Reduced Cholinergic Activity in the Hippocampus of Hippocampal Cholinergic Neurostimulating Peptide Precursor Protein Knockout Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Generation of HCNP-pp KO Mice
2.2. No Morphological Changes in HCNP-pp KO Mice Observed by Light Microscopy
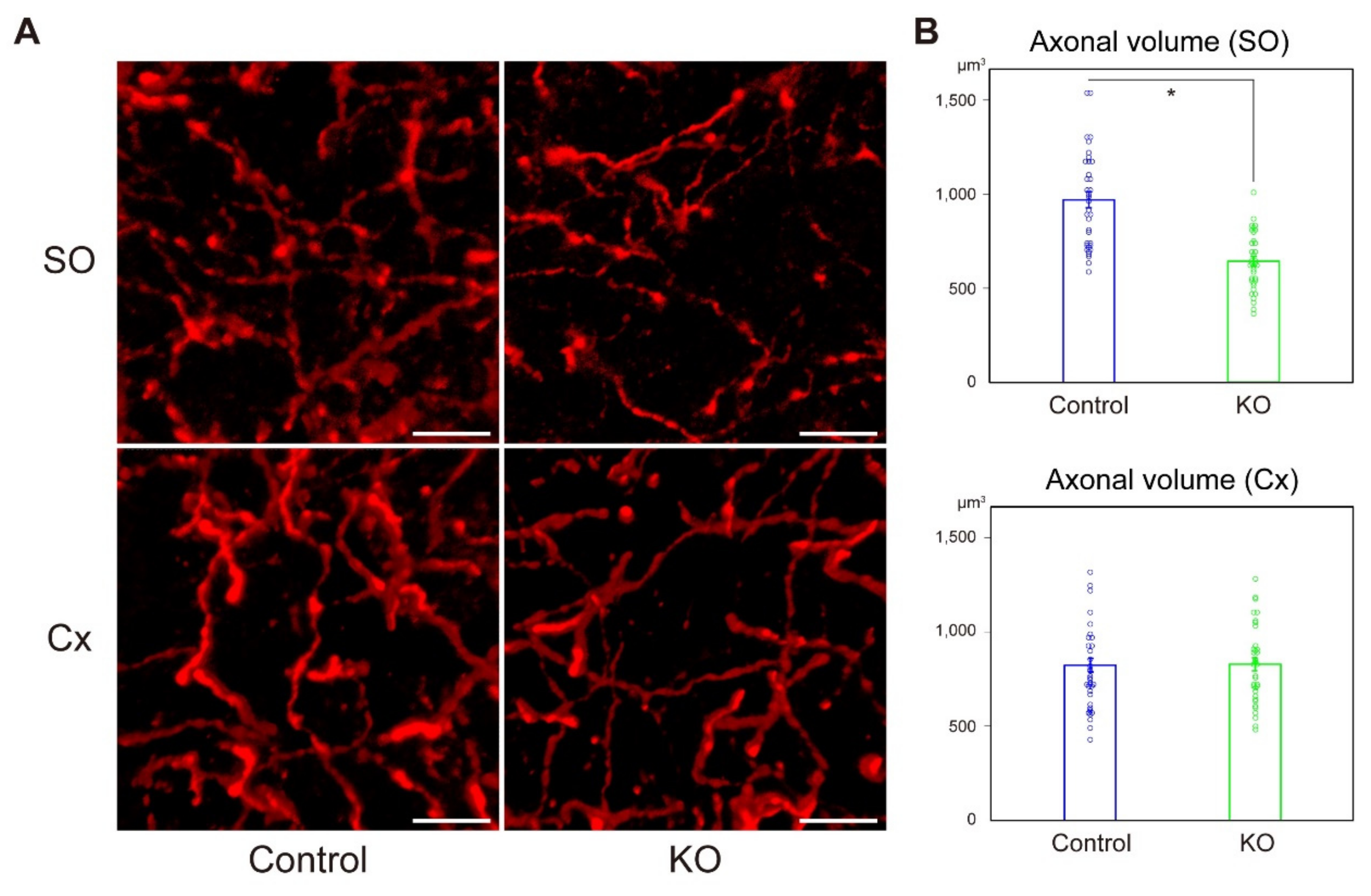
2.3. Diminished Cholinergic Projection to the CA1 Stratum Oriens in HCNP-pp KO Mice
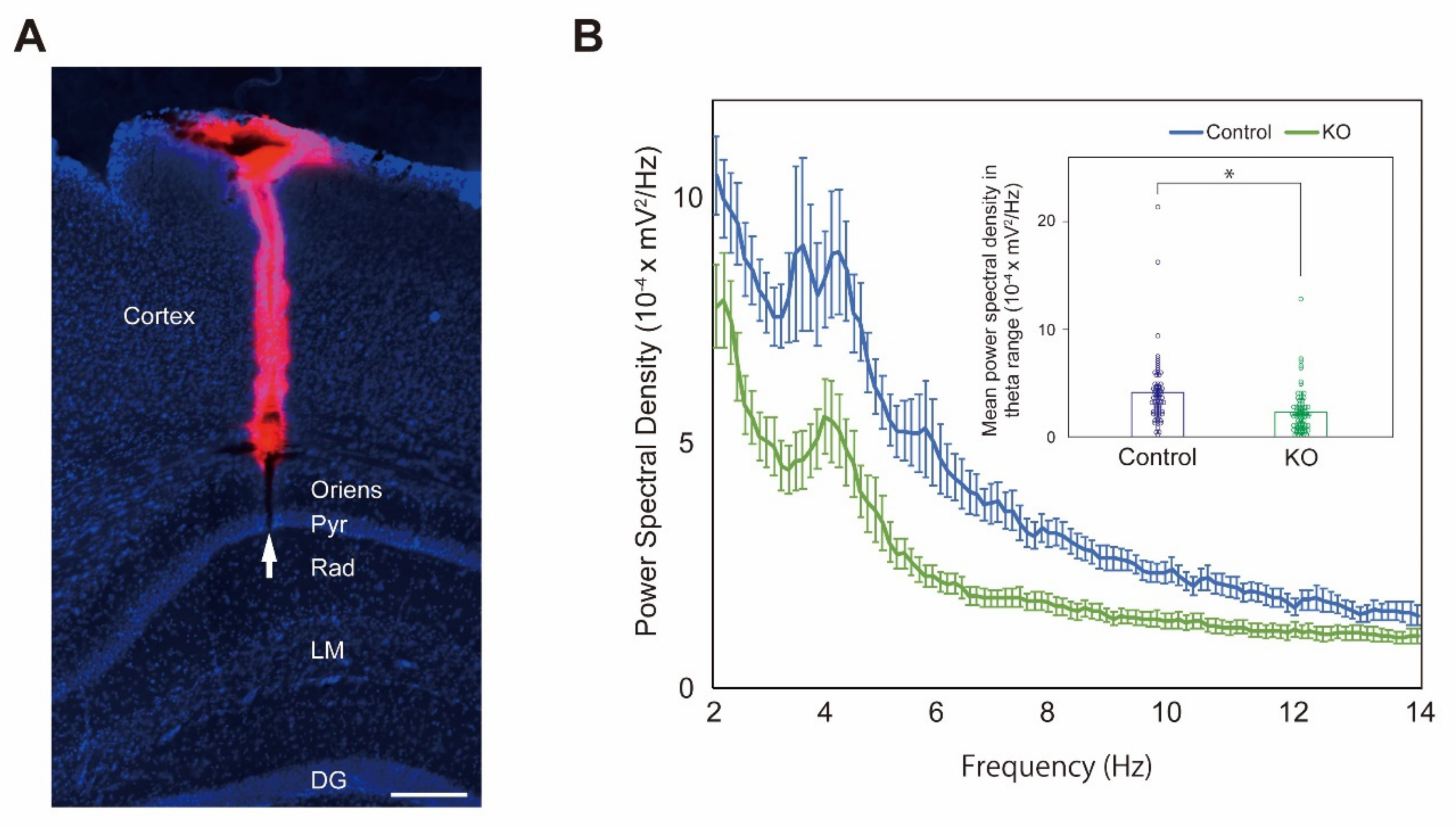
2.4. Reduction of Theta Activity in the CA1 Pyramidal Layer of HCNP-pp KO Mice
2.5. HCNP-pp KO Mice Show No Distinct Behavioral Phenotype
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Immunohistochemistry
4.3. Western Blot Analysis
4.4. Morphological Analysis
4.5. Quantification of Hippocampal Cholinergic Axons
4.6. Surgery
4.7. Electrophysiological Recordings
4.8. Behavioral Tests
4.8.1. Open Field Test
4.8.2. Y-Maze Test
4.8.3. Novel Object Recognition Test
4.8.4. Tail Suspension Test
4.8.5. Forced Swimming Test
4.9. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colgin, L.L. Rhythms of the hippocampal network. Nat. Rev. Neurosci. 2016, 17, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Mesulam, M.M.; Mufson, E.J.; Wainer, B.H.; Levey, A.I. Central cholinergic pathways in the rat: An overview based on an alternative nomenclature (Ch1-Ch6). Neuroscience 1983, 10, 1185–1201. [Google Scholar] [CrossRef]
- Ohi, Y.; Kato, D.; Mizuno, M.; Sato, T.; Ueki, Y.; Borlongan, C.V.; Ojika, K.; Haji, A.; Matsukawa, N. Enhancement of long-term potentiation via muscarinic modulation in the hippocampus of HCNP precursor transgenic mice. Neurosci. Lett. 2015, 597, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Ohi, Y.; Kato, D.; Mizuno, M.; Takase, H.; Kanamori, T.; Borlongan, C.V.; Haji, A.; Matsukawa, N. Hippocampal Cholinergic Neurostimulating Peptide as a Possible Modulating Factor against Glutamatergic Neuronal Disability by Amyloid Oligomers. Cell Transplant. 2017, 26, 1542–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, J.D.; Arduini, A.A. Hippocampal electrical activity in arousal. J. Neurophysiol. 1954, 17, 533–557. [Google Scholar] [CrossRef] [PubMed]
- Dupret, D.; O’Neill, J.; Csicsvari, J. Dynamic reconfiguration of hippocampal interneuron circuits during spatial learning. Neuron 2013, 78, 166–180. [Google Scholar] [CrossRef]
