Carotenoids as Novel Therapeutic Molecules Against Neurodegenerative Disorders: Chemistry and Molecular Docking Analysis



Abstract
:1. Introduction
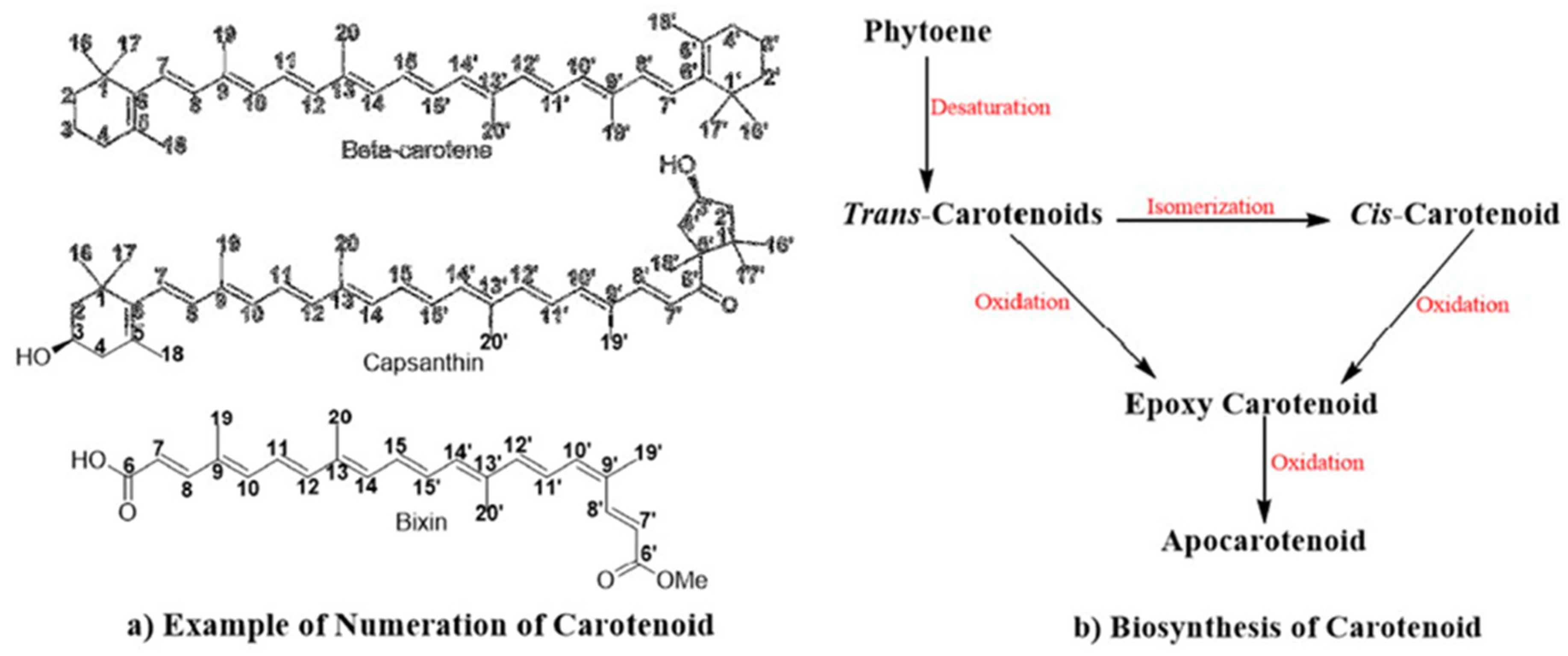
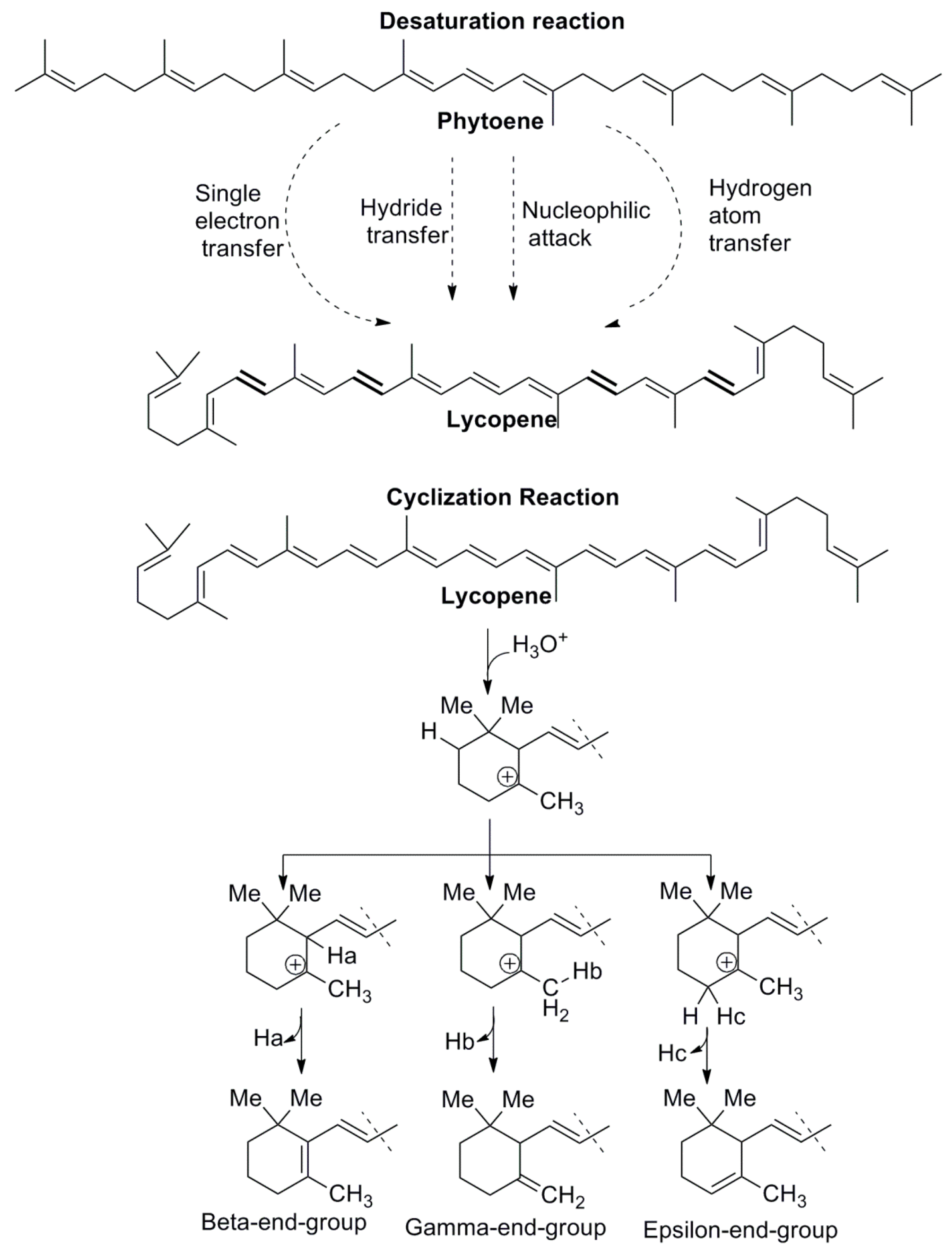
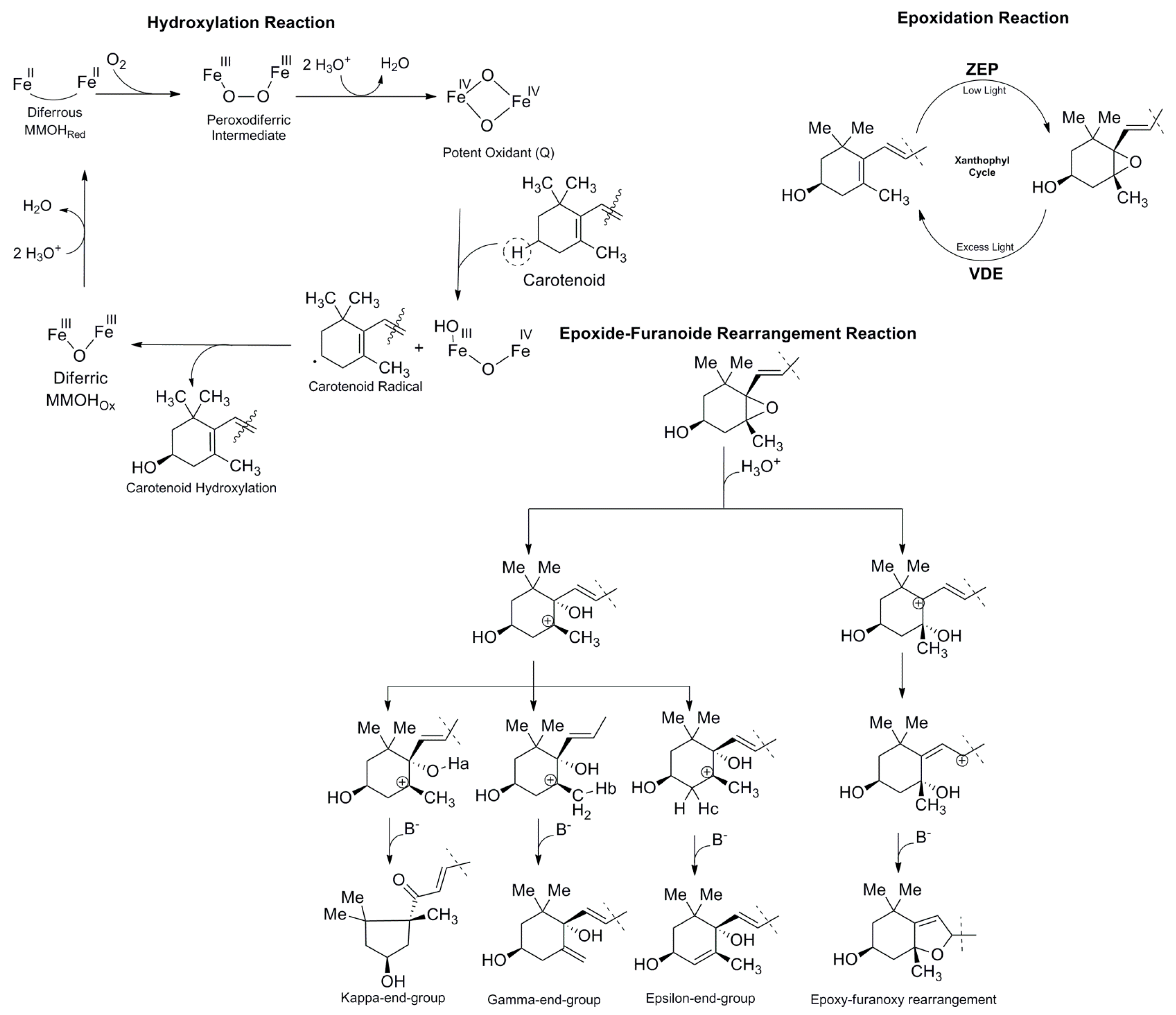
2. Chemistry of Carotenoid Compounds
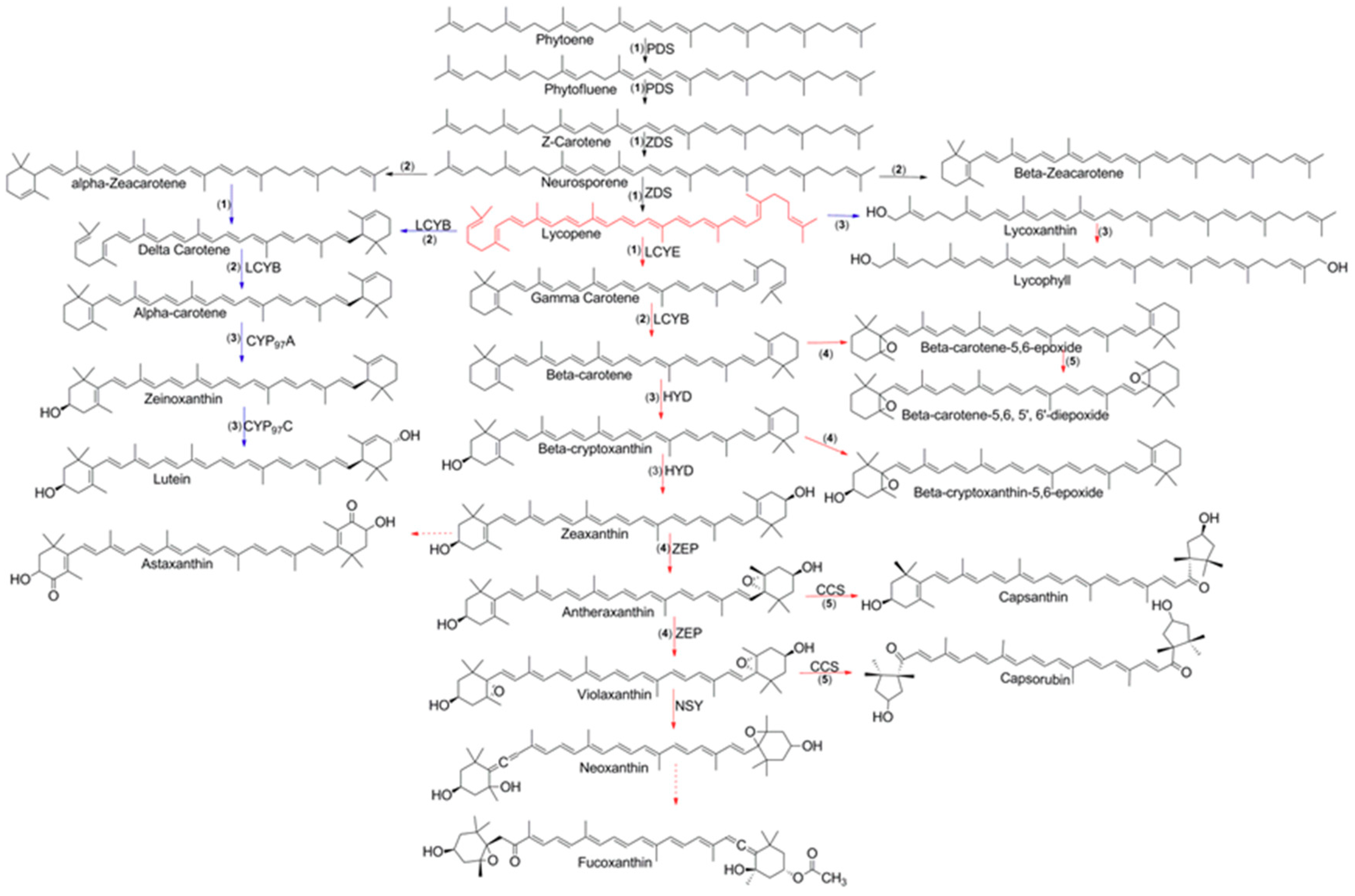
3. Biosynthesis of Carotenoids
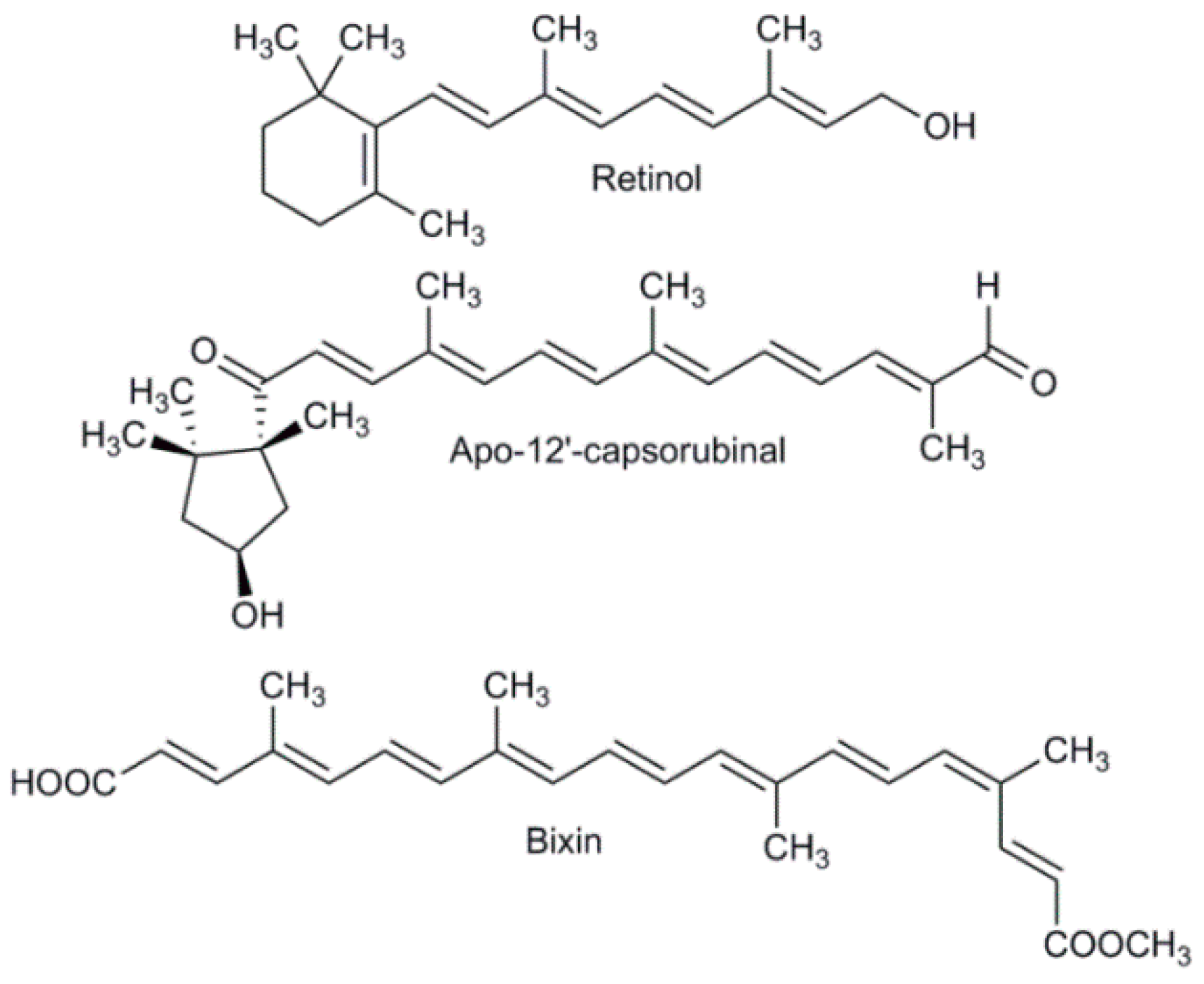
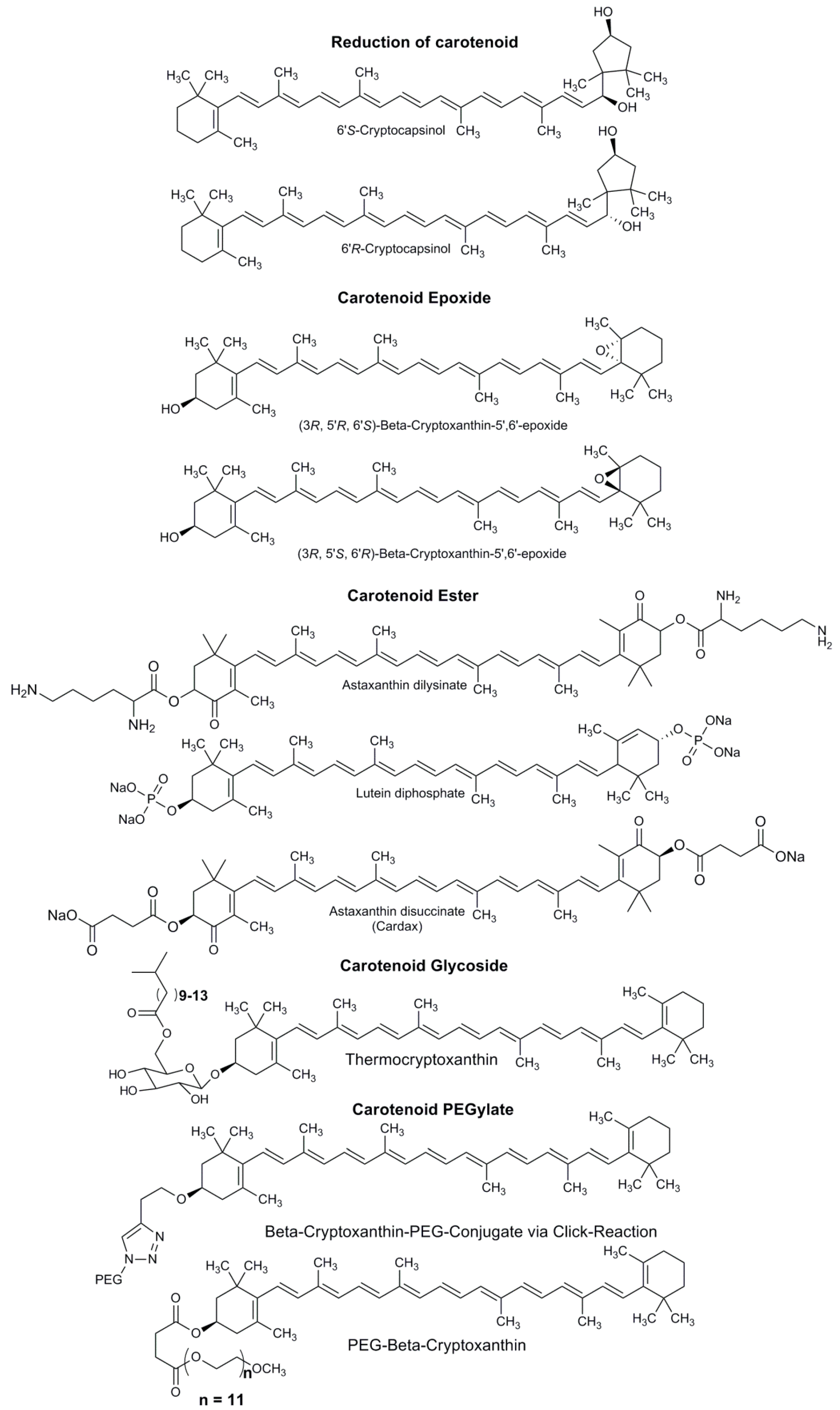
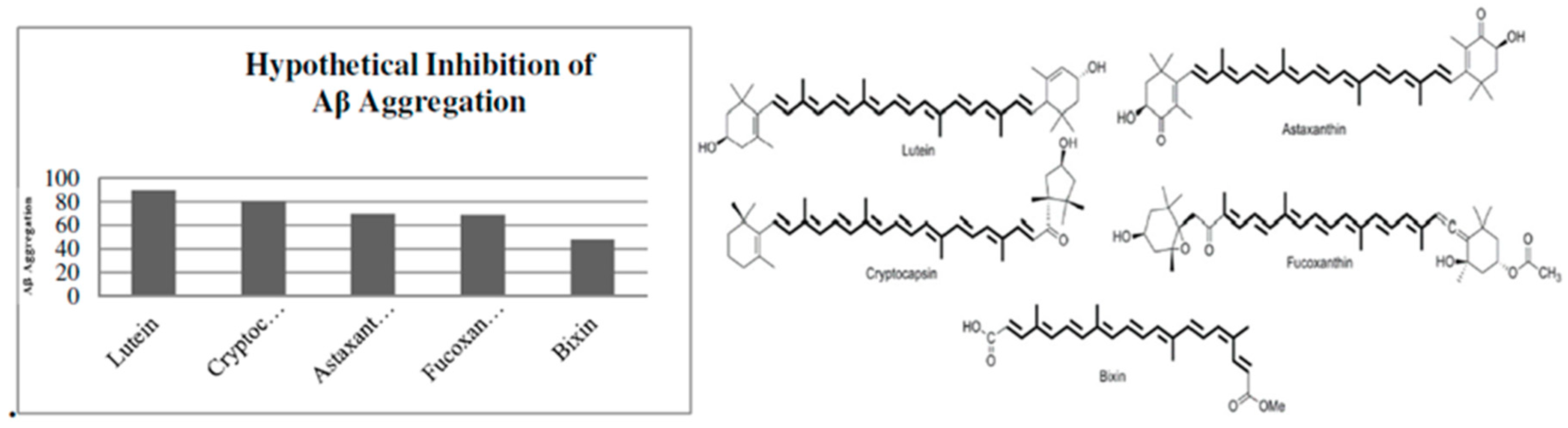
4. Types of Carotenoid Derivatives
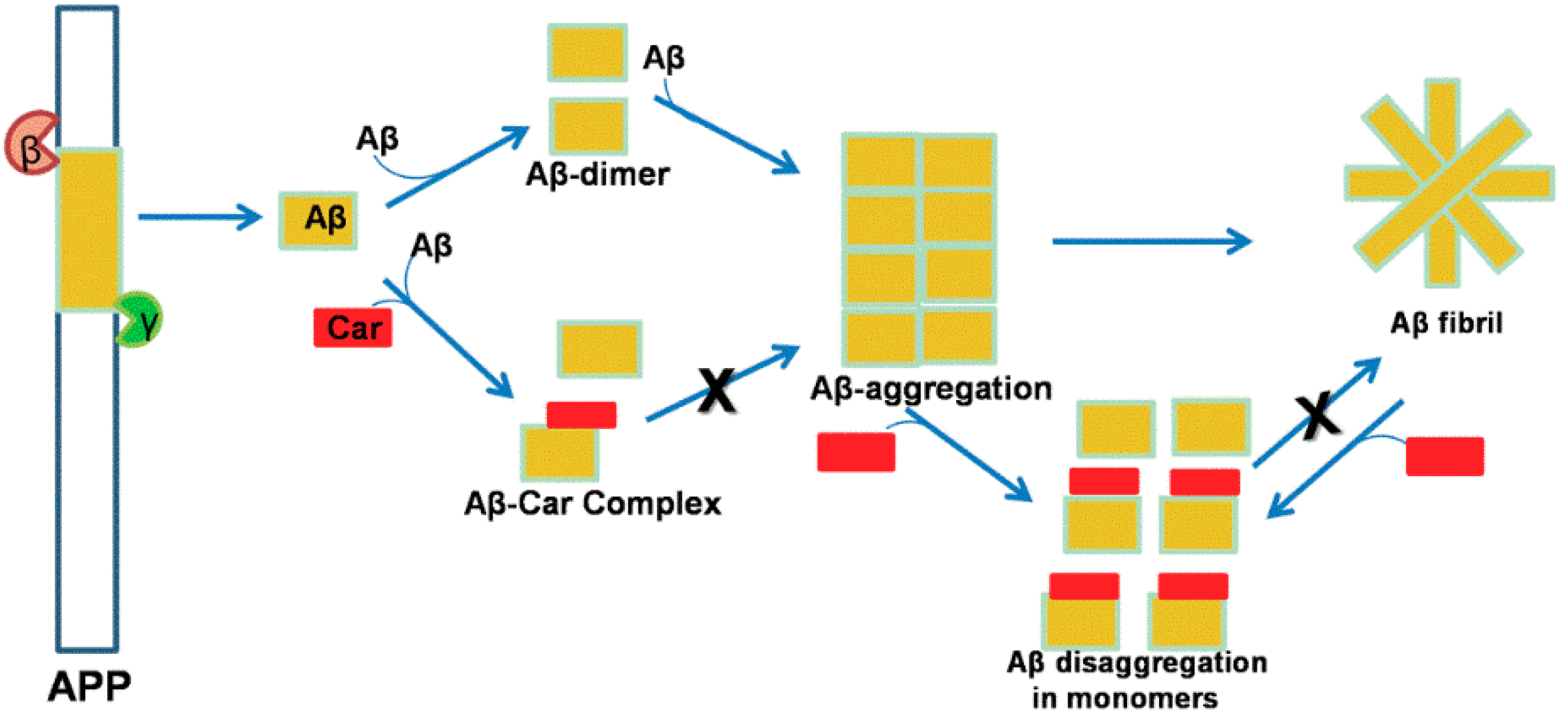
5. Neuroprotection by Carotenoids through Modulation of the App Pathway
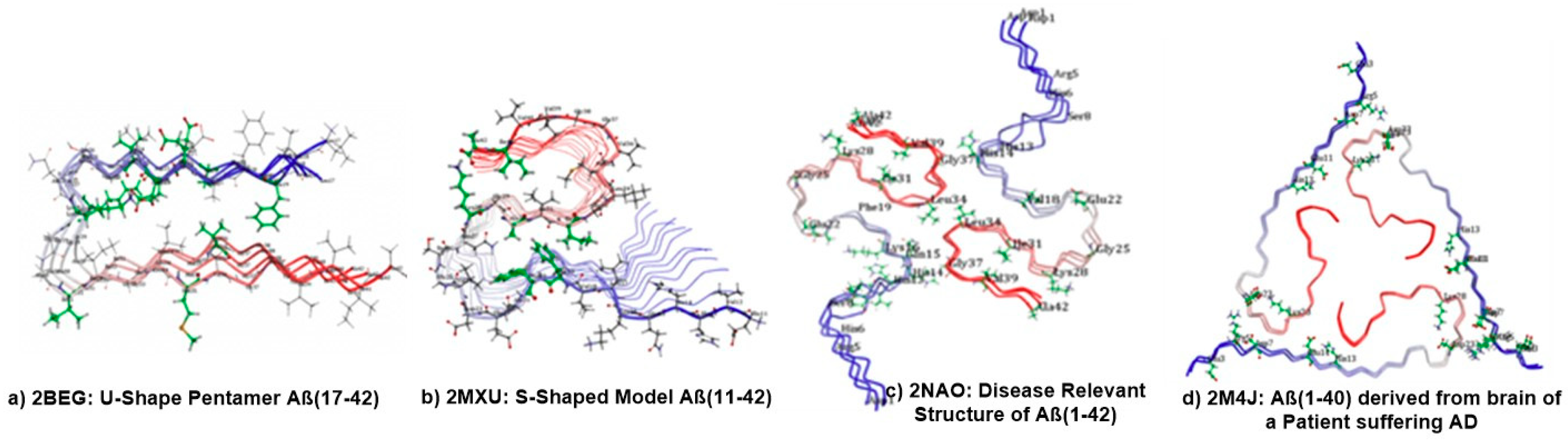
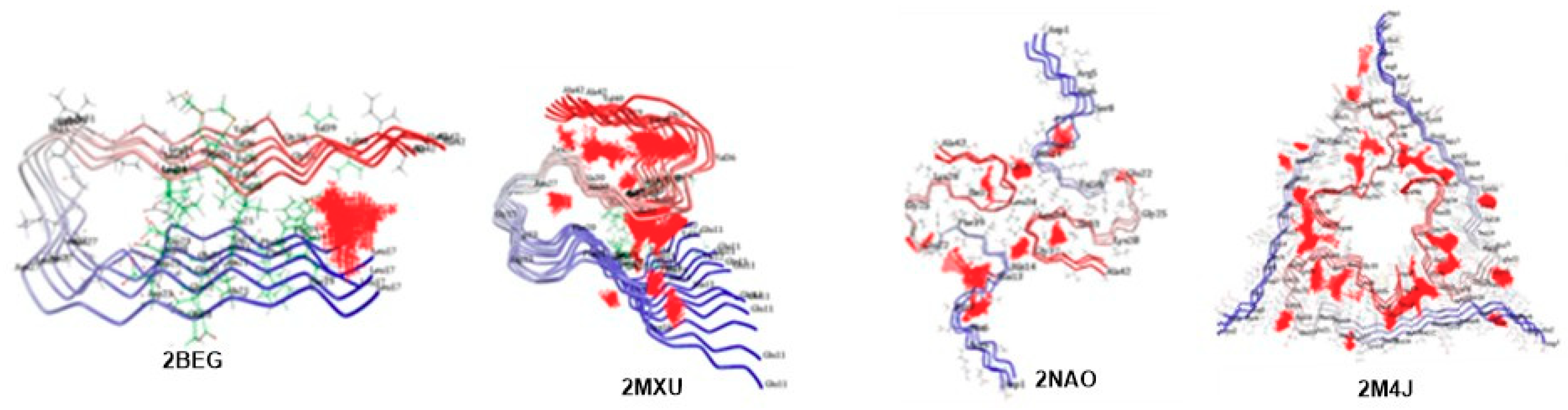
6. Molecular Docking Analysis of Carotenoid and Apocarotenoid against Aβ Aggregation
7. Aβ Aggregation and DNA Damage
8. Conclusions and New Directions in Research
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hashimoto, M.; Rockenstein, E.; Crews, L.; Masliah, E. Role of protein aggregation in mitochondrial dysfunction and neurodegeneration in Alzheimer’s and Parkinson’s diseases. Neuromol. Med. 2003, 4, 21–36. [Google Scholar] [CrossRef]
- Prado-Prado, F.; García, I. Review of theoretical studies for prediction of neurodegenerative inhibitors. Mini Rev. Med. Chem. 2012, 12, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.K.; Ryu, Y.B.; Curtis-Long, M.J.; Ryu, H.W.; Yuk, H.J.; Kim, D.W.; Kim, H.J.; Lee, W.S.; Park, K.H. Cholinestrase inhibitory effects of geranylated flavonoids from Paulownia tomentosa fruits. Bioorg. Med. Chem. 2012, 20, 2595–2602. [Google Scholar] [CrossRef] [PubMed]
- Majd, S.; Power, J.H.; Grantham, H.J.M. Neuronal response in Alzheimer’s and Parkinson’s disease: The effect of toxic proteins on intracellular pathways. BMC Neurosci. 2015, 16, 69. [Google Scholar] [CrossRef]
- Wang, S.S.; Hung, Y.T.; Wen, W.S.; Lin, K.C.; Chen, G.Y. Exploring the inhibitory activity of short-chain phospholipids against amyloid fibrillogenesis of hen egg-white lysozyme. Biochim. Biophys. Acta 2011, 1811, 301–313. [Google Scholar] [CrossRef]
- Lakey-Beitia, J.; Berrocal, R.; Rao, K.S.; Durant, A.A. Polyphenols as therapeutic molecules in Alzheimer’s disease through modulating amyloid pathways. Mol. Neurobiol. 2015, 51, 466–479. [Google Scholar] [CrossRef]
- Gadad, B.S.; Britton, G.B.; Rao, K.S. Targeting oligomers in neurodegenerative disorders: Lessons from α-synuclein, tau, and amyloid-β peptide. J. Alzheimer’s Dis. 2011, 24, 223–232. [Google Scholar] [CrossRef]
- Hegde, M.L.; Hegde, P.M.; Rao, K.S.; Mitra, S. Oxidative genome damage and its repair in neurodegenerative diseases: Function of transition metals as a double-edged sword. J. Alzheimer’s Dis. 2011, 24, 183–198. [Google Scholar] [CrossRef]
- Guerrero, E.; Padmaraju, V.; Hegde, M.L.; Britton, G.B.; Rao, K.S. Recent advances in α-synuclein functions, advanced glycation, and toxicity: Implications for Parkinson’s disease. Mol. Neurobiol. 2013, 47, 525–536. [Google Scholar] [CrossRef]
- Prakasam, A.; Muthuswamy, A.; Blonczy, Z.; Greig, N.H.; Fauq, A.; Rao, J.; Pappolla, M.A.; Sambamurti, K. Differential Accumulation of Secreted APP Metabolites in Ocular Fluids. J. Alzheimer’s Dis. 2010, 20, 1243–1253. [Google Scholar] [CrossRef]
- Suram, A.; Hegde, M.; Rao, K.S. A new evidence for DNA nicking property of amyloid b-peptide (1–42): Relevance to Alzheimer’s disease. Arch. Biochem. Biophys. 2007, 463, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Rambaran, R.N.; Serpell, L.C. Amyloid fibrils Abnormal protein assembly. Prion 2008, 2, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, B.N.; Raichurkar, K.P.; Shamasundar, N.M.; Rao, T.S.S.; Rao, K.S. Aβ(42) induced MRI changes in aged rabbit brain resembles AD brain. Neurochem. Int. 2011, 59, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef]
