Transcriptomic and Proteomic Analyses Reveal the Potential Mode of Action of Chondrocyte Sheets in Hyaline Cartilage Regeneration

 , ,
, , 
Abstract
:1. Introduction
2. Results
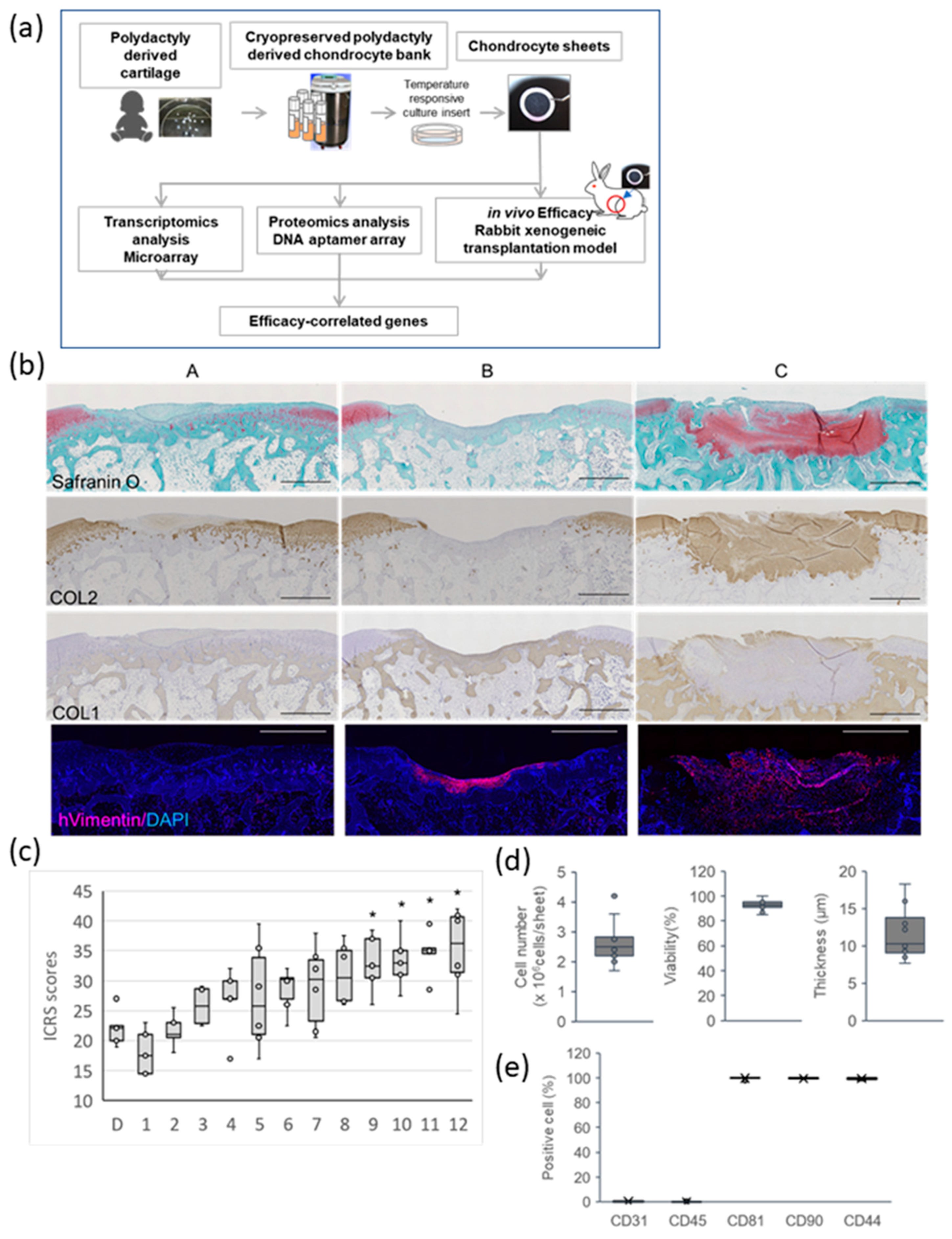
2.1. Effect of PD Sheets on Hyaline Cartilage Regeneration
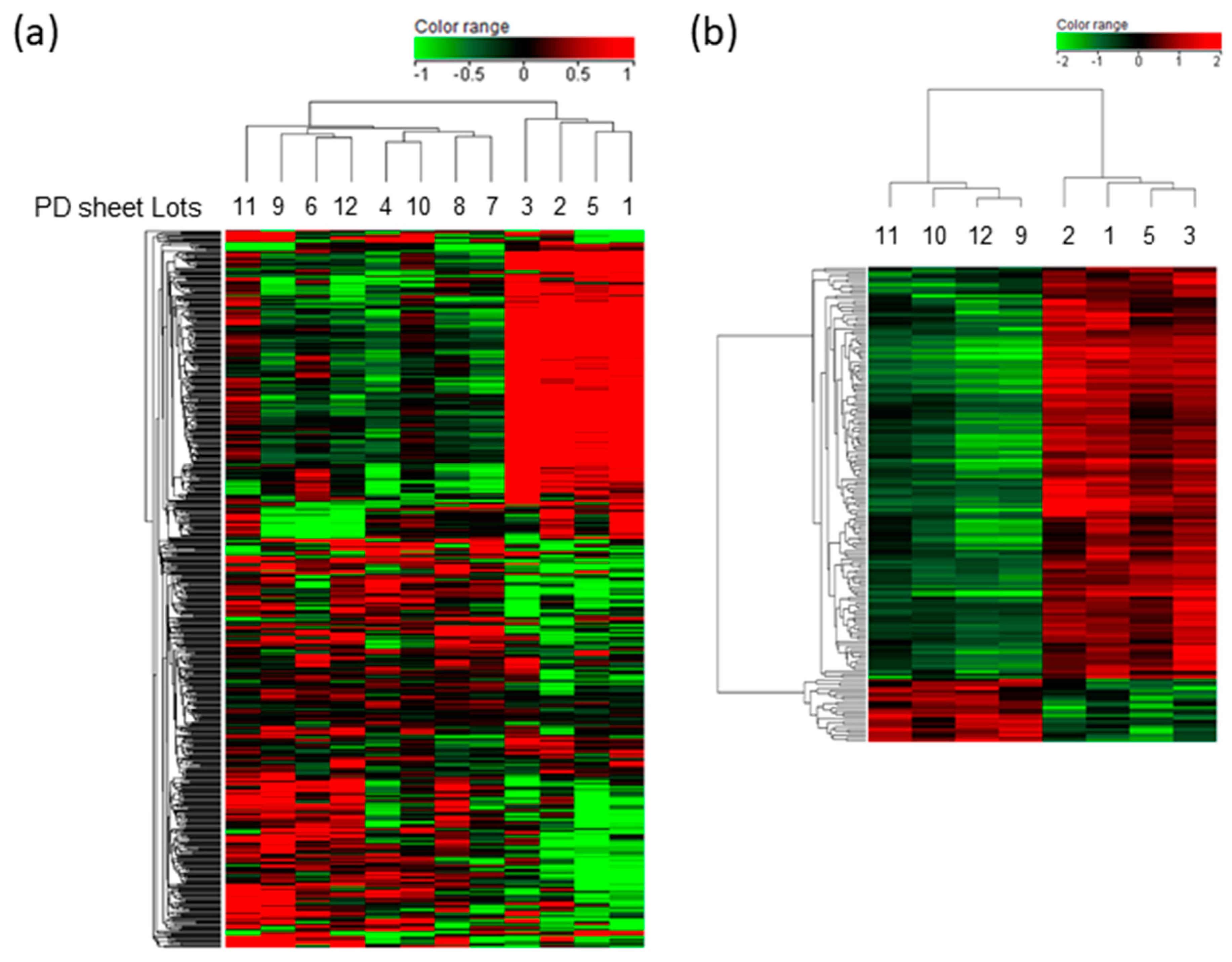
2.2. Gene Expression Profiles Associated with the Effectiveness of PD Sheets
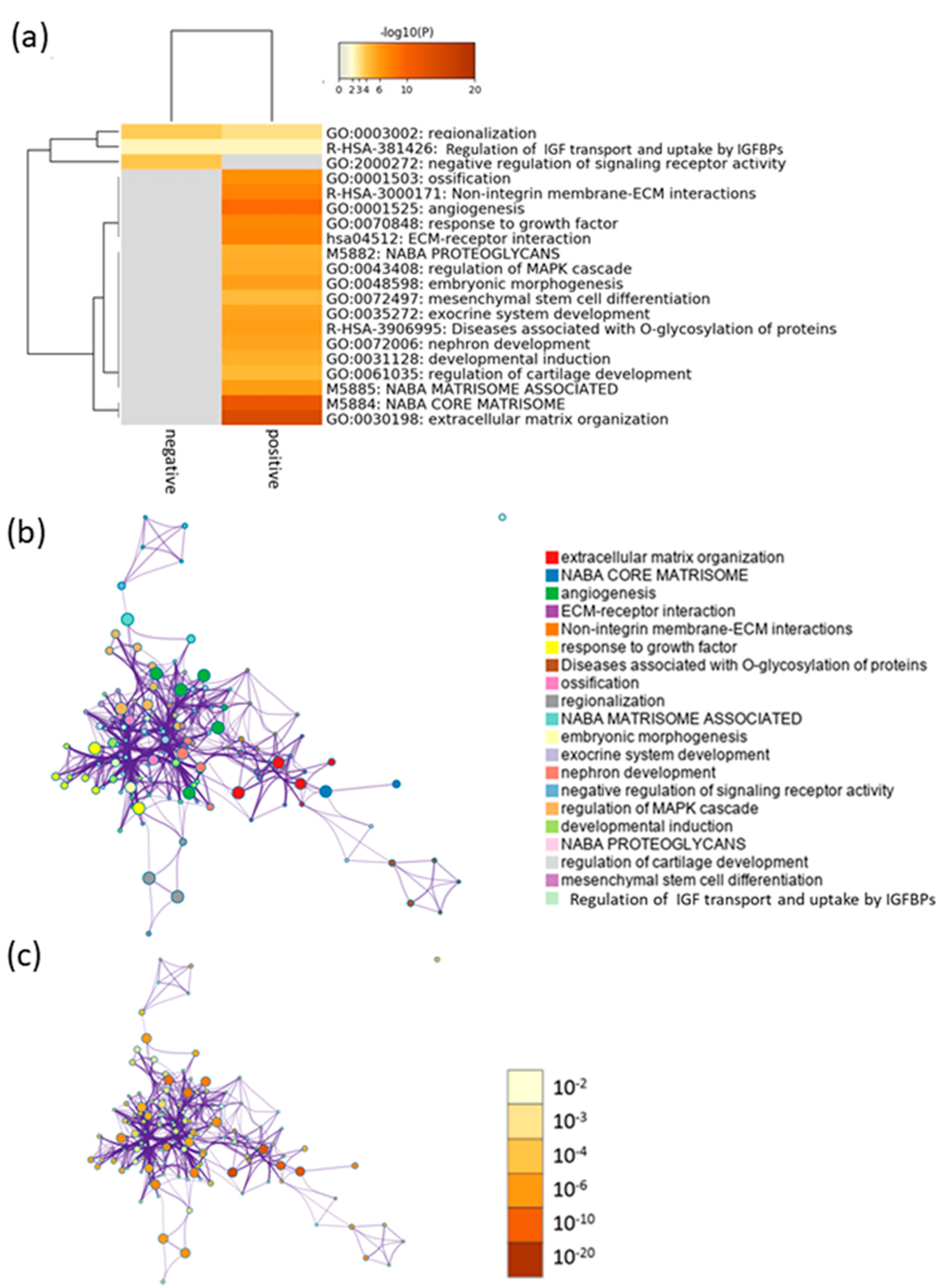
2.3. Function Prediction by Efficacy-Related Genes
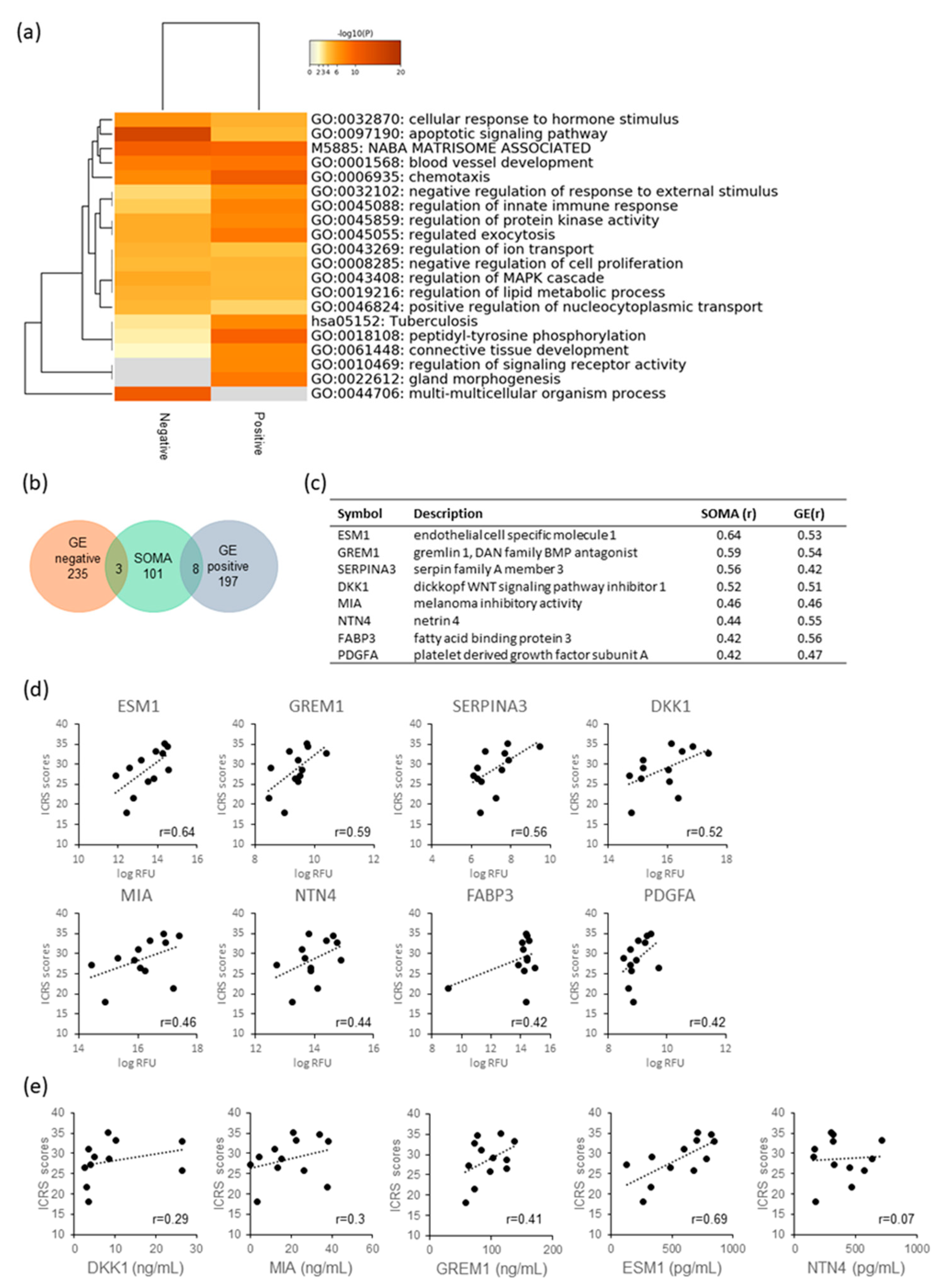
2.4. Analysis of Secretory Factors Related to PD Sheet Effectiveness
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Fabrication of PD Sheets
4.2.1. Preparation of Chondrocytes
4.2.2. Fabrication of PD Sheets
4.2.3. Evaluation of Characteristics of PD Sheets
4.3. Evaluation of the Efficacy of PD Sheets for Osteochondral Defects
4.3.1. Immunosuppressed Rabbit Osteochondral Defect Model
4.3.2. PD Sheet Transplantation
4.3.3. Histological Scoring for Regenerated Cartilage
4.3.4. Immunostaining
4.4. Gene Expression Analysis of PD Sheets
4.4.1. RNA Isolation
4.4.2. cRNA Amplification and Labeling
4.4.3. Sample Hybridization
4.4.4. Identification of Efficacy Correlation Genes
4.4.5. Enrichment Analysis
4.4.6. RT-PCR
4.5. Analysis of Proteins Secreted by PD Sheets
4.5.1. Preparation of the Supernatant of the PD sheets
4.5.2. Proteomic Analysis Using SOMAscan
4.5.3. Enzyme-Linked Immunosorbent Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACAN | Aggrecan |
| ACTB | Actin beta |
| APOE | Apolipoprotein E |
| CLS1 | Collagenase Type I |
| COL1 | Type I collagen |
| COL2 | Type II collagen |
| Ct | Cycle threshold |
| Cy3 | Cyanine 3 |
| ΔCt | Delta Ct |
| DKK1 | Dickkopf WNT signaling pathway inhibitor 1 |
| DMEM/F12 | Dulbecco’s modified Eagle’s medium–Nutrient Mixture F-12 |
| EDTA | Ethylenediaminetetraacetic acid |
| ELISA | Enzyme-linked immunosorbent assay |
| ESM1 | Endothelial cell-specific molecule 1 |
| FABP3 | Fatty acid-binding protein 3 |
| GDF5 | Growth differentiation factor 5 |
| GES | Gene expression score |
| GO | Gene ontology |
| GREM1 | Gremlin 1 |
| H&E | Hematoxylin and eosin |
| ICRS | International Cartilage Repair Society |
| ITGA10 | Integrin alpha 10 |
| MIA | Melanoma inhibitory activity |
| MSC | Mesenchymal stem cells |
| NGS | Normal goat serum |
| NTN4 | Netrin 4 |
| OA | Osteoarthritis |
| PBS | Phosphate buffer saline |
| PCR | Polymerase chain reaction |
| PD | Polydactyly-derived |
| PD sheet | PD chondrocyte sheet |
