Optimization of Mature Embryo-Based Tissue Culture and Agrobacterium-Mediated Transformation in Model Grass Brachypodium distachyon


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
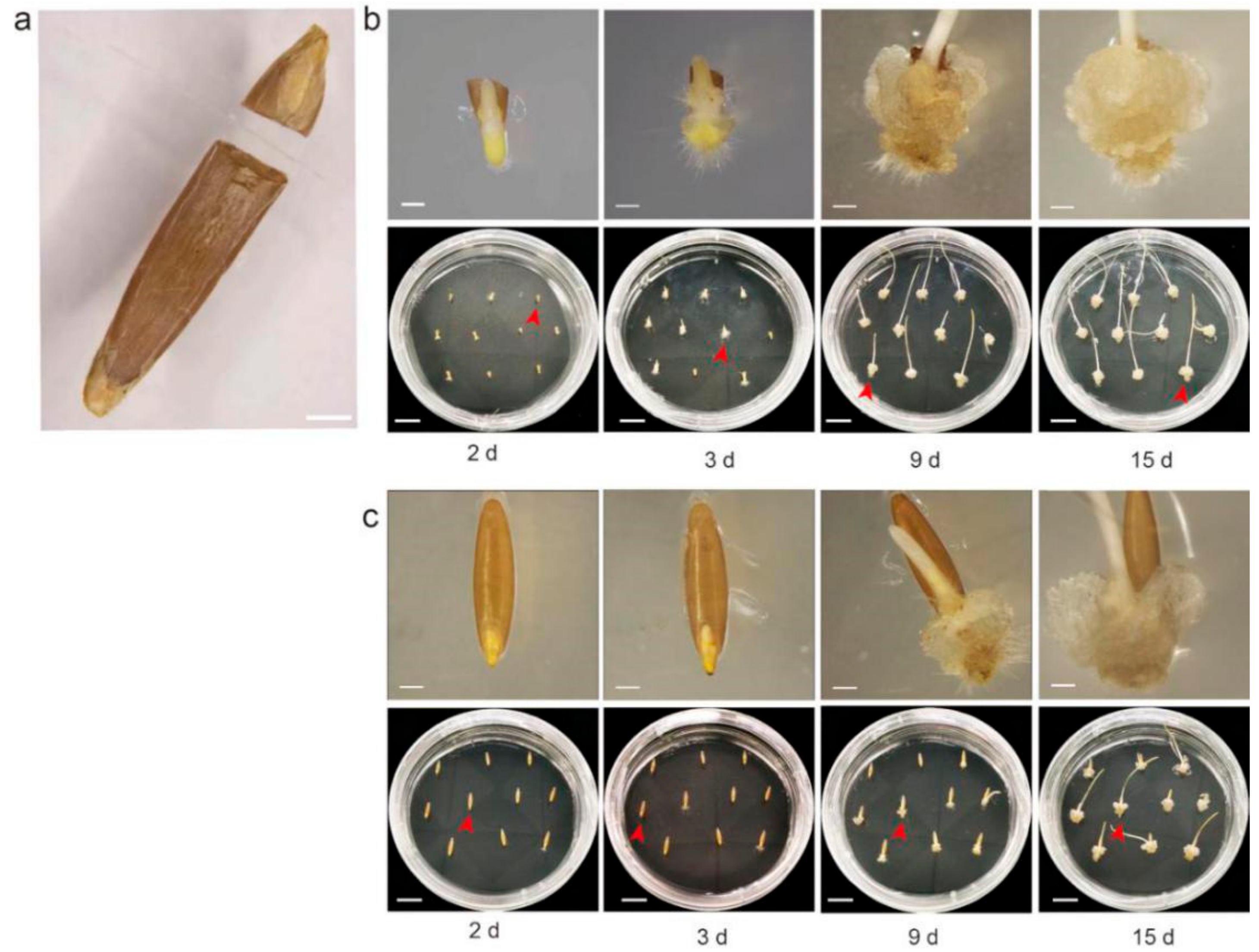
2.1. Shearing Promotes Callus Formation by Breaking Dormancy of Bd21 Mature Embryo
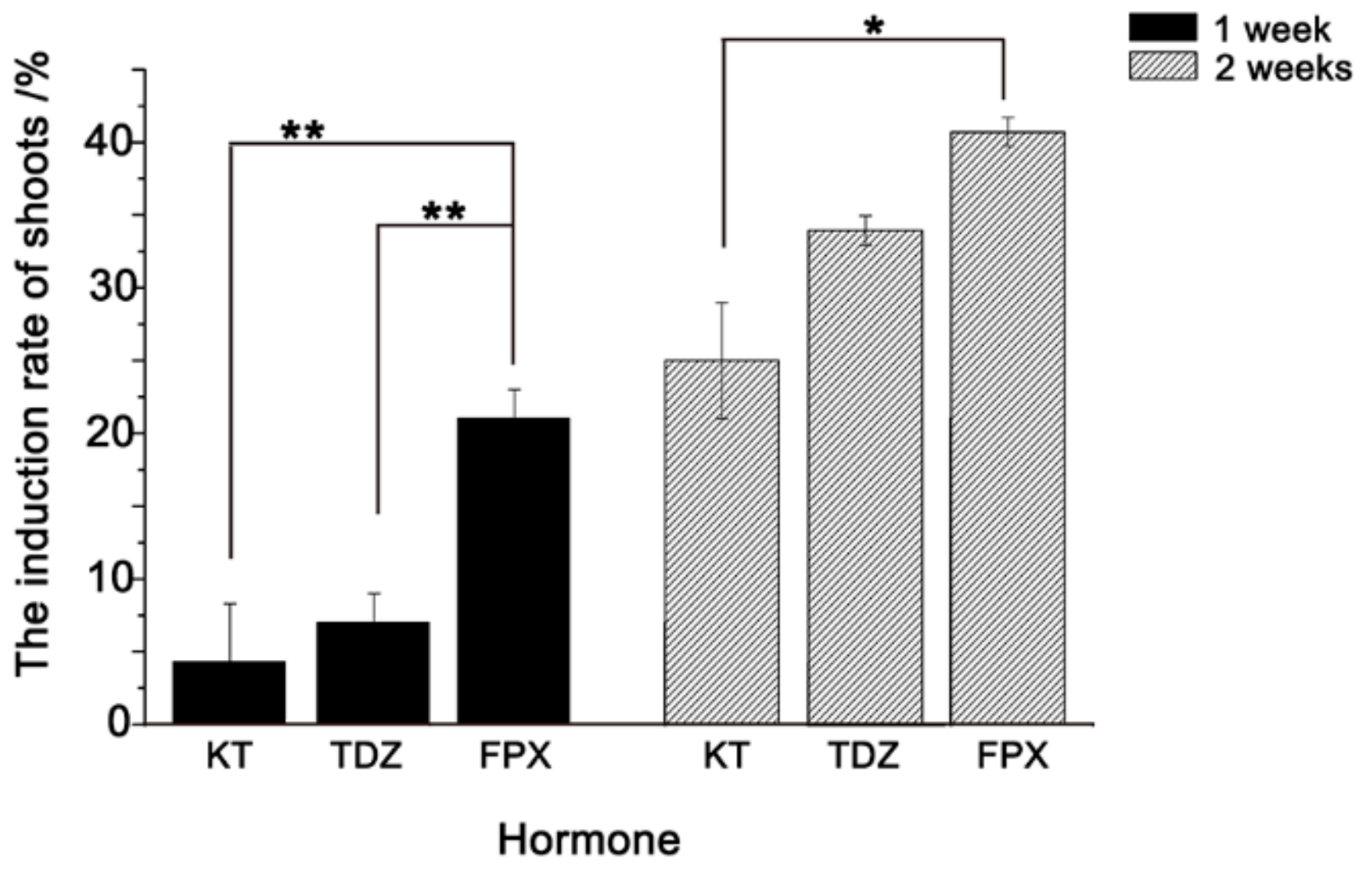
2.2. FPX Improves the Induction of Bd21 Shoot Regeneration
2.3. Photoautotrophic Rooting is a More Convenient and Efficient Rooting Method for Bd21
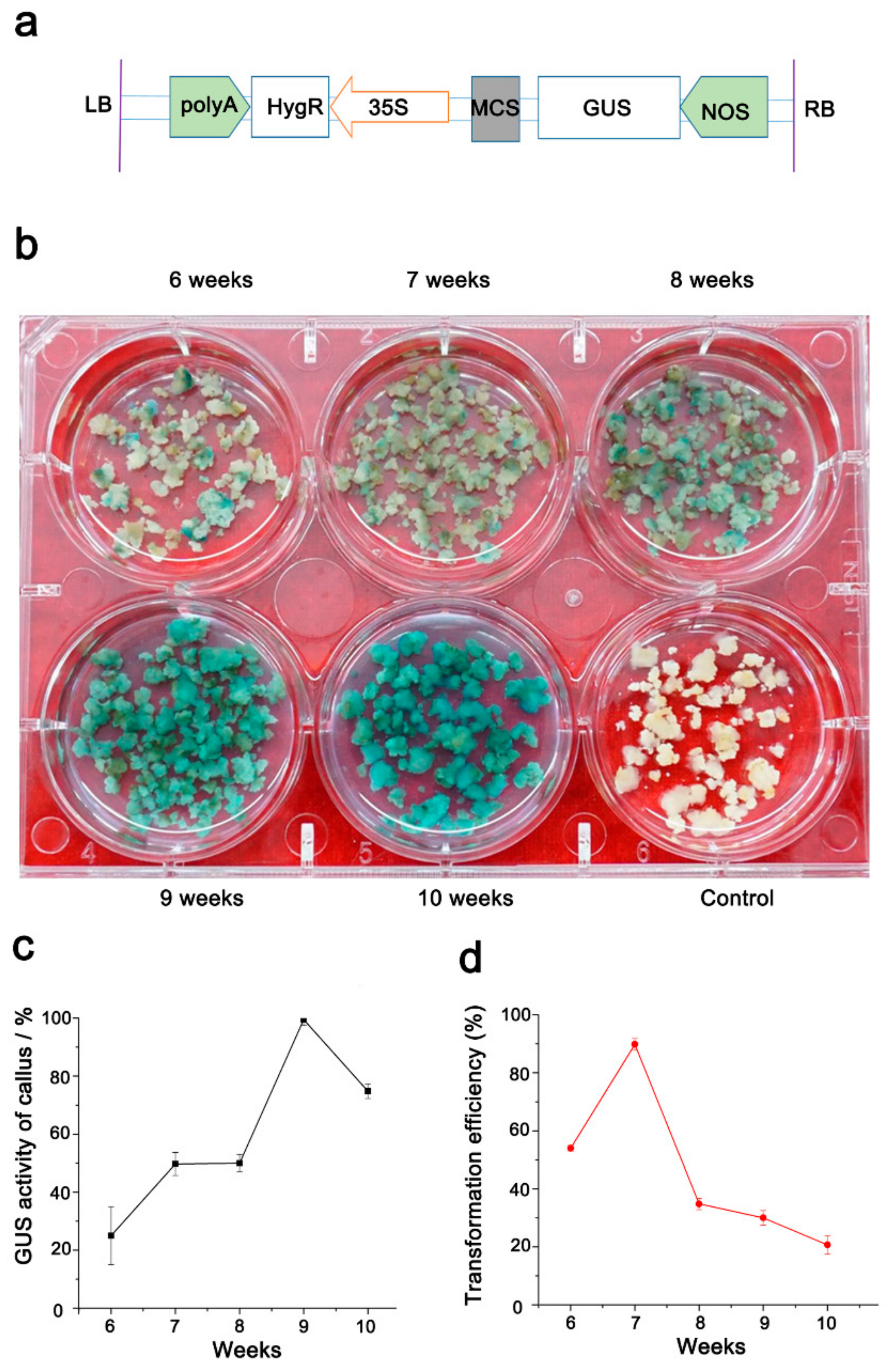
2.4. Optimization of Conditions for Efficient Transformation from Bd21 Callus
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Callus Induction and Regeneration
4.3. Photoautotrophic Rooting
4.4. Agrobacterium Strain and Plasmid Vector
4.5. Agrobacterium-Mediated Transformation
4.6. Histochemical Assay of GUS Activity
4.7. Genomic DNA Extraction and PCR Analysis
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Draper, J.; Mur, L.A.J.; Jenkins, G.; Ghosh-Biswas, G.C.; Bablak, P.; Hasterok, R.; Routledge, A.P.M. Brachypodium Distachyon. A New Model System for Functional Genomics in Grasses. Plant Physiol. 2001, 127, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.P.; Garvin, D.F.; Leong, O.M.; Hayden, D.M. Agrobacterium-Mediated Transformation and Inbred Line Development in the Model Grass Brachypodium Distachyon. Plant Cell Tissue Organ Cult. 2006, 84, 199–211. [Google Scholar] [CrossRef]
- Hammami, R.; Cuadrado, A.; Friero, E.; Jouve, N.; Soler, C.; González, J.M. Callus Induction and Plant Regeneration from Immature Embryos of Brachypodium Distachyon with Different Chromosome Numbers. Biol. Plant. 2011, 55, 797. [Google Scholar] [CrossRef]