- Jezek, K.; Henriksen, E.J.; Treves, A.; Moser, E.I.; Moser, M.B. Theta-paced flickering between place-cell maps in the hippocampus. Nature 2011, 478, 246–249. [Google Scholar] [CrossRef] [Green Version]
- Ojika, K.; Kojima, S.; Ueki, Y.; Fukushima, N.; Hayashi, K.; Yamamoto, M. Purification and structural analysis of hippocampal cholinergic neurostimulating peptide. Brain Res. 1992, 572, 164–171. [Google Scholar] [CrossRef]
- Ojika, K.; Mitake, S.; Tohdoh, N.; Appel, S.H.; Otsuka, Y.; Katada, E.; Matsukawa, N. Hippocampal cholinergic neurostimulating peptides (HCNP). Prog. Neurobiol. 2000, 60, 37–83. [Google Scholar] [CrossRef]
- Capecchi, M.R. Altering the genome by homologous recombination. Science 1989, 244, 1288–1292. [Google Scholar] [CrossRef]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Imai, H.; Matsukawa, M.; Okado, N. Lamina-selective changes in the density of synapses following perturbation of monoamines and acetylcholine in the rat medial prefrontal cortex. Brain Res. 2004, 1012, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Yakel, J.L. Timing-dependent septal cholinergic induction of dynamic hippocampal synaptic plasticity. Neuron 2011, 71, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Dannenberg, H.; Pabst, M.; Braganza, O.; Schoch, S.; Niediek, J.; Bayraktar, M.; Mormann, F.; Beck, H. Synergy of direct and indirect cholinergic septo-hippocampal pathways coordinates firing in hippocampal networks. J. Neurosci. 2015, 35, 8394–8410. [Google Scholar] [CrossRef]
- Vandecasteele, M.; Varga, V.; Berényi, A.; Papp, E.; Barthó, P.; Venance, L.; Freund, T.F.; Buzsáki, G. Optogenetic activation of septal cholinergic neurons suppresses sharp wave ripples and enhances theta oscillations in the hippocampus. Proc. Natl. Acad. Sci. USA 2014, 111, 13535–13540. [Google Scholar] [CrossRef] [Green Version]
- Hengst, U.; Albrecht, H.; Hess, D.; Monard, D. The phosphatidylethanolamine-binding protein is the prototype of a novel family of serine protease inhibitors. J. Biol. Chem. 2001, 276, 535–540. [Google Scholar] [CrossRef]