| PDGFA | Platelet-derived growth factor subunit A |
| PGF | Placental growth factor |
| RARRES2 | Retinoic acid receptor responder 2 |
| RT | Room temperature |
| RT-PCR | Reverse transcription PCR |
| SERPINA3 | Serpin family A member 3 |
| SOMA | Slow off-rate modified aptamers |
| SOX5 | SRY-box transcription factor 5 |
| SOX9 | SRY-box transcription factor 9 |
| SRY | Sex-determining region Y |
| TE | Tris and EDTA |
References
- Yoshimura, N.; Muraki, S.; Oka, H.; Mabuchi, A.; En-Yo, Y.; Yoshida, M.; Saika, A.; Yoshida, H.; Suzuki, T.; Yamamoto, S.; et al. Prevalence of knee osteoarthritis, lumbar spondylosis, and osteoporosis in Japanese men and women: The research on osteoarthritis/osteoporosis against disability study. J. Bone Miner. Metab. 2009, 27, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Yamada, N.; Sakai, H.; Sakurai, Y. A novel recovery system for cultured cells using plasma-treated polystyrene dishes grafted with poly (N-isopropylacrylamide). J. Biomed. Mater. Res. 1993, 27, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Yamada, N.; Okuhara, M.; Sakai, H.; Sakurai, Y. Mechanism of cell detachment from temperature-modulated, hydrophilic-hydrophobic polymer surfaces. Biomaterials 1995, 16, 297–303. [Google Scholar] [CrossRef]
- Kaneshiro, N.; Sato, M.; Ishihara, M.; Mitani, G.; Sakai, H.; Mochida, J. Bioengineered chondrocyte sheets may be potentially useful for the treatment of partial thickness defects of articular cartilage. Biochem. Biophys. Res. Commun. 2006, 349, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, G.; Sato, M.; Yamato, M.; Mitani, G.; Kutsuna, T.; Nagai, T.; Ito, S.; Ukai, T.; Kobayashi, M.; Kokubo, M.; et al. Cartilage repair in transplanted scaffold-free chondrocyte sheets using a minipig model. Biomaterials 2012, 33, 3846–3851. [Google Scholar] [CrossRef]
- Ito, S.; Sato, M.; Yamato, M.; Mitani, G.; Kutsuna, T.; Nagai, T.; Ukai, T.; Kobayashi, M.; Kokubo, M.; Okano, T.; et al. Repair of articular cartilage defect with layered chondrocyte sheets and cultured synovial cells. Biomaterials 2012, 33, 5278–5286. [Google Scholar] [CrossRef]
- Tani, Y.; Sato, M.; Maehara, M.; Nagashima, H.; Yokoyama, M.; Yokoyama, M.; Yamato, M.; Okano, T.; Mochida, J. The effects of using vitrified chondrocyte sheets on pain alleviation and articular cartilage repair. J. Tissue Eng. Regen. Med. 2017, 11, 3437–3444. [Google Scholar] [CrossRef]