- Chiang, K.; Koo, E. Emerging therapeutics for Alzheimer’s disease. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 381–405. [Google Scholar] [CrossRef]
- Pillai, J.A.; Cummings, J.L. Clinical trials in predementia stages of Alzheimer disease. Med. Clin. N. Am. 2013, 97, 439–457. [Google Scholar] [CrossRef]
- Schenk, D.; Basi, G.S.; Pangalos, M.N. Treatment strategies targeting amyloid β-protein. Cold Spring Harb. Perspect. Med. 2012, 2, a006387. [Google Scholar] [CrossRef]
- Shi, M.; Caudle, W.M.; Zhang, J. Biomarker discovery in neurodegenerative diseases: A proteomic approach. Neurobiol. Dis. 2009, 35, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.P.; Khanum, F. Neuroprotective potential of phytochemicals. Pharmacogn. Rev. 2012, 6, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, H.; Tabuchi, M.; Yamaguchi, T.; Iwasaki, K.; Furukawa, K.; Sekiguchi, K.; Ikarashi, Y.; Kudo, Y.; Higuchi, M.; Saido, T.; et al. A traditional medicinal herb Paeonia suffruticosa and its active constituent 1,2,3,4,6-penta-O-galloyl-beta-D-glucopyranose have potent anti-aggregation effects on Alzheimer’s amyloid beta proteins in vitro and in vivo. J. Neurochem. 2009, 169, 1648–1657. [Google Scholar] [CrossRef]
- Ramesh, B.N.; Indi, S.S.; Rao, K.S. Anti-amyloidogenic property of leaf aqueous extract of Caesalpinia crista. Neurosci. Lett. 2010, 475, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Murillo, E.; Britton, G.B.; Durant, A.A. Antioxidant activity and polyphenol content in cultivated and wild edible fruits grown in Panama. J. Pharm. Bioallied Sci. 2012, 4, 313–317. [Google Scholar] [PubMed]
- Obulesu, M.; Dowlathabad, M.R.; Bramhachari, P.V. Carotenoids and Alzheimer’s disease: An insight into therapeutic role of retinoids in animal models. Neurochem. Int. 2011, 59, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Lakey-Beitia, J.; Gonzalez, Y.; Doens, D.; Stephens, D.E.; Santamaria, R.; Murillo, E.; Gutierrez, M.; Fernandez, P.L.; Rao, K.S.; Larionov, O.V.; et al. Assessment of Novel Curcumin Derivatives as Potent Inhibitors of Inflammation and Amyloid-Beta Aggregation in Alzheimer Disease. J. Alzheimers Dis. 2017, 60, S59–S68. [Google Scholar] [CrossRef] [PubMed]
- Lakey-Beitia, J.; Doens, D.; Kumar, D.J.; Murillo, E.; Fernandez, P.L.; Rao, K.S.; Durant-Archibold, A.A. Anti-amyloid aggregation activity of novel carotenoids: Implications for Alzheimer’s drug discovery. Clin. Aging Interv. 2017, 12, 815–822. [Google Scholar] [CrossRef]
- Parker, R.S.; Swanson, J.E.; You, C.-S.; Edwards, A.J.; Huang, T. Bioavailability of carotenoids in human subjects. Proc. Nutr. Soc. 1999, 58, 155–162. [Google Scholar] [CrossRef]
- Zern, T.L.; Fernandez, M.L. Cardioprotective Effects of Dietary Polyphenols. J. Nutr. 2005, 135, 2291–2294. [Google Scholar] [CrossRef] [Green Version]
- Rivera Vélez, S.M. Guide for Carotenoid Identification in Biological Samples. J. Nat. Prod. 2016, 79, 1473–1484. [Google Scholar] [CrossRef]
- Choi, S.K.; Osawa, A.; Maoka, T.; Hattan, J.I.; Ito, K.; Uchiyama, A.; Suzuki, M.; Shindo, K.; Misawa, N. 3-β-Glucosyl-3′-β-quinovosyl zeaxanthin, a novel carotenoid glycoside synthesized by Escherichia coli cells expressing the Pantoea ananatis carotenoid biosynthesis gene cluster. Appl. Microbiol. Biotechnol. 2013, 97, 8479–8486. [Google Scholar] [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Polotow, T.G.; Poppe, S.C.; Vardaris, C.V.; Ganini, D.; Guariroba, M.; Mattei, R.; Hatanaka, E.; Martins, M.F.; Bondan, E.F.; Barros, M.P. Redox Status and Neuro Inflammation Indexes in Cerebellum and Motor Cortex of Wistar Rats Supplemented with Natural Sources of Omega-3 Fatty Acids and Astaxanthin: Fish Oil, Krill Oil, and Algal Biomass. Mar. Drugs 2015, 13, 6117–6137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaichi, S. Carotenoids in Algae: Distributions, Biosyntheses and Functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Amorim-Carrilho, K.T.; Cepeda, A.; Fente, C.; Regal, P. Review of methods for analysis of carotenoids. Trends Anal. Chem. 2014, 56, 49–73. [Google Scholar] [CrossRef]
