Ouabain Enhances Cell-Cell Adhesion Mediated by β1 Subunits of the Na+,K+-ATPase in CHO Fibroblasts

 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
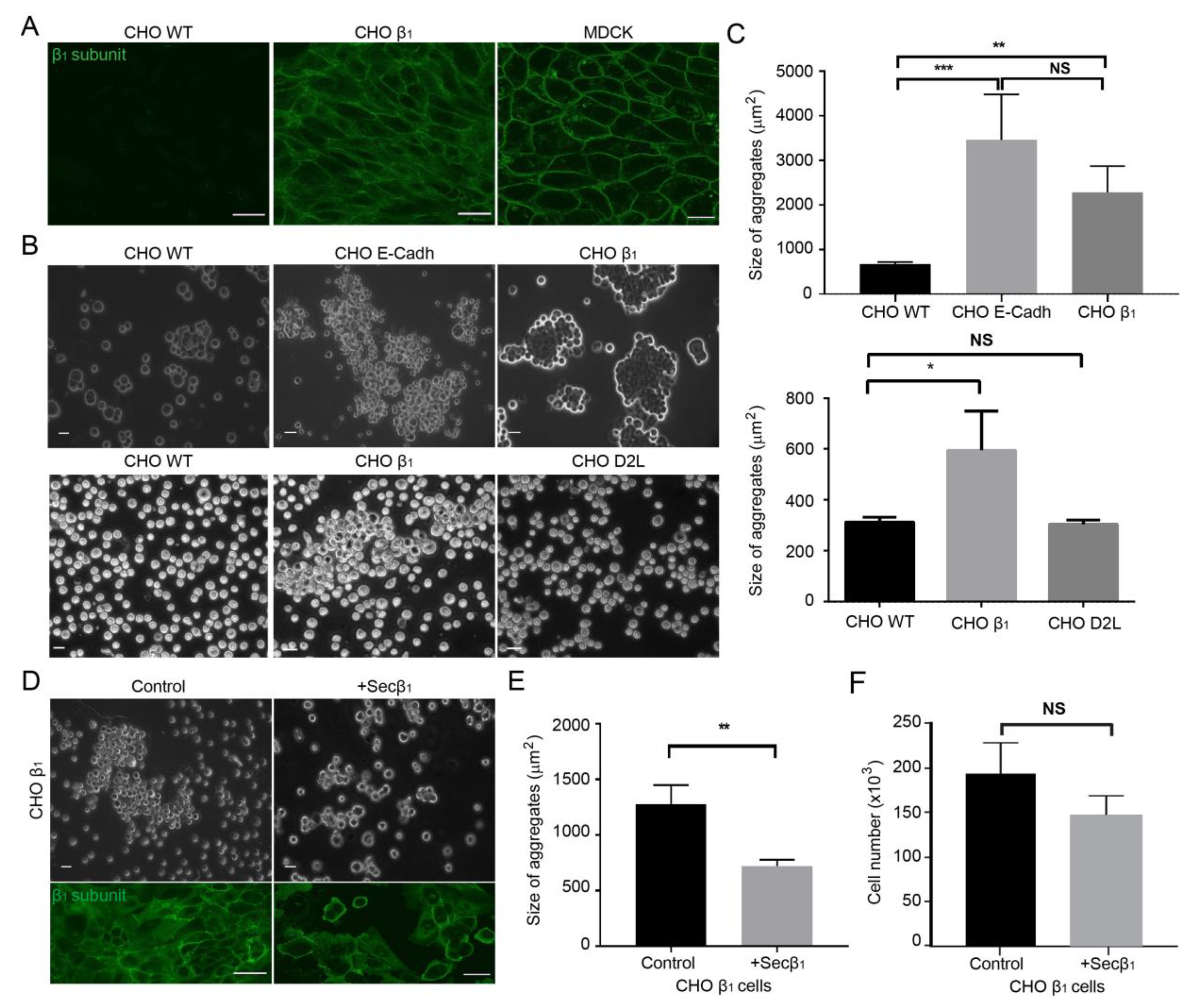
2.1. Cell-Cell Adhesion of CHO Fibroblasts Expressing Canine β1 Subunit of Na+,K+-ATPase is Mediated by β1 Homotypic Interactions in-trans
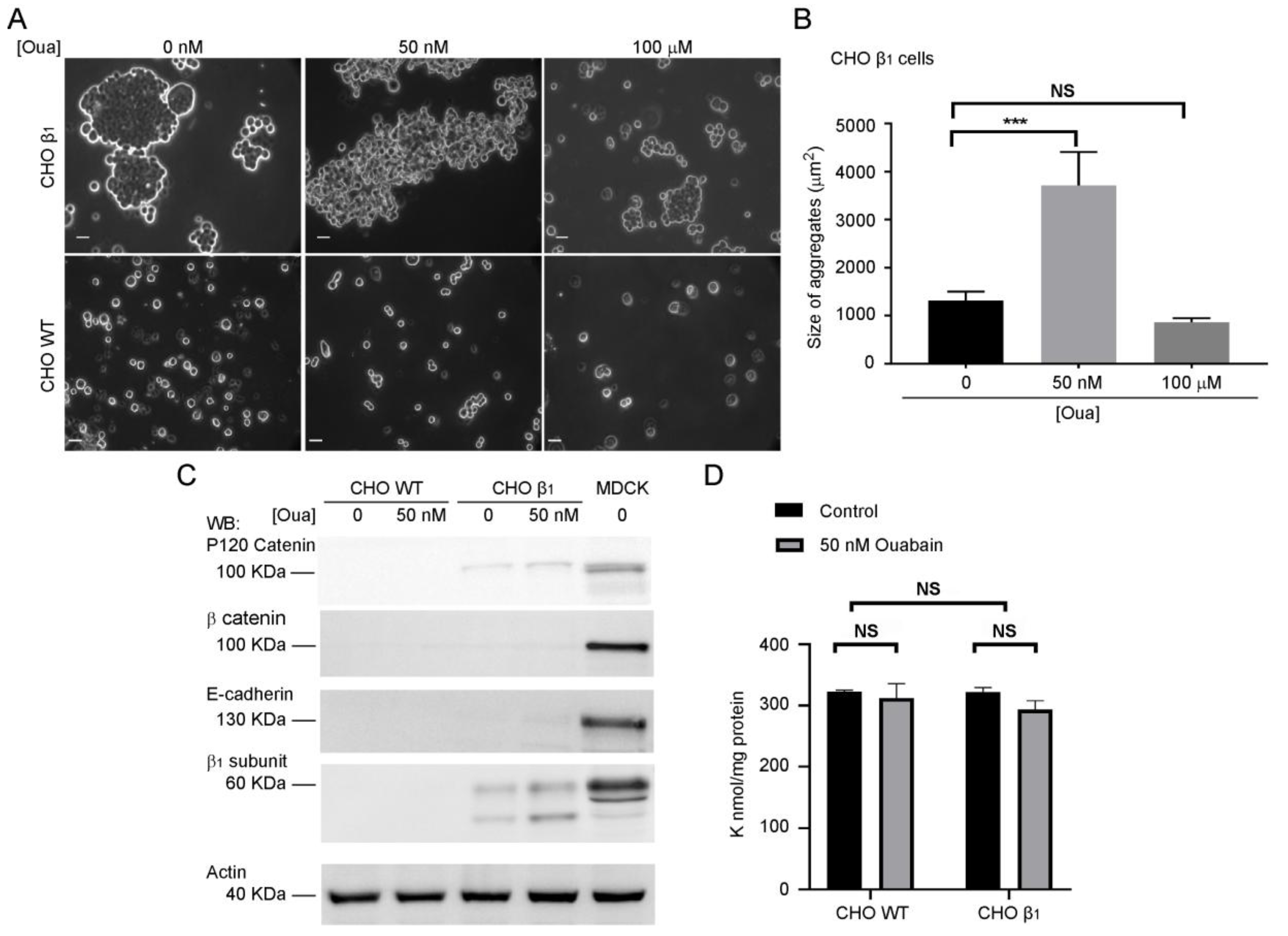
2.2. Ouabain Increases Cell-Cell Adhesion of CHO β1 Cells
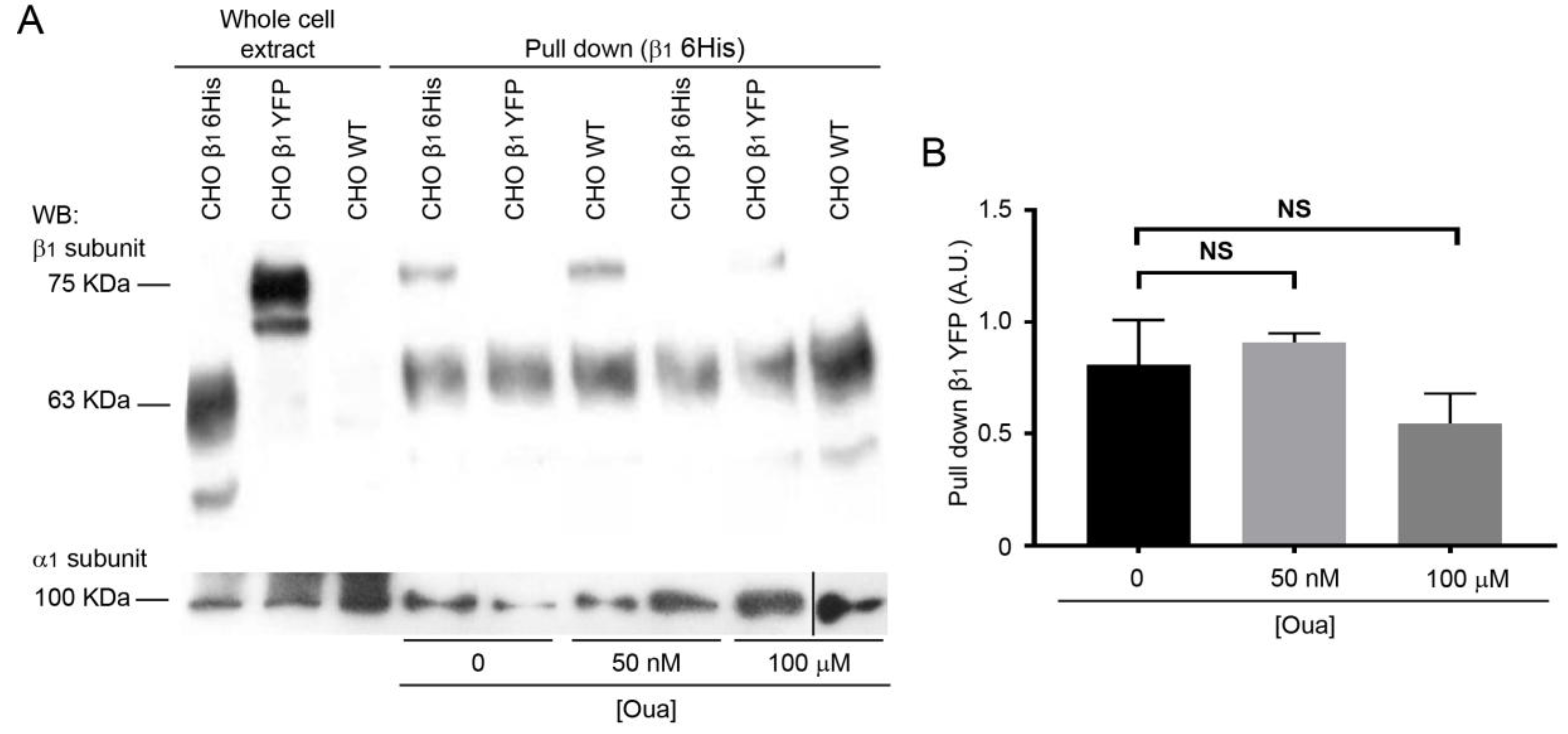
2.3. The Interactions of β1-β1 Subunits are Stable in vitro Independently of Ouabain Treatment
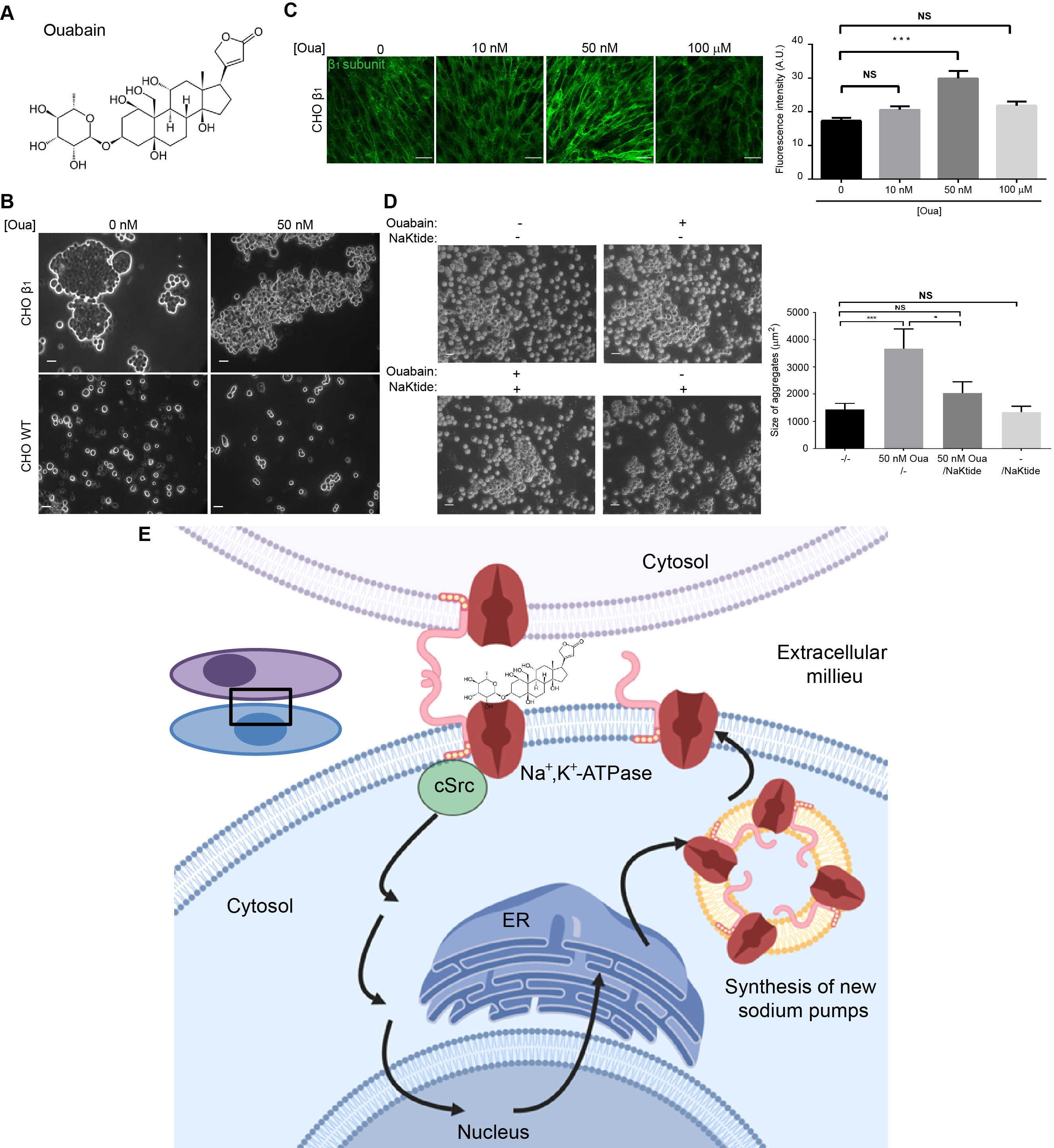
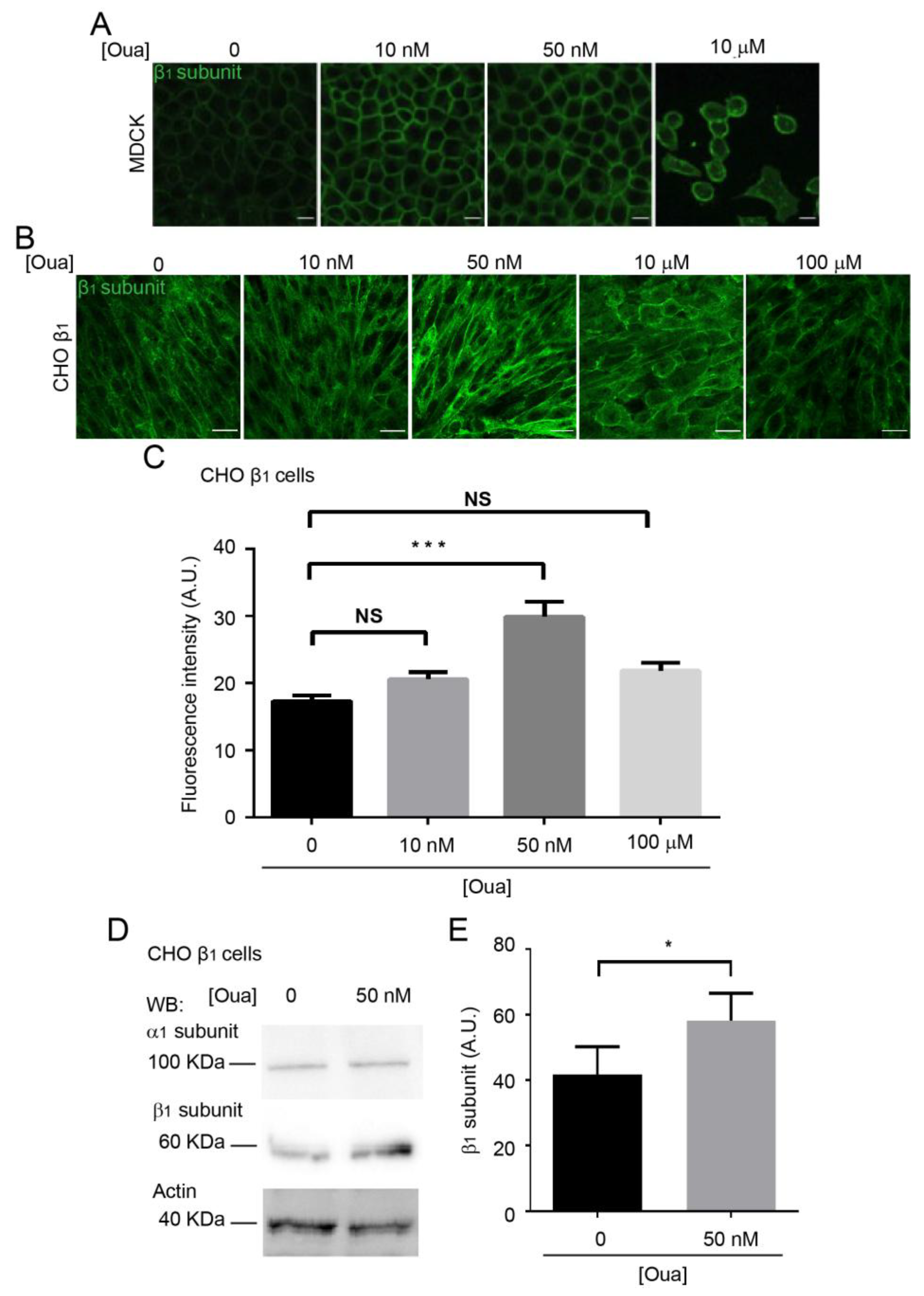
2.4. Ouabain Increases the Expression and Localization of the Sodium Pump at the Plasma Membrane
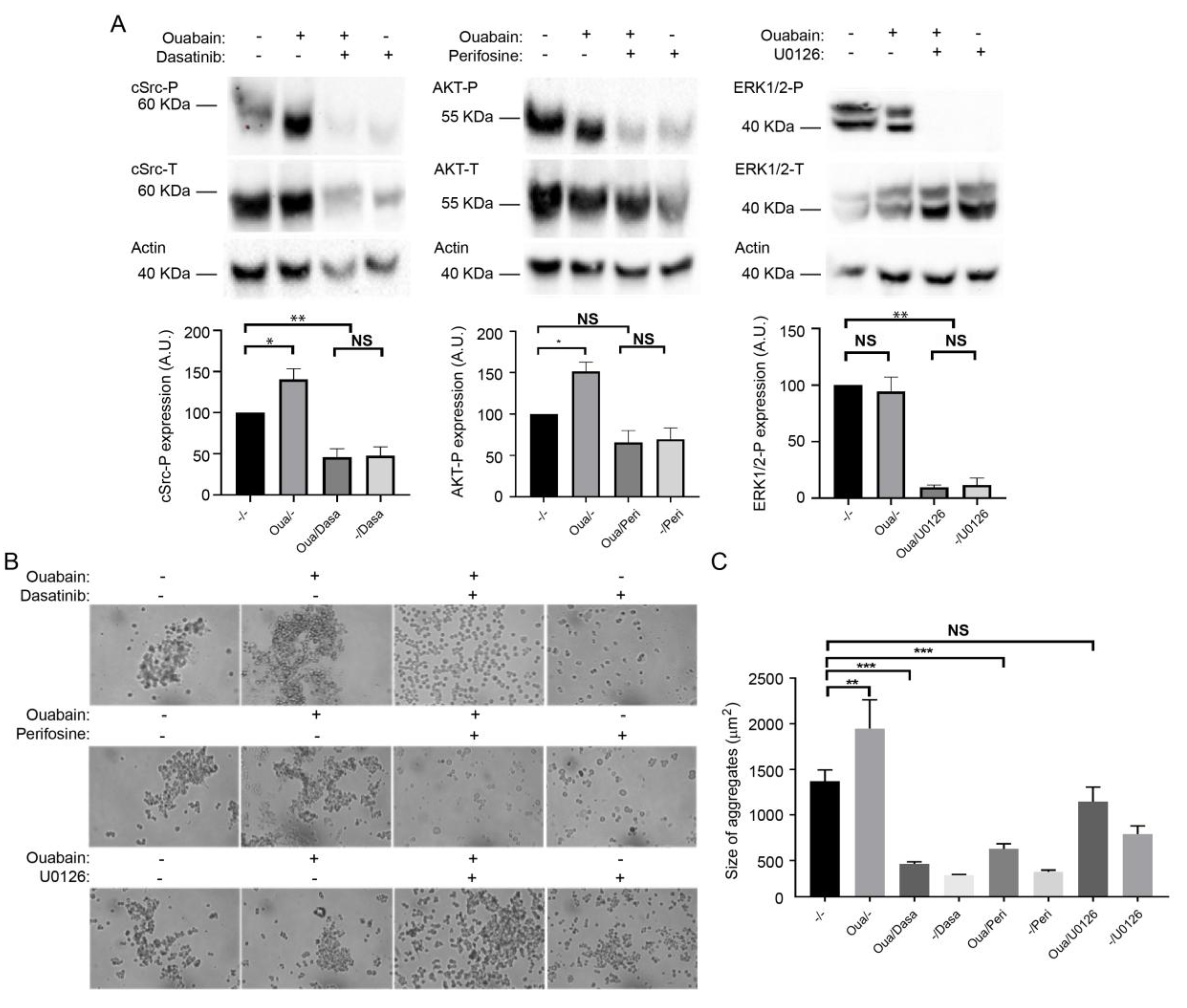