- Takatori, N.; Sato, M.; Toyoda, E.; Takahashi, T.; Okada, E.; Maehara, M.; Watanabe, M. Cartilage repair and inhibition of the progression of cartilage degeneration after transplantation of allogeneic chondrocyte sheets in a nontraumatic early arthritis model. Regen. Ther. 2018, 9, 24–31. [Google Scholar] [CrossRef]
- Sato, M.; Yamato, M.; Mitani, G.; Takagaki, T.; Hamahashi, K.; Nakamura, Y.; Ishihara, M.; Matoba, R.; Kobayashi, H.; Okano, T.; et al. Combined surgery and chondrocyte cell-sheet transplantation improves clinical and structural outcomes in knee osteoarthritis. NPJ Regen. Med. 2019, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Yanke, A.B.; Tilton, A.K.; Wetters, N.G.; Merkow, D.B.; Cole, B.J. DeNovo NT Particulated Juvenile Cartilage Implant. Sports Med. Arthrosc. 2015, 23, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Maehara, M.; Sato, M.; Toyoda, E.; Takahashi, T.; Okada, E.; Kotoku, T.; Watanabe, M. Characterization of polydactyly-derived chondrocyte sheets versus adult chondrocyte sheets for articular cartilage repair. Inflamm. Regen. 2017, 37, 22. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Sato, M.; Toyoda, E.; Maehara, M.; Takizawa, D.; Maruki, H.; Tominaga, A.; Okada, E.; Okazaki, K.; Watanabe, M. Rabbit xenogeneic transplantation model for evaluating human chondrocyte sheets used in articular cartilage repair. J. Tissue Eng. Regen. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 5, e15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, L.; Walker, J.J.; Wilcox, S.K.; Williams, S. Advances in human proteomics at high scale with the SOMAscan proteomics platform. New Biotechnol. 2012, 29, 543–549. [Google Scholar] [CrossRef]
- Oh, H.; Chun, C.-H.; Chun, J.-S. Dkk-1 expression in chondrocytes inhibits experimental osteoarthritic cartilage destruction in mice. Arthr. Rheum. 2012, 64, 2568–2578. [Google Scholar] [CrossRef]
- Leijten, J.C.H.; Moreira Teixeira, L.S.; Landman, E.B.M.; van Blitterswijk, C.A.; Karperien, M. Hypoxia inhibits hypertrophic differentiation and endochondral ossification in explanted tibiae. PLoS ONE 2012, 7, e49896. [Google Scholar] [CrossRef] [Green Version]
- Bosserhoff, A.K.; Buettner, R. Establishing the protein MIA (melanoma inhibitory activity) as a marker for chondrocyte differentiation. Biomaterials 2003, 24, 3229–3234. [Google Scholar] [CrossRef]
- Schubert, T.; Schlegel, J.; Schmid, R.; Opolka, A.; Grassel, S.; Humphries, M.; Bosserhoff, A.-K. Modulation of cartilage differentiation by melanoma inhibiting activity/cartilage-derived retinoic acid-sensitive protein (MIA/CD-RAP). Exp. Mol. Med. 2010, 42, 166–174. [Google Scholar] [CrossRef]