- Thole, V.; Peraldi, A.; Worland, B.; Nicholson, P.; Doonan, J.H.; Vain, P. T-DNA Mutagenesis in Brachypodium Distachyon. J. Exp. Bot. 2011, 63, 567–576. [Google Scholar] [CrossRef]
- Scholthof, K.G.; Irigoyen, S.; Catalan, P.; Mandadi, K. Brachypodium: A Monocot Grass Model System for Plant Biology. Plant Cell 2018. [Google Scholar] [CrossRef]
- Garvin, D.F.; Gu, Y.Q.; Hasterok, R.; Hazen, S.P.; Jenkins, G.; Mockler, T.C.; Mur Luis, A.J.; Vogel, J.P. Development of Genetic and Genomic Research Resources for Brachypodium distachyon, a New Model System for Grass Crop Research. Crop Sci. 2008, 48, S69–S84. [Google Scholar] [CrossRef]
- Mur, L.A.; Allainguillaume, J.; Catalan, P.; Hasterok, R.; Jenkins, G.; Lesniewska, K.; Thomas, I.; Vogel, J. Exploiting the Brachypodium Tool Box in Cereal and Grass Research. New Phytol. 2011, 191, 334–347. [Google Scholar] [CrossRef]
- Kellogg, E.A. Brachypodium Distachyon as a Genetic Model System. Annu. Rev. Genet. 2015, 49, 1–20. [Google Scholar] [CrossRef]
- Lee, M.B.; Jeon, W.B.; Kim, D.Y.; Bold, O.; Hong, M.J.; Lee, Y.J.; Park, J.H.; Seo, Y.W. Agrobacterium-Mediated Transformation of Brachypodium Distachyon Inbred Line Bd21 with Two Binary Vectors Containing Hygromycin Resistance and Gus Reporter Genes. J. Crop Sci. Biotechnol. 2011, 14, 233–238. [Google Scholar] [CrossRef]
- International Brachypodium Initiative. Genome Sequencing and Analysis of the Model Grass Brachypodium Distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef]