2.5. The β1-β1 Adhesion Enhanced by Ouabain Depends on the Activation of cSrc and AKT Signaling Pathway
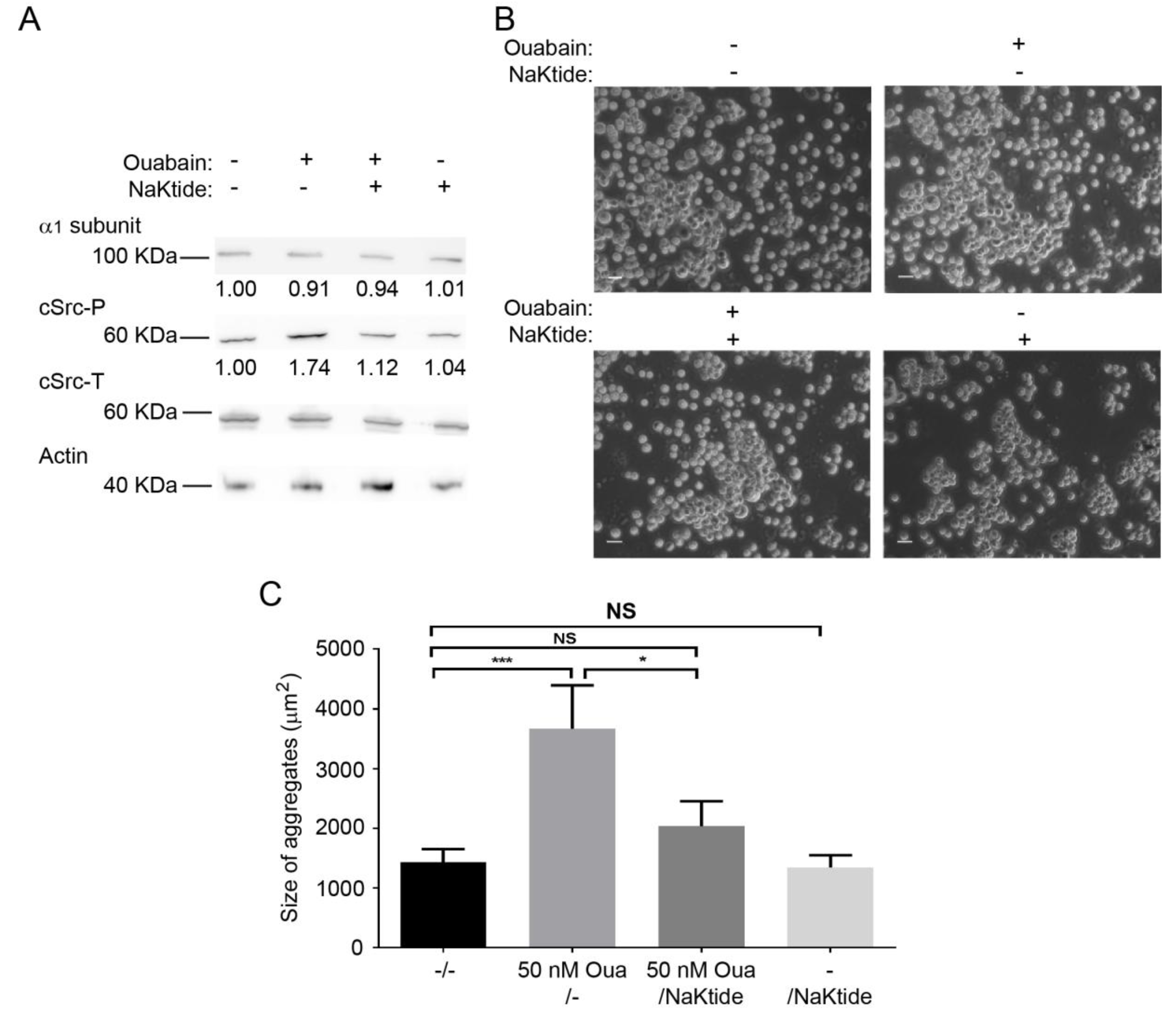
2.6. pNaKtide Reduces Cell Adhesion Effect Exerted by Ouabain
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Antibodies
4.3. Immunofluorescence and Confocal Microscopy Analyses
4.4. Western Blot Analysis
4.5. Cell Surface Biotinylation
4.6. Cell Adhesion Assay (Dispase Assay)
4.7. Pull Down Assay (PD)
4.8. Whole Cell K+ and Na+ Determinations by Atomic Absorbance Spectroscopy
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Skou, J.C. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochim. Biophys. Acta 1957, 23, 394–401. [Google Scholar] [CrossRef]