- Tscheudschilsuren, G.; Bosserhoff, A.K.; Schlegel, J.; Vollmer, D.; Anton, A.; Alt, V.; Schnettler, R.; Brandt, J.; Proetzel, G. Regulation of mesenchymal stem cell and chondrocyte differentiation by MIA. Exp. Cell Res. 2006, 312, 63–72. [Google Scholar] [CrossRef]
- Takaku, Y.; Murai, K.; Ukai, T.; Ito, S.; Kokubo, M.; Satoh, M.; Kobayashi, E.; Yamato, M.; Okano, T.; Takeuchi, M.; et al. In vivo cell tracking by bioluminescence imaging after transplantation of bioengineered cell sheets to the knee joint. Biomaterials 2014, 35, 2199–2206. [Google Scholar] [CrossRef] [PubMed]
- Kaneshiro, N.; Sato, M.; Ishihara, M.; Mitani, G.; Sakai, H.; Kikuchi, T.; Mochida, J. Cultured articular chondrocytes sheets for partial thickness cartilage defects utilizing temperature-responsive culture dishes. Eur. Cell Mater. 2007, 13, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Hamahashi, K.; Sato, M.; Yamato, M.; Kokubo, M.; Mitani, G.; Ito, S.; Nagai, T.; Ebihara, G.; Kutsuna, T.; Okano, T.; et al. Studies of the humoral factors produced by layered chondrocyte sheets. J. Tissue Eng. Regen. Med. 2015, 9, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Kokubo, M.; Sato, M.; Yamato, M.; Mitani, G.; Kutsuna, T.; Ebihara, G.; Okano, T.; Mochida, J. Characterization of chondrocyte sheets prepared using a co-culture method with temperature-responsive culture inserts. J. Tissue Eng. Regen. Med. 2016, 10, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Dell’accio, F.; De Bari, C.; Eltawil, N.M.; Vanhummelen, P.; Pitzalis, C. Identification of the molecular response of articular cartilage to injury, by microarray screening: Wnt-16 expression and signaling after injury and in osteoarthritis. Arthr. Rheum. 2008, 58, 1410–1421. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhong, L.; van Helvoort, E.; Lafeber, F.; Mastbergen, S.; Hendriks, J.; Post, J.N.; Karperien, M. The Expressions of Dickkopf-Related Protein 1 and Frizzled-Related Protein Are Negatively Correlated to Local Inflammation and Osteoarthritis Severity. Cartilage 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leijten, J.C.H.; Bos, S.D.; Landman, E.B.M.; Georgi, N.; Jahr, H.; Meulenbelt, I.; Post, J.N.; van Blitterswijk, C.A.; Karperien, M. GREM1, FRZB and DKK1 mRNA levels correlate with osteoarthritis and are regulated by osteoarthritis-associated factors. Arthr. Res. Ther. 2013, 15, R126. [Google Scholar] [CrossRef] [Green Version]