- Tzfira, T.; Citovsky, V. Agrobacterium-Mediated Genetic Transformation of Plants: Biology and Biotechnology. Curr. Opin. Biotechnol. 2006, 17, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.H.; Yu, M.; Lai, E.M. Agrobacterium-Mediated Plant Transformation: Biology and Applications. Arabidopsis Book 2017, 15, e0186. [Google Scholar] [CrossRef] [PubMed]
- Altpeter, F.; Springer, N.M.; Bartley, L.E.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing Crop Transformation in the Era of Genome Editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sogutmaz, O.B.; Budak, H. Application of Tissue Culture and Transformation Techniques in Model Species Brachypodium Distachyon. In Brachypodium Genomics: Methods and Protocols; Springer: New York, NY, USA, 2018; pp. 289–310. [Google Scholar]
- Alves, S.C.; Worland, B.; Thole, V.; Snape, J.W.; Bevan, M.W.; Vain, P. A Protocol for Agrobacterium-Mediated Transformation of Brachypodium Distachyon Community Standard Line Bd21. Nat. Protoc. 2009, 4, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.; Hill, T. High-Efficiency Agrobacterium-Mediated Transformation of Brachypodium Distachyon Inbred Line Bd21–3. Plant Cell Rep. 2008, 27, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Bragg, J.N.; Anderton, A.; Nieu, R.; Vogel, J.P. Brachypodium Distachyon. Methods Mol. Biol. 2015, 1223, 17–33. [Google Scholar]
- Sogutmaz Ozdemir, B.; Budak, H. Application of Tissue Culture and Transformation Techniques in Model Species Brachypodium distachyon. Methods Mol. Biol. 2018, 1667, 289–310. [Google Scholar]
- Ashley, J.A.; Preece, J.E. Seed Cutting Treatments Stimulate Germination and Elucidate a Dormancy Gradient in Dormant Fraxinus Americana, L. And Fraxinus Pennsylvanica Marsh. Propag. Ornam. Plants 2009, 9, 122–128. [Google Scholar]