- Blanco, G.; Mercer, R.W. Isozymes of the Na-K-atpase: Heterogeneity in structure, diversity in function. Am. J. Physiol. 1998, 275, F633–F650. [Google Scholar]
- Post, R.L.; Sen, A.K.; Rosenthal, A.S. A phosphorylated intermediate in adenosine triphosphate-dependent sodium and potassium transport across kidney membranes. J. Biol. Chem. 1965, 240, 1437–1445. [Google Scholar]
- Pedersen, P.L.; Carafoli, E. Ion motive atpases. I. Obiquity, properties, and significance to cell function. Trends Biochem. Sci. 1987, 12, 146–150. [Google Scholar]
- Post, R.L.; Hegyvary, C.; Kume, S. Activation by adenosine triphosphate in the phosphorylation kinetics of sodium and potassium ion transport adenosine triphosphatase. J. Biol. Chem. 1972, 247, 6530–6540. [Google Scholar]
- Therien, A.G.; Blostein, R. Mechanisms of sodium pump regulation. Am. J. Physiol. Cell Physiol. 2000, 279, C541–C566. [Google Scholar] [CrossRef]
- Morth, J.P.; Pedersen, B.P.; Toustrup-Jensen, M.S.; Sorensen, T.L.; Petersen, J.; Andersen, J.P.; Vilsen, B.; Nissen, P. Crystal structure of the sodium-potassium pump. Nature 2007, 450, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Geering, K. Functional roles of Na,K-atpase subunits. Curr. Opin. Nephrol. Hypertens. 2008, 17, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Bagrov, A.Y.; Shapiro, J.I.; Fedorova, O.V. Endogenous cardiotonic steroids: Physiology, pharmacology, and novel therapeutic targets. Pharmacol. Rev. 2009, 61, 9–38. [Google Scholar] [CrossRef] [PubMed]
- Chow, D.C.; Forte, J.G. Functional significance of the beta-subunit for heterodimeric p-type atpases. J. Exp. Biol. 1995, 198, 1–17. [Google Scholar]