- Yeung, K.; Seitz, T.; Li, S.; Janosch, P.; McFerran, B.; Kaiser, C.; Fee, F.; Katsanakis, K.D.; Rose, D.W.; Mischak, H.; et al. Suppression of Raf-1 kinase activity and MAP kinase signalling by RKIP. Nature 1999, 401, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, M.; Johnson-Venkatesh, E.M.; Zhang, H.; Parent, J.M.; Sutton, M.A.; Umemori, H. Multiple forms of activity-dependent competition refine hippocampal circuits in vivo. Neuron 2011, 70, 1128–1142. [Google Scholar] [CrossRef]
- Yuasa, H.; Ojika, K.; Mitake, S.; Katada, E.; Matsukawa, N.; Otsuka, Y.; Fujimori, O.; Hirano, A. Age-dependent changes in HCNP-related antigen expression in the human hippocampus. Dev. Brain Res. 2001, 127, 1–7. [Google Scholar] [CrossRef]
- Mitake, S.; Ojika, K.; Katada, E.; Otsuka, Y.; Matsukawa, N.; Fujimori, O. Distribution of hippocampal cholinergic neurostimulating peptide (HCNP) immunoreactivity in the central nervous system of the rat. Brain Res. 1996, 706, 57–70. [Google Scholar] [CrossRef]
- Taniguchi, H.; He, M.; Wu, P.; Kim, S.; Paik, R.; Sugino, K.; Kvitsiani, D.; Fu, Y.; Lu, J.; Lin, Y.; et al. A resource of Cre driver lines for genetic targeting of GABAergic neurons in cerebral cortex. Neuron 2011, 71, 995–1013. [Google Scholar] [CrossRef] [PubMed]
- Uematsu, N.; Matsukawa, N.; Kanamori, T.; Arai, Y.; Sagisaka, T.; Toyoda, T.; Yoshida, M.; Ojika, K. Overexpression of hippocampal cholinergic neurostimulating peptide in heterozygous transgenic mice increases the amount of ChAT in the medial septal nucleus. Brain Res. 2009, 1305, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Mele, T.; Juric, D.M. Metrifonate, like acetylcholine, up-regulates neurotrophic activity of cultured rat astrocytes. Pharmacol. Rep. 2014, 66, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Di Liberto, V.; Borroto-Escuela, D.O.; Frinchi, M.; Verdi, V.; Fuxe, K.; Belluardo, N.; Mudò, G. Existence of muscarinic acetylcholine receptor (mAChR) and fibroblast growth factor receptor (FGFR) heteroreceptor complexes and their enhancement of neurite outgrowth in neural hippocampal cultures. Biochim. Biophys. Acta. Gen. Subj. 2017, 1861, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Madziar, B.; Lopez-Coviella, I.; Zemelko, V.; Berse, B. Regulation of cholinergic gene expression by nerve growth factor depends on the phosphatidylinositol-3’-kinase pathway. J. Neurochem. 2005, 92, 767–779. [Google Scholar] [CrossRef]
- Lee, M.G.; Chrobak, J.J.; Sik, A.; Wiley, R.G.; Buzsáki, G. Hippocampal theta activity following selective lesion of the septal cholinergic system. Neuroscience 1994, 62, 1033–1047. [Google Scholar] [CrossRef]