- Loughlin, J.; Dowling, B.; Chapman, K.; Marcelline, L.; Mustafa, Z.; Southam, L.; Ferreira, A.; Ciesielski, C.; Carson, D.A.; Corr, M. Functional variants within the secreted frizzled-related protein 3 gene are associated with hip osteoarthritis in females. Proc. Natl. Acad. Sci. USA 2004, 101, 9757–9762. [Google Scholar] [CrossRef] [Green Version]
- He, F.; Soriano, P. Dysregulated PDGFRα signaling alters coronal suture morphogenesis and leads to craniosynostosis through endochondral ossification. Development 2017, 144, 4026–4036. [Google Scholar] [CrossRef] [Green Version]
- Peracchia, F.; Ferrari, G.; Poggi, A.; Rotilio, D. IL-1 beta-induced expression of PDGF-AA isoform in rabbit articular chondrocytes is modulated by TGF-beta 1. Exp. Cell Res. 1991, 193, 208–212. [Google Scholar] [CrossRef]
- Yao, Z.; Chen, P.; Wang, S.; Deng, G.; Hu, Y.; Lin, Q.; Zhang, X.; Yu, B. Reduced PDGF-AA in subchondral bone leads to articular cartilage degeneration after strenuous running. J. Cell. Physiol. 2019, 234, 17946–17958. [Google Scholar] [CrossRef] [PubMed]
- Enoki, Y.; Sato, T.; Kokabu, S.; Hayashi, N.; Iwata, T.; Yamato, M.; Usui, M.; Matsumoto, M.; Tomoda, T.; Ariyoshi, W.; et al. Netrin-4 Promotes Differentiation and Migration of Osteoblasts. In Vivo 2017, 31, 793–799. [Google Scholar] [PubMed] [Green Version]
- Zhou, Z.; Xie, J.; Lee, D.; Liu, Y.; Jung, J.; Zhou, L.; Xiong, S.; Mei, L.; Xiong, W.-C. Neogenin regulation of BMP-induced canonical Smad signaling and endochondral bone formation. Dev. Cell 2010, 19, 90–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Shao, Y.; Liu, T.; Qu, Y.-L.; Li, W.; Liu, Z. Therapeutic effects of topical netrin-4 inhibits corneal neovascularization in alkali-burn rats. PLoS ONE 2015, 10, e0122951. [Google Scholar] [CrossRef] [PubMed]
- Lejmi, E.; Leconte, L.; Pédron-Mazoyer, S.; Ropert, S.; Raoul, W.; Lavalette, S.; Bouras, I.; Feron, J.-G.; Maitre-Boube, M.; Assayag, F.; et al. Netrin-4 inhibits angiogenesis via binding to neogenin and recruitment of Unc5B. Proc. Natl. Acad. Sci. USA 2008, 105, 12491–12496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechard, D.; Meignin, V.; Scherpereel, A.; Oudin, S.; Kervoaze, G.; Bertheau, P.; Janin, A.; Tonnel, A.; Lassalle, P. Characterization of the secreted form of endothelial-cell-specific molecule 1 by specific monoclonal antibodies. J. Vasc. Res. 2000, 37, 417–425. [Google Scholar] [CrossRef]
