A Novel HIF Inhibitor Halofuginone Prevents Neurodegeneration in a Murine Model of Retinal Ischemia-Reperfusion


 and
and
Abstract
:1. Introduction
2. Results
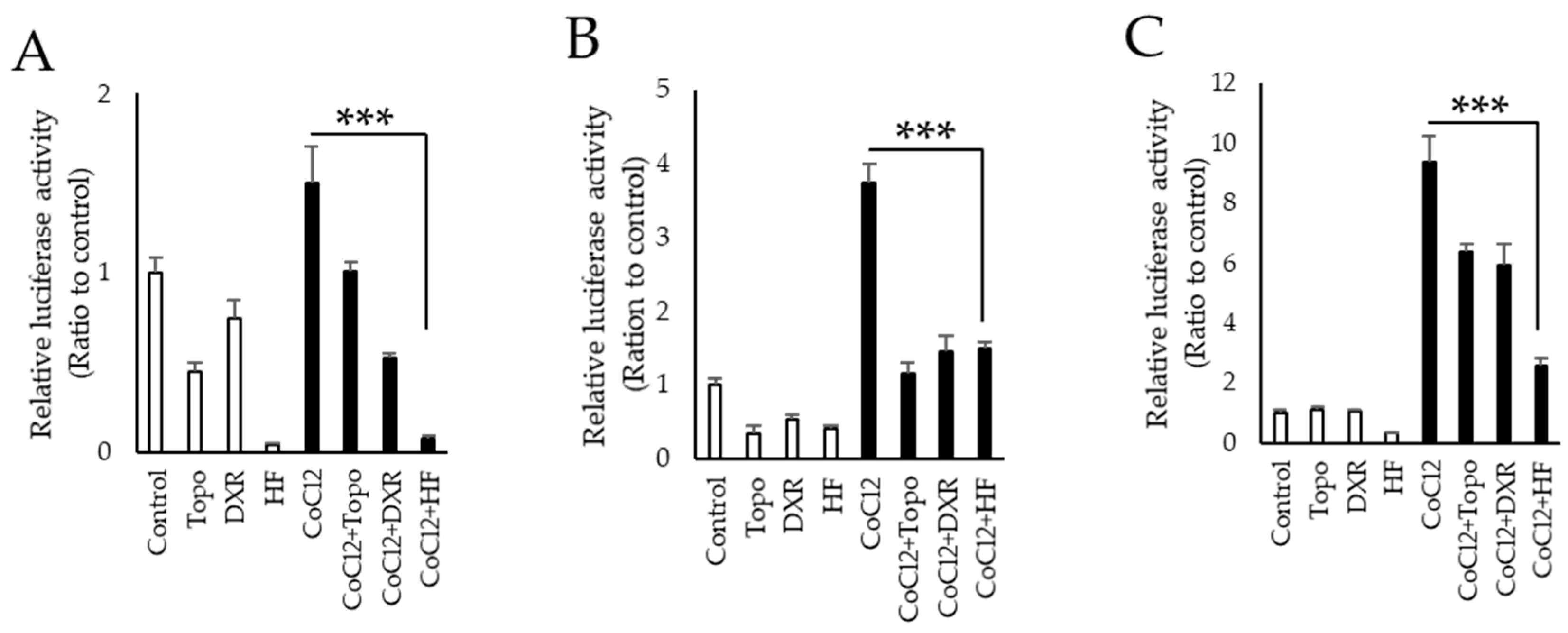
2.1. Screening for Novel HIF Inhibitors
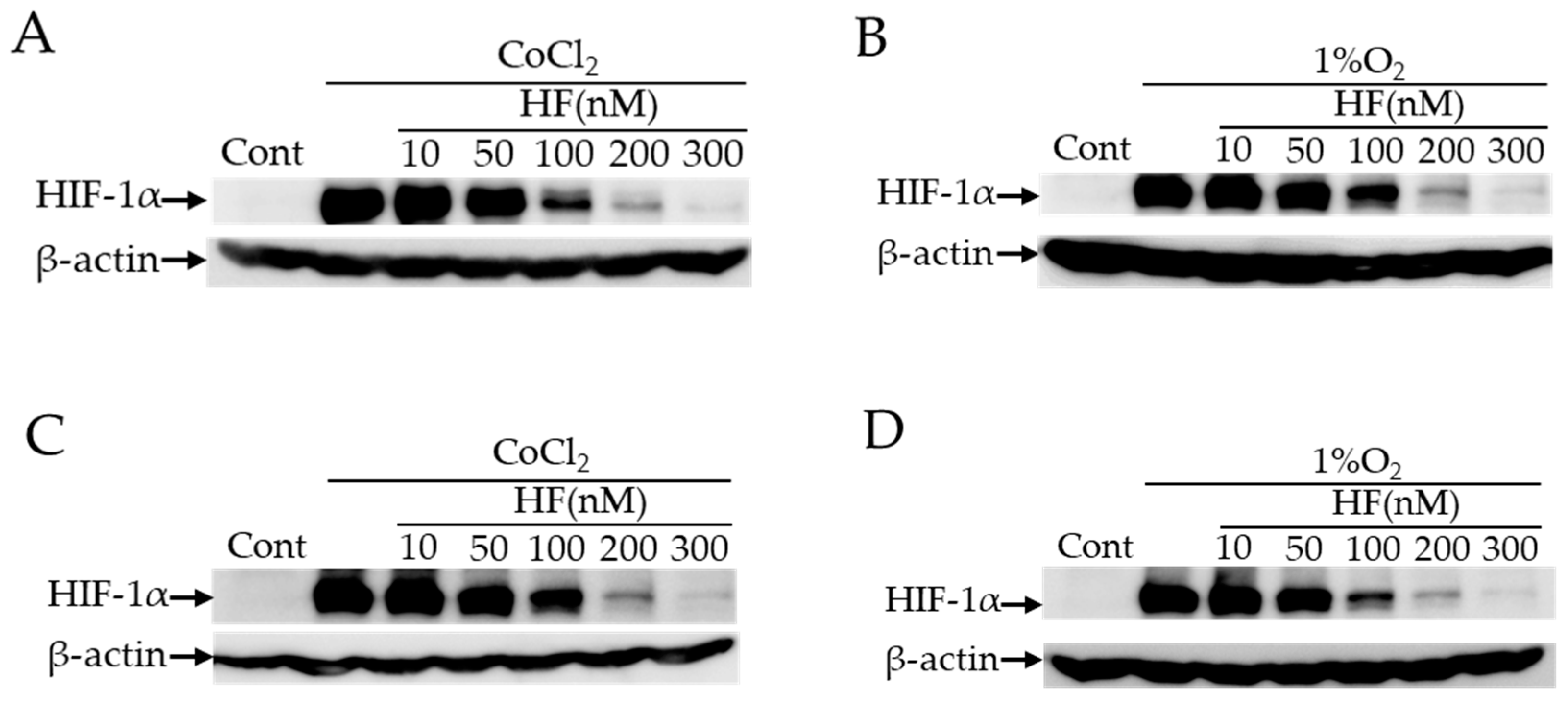
2.2. HIF Inhibitory Effects of Halofuginone In Vitro
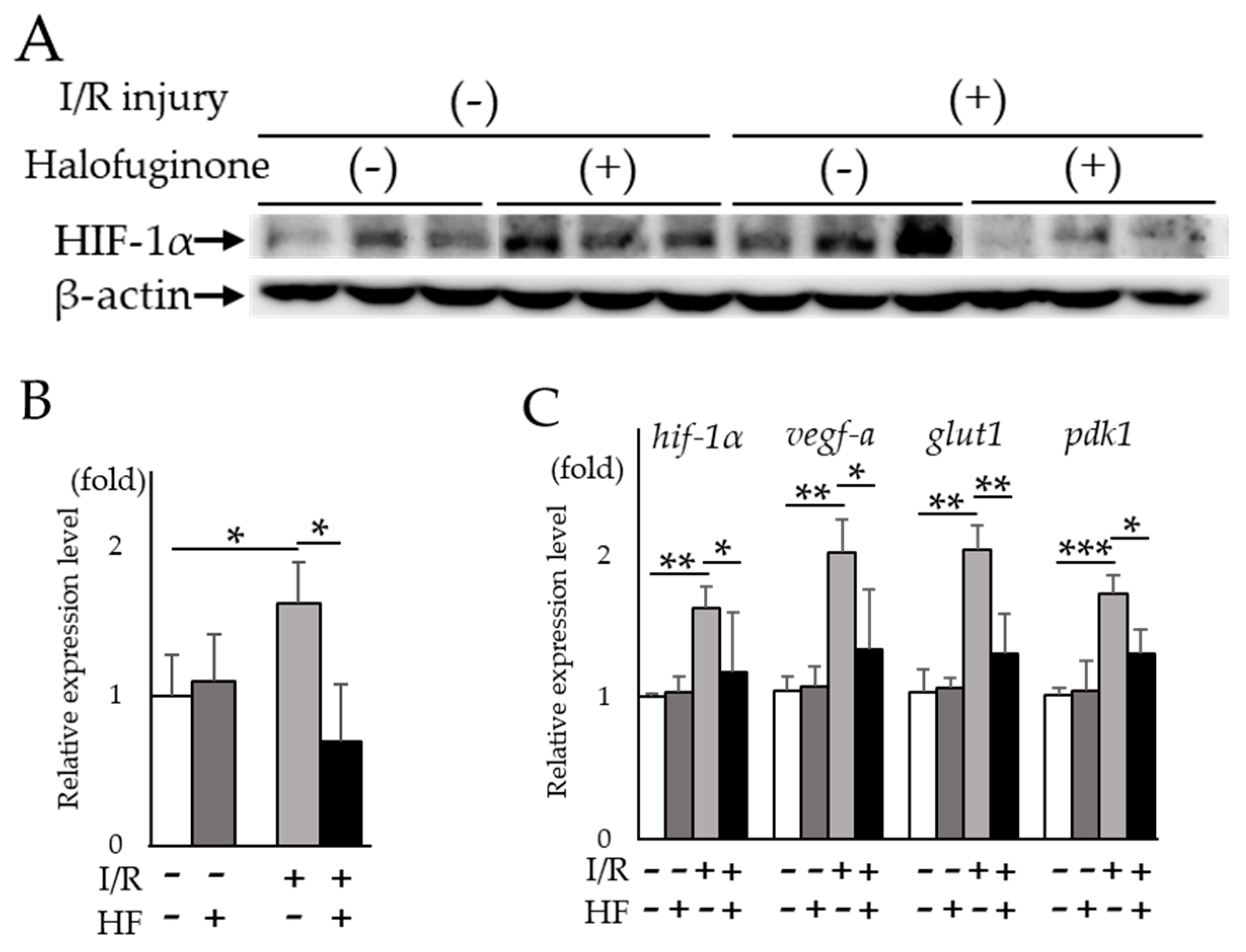
2.3. Halofuginone Inhibits Retinal HIF-1α and Target Genes In Vivo
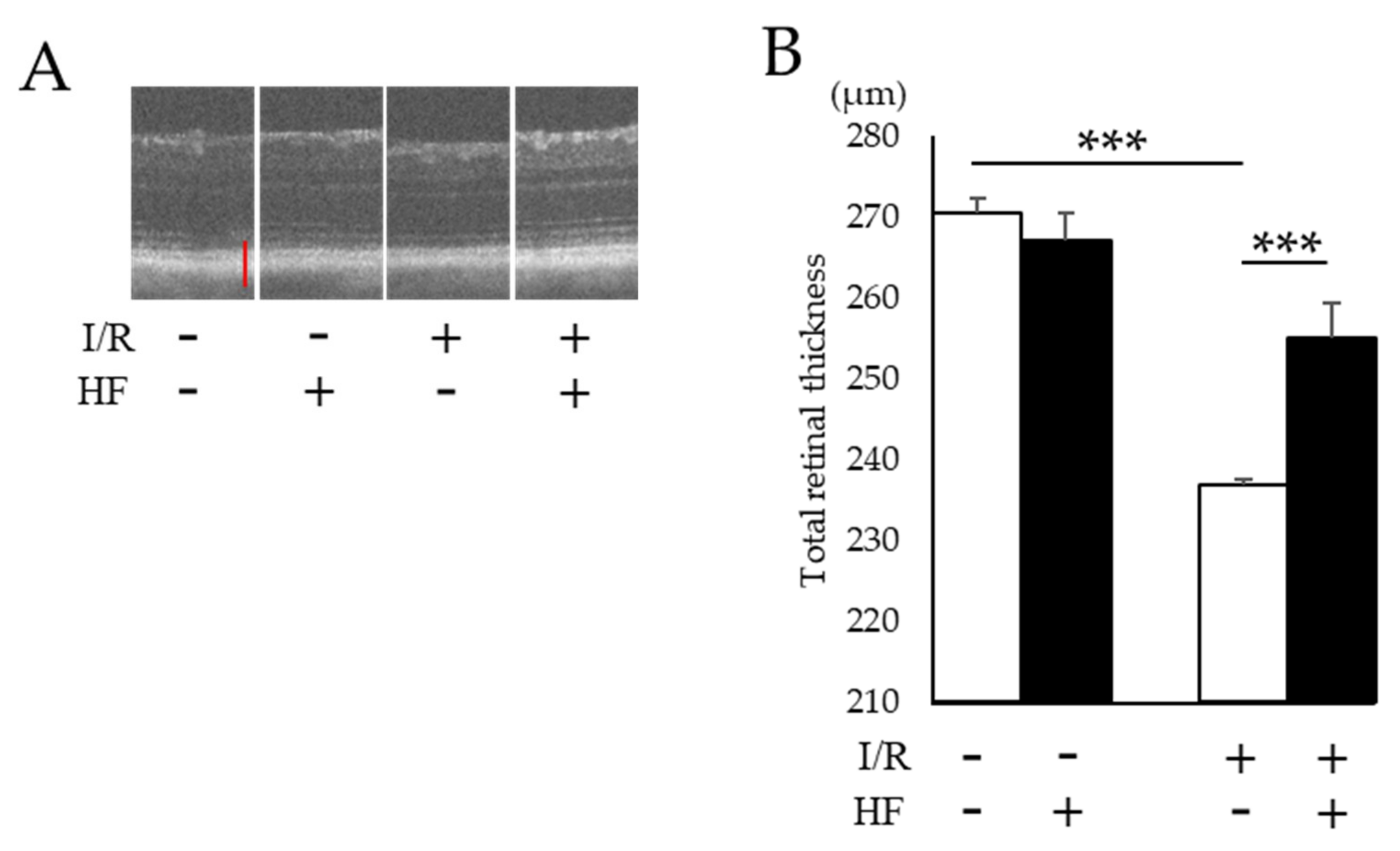
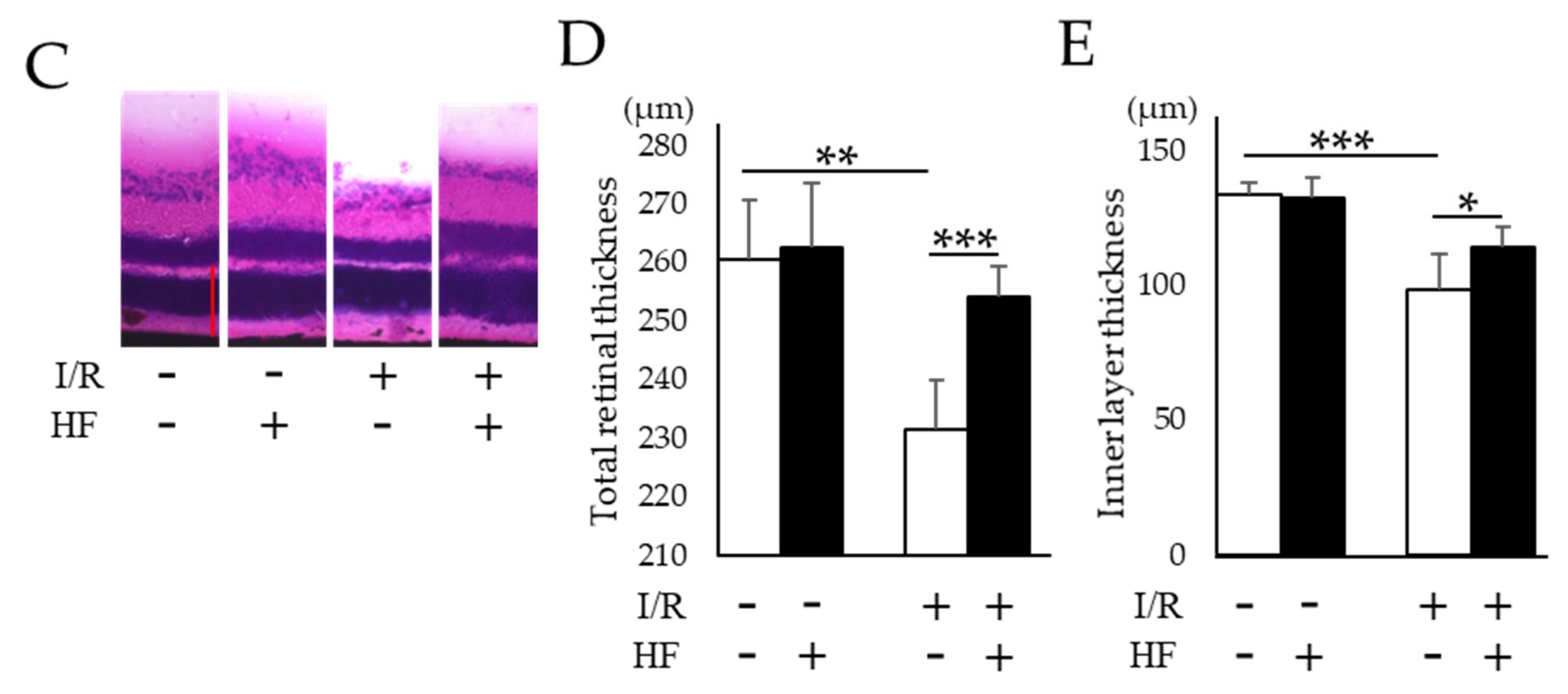
2.4. Improvement of RGC Survival with Halofuginone Administration in Post-I/R Retinas
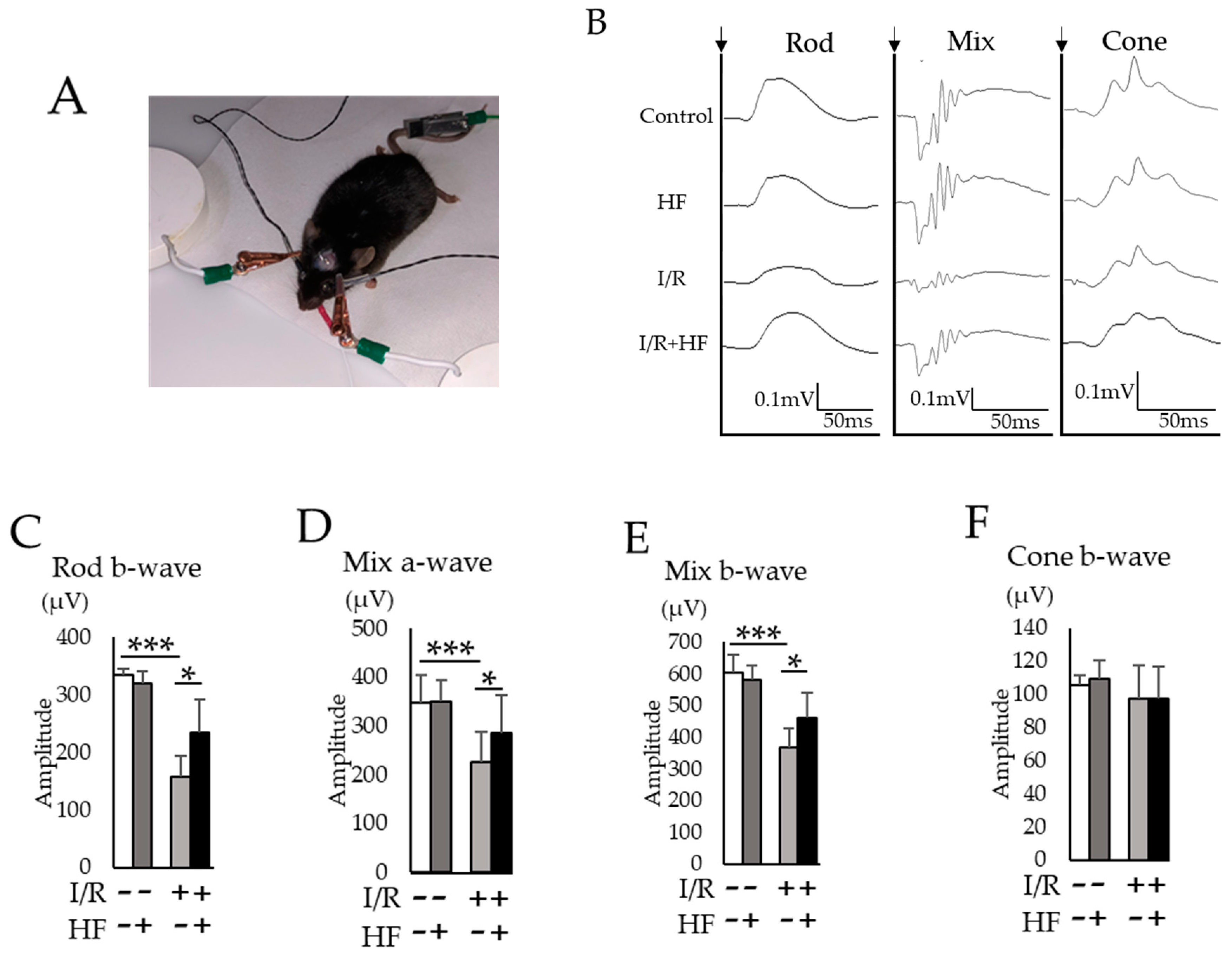
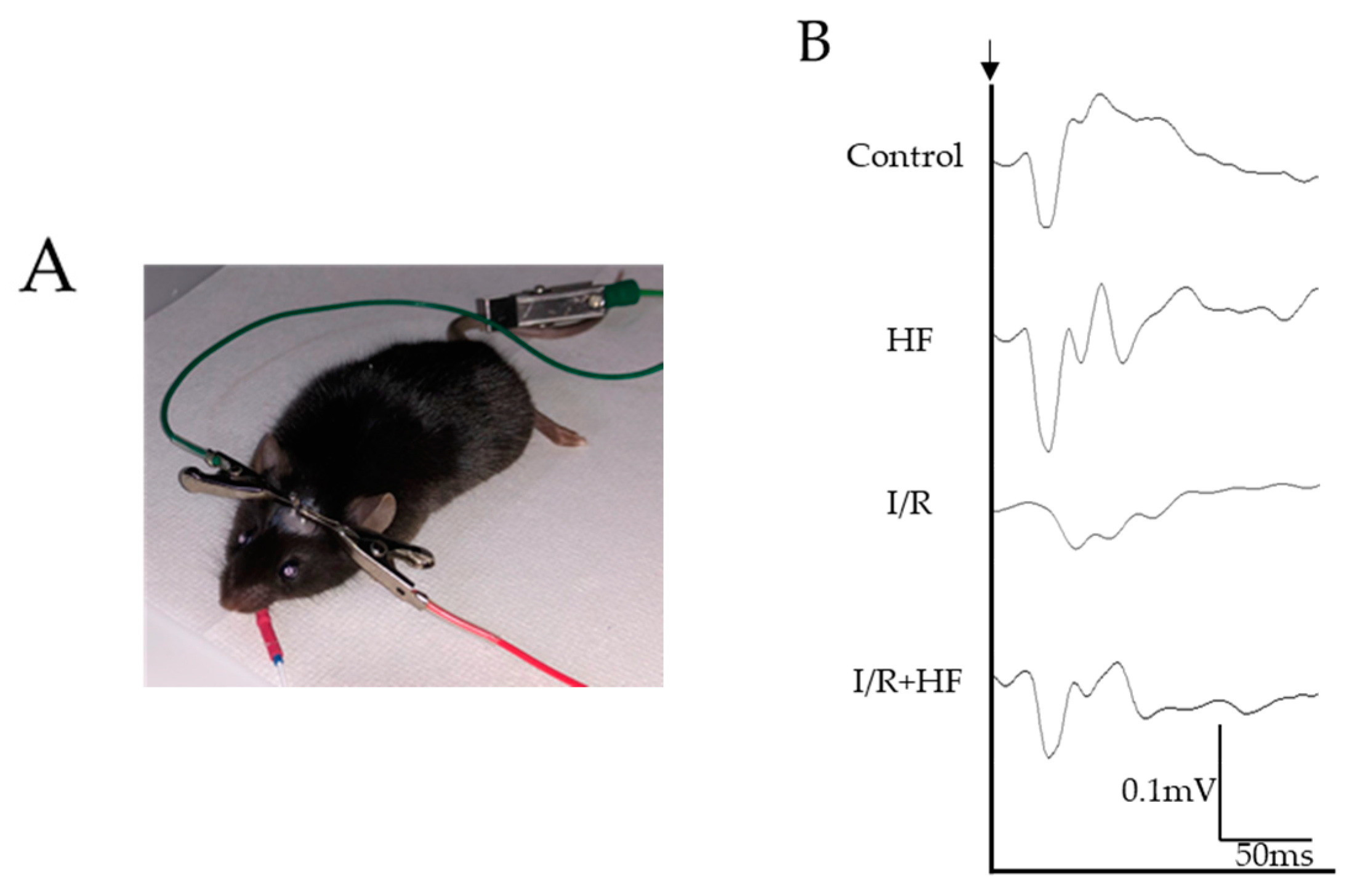
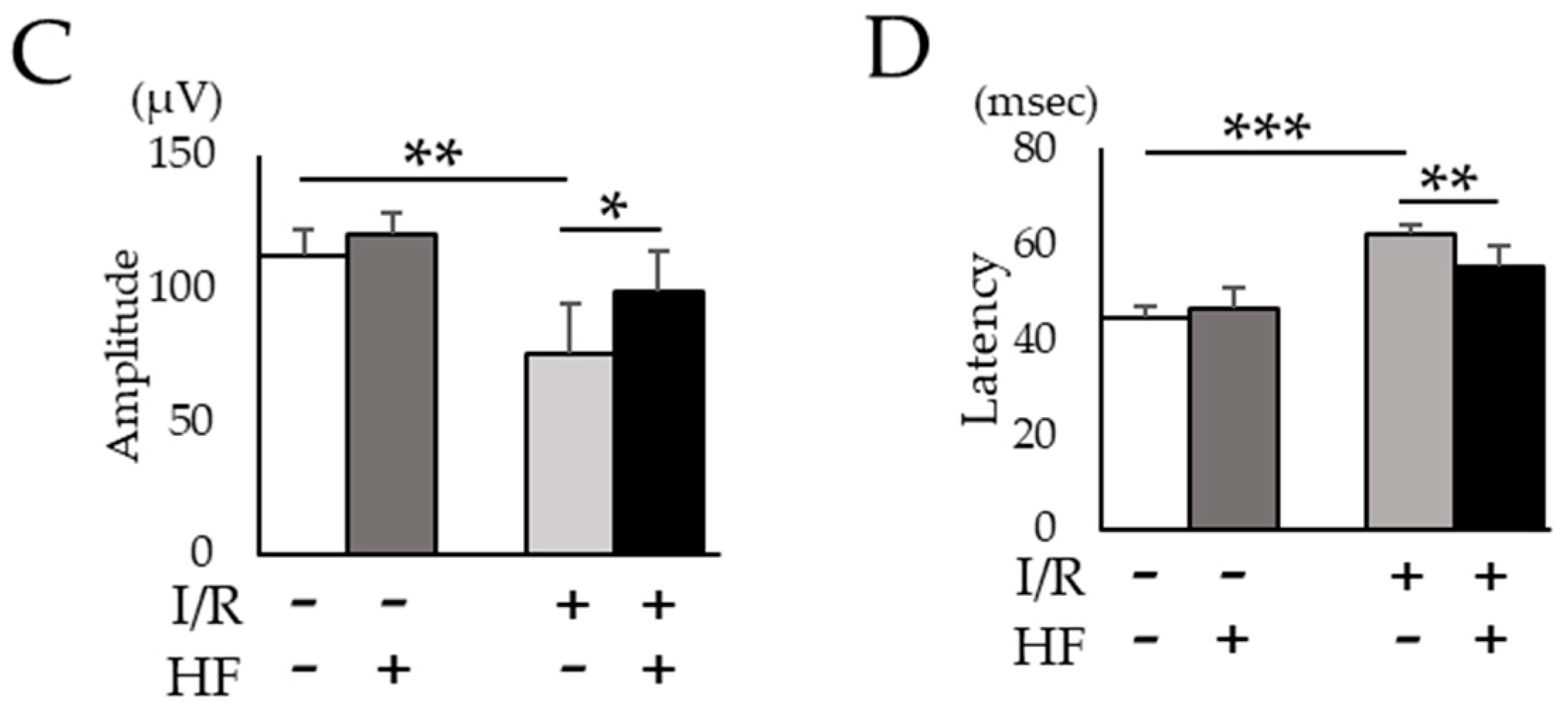
2.5. Halofuginone Administration Protects I/R-Damaged Retina Electrophysiologically
3. Discussion
4. Materials and Methods
4.1. Luciferase Assay for Drug Screening
4.2. Western Blotting
4.3. RNA Extraction and qPCR
4.4. Animals
4.5. Retinal Ischemia/Reperfusion (I/R) Injury
4.6. Evaluation of Retinal Thickness
4.7. Retrograde Labeling of Retinal Ganglion Cells (RGCs)
4.8. Electrophysiological Examination
4.9. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HIF | Hypoxia inducible factor |
| PCR | Polymerase chain reaction |
| EGR1 | Early growth response protein 1 |
| I/R | Ischemia-reperfusion |
| CoCl2 | Cobalt (II) chloride hexahydrate |
| VEGF-A | Vascular endothelial growth factor A |
| GLUT1 | Glucose transporter 1 |
| PDK1 | Phosphoinositide-dependent kinase 1 |
| OCT | Optic coherence tomography |
| RGC | Retinal ganglion cell |
| ERG | Electroretinogram |
| VEP | Visual evoked potential |
| CMV | Cytomegalovirus |
| PBS | Phosphate Buffered Saline |
| IOP | Intraocular pressure |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Products | Relative Intensity |
|---|---|
| Hydrangea stalk extract | 0.03 |
| Narcissus leaf extract | 0.03 |
| Hydrangea petal and calyx extract | 0.04 |
| Narcissus bulb extract | 0.06 |
| Geranium root extract | 0.20 |
| Camellia leaf extract | 0.23 |
| Growth factor concentrate | 0.26 |
| Camellia ‘Beniwabisuke’ calyx extract | 0.26 |
| Japanese butterfish extract | 0.29 |
| Nettle extract | 0.29 |
| Broccoli root extract | 0.31 |
| Geranium stalk extract | 0.34 |
| Fish protein | 0.35 |
| Turmeric extract | 0.36 |
| Tea plant branch extract | 0.37 |
| Saffron cod egg extract | 0.47 |
| Mango seed extract | 0.61 |
| Camellia ‘Hatsuarashi’ stamen and pistil extract | 0.62 |
| Echinocactus grusonii var.albispinus root extract | 0.63 |
| Camellia ‘Hatsuarashi’ petal extract | 0.80 |
| Sansevieria extract | 0.80 |
| Wax gourd flesh and seed extract | 0.86 |
| Bio-active shark cartilage powder | 0.90 |
| Japanese pepper extract | 0.94 |