- Bender, F.; Gorbati, M.; Cadavieco, M.C.; Denisova, N.; Gao, X.; Holman, C.; Korotkova, T.; Ponomarenko, A. Theta oscillations regulate the speed of locomotion via a hippocampus to lateral septum pathway. Nat. Commun. 2015, 6, 8521. [Google Scholar] [CrossRef] [Green Version]
- Marrosu, F.; Portas, C.; Mascia, M.S.; Casu, M.A.; Fà, M.; Giagheddu, M.; Imperato, A.; Gessa, G.L. Microdialysis measurement of cortical and hippocampal acetylcholine release during sleep-wake cycle in freely moving cats. Brain Res. 1995, 671, 329–332. [Google Scholar] [CrossRef]
- Mamad, O.; McNamara, H.M.; Reilly, R.B.; Tsanov, M. Medial septum regulates the hippocampal spatial representation. Front. Behav. Neurosci. 2015, 9, 166. [Google Scholar] [CrossRef]
- Whittington, R.A.; Virag, L. The differential effects of equipotent doses of isoflurane and desflurane on hippocampal acetylcholine levels in young and aged rats. Neurosci. Lett. 2010, 471, 166–170. [Google Scholar] [CrossRef]
- Ishisaka, M.; Kakefuda, K.; Yamauchi, M.; Tsuruma, K.; Shimazawa, M.; Tsuruta, A.; Hara, H. Luteolin shows an antidepressant-like effect via suppressing endoplasmic reticulum stress. Biol. Pharm. Bull. 2011, 34, 1481–1486. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, N.; Furuya, Y.; Ogura, H.; Ojika, K. HCNP precursor protein transgenic mice display a depressive-like phenotype in old age. Brain Res. 2010, 1349, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Wess, J.; Duttaroy, A.; Zhang, W.; Gomeza, J.; Cui, Y.; Miyakawa, T.; Bymaster, F.P.; McKinzie, L.; Felder, C.C.; Lamping, K.G.; et al. M1-M5 muscarinic receptor knockout mice as novel tools to study the physiological roles of the muscarinic cholinergic system. Recept. Channels 2003, 9, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Leaderbrand, K.; Chen, H.J.; Corcoran, K.A.; Guedea, A.L.; Jovasevic, V.; Wess, J.; Radulovic, J. Muscarinic acetylcholine receptors act in synergy to facilitate learning and memory. Learn. Mem. 2016, 23, 631–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, R.M.; Norman, T.A.; Haydar, T.F.; Slack, B.E.; Leeman, S.E.; Blusztajn, J.K.; Mellott, T.J. BMP9 ameliorates amyloidosis and the cholinergic defect in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2013, 110, 19567–19572. [Google Scholar] [CrossRef] [PubMed]
