |
| Energy balance | OEC gene expression | Cattle | Comparison between OECs from postpartum lactating and non-lactating dairy cows evidenced 15 DEGs in the isthmus and none in the ampulla | [66] |
| Energy balance | OF proteins | Cattle | Comparison between OF from postpartum lactating and non-lactating dairy cows evidenced 12 differentially abundant proteins | [102] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saint-Dizier, M.; Schoen, J.; Chen, S.; Banliat, C.; Mermillod, P. Composing the Early Embryonic Microenvironment: Physiology and Regulation of Oviductal Secretions. Int. J. Mol. Sci. 2020, 21, 223. https://doi.org/10.3390/ijms21010223
Saint-Dizier M, Schoen J, Chen S, Banliat C, Mermillod P. Composing the Early Embryonic Microenvironment: Physiology and Regulation of Oviductal Secretions. International Journal of Molecular Sciences. 2020; 21(1):223. https://doi.org/10.3390/ijms21010223
Chicago/Turabian StyleSaint-Dizier, Marie, Jennifer Schoen, Shuai Chen, Charles Banliat, and Pascal Mermillod. 2020. "Composing the Early Embryonic Microenvironment: Physiology and Regulation of Oviductal Secretions" International Journal of Molecular Sciences 21, no. 1: 223. https://doi.org/10.3390/ijms21010223