- Shin, J.W.; Huggenberger, R.; Detmar, M. Transcriptional profiling of VEGF-A and VEGF-C target genes in lymphatic endothelium reveals endothelial-specific molecule-1 as a novel mediator of lymphangiogenesis. Blood 2008, 112, 2318–2326. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Chung, U.; Tanaka, S.; Saito, T. Determination of differential gene expression profiles in superficial and deeper zones of mature rat articular cartilage using RNA sequencing of laser microdissected tissue specimens. Biomed. Res. 2014, 35, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Rocha, S.F.; Schiller, M.; Jing, D.; Li, H.; Butz, S.; Vestweber, D.; Biljes, D.; Drexler, H.C.A.; Nieminen-Kelhä, M.; Vajkoczy, P.; et al. Esm1 modulates endothelial tip cell behavior and vascular permeability by enhancing VEGF bioavailability. Circ. Res. 2014, 115, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Furukawa, K.S.; Sato, M.; Ushida, T.; Mochida, J. Characteristics of a scaffold-free articular chondrocyte plate grown in rotational culture. Tissue Eng. Part A 2008, 14, 1183–1193. [Google Scholar] [CrossRef]
- Nagai, T.; Sato, M.; Kutsuna, T.; Kokubo, M.; Ebihara, G.; Ohta, N.; Mochida, J. Intravenous administration of anti-vascular endothelial growth factor humanized monoclonal antibody bevacizumab improves articular cartilage repair. Arthr. Res. Ther. 2010, 12, R178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeuf, S.; Steck, E.; Pelttari, K.; Hennig, T.; Buneb, A.; Benz, K.; Witte, D.; Sültmann, H.; Poustka, A.; Richter, W. Subtractive gene expression profiling of articular cartilage and mesenchymal stem cells: Serpins as cartilage-relevant differentiation markers. Osteoarthr. Cartil. 2008, 16, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensaad, K.; Favaro, E.; Lewis, C.A.; Peck, B.; Lord, S.; Collins, J.M.; Pinnick, K.E.; Wigfield, S.; Buffa, F.M.; Li, J.-L.; et al. Fatty acid uptake and lipid storage induced by HIF-1α contribute to cell growth and survival after hypoxia-reoxygenation. Cell Rep. 2014, 9, 349–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhou, Y.; Andreyev, O.; Hoyt, R.F.; Singh, A.; Hunt, T.; Horvath, K.A. Overexpression of FABP3 inhibits human bone marrow derived mesenchymal stem cell proliferation but enhances their survival in hypoxia. Exp. Cell Res. 2014, 323, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Hunt, H.E.; Sadr, K.; Deyoung, A.J.; Gortz, S.; Bugbee, W.D. The role of immunologic response in fresh osteochondral allografting of the knee. Am. J. Sports Med. 2014, 42, 886–891. [Google Scholar] [CrossRef]