- Yan, J.; Wang, Y.; Gutterman, Y.; Nevo, E.; Cheng, J. Effects of Seed Cutting on Seed Dormancy and Germination of Brochypodium Distachyon. Seed 2010, 29, 15–18. [Google Scholar]
- Nakano, T.; Tanaka, S.; Ohtani, M.; Yamagami, A.; Takeno, S.; Hara, N.; Mori, A.; Nakano, A.; Hirose, S.; Himuro, Y.; et al. Fpx Is a Novel Chemical Inducer That Promotes Callus Formation and Shoot Regeneration in Plants. Plant Cell Physiol. 2018, 59, 1555–1567. [Google Scholar] [CrossRef]
- Ragonezi, C.; Klimaszewska, K.; Castro, M.R.; Lima, M.; De, O.P.; Zavattieri, M.A. Adventitious Rooting of Conifers: Influence of Physical and Chemical Factors. Trees 2010, 24, 975–992. [Google Scholar] [CrossRef]
- Díaz-Sala, C. Direct Reprogramming of Adult Somatic Cells toward Adventitious Root Formation in Forest Tree Species: The Effect of the Juvenile-Adult Transition. Front. Plant Sci. 2014, 5, 310. [Google Scholar] [CrossRef] [PubMed]
- Vunsh, R. Methods for Functional Transgenics: Development of Highly Efficient Transformation Protocol in Brachypodium and Its Suitability for Advancing Brachypodium Transgenics. Methods Mol. Biol. 2018, 1667, 101–117. [Google Scholar] [PubMed]
- Bablak, P.; Draper, J.; Davey, M.R.; Lynch, P.T. Plant Regeneration and Micropropagation of Brachypodium Distachyon. Plant Cell Tissue Organ. Cult. 1995, 42, 97–107. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Types of Seeds and Kinds of Seed Dormancy. In Ecology, Biogeography and Evolution of Dormancy and Germination; Elsevier: Amsterdam, The Netherlands, 2014; pp. 37–78. [Google Scholar]