- Geering, K. The functional role of beta subunits in oligomeric p-type atpases. J. Bioenerg. Biomembr. 2001, 33, 425–438. [Google Scholar] [CrossRef] [PubMed]
- Dempski, R.E.; Friedrich, T.; Bamberg, E. The beta subunit of the Na+/K+-atpase follows the conformational state of the holoenzyme. J. Gen. Physiol. 2005, 125, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Crambert, G.; Hasler, U.; Beggah, A.T.; Yu, C.; Modyanov, N.N.; Horisberger, J.D.; Lelievre, L.; Geering, K. Transport and pharmacological properties of nine different human Na,K-atpase isozymes. J. Biol. Chem. 2000, 275, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, S.A.; Palmer, L.G.; Quan, K.; Harper, J.F.; Ball, W.J., Jr.; Bander, N.H.; Peralta Soler, A.; Rajasekaran, A.K. Na,K-atpase beta-subunit is required for epithelial polarization, suppression of invasion, and cell motility. Mol. Biol. Cell 2001, 12, 279–295. [Google Scholar] [CrossRef]
- Espineda, C.E.; Chang, J.H.; Twiss, J.; Rajasekaran, S.A.; Rajasekaran, A.K. Repression of Na,K-atpase beta1-subunit by the transcription factor snail in carcinoma. Mol. Biol. Cell 2004, 15, 1364–1373. [Google Scholar] [CrossRef]
- Madan, P.; Rose, K.; Watson, A.J. Na/K-atpase beta1 subunit expression is required for blastocyst formation and normal assembly of trophectoderm tight junction-associated proteins. J. Biol. Chem. 2007, 282, 12127–12134. [Google Scholar] [CrossRef]
- Padilla-Benavides, T.; Roldan, M.L.; Larre, I.; Flores-Benitez, D.; Villegas-Sepulveda, N.; Contreras, R.G.; Cereijido, M.; Shoshani, L. The