- Ryan, P.M.; Turner, R.C.; Anderson, C.D.; Groth, A.T. Comparative Outcomes for the Treatment of Articular Cartilage Lesions in the Ankle with a DeNovo NT Natural Tissue Graft: Open Versus Arthroscopic Treatment. Orthop. J. Sports Med. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Mainil-Varlet, P.; Aigner, T.; Brittberg, M.; Bullough, P.; Hollander, A.; Hunziker, E.; Kandel, R.; Nehrer, S.; Pritzker, K.; Roberts, S.; et al. Histological Assessment of Cartilage Repair. J. Bone Jt. Surg. 2003, 85, 45–57. [Google Scholar] [CrossRef]
- O’Driscoll, S.W.; Keeley, F.W.; Salter, R.B. The chondrogenic potential of free autogenous periosteal grafts for biological resurfacing of major full-thickness defects in joint surfaces under the influence of continuous passive motion. An experimental investigation in the rabbit. J. Bone Jt. Surg. Am. 1986, 68, 1017–1035. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Description | LogP | Log (q-Value) | Symbols |
|---|---|---|---|---|
| GO:0030198 | extracellular matrix organization | −14.18 | −9.87 | ACAN, COL4A2, COL9A3, HAPLN1, DAG1, FOXC1, HSPG2, CCN1, ITGA6, ITGA3, LAMB3, MELTF, PDGFA, SOX9, TNXB, ITGA10, ADAMTSL2, SPINT2, ABI3BP, GREM1, NTN4, THSD4, SCUBE1, OLFML2A, DMD, GDF5, ADAMTS1, P3H2, COL10A1, MMP17, CAPNS2, CTRB2 |
| GO:0001525 | angiogenesis | −9.15 | −5.53 | COL4A2, CSPG4, DAG1, FGF1, FOXC1, HGF, HOXA3, HSPG2, CCN1, LEPR, MFGE8, NOTCH3, PDGFA, PTGIS, FZD8, APLN, ADGRG1, RAMP1, ESM1, GREM1, HIPK2, RASIP1, ISM1, WNT11, ERRFI1, ADRB2, DOK1, EFNB1, GALNT3, IGFBP2, PTGIR, SOX9, MTSS1, PLCB1, NGEF, RHBDF2, SPRY4 |
| GO:0070848 | response to growth factor | −7.29 | −4.01 | RUNX3, COL4A2, FGF1, FOXC1, GALNT3, GDF10, HGF, CCN1, ITGA3, PENK, SOX5, SOX9, GDF5, APLN, ADAMTSL2, SPINT2, DKK1, BAMBI, GREM1, DKK3, HIPK2, ERRFI1, SPRY4, INHBB |
| GO:0001503 | ossification | −6.64 | −3.50 | ACP5, ADRB2, FOXC1, GDF10, HGF, CCN1, P2RX7, PENK, SRGN, SOX9, WNT3, WNT11, GDF5, DKK1, GREM1, SMOC1, ACAN, RUNX3, HAPLN1, HOXA3, HOXD10, SOX5, SPNS2, CSPG4, LEPR |
| GO:0003002 | regionalization | −6.33 | −2.93 | EMX1, EVX1, FGF1, FOXC1, HOXA3, HOXA10, HOXD10, LFNG, WNT3, LHX3, ADGRG1, LHX2, ZEB2, SPRY1, DKK1, GREM1, FOXB1, HIPK2, HES2, GSC, SP8, EFNB1, SHROOM3 |
| GO:0048598 | embryonic morphogenesis | −5.72 | −2.71 | COL4A2, DAG1, FOXC1, HOXA3, HOXD10, CCN1, ITGA3, LAMB3, SOX9, WNT3, WNT11, GDF5, APLN, LHX2, SPINT2, DKK1, HIPK2, SHROOM3, ACAN, ADAMTS1, GREM1, HSPB7, ALPK3, SCUBE1, PDGFA |