| Camellia stamen and pistil extract | 1.02 |
| Taheebo tea | 1.05 |
| Canadian goldenrod root extract | 1.11 |
| Japanese cleyera stalk extract | 1.24 |
| Camellia ‘Hatsuarashi’ calyx extract | 1.27 |
| Banded blue sprat extract | 1.30 |
| Chinese fringe tree leaf extract | 1.31 |
| Saxifrage root extract | 1.53 |
| Aso pickled mustard leaf extract | 1.53 |
| Poria Sclerotium extract | 1.56 |
| Baked flying fish extract | 1.59 |
| Plaster | 1.63 |
| Taro leaf extract | 1.65 |
| Almond shell extract | 1.67 |
| Tulip leaf extract | 1.70 |
| Camellia ‘Koiso’ calyx extract | 1.72 |
| Sea grapes extract | 1.73 |
| Wild sesame extract | 1.74 |
| Green soybeans pod extract | 1.80 |
| Radish leaf extract | 1.81 |
| Camellia ‘Beniwabisuke’ petal extract | 1.84 |
| Nam prik (pepper seasoning) extract | 1.85 |
| Blue tea | 1.86 |
| Radish fleash extract | 1.87 |
| Hosta sieboldiana extract | 1.87 |
| Tulip petal extract | 1.89 |
| Tea plant leaf extract | 1.91 |
| Viola leaf extract | 1.97 |
| Baby leaf extract | 1.99 |
| Ginseng root extract | 2.00 |
| Spinach leaf extract | 2.01 |
| Taro bulb extract | 2.01 |
| Passionflower extract | 2.09 |
| Golden brown rice tea | 2.10 |
| Allium root extract | 2.10 |
| Viola stalk extract | 2.11 |
| Boldo tea | 2.11 |
| Camellia ‘Koiso’ petal extract | 2.12 |
| Hemp palm leaf extract | 2.12 |
| Green tea | 2.14 |
| Brown rice germ extract | 2.15 |
| Monostroma nitidum extract | 2.18 |
| Soybean sprout extract | 2.19 |
| Chinese chive extract | 2.20 |
| Corn flour extract | 2.30 |
| Japanese dock root extract | 2.37 |
| Potherb mustard extract | 2.38 |
| Tulip stalk extract | 2.42 |
| Tree onion (lower part) extract | 2.49 |
| Geranium leaf extract | 2.49 |
| Long pepper (dry) extract | 2.58 |
| Pachira extract | 2.60 |
| Squid ink dye | 2.63 |
| Citrus hassaku leaf extract | 2.66 |
| Topotecan* | 2.68 |
| Camellia branch extract | 2.69 |
| Doxorubicin* | 2.70 |
| Spinach stalk extract | 2.73 |
| Buckwheat seed extract | 2.81 |
| Yuzu leaf extract | 2.81 |
| Red bayberry leaf extract | 2.83 |
| Sunflower leaf extract | 2.92 |
| Green soybeans root extract | 2.93 |
| Japanese yam bulbil extract | 2.95 |
| Asiatic dayflower stalk extract | 2.96 |
| Chlorophyll20 | 3.00 |
| Cedar leaf extract | 3.01 |
| Elder extract | 3.02 |
| Sunflower stalk extract | 3.03 |
| Canadian goldenrod leaf extract | 3.06 |
| Curry plant extract | 3.09 |
| Canola flower seed extract | 3.09 |
| Romanesco broccoli petal extract | 3.09 |
| White clover leaf extract | 3.11 |
| Oyster extract | 3.13 |
| Pen shell extract | 3.17 |
| Carqueja tea | 3.18 |
| Japanese dock spike extract | 3.20 |
| Valerian extract | 3.20 |
| Coix seed extract | 3.23 |
| Safflower extract | 3.23 |
| Petit vert extract | 3.24 |
| Parsley extract | 3.29 |