- Crutcher, K.A.; Collins, F. In vitro Evidence for two distinct hippocampal growth factors: Basis of neuronal plasticity? Science 1982, 217, 67–68. [Google Scholar] [CrossRef] [PubMed]
- Korsching, S.; Auburger, G.; Heumann, R.; Scott, J.; Thoenen, H. Levels of nerve growth factor and its mRNA in the central nervous system of the rat correlate with cholinergic innervation. EMBO J. 1985, 4, 1389–1393. [Google Scholar] [CrossRef]
- Holtzman, D.M.; Li, Y.; Parada, L.F.; Kinsman, S.; Chen, C.K.; Valletta, J.S.; Zhou, J.; Long, J.B.; Mobley, W.C. p140trk mRNA marks NGF-responsive forebrain neurons: Evidence that trk gene expression is induced by NGF. Neuron 1992, 9, 465–478. [Google Scholar] [CrossRef]
- Koh, S.; Higgins, G.A. Differential regulation of the low-affinity nerve growth factor receptor during postnatal development of the rat brain. J. Comp. Neurol. 1991, 313, 494–508. [Google Scholar] [CrossRef]
- Pioro, E.P.; Cuello, A.C. Distribution of nerve growth factor receptor-like immunoreactivity in the adult rat central nervous system. Effect of colchicine and correlation with the cholinergic system--I. Forebrain. Neuroscience 1990, 34, 57–87. [Google Scholar] [CrossRef]
- Alderson, R.F.; Alterman, A.L.; Barde, Y.A.; Lindsay, R.M. Brain-derived neurotrophic factor increases survival and differentiated functions of rat septal cholinergic neurons in culture. Neuron 1990, 5, 297–306. [Google Scholar] [CrossRef]
- Nonner, D.; Barrett, E.F.; Barrett, J.N. Neurotrophin effects on survival and expression of cholinergic properties in cultured rat septal neurons under normal and stress conditions. J. Neurosci. 1996, 16, 6665–6675. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Triaca, V.; Besusso, D.; Costanzi, M.; Horn, J.M.; Koudelka, J.; Geibel, M.; Cestari, V.; Minichiello, L. Loss of NGF-TrkA signaling from the CNS is not sufficient to induce cognitive impairments in young adult or intermediate-aged mice. J. Neurosci. 2012, 32, 14885–14898. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ortiz, E.; Yui, D.; Song, D.; Li, Y.; Rubenstein, J.L.; Reichardt, L.F.; Parada, L.F. TrkA gene ablation in basal forebrain results in dysfunction of the cholinergic circuitry. J. Neurosci. 2012, 32, 4065–4079. [Google Scholar] [CrossRef]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Academic Press: New York, NY, USA, 2008; Figure 48. [Google Scholar]