- Cong, L.; Wang, C.; Chen, L.; Liu, H.; Yang, G.; He, G. Expression of phytoene synthase1 and Carotene Desaturase crtI Genes Result in an Increase in the Total Carotenoids Content in Transgenic Elite Wheat (Triticum aestivum L.). J. Agric. Food Chem. 2009, 57, 8652–8660. [Google Scholar] [CrossRef] [PubMed]
- Bolhassani, A.; Khavari, A.; Bathaie, S.Z. Saffron and natural carotenoids: Biochemical activities and anti-tumor effects. Biochim. Biophys. Acta 2014, 1845, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.; Meléndez-Martínez, A.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Failla, M.L.; Huo, T.; Thakkar, S.K. In vitro screening of relative bioaccessibility of carotenoids from foods. Asia Pac. J. Clin. Nutr. 2008, 17, 200–203. [Google Scholar] [PubMed]
- Marin, E.; Nussaume, L.; Quesada, A.; Gonneau, M.; Sotta, B.; Hugueney, P.; Frey, A.; Marion-Poll, A. Molecular identification of zeaxanthin epoxidase of Nicotiana plumbaginifolia, a gene involved in abscisic acid biosynthesis and corresponding to the ABA locus of Arabidopsis thaliana. EMBO J. 1996, 15, 2331–2342. [Google Scholar] [CrossRef]
- Háda, M.; Nagy, V.; Gulyás-Fekete, G.; Deli, J.; Agócs, A. Towards Carotenoid Dendrimers: Carotenoid Diesters and Triesters with Aromatic Cores. Helv. Acta 2010, 93, 1149–1155. [Google Scholar] [CrossRef]
- Shaish, A.; Harari, A.; Kamari, Y.; Soudant, E.; Harats, D.; Ben-Amotz, A. A carotenoid algal preparation containing phytoene and phytofluene inhibited LDL oxidation in vitro. Plant Foods Hum. Nutr. 2008, 63, 83–86. [Google Scholar] [CrossRef]
- Chisté, R.C.; Freitas, M.; Mercadante, A.Z.; Fernandes, E. Carotenoids inhibit lipid peroxidation and hemoglobin oxidation, but not the depletion of glutathione induced by ROS in human erythrocytes. Life Sci. 2014, 99, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gálvez, A.; Mínguez-Mosquera, M.I. Esterification of xanthophylls and its effect on chemical behavior and bioavailability of carotenoids in the human. Nutr. Res. 2005, 25, 631–640. [Google Scholar] [CrossRef]
- Papandreou, M.A.; Kanakis, C.D.; Polissiou, M.G.; Efthimiopoulos, S.; Cordopatis, P.; Margarity, M.; Lamari, F.N. Inhibitory Activity on Amyloid-Beta Aggregation and Antioxidant Properties of Crocus sativus Stigmas Extract and Its Crocin Constituents. J. Agric. Food Chem. 2006, 54, 8762–8768. [Google Scholar] [CrossRef]
- Firdous, A.P.; Preethi, K.C.; Kuttan, R. Antioxidant potential of meso-zeaxanthin a semi synthetic carotenoid. Food Chem. 2010, 119, 1096–1101. [Google Scholar] [CrossRef]
- Sachdeva, A.K.; Chopra, K. Lycopene abrogates Aβ(1-42)-mediated neuroinflammatory cascade in an experimental model of Alzheimer’s disease. J. Nutr. Biochem. 2015, 26, 736–744. [Google Scholar] [CrossRef]
- Britton, G. Functions of Intact Carotenoids. In Carotenoids; Birkhäuser: Basel, Switzerland, 2008; Volume 4, pp. 189–212. [Google Scholar]
- Kopsell, D.A.; Kopsell, D.E. Accumulation and bioavailability of dietary carotenoids in vegetable crops. Trends Plant Sci. 2006, 11, 499–507. [Google Scholar] [CrossRef]
- Hirschberg, J. Carotenoid biosynthesis in flowering plants. Curr. Opin. Plant Biol. 2001, 4, 210–218. [Google Scholar] [CrossRef]
- Chemler, J.A.; Yan, Y.; Koffas, M.A.G. Biosynthesis of isoprenoids, polyunsaturated fatty acids and flavonoids in Saccharomyces cerevisiae. Microb. Cell Fact. 2006, 5, 20. [Google Scholar] [CrossRef]