- Barrero, J.M.; Jacobsen, J.V.; Talbot, M.J.; White, R.G.; Swain, S.M.; Garvin, D.F.; Gubler, F. Grain Dormancy and Light Quality Effects on Germination in the Model Grass Brachypodium Distachyon. New Phytol. 2012, 193, 376–386. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J. Molecular Mechanisms of Seed Dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef]
- Finch Savage, W.E.; Leubner Metzger, G. Seed Dormancy and the Control of Germination. New Phytol. 2010, 171, 501–523. [Google Scholar] [CrossRef]
- Adelberg, J.; Fujiwara, K.; Kirdmanee, C.; Kozai, T. Photoautotrophic Shoot and Root Development for Triploid Melon. Plant Cell Tissue Organ. Cult. 1999, 57, 95–104. [Google Scholar] [CrossRef]
- Kozai, T. Autotrophic Micropropagation. In High-Tech and Micropropagation I; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1991; pp. 313–343. [Google Scholar]
- Sutter, E. Stomatal and Cuticular Water Loss from Apple, Cherry and Sweetgum Plants after Removal from in Vitro Culture. J. Am. Soc. Hortic. Sci. 1988, 113, 234–238. [Google Scholar]
- De Klerk, G.J. Rooting of Microcuttings: Theory and Practice. In Vitro Cell. Dev. Biol. Plant 2002, 38, 415–422. [Google Scholar] [CrossRef]
- Wu, X.; Liu, J.; Nielsen, K.; Yang, Z.; Gao, C. Agrobacterium-Mediated Transformation of Brachypodium Distachyon through Embryogenic Calli Derived from Immature Embryos. Acta Prat. Sin. 2010, 19, 9–16. [Google Scholar]
- Chen, F.; Liu, Q.; Vogel, J.P.; Wu, J. Agrobacterium-Mediated Transformation of Brachypodium Distachyon. Curr. Protoc. Plant Biol. 2019, e20088. [Google Scholar] [CrossRef] [PubMed]
- Thole, V.; Vain, P. Agrobacterium-Mediated Transformation of Brachypodium Distachyon. In Transgenic Plants; Jim, M.D., Andy, C.W., Eds.; Humana Press: Berlin, Germany, 2012; pp. 137–149. [Google Scholar]