- Takata, N.; Yoshida, K.; Komaki, Y.; Xu, M.; Sakai, Y.; Hikishima, K.; Mimura, M.; Okano, H.; Tanaka, K.F. Optogenetic activation of CA1 pyramidal neurons at the dorsal and ventral hippocampus evokes distinct brain-wide responses revealed by mouse fMRI. PLoS ONE 2015, 10, e0121417. [Google Scholar] [CrossRef]
- Aitken, P.; Zheng, Y.; Smith, P.F. The modulation of hippocampal theta rhythm by the vestibular system. J. Neurophysiol. 2018, 119, 548–562. [Google Scholar] [CrossRef] [Green Version]
- Kakefuda, K.; Ishisaka, M.; Tsuruma, K.; Shimazawa, M.; Hara, H. Memantine, an NMDA receptor antagonist, improves working memory deficits in DGKbeta knockout mice. Neurosci. Lett. 2016, 630, 228–232. [Google Scholar] [CrossRef]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- Kakefuda, K.; Oyagi, A.; Ishisaka, M.; Tsuruma, K.; Shimazawa, M.; Yokota, K.; Shirai, Y.; Horie, K.; Saito, N.; Takeda, J.; et al. Diacylglycerol kinase beta knockout mice exhibit lithium-sensitive behavioral abnormalities. PLoS ONE 2010, 5, e13447. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madokoro, Y.; Yoshino, Y.; Kato, D.; Sato, T.; Mizuno, M.; Kanamori, T.; Shimazawa, M.; Hida, H.; Hara, H.; Yoshida, M.; et al. Reduced Cholinergic Activity in the Hippocampus of Hippocampal Cholinergic Neurostimulating Peptide Precursor Protein Knockout Mice. Int. J. Mol. Sci. 2019, 20, 5367. https://doi.org/10.3390/ijms20215367
Madokoro Y, Yoshino Y, Kato D, Sato T, Mizuno M, Kanamori T, Shimazawa M, Hida H, Hara H, Yoshida M, et al. Reduced Cholinergic Activity in the Hippocampus of Hippocampal Cholinergic Neurostimulating Peptide Precursor Protein Knockout Mice. International Journal of Molecular Sciences. 2019; 20(21):5367. https://doi.org/10.3390/ijms20215367
Chicago/Turabian StyleMadokoro, Yuta, Yuta Yoshino, Daisuke Kato, Toyohiro Sato, Masayuki Mizuno, Tetsuko Kanamori, Masamitsu Shimazawa, Hideki Hida, Hideaki Hara, Mari Yoshida, and et al. 2019. "Reduced Cholinergic Activity in the Hippocampus of Hippocampal Cholinergic Neurostimulating Peptide Precursor Protein Knockout Mice" International Journal of Molecular Sciences 20, no. 21: 5367. https://doi.org/10.3390/ijms20215367