- Werner, S.; Böhm, V. Bioaccessibility of carotenoids and vitamin e from pasta: Evaluation of an in vitro digestion model. J. Agric. Food Chem. 2011, 59, 1163–1170. [Google Scholar] [CrossRef]
- Wolak, T.; Hamias, R.; Volvich, L.; Paran, E. Carotenoids and flavonoids synergistically attenuate inflammation in endothelial cells induced by TNF-α. J. Am. Soc. Hypertens. 2014, 8, e77. [Google Scholar] [CrossRef]
- Britton, G. Structure and properties of carotenoids in relation to function. FASEB J. 1995, 15, 1551–1558. [Google Scholar] [CrossRef]
- Britton, G.; Khachik, F. Carotenoids in Food. In Carotenoids; Birkhäuser: Basel, Switzerland, 2009; Volume 5, pp. 45–66. [Google Scholar]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Natural Functions. In Carotenoids; Birkhäuser: Basel, Switzerland, 2008; pp. 1–370. [Google Scholar]
- Rodriguez-Amaya, D.B. A Guide to carotenoid Analysis in Foods; ILSI Press: Washington, DC, USA, 2001; pp. 1–64. [Google Scholar]
- Katayama, S.; Ogawa, H.; Nakamura, S. Apricot carotenoids possess potent anti-amyloidogenic activity in vitro. J. Agric. Food Chem. 2011, 59, 12691–12696. [Google Scholar] [CrossRef] [PubMed]
- Faulks, R.M.; Southon, S. Challenges to understanding and measuring carotenoid bioavailability. Biochim. Biophys. Acta 2005, 1740, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vershinin, A. Biological functions of carotenoids-diversity and evolution. Biofactors 1999, 10, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Hornero-Méndez, D.; Gómez-Ladrón De Guevara, R.; Mínguez-Mosquera, M.I. Carotenoid biosynthesis changes in five red pepper (Capsicum annuum L.) cultivars during ripening. Cultivar selection for breeding. J. Agric. Food Chem. 2000, 48, 3857–3864. [Google Scholar] [CrossRef] [PubMed]
- Maiani, G.; Castón, M.J.P.; Catasta, G.; Toti, E.; Cambrodón, I.G.; Bysted, A.; Granado-Lorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M.; et al. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food Res. 2009, 53, S194–218. [Google Scholar] [CrossRef]
- Murillo, E.; Giuffrida, D.; Menchaca, D.; Dugo, P.; Torre, G.; Meléndez-Martinez, A.J.; Mondello, L. Native carotenoids composition of some tropical fruits. Food Chem. 2013, 140, 825–836. [Google Scholar] [CrossRef]
- Che, H.; Li, Q.; Zhang, T.; Wang, D.; Yang, L.; Xu, J.; Yanagita, T.; Xue, C.; Chang, Y.; Wang, Y. The effects of astaxanthin and docosahexaenoic acid-acylated astaxanthin on Alzheimer’s disease in APP/PS1 double transgenic mice. J. Agric. Food Chem. 2018, 66, 4948–4957. [Google Scholar] [CrossRef]
- Moise, A.R.; Al-Babili, S.; Wurtzel, E.T. Mechanistic aspects of carotenoid biosynthesis. Chem. Rev. 2014, 114, 164–193. [Google Scholar] [CrossRef]
- Kita, M.; Kato, M.; Ban, Y.; Honda, C.; Yaegaki, H.; Ikoma, Y.; Moriguchi, T. Carotenoid accumulation in Japanese apricot (Prunus mume Siebold & Zucc.): Molecular analysis of carotenogenic gene expression and ethylene regulation. J. Agric. Food Chem. 2007, 55, 3414–3420. [Google Scholar]
- Gulyás-Fekete, G.; Murillo, E.; Kurtán, T.; Papp, T.; Illyés, T.-Z.; Drahos, L.; Visy, J.; Agócs, A.; Turcsi, E.; Deli, J. Cryptocapsinepoxide-type carotenoids from red mamey, Pouteria sapota. J. Nat. Prod. 2013, 76, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Scotter, M.J.; Wilson, L.A.; Appleton, G.P.; Castle, L. Analysis of Annatto (Bixa orellana) Food Coloring Formulations. Determination of Aromatic Hydrocarbon Thermal Degradation Products by Gas Chromatography. J. Agric. Food Chem. 2000, 48, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Murillo, E.; Nagy, V.; Agócs, A.; Deli, J. Carotenoids with κ -End Group. In Carotenoids; Nova Science Publishers: New York, NY, USA, 2013; pp. 1–78. [Google Scholar]
- Gaweska, H.; Fitzpatrick, P.F. Structures and Mechanism of the Monoamine Oxidase Family. Biomol. Concepts 2011, 2, 365–377. [Google Scholar] [CrossRef]