| Satsuma (orange) extract | 3.29 |
| Tree onion (upper part) extract | 3.34 |
| Kyoho grape branch extract | 3.34 |
| Yuzu seed extract | 3.36 |
| Pine tree leaf extract | 3.37 |
| Canadian goldenrod stalk extract | 3.38 |
| Italian parsley extract | 3.39 |
| Prune seed extract | 3.45 |
| Japanese apricot seed coat extract | 3.54 |
| Broccoli leaf extract | 3.57 |
| Acidanthera bulb extract | 3.58 |
| Trachycarpus stalk extract | 3.60 |
| American ginseng extract | 3.64 |
| Baby corn extract | 3.69 |
| Corn silk extract | 3.73 |
| Crown daisy stalk extract | 3.77 |
| Yuzu peel extract | 3.77 |
| Su-Wu tea | 3.80 |
| Salix gracilistyla stalk extract | 3.84 |
| Yellowtail egg extract | 3.86 |
| Japanese Aucuba stalk extract | 3.86 |
| Japanese cleyera leaf extract | 3.88 |
| dandelion extract | 3.90 |
| Colocasia gigantea bulb extract | 3.90 |
| Goji chrysanthemum tea | 3.92 |
| Flax seed extract | 3.93 |
| Paeonia lactiflora extract | 3.97 |
| Sunflower patel extract | 3.98 |
| Japanese dock leaf extract | 4.01 |
| Nalta jute noodle extract | 4.01 |
| Yacón extract | 4.03 |
| Allium leaf extract | 4.06 |
| Lemon peel extract | 4.11 |
| Rapeseed oil | 4.11 |
| Pumpkin seed extract | 4.14 |
| Abalone powder | 4.16 |
| Oats fiber extract | 4.16 |
| Echinocactus grusonii var.albispinus flesh extract | 4.18 |
| White clover stalk extract | 4.21 |
| Chinese Plantain extract | 4.25 |
| Senna obtusifolia extract | 4.27 |
| Green pea extract | 4.28 |
| Echinacea extract | 4.28 |
| Rheum extract | 4.30 |
| Hemp Seed extract | 4.34 |
| Lime peel extract | 4.37 |
| Giant Butterbur stalk extract | 4.38 |
| Japanese hawthorn leaf extract | 4.39 |
| Green leaf extract | 4.48 |
| Florist’s Cineraria stalk extract | 4.48 |
| Salix gracilistyla leaf extract | 4.51 |
| Viola petal extract | 4.52 |
| Green shell mussel powder | 4.54 |
| Sea mustard root extract | 4.54 |
| Panax notoginseng (powder) | 4.57 |
| Canola flower stalk extract | 4.63 |
| Broad bean extract | 4.63 |
| Bay laurel leaf extract | 4.65 |
| Tulip root extract | 4.67 |
| Sunflower stamen and pistil extract | 4.68 |
| Common sage extract | 4.77 |
| White clover petal extract | 4.79 |
| Young corn extract | 4.86 |
| Marguerite stalk extract | 4.93 |
| Loquat leaf extract | 4.94 |
| Marguerite petal extract | 4.97 |
| Florist’s Cineraria root extract | 4.97 |
| Sweetleaf extract | 5.04 |
| Black tea | 5.05 |
| Catnip extract | 5.06 |
| Broad bean shell extract | 5.06 |
| Marjoram extract | 5.26 |
| Red Shiso stalk extract | 5.38 |
| Japanese wisteria stalk extract | 5.40 |
| Ginseng root and astragalus root tea | 5.40 |
| Lilium ‘Casa Blanca’ bulb extract | 5.41 |
| Tarragon extract | 5.42 |
| Florist’s Cineraria petal extract | 5.45 |
| Crown daisy leaf extract | 5.50 |