- Li, D.Z.; Xu, L.; Pang, S.; Liu, Z.Q.; Zhao, W.S.; Wang, C.J. Multiple Pesticides Detoxification Function of Maize (Zea Mays) Gst34. J. Agric. Food Chem. 2017, 65, 1847–1853. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A Modified Protocol for Rapid DNA Isolation from Plant Tissues Using Cetyltrimethylammonium Bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef] [PubMed]
- Collier, R.; Bragg, J.; Hernandez, B.T.; Vogel, J.P.; Thilmony, R. Use of Agrobacterium Rhizogenes Strain 18r12v and Paromomycin Selection for Transformation of Brachypodium Distachyon and Brachypodium Sylvaticum. Front. Plant Sci. 2016, 7, 716. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, G.; Wang, J.; Miao, L.; Xi, M.; Wang, Q.; Wang, K. Optimization of Mature Embryo-Based Tissue Culture and Agrobacterium-Mediated Transformation in Model Grass Brachypodium distachyon. Int. J. Mol. Sci. 2019, 20, 5448. https://doi.org/10.3390/ijms20215448
Yu G, Wang J, Miao L, Xi M, Wang Q, Wang K. Optimization of Mature Embryo-Based Tissue Culture and Agrobacterium-Mediated Transformation in Model Grass Brachypodium distachyon. International Journal of Molecular Sciences. 2019; 20(21):5448. https://doi.org/10.3390/ijms20215448
Chicago/Turabian StyleYu, Guangrun, Jianyong Wang, Li Miao, Mengli Xi, Qiongli Wang, and Kai Wang. 2019. "Optimization of Mature Embryo-Based Tissue Culture and Agrobacterium-Mediated Transformation in Model Grass Brachypodium distachyon" International Journal of Molecular Sciences 20, no. 21: 5448. https://doi.org/10.3390/ijms20215448