| GO:0035272 | exocrine system development | −5.57 | −2.58 | DAG1, FOXC1, HGF, PDGFA, SOX9, NTN4, FGF1, SPINT2, RASIP1, WNT11, DKK1, GREM1, CYP19A1, HOXA3, WNT3, APLN, DKK3, PLCB1, PRR5L, PEAR1, LHX2 |
| GO:0072006 | nephron development | −5.52 | −2.55 | FGF1, FOXC1, ITGA3, NOTCH3, PDGFA, PODXL, SOX9, WNT11, MTSS1, DAG1, HGF, CCN1, WNT3, LHX2, SPINT2, DKK1, RIPK4, RASIP1, SHROOM3, NTN4, CYP19A1, ITGA6, CRLF1, ADAMTS1, ADAMTSL2, ERRFI1, PGF, SPRY1 |
| GO:2000272 | negative regulation of signaling receptor activity | −5.38 | −2.19 | DKK1, DKK3, ERRFI1, LYNX1, MTRNR2L7, MTRNR2L3, MTRNR2L8, MTRNR2L9, ADRB2, MUC4, SHANK2, GREM1 |
| GO:0043408 | regulation of mitogen-activated protein kinase cascade | −5.17 | −2.35 | ADRB2, CSPG4, DAG1, FGF1, GDF10, HGF, CCN1, INHBB, IRAK2, P2RX7, PDGFA, GDF5, FZD8, DKK1, PLCB1, GREM1, HIPK2, ERRFI1, NDRG2, SPRY4, DMD, RASIP1, PRR5L, CAMK2N2, WNT11 |
| GO:0031128 | developmental induction | −5.08 | −2.30 | FGF1, SOX9, WNT3, DKK1, HIPK2, COL4A2, DAG1, FOXC1, ITGA3, LAMB3, WNT11, APLN, ADRB2, IGFBP2, P2RX7, FZD8, SHANK2, PLCB1, BAMBI, GREM1, DKK3, NDRG2, LHX2, ANK1, HGF, HOXD10, HSPG2, NOTCH3, SOX5, RAMP1, GDF5 |
| GO:0061035 | regulation of cartilage development | −4.54 | −1.84 | CCN1, SOX5, SOX9, WNT11, GDF5, GREM1, ACAN, RUNX3, HOXA3, PDGFA, ANXA2, GDF10, SRGN, DKK1 |
| GO:0072497 | mesenchymal stem cell differentiation | −4.52 | −1.83 | SOX5, SOX9, WNT3, FOXC1, TNXB, THSD4, ADAMTSL2, ERRFI1, DKK1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toyoda, E.; Sato, M.; Takahashi, T.; Maehara, M.; Okada, E.; Wasai, S.; Iijima, H.; Nonaka, K.; Kawaguchi, Y.; Watanabe, M. Transcriptomic and Proteomic Analyses Reveal the Potential Mode of Action of Chondrocyte Sheets in Hyaline Cartilage Regeneration. Int. J. Mol. Sci. 2020, 21, 149. https://doi.org/10.3390/ijms21010149
Toyoda E, Sato M, Takahashi T, Maehara M, Okada E, Wasai S, Iijima H, Nonaka K, Kawaguchi Y, Watanabe M. Transcriptomic and Proteomic Analyses Reveal the Potential Mode of Action of Chondrocyte Sheets in Hyaline Cartilage Regeneration. International Journal of Molecular Sciences. 2020; 21(1):149. https://doi.org/10.3390/ijms21010149
Chicago/Turabian StyleToyoda, Eriko, Masato Sato, Takumi Takahashi, Miki Maehara, Eri Okada, Shiho Wasai, Hiroshi Iijima, Ken Nonaka, Yuka Kawaguchi, and Masahiko Watanabe. 2020. "Transcriptomic and Proteomic Analyses Reveal the Potential Mode of Action of Chondrocyte Sheets in Hyaline Cartilage Regeneration" International Journal of Molecular Sciences 21, no. 1: 149. https://doi.org/10.3390/ijms21010149