| Swiss chard leaf extract | 5.50 |
| Broccoli flower extract | 5.51 |
| Pistachio nut extract | 5.64 |
| CoCl2 | 5.66 |
| Walnut shell powder | 5.67 |
| Bush groundsel tea | 5.71 |
| Japanese hawthorn flesh extract | 5.71 |
| savory extract | 5.73 |
| Asiatic dayflower leaf extract | 5.92 |
| Fish collagen | 5.93 |
| Swertia japonica extract | 6.00 |
| Japanese Aucuba leaf extract | 6.07 |
| Fragrant orange-colored olive leaf extract | 6.24 |
| Oolong tea | 6.24 |
| Marguerite leaf extract | 6.26 |
| Camellia petal extract | 6.34 |
| Hyssop extract | 6.36 |
| Peppermint stalk extract | 6.70 |
| Simon leaf extract | 6.71 |
| Pistachio shell extract | 6.80 |
| Larvae of a bee extract | 6.83 |
| Saxifrage leaf extract | 6.90 |
| Camellia ‘Koiso’ stamen and pistil extract | 7.03 |
| Saxifrage stalk extract | 7.12 |
| Eye bright tea | 7.26 |
| Field Horsetail extract | 7.29 |
| Pineapple enzyme | 7.32 |
| Giant Butterbur leaf extract | 7.41 |
| Peppermint leaf extract | 7.49 |
| Chinese Plantain seed extract | 7.56 |
| Japanese photinia leaf extract | 7.59 |
| Green oolong tea | 7.68 |
| Eucalyptus stalk extract | 7.77 |
| Cleyera japonica leaf extract | 7.85 |
| Açaí berry purée | 7.86 |
| Red Shiso leaf extract | 8.04 |
| Henbit extract | 8.08 |
| Roselle tea | 8.14 |
| Western bracken fern extract | 8.19 |
| Cashew nut extract | 8.27 |
| Azalea petal extract | 8.47 |
| Florist’s Cineraria leaf extract | 8.75 |
| Japanese dock stalk extract | 8.79 |
| Eucalyptus leaf extract | 9.19 |
| Japanese photinia branch extract | 9.59 |
| Organic oolong tea | 10.10 |
| Colocasia gigantea stalk extract | 10.36 |
| Propolis powder | 10.37 |
| Fennel extract | 11.16 |
| Camellia bud extract | 11.91 |
| Organic coffee | 12.54 |
| Dayuling oolong tea | 13.03 |
| Rubber tree leaf extract | 14.56 |
| Japanese wisteria leaf extract | 25.22 |
| Sunflower seed extract | 56.48 |
| Products | Relative Intensity |
|---|---|
| Hydrangea stalk extract | 0.02 |
| Hydrangea petal and calyx extract | 0.03 |
| Narcissus leaf extract | 0.06 |
| Narcissus bulb extract | 0.09 |
| Camellia ‘Koiso’ calyx extract | 0.18 |
| Geranium root extract | 0.23 |
| Fish protein | 0.24 |
| Turmeric extract | 0.31 |
| Growth factor concentrate | 0.33 |
| Taheebo tea | 0.45 |
| Camellia ‘Beniwabisuke’ calyx extract | 0.46 |
| Broccoli root extract | 0.47 |
| Bio-active shark cartilage powder | 0.48 |
| Red bayberry leaf extract | 0.52 |
| Echinocactus grusonii var.albispinus root extract | 0.61 |
| Mango seed extract | 0.67 |
| Aso pickled mustard leaf extract | 0.72 |
| Tea plant branch extract | 0.73 |
| Japanese butterfish extract | 0.78 |
| Prune seed extract | 0.80 |
| Kyoho grape branch extract | 0.81 |
| Saxifrage root extract | 0.97 |