- Deli, J.; Molnár, P.; Matus, Z.; Tóth, G. Carotenoid composition in the fruits of red paprika (Capsicum annuum var. lycopersiciforme rubrum) during ripening; biosynthesis of carotenoids in Red Paprika. J. Agric. Food Chem. 2001, 49, 1517–1523. [Google Scholar] [CrossRef]
- Wallar, B.J.; Lipscomb, J.D. Dioxygen Activation by Enzymes Containing Binuclear Non-Heme Iron Clusters. Chem. Rev. 1996, 96, 612–625. [Google Scholar] [CrossRef]
- Baik, M.; Newcomb, M.; Friesner, R.A.; Lippard, S.J. Mechanistic Studies on the Hydroxylation of Methane by Methane Monooxygenase. Chem. Rev. 2003, 103, 2385–2419. [Google Scholar] [CrossRef]
- Yun, D.; Garcia-Serres, R.; Chicalese, B.M.; An, Y.H.; Huynh, B.H.; Bollinger, J.M. (μ-1,2-Peroxo)diiron(III/III) Complex as a Precursor to the Diiron (III/IV) Intermediate X in the Assembly of the Iron-radical Cofactor of Ribonucleotide Reductase from Mouse. Biochemistry 2007, 46, 1925–1932. [Google Scholar] [CrossRef]
- Demming-Adams, B.; Adams, W.W., III. The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends Plant Sci. 1996, 1, 21–26. [Google Scholar] [CrossRef]
- Háda, M.; Petrovics, D.; Nagy, V.; Böddi, K.; Deli, J.; Agócs, A. The first synthesis of PEG–carotenoid conjugates. Tetrahedron Lett. 2011, 52, 3195–3197. [Google Scholar] [CrossRef]
- Háda, M.; Nagy, V.; Deli, J.; Agócs, A. Hydrophilic carotenoids: Recent progress. Molecules 2012, 17, 5003–5012. [Google Scholar] [CrossRef]
- Deli, J.; Ősz, E.; Molnár, P.; Zsila, F.; Simonyi, M.; Tóth, G. Reduction of Capsorubin and Cryptocapsin. Helv. Chim. Acta 2001, 84, 3810–3817. [Google Scholar] [CrossRef]
- Turcsi, E.; Murillo, E.; Kurtán, T.; Szappanos, Á.; Illyés, T.-Z.; Gulyás-Fekete, G.; Agócs, A.; Avar, P.; Deli, J. Isolation of β-Cryptoxanthin-epoxides, Precursors of Cryptocapsin and 3′-Deoxycapsanthin, from Red Mamey (Pouteria sapota). J. Agric. Food Chem. 2015, 63, 6059–6065. [Google Scholar] [CrossRef] [PubMed]
- Molnar, P.; Deli, J.; Matma, Z.; Totha, G.; Steckb, A.; Pfanderb, H. Partial Synthesis and Characterization of the Mono-and Diepoxides of p-Cryptoxanthin. Helv. Chim. Acta 1997, 80, 221–229. [Google Scholar] [CrossRef]
- Cardounel, A.J.; Dumitrescu, C.; Zweier, J.L.; Lockwood, S.F. Direct superoxide anion scavenging by a disodium disuccinate astaxanthin derivative: Relative efficacy of individual stereoisomers versus the statistical mixture of stereoisomers by electron paramagnetic resonance imaging. Biochem. Biophys. Res. Commun. 2003, 307, 704–712. [Google Scholar] [CrossRef]
- Frey, D.A.; Kataisto, E.W.; Ekmanis, J.L.; O’Malley, S.; Lockwood, S.F. The Efficient Synthesis of Disodium Disuccinate Astaxanthin (Cardax). Org. Process Res. Dev. 2004, 8, 796–801. [Google Scholar] [CrossRef]
- Háda, M.; Nagy, V.; Takátsy, A.; Deli, J.; Agócs, A. Dicarotenoid esters of bivalent acids. Tetrahedron Lett. 2008, 49, 3524–3526. [Google Scholar] [CrossRef]
- Nadolski, G.; Cardounel, A.J.; Zweier, L.; Lockwood, S.F. The synthesis and aqueous superoxide anion scavenging of water-dispersible lutein esters. Bioorg. Med. Chem. Lett. 2006, 16, 775–781. [Google Scholar] [CrossRef]
- Jackson, H.L.; Cardounel, A.J.; Zweier, L.; Lockwood, S.F. Synthesis, characterization, and direct aqueous superoxide anion scavenging of a highly water-dispersible astaxanthin-amino acid conjugate. Bioorg. Med. Chem. Lett. 2004, 14, 3985–3991. [Google Scholar] [CrossRef]
- Yamano, Y.; Sakai, Y.; Hara, M.; Ito, M. Carotenoids and related polyenes. Part 9. 1 Total synthesis of thermozeaxanthin and thermocryptoxanthin and the stabilizing effect of thermozeaxanthin on liposomes. J. Chem. Soc. Perkin Trans. 1 2013, 2, 2006–2013. [Google Scholar]
- Neises, V.B.; Steglich, W. Einfaches Verfahren zur Veresterung von Carbonsäuren. Angew. Chem. 1978, 90, 556–557. [Google Scholar] [CrossRef]