| Japanese yam bulbil extract | 0.98 |
| Salix gracilistyla leaf extract | 1.11 |
| Banded blue sprat extract | 1.14 |
| Poria Sclerotium extract | 1.14 |
| Green soybeans pod extract | 1.21 |
| Camellia ‘Hatsuarashi’ stamen and pistil extract | 1.26 |
| Camellia ‘Hatsuarashi’ petal extract | 1.28 |
| Spinach leaf extract | 1.31 |
| Wax gourd flesh and seed extract | 1.33 |
| Nettle extract | 1.35 |
| Radish leaf extract | 1.40 |
| Japanese cleyera stalk extract | 1.42 |
| Nam prik (pepper seasoning) extract | 1.44 |
| Broccoli root extract | 1.45 |
| Radish fleash extract | 1.57 |
| Plaster | 1.74 |
| Baked flying fish extract | 1.80 |
| Boldo tea | 1.82 |
| Almond shell extract | 1.91 |
| Passionflower extract | 1.91 |
| Taro bulb extract | 1.98 |
| Taro leaf extract | 2.01 |
| Monostroma nitidum extract | 2.16 |
| Blue tea | 2.29 |
| Japanese pepper extract | 2.32 |
| Cedar leaf extract | 2.37 |
| Wild sesame extract | 2.40 |
| Tea plant leaf extract | 2.41 |
| Geranium stalk extract | 2.50 |
| Tulip leaf extract | 2.66 |
| Potherb mustard extract | 2.66 |
| Ginseng root extract | 2.69 |
| Camellia ‘Beniwabisuke’ petal extract | 2.72 |
| Tea plant branch extract | 2.88 |
| Topotecan* | 2.90 |
| Sansevieria extract | 3.12 |
| Allium root extract | 3.18 |
| Doxorubicin* | 3.25 |
| Hemp palm leaf extract | 3.39 |
| Golden brown rice tea | 3.41 |
| Chinese fringe tree leaf extract | 3.52 |
| Saffron cod egg extract | 3.55 |
| Pachira extract | 3.57 |
| Viola leaf extract | 3.60 |
| Japanese dock root extract | 3.88 |
| Camellia leaf extract | 4.04 |
| Camellia ‘Koiso’ petal extract | 4.08 |
| Sea grapes extract | 4.30 |
| Squid ink dye | 4.67 |
| Tulip petal extract | 4.80 |
| Camellia ‘Beniwabisuke’ calyx extract | 4.93 |
| Tulip stalk extract | 4.95 |
| Asiatic dayflower stalk extract | 5.78 |
| Camellia ‘Hatsuarashi’ calyx extract | 6.27 |
| CoCl2 | 6.64 |
| Long pepper (dry) extract | 6.69 |
| Camellia stamen and pistil extract | 12.08 |
| Saxifrage root extract | 13.04 |
| Green tea | 23.69 |
References
- Semenza, G.L. Regulation of Metabolism by Hypoxia-Inducible Factor 1. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, H.; Yu, A.Y.; Della, N.; Ozaki, K.; Luna, J.D.; Yamada, H.; Hackett, S.F.; Okamoto, N.; Zack, D.J.; Semenza, G.L.; et al. Hypoxia inducible factor-1α is increased in ischemic retina: Temporal and spatial correlation with VEGF expression. Investig. Ophthalmol. Vis. Sci. 1999, 40, 182–189. [Google Scholar]
- Tezel, G.; Wax, M.B. Hypoxia-inducible factor 1α in the glaucomatous retina and optic nerve head. Arch. Ophthalmol. 2004, 122, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Rapisarda, A.; Uranchimeg, B.; Scudiero, D.A.; Selby, M.; Sausville, E.A.; Shoemaker, R.H.; Melillo, G. Identification of small molecule inhibitors of hypoxia-inducible factor 1 transcriptional activation pathway. Cancer Res. 2002, 62, 4316–4324. [Google Scholar] [PubMed]
- Lee, K.; Qian, D.Z.; Rey, S.; Wei, H.; Liu, J.O.; Semenza, G.L. Anthracycline chemotherapy inhibits HIF-1 transcriptional activity and tumor-induced mobilization of circulating angiogenic cells. Proc. Natl. Acad. Sci. USA 2009, 106, 2353–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.; Kersten, S. Nutrigenomics: Goals and strategies. Nat. Rev. Genet. 2003, 4, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P. Nutritional genomics – “Nutrigenomics”. Br. J. Nutr. 2003, 89, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Chuaqui, R.F.; Bonner, R.F.; Best, C.J.M.; Gillespie, J.W.; Flaig, M.J.; Hewitt, S.M.; Phillips, J.L.; Krizman, D.B.; Tangrea, M.A.; Ahram, M.; et al. Post-analysis follow-up and validation of microarray experiments. Nat. Genet. 2002, 32, 509. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, C. Application of Nutrigenomics in Eye Health. In Forum of Nutrition; Karger Publishers: Basel, Switzerland, 2007; Volume 60, pp. 168–175. ISBN 1660-0347. [Google Scholar]
- Fine, I.H. A Randomized, Placebo-Controlled, Clinical Trial of High-Dose Supplementation with Vitamins C and E and Beta-Carotene for Age-Related Cataract and Vision Loss. Evidence-Based Eye Care 2003, 3, 80–81. [Google Scholar] [CrossRef]
- Schalch, W.; Chylack, L.T. Antioxidative Mikronährstoffe und Katarakt. Der. Ophthalmol. 2003, 100, 181–189. [Google Scholar] [CrossRef]
- Dátilo, M.N.; Sant’Ana, M.R.; Formigari, G.P.; Rodrigues, P.B.; de Moura, L.P.; da Silva, A.S.R.; Ropelle, E.R.; Pauli, J.R.; Cintra, D.E. Omega-3 from Flaxseed Oil Protects Obese Mice Against Diabetic Retinopathy Through GPR120 Receptor. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Kurihara, T.; Miyauchi, M.; Ishida, A.; Jiang, X.; Ikeda, S.; Torii, H.; Tsubota, K. Oral crocetin administration suppressed refractive shift and axial elongation in a murine model of lens-induced myopia. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pines, M.; Nagler, A. Halofuginone: A novel antifibrotic therapy. Gen. Pharmacol. 1998, 30, 445–450. [Google Scholar] [CrossRef]
- McGaha, T.L.; Kodera, T.; Spiera, H.; Stan, A.C.; Pines, M.; Bona, C.A. Halofuginone inhibition of COL1A2 promoter activity via a c-Jun-dependent mechanism. Arthritis Rheum. 2002, 46, 2748–2761. [Google Scholar] [CrossRef] [PubMed]
- Levi-Schaffer, F.; Nagler, A.; Slavin, S.; Knopov, V.; Pines, M. Inhibition of collagen synthesis and changes in skin morphology in murine graft-versus-host disease and tight skin mice: Effect of halofuginone. J. Invest. Dermatol. 1996, 106, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Sundrud, M.S.; Koralov, S.B.; Feuerer, M.; Calado, D.P.; Elhed, A.; Rhule-smith, A.; Lefebvre, R.E.; Unutmaz, D.; Waldner, H.; Whitman, M.; et al. Halofuginone inhibits TH17 cell differentiation by activating the amino acid starvation response. Science. 2010, 324, 1334–1338. [Google Scholar] [CrossRef] [PubMed]