- Shi, W.; Dolai, S.; Rizk, S.; Hussain, A.; Tariq, H.; Averick, S.; L’Amoreaux, W.; El Ldrissi, A.; Banerjee, P.; Raja, K. Synthesis of monofunctional curcumin derivatives, clicked curcumin dimer, and a PAMAM dendrimer curcumin conjugate for therapeutic applications. Org. Lett. 2007, 9, 5461–5464. [Google Scholar] [CrossRef] [PubMed]
- Convertino, M.; Pellarin, R.; Catto, M.; Carotti, A.; Caflisch, A. 9,10-Anthraquinone hinders beta β-aggregation: How does a small molecule interfere with Aβ-peptide amyloid fibrillation? Protein Sci. 2009, 18, 792–800. [Google Scholar] [PubMed]
- Shih, P.H.; Wu, C.H.; Yeh, C.T.; Yen, G.C. Protective effects of anthocyanins against amyloid β-peptide-induced damage in neuro-2A cells. J. Agric. Food Chem. 2011, 59, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Esler, W.P.; Wolfe, M.S. A portrait of Alzheimer secretases-new features and familiar faces. Science 2001, 293, 1449–1454. [Google Scholar] [CrossRef]
- Zhou, Y.; Suram, A.; Venugopal, C.; Prakasam, A.; Lin, S.; Su, Y.; Li, B.; Paul, S.M.; Sambamurti, K. Geranylgeranyl pyrophosphate stimulates gamma-secretase to increase the generation of Abeta and APP-CTFgamma. FASEB J. 2008, 22, 47–54. [Google Scholar] [CrossRef]
- Hamley, I.W. The amyloid beta peptide: A chemist’s perspective. Role in Alzheimer’s and fibrillization. Chem. Rev. 2012, 112, 5147–5192. [Google Scholar] [CrossRef]
- Venugopal, C.; Demos, C.M.; Rao, K.S.; Pappolla, M.A. Beta-secretase: Structure, function, and evolution. CNS Neurol. Disord. Drug Targets 2008, 7, 278–294. [Google Scholar] [CrossRef]
- Rajendran, L.; Schneider, A.; Schlechtingen, G.; Weidlich, S.; Ries, J.; Braxmeier, T.; Schwille, P.; Schulz, J.B.; Schroeder, C.; Simons, M.; et al. Efficient inhibition of the Alzheimer’s disease β-secretase by membrane targeting. Science 2008, 320, 520–523. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Wang, Z.; Cui, Y.; Tan, X.; Yuan, T.; Liu, Q.; Liu, Z.; Liu, X. Supplementation of lycopene attenuates lipopolysaccharide-induced amyloidogenesis and cognitive impairments via mediating neuroinflammation and oxidative stress. J. Nutr. Biochem. 2018, 56, 16–25. [Google Scholar] [CrossRef]
- Reboul, E.; Borel, P.; Abou, L.; Charbonnier, M.; Caris-Veyrat, C.; Goupy, P.; Lairon, D. Human Nutrition and Metabolism Enrichment of Tomato Paste with 6% Tomato Peel Increases Lycopene and Beta-Carotene Bioavailability in Men. J. Nutr. 2005, 135, 790–794. [Google Scholar] [CrossRef]
- Cho, K.S.; Shin, M.; Kim, S.; Lee, S.B. Recent Advances in Studies on the Therapeutic Potential of Dietary Carotenoids in Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2018, 2018, 4120458. [Google Scholar] [CrossRef] [PubMed]
- Ghahghaei, A.; Bathaie, S.Z.; Kheirkhah, H.; Bahraminejad, E. The protective effect of crocin on the amyloid fibril formation of Aβ42 peptide in vitro. Cell. Mol. Biol. Lett. 2013, 18, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Lee, Y.J.; Kwon, K.H. Neuroprotective Effects of Astaxanthin in Oxygen-Glucose Deprivation in SH-SY5Y Cells and Global Cerebral Ischemia in Rat. J. Clin. Biochem. Nutr. 2010, 47, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wu, H.; Wen, H.; Fang, H.; Hong, Z.; Yi, R.; Liu, R. Simultaneous Determination of Fucoxanthin and Its Deacetylated Metabolite Fucoxanthinol in Rat Plasma by Liquid Chromatography-Tandem Mass Spectrometry. Mar. Drugs 2015, 13, 6521–6536. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Liu, F.; Lin, J.; Chen, H.; Huang, C.; Chen, L.; Zhou, Y.; Ye, L.; Zhang, K.; Jin, J.; et al. Fucoxanthin Inhibits β-Amyloid Assembly and Attenuates β-Amyloid Oligomer-Induced Cognitive Impairments. J. Agric. Food Chem. 2017, 65, 4092–4102. [Google Scholar] [CrossRef]