- Pines, M.; Halevy, O. Halofuginone and muscular dystrophy. Histol. Histopathol. 2011, 26, 135–146. [Google Scholar]
- Nagler, A.; Ohana, M.; Shibolet, O.; Shapira, M.Y.; Alper, R.; Vlodavsky, I.; Pines, M.; Ilan, Y. Suppression of hepatocellular carcinoma growth in mice by the alkaloid coccidiostat halofuginone. Eur. J. Cancer 2004, 40, 1397–1403. [Google Scholar] [CrossRef]
- Taras, D.; Blanc, J.-F.; Rullier, A.; Dugot-Senant, N.; Laurendeau, I.; Bièche, I.; Pines, M.; Rosenbaum, J. Halofuginone suppresses the lung metastasis of chemically induced hepatocellular carcinoma in rats through MMP inhibition. Neoplasia 2006, 8, 312–318. [Google Scholar] [CrossRef] [Green Version]
- Gavish, Z.; Pinthus, J.H.; Barak, V.; Ramon, J.; Nagler, A.; Eshhar, Z.; Pines, M. Growth inhibition of prostate cancer xenografts by halofuginone. Prostate 2002, 51, 73–83. [Google Scholar] [CrossRef]
- Gross, D.J.; Reibstein, I.; Weiss, L.; Slavin, S.; Dafni, H.; Neeman, M.; Pines, M.; Nagler, A. Treatment with Halofuginone Results in Marked Growth Inhibition of a von Hippel-Lindau Pheochromocytoma in vivo. Clin. Cancer Res. 2003, 9, 3788–3793. [Google Scholar] [PubMed]
- Pinthus, H.J.; Sheffer, Y.; Nagler, A.; Fridman, E.; Mor, Y.; Genina, O.; Pines, M. Inhibition of Wilms tumor xenograft progression by halofuginone is accompanied by activation of WT-1 gene expression. J. Urol. 2005, 174, 1527–1531. [Google Scholar] [CrossRef] [PubMed]
- Pines, M.; Snyder, D.; Yarkoni, S.; Nagler, A. Halofuginone to treat fibrosis in chronic graft-versus-host disease and scleroderma. Biol. Blood Marrow Transplant. 2003, 9, 417–425. [Google Scholar] [CrossRef] [Green Version]
- 25. Onishi, S.; Meguro, S.; Pervin, M.; Kitazawa, H.; Yoto, A.; Ishino, M.; Shimba, Y.; Mochizuki, Y.; Miura, S.; Tokimitsu, I.; et al. Green Tea Extracts Attenuate Brain Dysfunction in High-Fat-Diet-Fed SAMP8 Mice. Nutrients 2019, 11, 821. [Google Scholar] [CrossRef] [PubMed]
- Faggi, L.; Porrini, V.; Lanzillotta, A.; Benarese, M.; Mota, M.; Tsoukalas, D.; Parrella, E.; Pizzi, M. A Polyphenol-Enriched Supplement Exerts Potent Epigenetic-Protective Activity in a Cell-Based Model of Brain Ischemia. Nutrients 2019, 11, 345. [Google Scholar] [CrossRef] [PubMed]
- Gorji, N.; Moeini, R.; Memariani, Z. Almond, hazelnut and walnut, three nuts for neuroprotection in Alzheimer’s disease: A neuropharmacological review of their bioactive constituents. Pharmacol. Res. 2018, 129, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.L.; Park, H.Y.; Dasilva, N.A.; Vattem, D.A.; Ma, H.; Seeram, N.P. Levodopa-reduced mucuna pruriens seed extract shows neuroprotective effects against parkinson’s disease in murine microglia and human neuroblastoma cells, Caenorhabditis elegans, and Drosophila melanogaster. Nutrients 2018, 10, 1139. [Google Scholar] [CrossRef] [PubMed]
- Altieri, F.; Cairone, F.; Giamogante, F.; Carradori, S.; Locatelli, M.; Chichiarelli, S.; Cesa, S. Influence of Ellagitannins Extracted by Pomegranate Fruit on Disulfide Isomerase PDIA3 Activity. Nutrients 2019, 11, 186. [Google Scholar] [CrossRef]
- Locri, F.; Cammalleri, M.; Pini, A.; Dal Monte, M.; Rusciano, D.; Bagnoli, P. Further Evidence on Efficacy of Diet Supplementation with Fatty Acids in Ocular Pathologies: Insights from the EAE Model of Optic Neuritis. Nutrients 2018, 10, 1447. [Google Scholar] [CrossRef]
- Lu, C.; Wang, Y.; Wang, D.; Zhang, L.; Lv, J.; Jiang, N.; Fan, B.; Liu, X.; Wang, F. Neuroprotective effects of soy isoflavones on scopolamine-induced amnesia in mice. Nutrients 2018, 10, 853. [Google Scholar] [CrossRef] [PubMed]
- Miwa, Y.; Hoshino, Y.; Shoda, C.; Jiang, X.; Tsubota, K.; Kurihara, T. Pharmacological HIF inhibition prevents retinal neovascularization with improved visual function in a murine oxygen-induced retinopathy model. Neurochem. Int. 2019, 128, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Wang, L.L.; Abdelmaksoud, S.; Aboelgheit, A.; Saeed, S.; Zhang, C.L. Minocycline modulates microglia polarization in ischemia-reperfusion model of retinal degeneration and induces neuroprotection. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Schultz, R.; Krug, M.; Precht, M.; Wohl, S.G.; Witte, O.W.; Schmeer, C. Frataxin overexpression in Müller cells protects retinal ganglion cells in a mouse model of ischemia/reperfusion injury in vivo. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunimi, H.; Miwa, Y.; Inoue, H.; Tsubota, K.; Kurihara, T. A Novel HIF Inhibitor Halofuginone Prevents Neurodegeneration in a Murine Model of Retinal Ischemia-Reperfusion. Int. J. Mol. Sci. 2019, 20, 3171. https://doi.org/10.3390/ijms20133171
Kunimi H, Miwa Y, Inoue H, Tsubota K, Kurihara T. A Novel HIF Inhibitor Halofuginone Prevents Neurodegeneration in a Murine Model of Retinal Ischemia-Reperfusion. International Journal of Molecular Sciences. 2019; 20(13):3171. https://doi.org/10.3390/ijms20133171
Chicago/Turabian StyleKunimi, Hiromitsu, Yukihiro Miwa, Hiroyoshi Inoue, Kazuo Tsubota, and Toshihide Kurihara. 2019. "A Novel HIF Inhibitor Halofuginone Prevents Neurodegeneration in a Murine Model of Retinal Ischemia-Reperfusion" International Journal of Molecular Sciences 20, no. 13: 3171. https://doi.org/10.3390/ijms20133171