- Jung, H.; Ali, M.; Choi, R.; Jeong, H.; Chung, H.; Choi, J. Kinetics and molecular docking studies of fucosterol and fucoxanthin, BACE1 inhibitors from brown algae Undaria pinnatifida and Ecklonia stolonifera. Food Chem. Toxicol. 2016, 89, 104–111. [Google Scholar] [CrossRef]
- Murillo, E.; Mclean, R.; Britton, G.; Agócs, A.; Nagy, V.; Deli, J. Sapotexanthin, an A-Provitamin Carotenoid from Red Mamey (Pouteria sapota). J. Nat. Prod. 2011, 74, 283–285. [Google Scholar] [CrossRef]
- Chacón-Ordóñez, T.; Esquivel, P.; Jiménez, V.M.; Carle, R.; Schweiggert, R.M. Deposition Form and Bioaccessibility of Keto-carotenoids from Mamey Sapote (Pouteria sapota), Red Bell Pepper (Capsicum annuum), and Sockeye Salmon (Oncorhynchus nerka) Filet. J. Agric. Food Chem. 2016, 64, 1989–1998. [Google Scholar] [CrossRef]
- Miyake, S.; Kobayashi, S.; Tsubota, K.; Ozawa, Y. Phase II enzyme induction by a carotenoid, lutein, in a PC12D neuronal cell line. Biochem. Biophys. Res. Commun. 2014, 446, 535–540. [Google Scholar] [CrossRef] [Green Version]
- Nishino, A.; Yasui, H.; Maoka, T. Reaction of Paprika Carotenoids, Capsanthin and Capsorubin, with Reactive Oxygen Species. J. Agric. Food Chem. 2016, 64, 4786–4792. [Google Scholar] [CrossRef]
- Nakagawa, K.; Kiko, T.; Miyazawa, T.; Sookwong, P.; Tsuduki, T.; Satoh, A.; Miyazawa, T. Amyloid β-induced erythrocytic damage and its attenuation by carotenoids. FEBS Lett. 2011, 585, 1249–1254. [Google Scholar] [CrossRef] [PubMed]
- Maksimov, G.V.; Volkov, V.V.; Parshina, E.Y.; Akhalaia, M.I.; Kozlova, O.V.; Derinskaya, E.V.; Revin, V.V.; Rubin, A.B. Investigation of carotenoid conformations in myelin nerve upon changes in oxygen content. Biochem. Biophys. Mol. Biol. 2007, 417, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Thiessen, P.; Bolton, E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Berman, H.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.; Weissig, H.; Shindyalov, I.; Bourne, P. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Momany, F.A.; Rone, R. Validation of the general purpose QUANTA® 3.2/CHARMm® force field. J. Comput. Chem. 1992, 13, 888–900. [Google Scholar] [CrossRef]
- Tjernberg, L.O.; Na, J.; Lindqvist, F.; Johansson, J.; Karlstro, A.R.; Thyberg, J. Arrest of Beta-Amyloid Fibril Formation by a Pentapeptide Ligand. J. Biol. Chem. 1996, 271, 8545–8548. [Google Scholar] [CrossRef]
- Kumar, J.; Namsechi, R.; Sim, V.L. Structure-Based Peptide Design to Modulate Amyloid Beta Aggregation and Reduce Cytotoxicity. PLoS ONE 2015, 12, e0129087. [Google Scholar] [CrossRef]
- Hetényi, C.; Körtvélyesi, T.; Penke, B. Mapping of Possible Binding Sequences of Two Beta-Sheet Breaker Peptides on Beta Amyloid Peptide of Alzheimer’s Disease. Bioorg. Med. Chem. 2002, 10, 1587–1593. [Google Scholar] [CrossRef]
- Kepp, K.P. Alzheimer’s disease: How metal ions define β-amyloid function. Coord. Chem. Rev. 2017, 351, 127–159. [Google Scholar] [CrossRef]
- Azqueta, A.; Collins, A.R. Carotenoids and DNA damage. Mutat. Res. Mol. Mech. Mutagen. 2012, 733, 4–13. [Google Scholar] [CrossRef]
- Torbergsen, A.C.; Collins, A.R. Recovery of human lymphocytes from oxidative DNA damage; the apparent enhancement of DNA repair by carotenoids is probably simply an antioxidant effect. Eur. J. Nutr. 2000, 39, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Aldini, G.; Johnson, E.J.; Rasmussen, H.; Kraemer, K.; Woolf, H.; Musaeus, N.; Krinsky, N.I.; Russell, R.M.; Yeum, K. Modification of lymphocyte DNA damage by carotenoid supplementation in postmenopausal women 1-4. Am. J. Clin. Nutr. 2018, 83, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Serpeloni, J.M.; Mazzaron, G.; Friedmann, J.; Zerlotti, A.; Lourdes, M.; Bianchi, P.; Pires, M.; Greggi, L. Dietary carotenoid lutein protects against DNA damage and alterations of the redox status induced by cisplatin in human derived HepG2 cells. Toxicol. In Vitro 2012, 26, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Santocono, M.; Zurria, M.; Berrettini, M.; Fedeli, D.; Falcioni, G. Influence of astaxanthin, zeaxanthin and lutein on DNA damage and repair in UVA-irradiated cells. J. Photochem. Photobiol. B Biol. 2006, 85, 205–215. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligands | Highest -CDOCKER Energy (kcal/mol) | Highest -CDOCKER Interaction Energy (kcal/mol) | Amino Acid Residues Interacting with Aβ Chains | Common Residues Interacting Across all Ligands |
|---|---|---|---|---|
| Astaxanthin | −83.0351 | −17.3682 | Gly38, Val39 | Ala21, Glu22 *, Val24 *, Gly25 *, Asp23 #, Ser26 #, Lys28 #, Ile32, Gly33; Leu34 *, Gly38, Val39 |
| Bixin | −85.044 | −127.336 | Glu22 * | |
| Cryptocapsin | −83.6518 | −14.4395 | Leu34 *, Ile32, Gly33, Gly33 | |
| Fucoxanthin | −69.4873 | −39.3977 | Asp23 #, Glu22, Ala21 | |
| Lutein | −97.6659 | −20.8785 | Asp23 *, Val24 *,#, Gly25 *,#, Lys28 #, Ser26 # |
| Ligands | Highest -CDOCKER Energy (kcal/mol) | Highest -CDOCKER Interaction Energy (kcal/mol) | Amino Acid Residues Interacting with Aβ Chains | Common Residues Interacting Across all Ligands |
|---|---|---|---|---|
| Astaxanthin | −36.1994 | −60.0733 | Val2, His14 | Val2, His14 Ile32, Gly33, Leu34 |
| Bixin | 10.2499 | −45.5219 | Leu34, Gly33 | |
| Cryptocapsin | −44.9219 | −60.0403 | Gly33, Ile32, Gly33 | |
| Fucoxanthin | −40.2796 | −60.6453 | Leu34, His14 | |
| Lutein | −60.6453 | −50.7407 | His14 |
| Ligands | Highest -CDOCKER Energy (kcal/mol) | Highest -CDOCKER Interaction Energy (kcal/mol) | Amino Acid Residues Interacting with Aβ Chains | Common Residues Interacting Across all Ligands |
|---|---|---|---|---|
| Astaxanthin | −76.0481 | −29.5232 | Ser8, Glu11, Gly9 Tyr10 | Ser8, Glu11, Lys16 |
| Bixin | −74.169 | −104.431 | Glu22, Glu11 | |
| Cryptocapsin | −84.3430 | −33.051 | Lys16, Glu18 | |
| Fucoxanthin | −75.0944 | −37.7015 | Lys16, His13 | |
| Lutein | −88.1515 | −30.5548 | Ser8 |
| Ligands | Highest -CDOCKER Energy (kcal/mol) | Highest -CDOCKER Interaction Energy (kcal/mol) | Amino Acid Residues Interacting with amyloid-β chains |
|---|---|---|---|
| Astaxanthin | −82.9702 | −18.1365 | - |
| Bixin | −22.6447 | −16.4999 | Lys28 |
| Cryptocapsin | −101.782 | −14.0123 | Gly37 |
| Fucoxanthin | −89.3804 | −18.9031 | Gly37 |
| Lutein | −110.897 | −16.1726 | Gly37 |
| Ligands | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 16 | 18 | 21 | 22 | 23 | 24 | 25 | 26 | 28 | 32 | 33 | 34 | 37 | 38 | 39 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S | G | Y | E | V | H | H | K | V | A | E | D | V | G | S | K | I | G | L | G | G | V | |
| Astaxanthin | ||||||||||||||||||||||
| Bixin | ||||||||||||||||||||||
| Cryptocapsin | ||||||||||||||||||||||
| Fucoxanthin | ||||||||||||||||||||||
| Lutein |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lakey-Beitia, J.; Kumar D., J.; Hegde, M.L.; Rao, K.S. Carotenoids as Novel Therapeutic Molecules Against Neurodegenerative Disorders: Chemistry and Molecular Docking Analysis. Int. J. Mol. Sci. 2019, 20, 5553. https://doi.org/10.3390/ijms20225553
Lakey-Beitia J, Kumar D. J, Hegde ML, Rao KS. Carotenoids as Novel Therapeutic Molecules Against Neurodegenerative Disorders: Chemistry and Molecular Docking Analysis. International Journal of Molecular Sciences. 2019; 20(22):5553. https://doi.org/10.3390/ijms20225553
Chicago/Turabian StyleLakey-Beitia, Johant, Jagadeesh Kumar D., Muralidhar L. Hegde, and K.S. Rao. 2019. "Carotenoids as Novel Therapeutic Molecules Against Neurodegenerative Disorders: Chemistry and Molecular Docking Analysis" International Journal of Molecular Sciences 20, no. 22: 5553. https://doi.org/10.3390/ijms20225553