polarized distribution of Na+,K+-atpase: Role of the interaction between {beta} subunits. Mol. Biol. Cell 2010, 21, 2217–2225. [Google Scholar] [CrossRef]
- Tokhtaeva, E.; Sachs, G.; Souda, P.; Bassilian, S.; Whitelegge, J.P.; Shoshani, L.; Vagin, O. Epithelial junctions depend on intercellular trans-interactions between the Na,K-atpase beta(1) subunits. J. Biol. Chem. 2011, 286, 25801–25812. [Google Scholar] [CrossRef]
- Shoshani, L.; Contreras, R.G.; Roldan, M.L.; Moreno, J.; Lazaro, A.; Balda, M.S.; Matter, K.; Cereijido, M. The polarized expression of Na+,K+-atpase in epithelia depends on the association between beta-subunits located in neighboring cells. Mol. Biol. Cell 2005, 16, 1071–1081. [Google Scholar] [CrossRef]
- Gloor, S.; Antonicek, H.; Sweadner, K.J.; Pagliusi, S.; Frank, R.; Moos, M.; Schachner, M. The adhesion molecule on glia (amog) is a homologue of the beta subunit of the Na,K-atpase. J. Cell Biol. 1990, 110, 165–174. [Google Scholar] [CrossRef]
- Askari, A.; Kakar, S.S.; Huang, W.H. Ligand binding sites of the ouabain-complexed (Na+ + K+)-atpase. J. Biol. Chem. 1988, 263, 235–242. [Google Scholar] [PubMed]
- Hamlyn, J.M.; Blaustein, M.P.; Bova, S.; DuCharme, D.W.; Harris, D.W.; Mandel, F.; Mathews, W.R.; Ludens, J.H. Identification and characterization of a ouabain-like compound from human plasma. Proc. Natl. Acad. Sci. USA 1991, 88, 6259–6263. [Google Scholar] [CrossRef] [PubMed]
- Laursen, M.; Gregersen, J.L.; Yatime, L.; Nissen, P.; Fedosova, N.U. Structures and characterization of digoxin- and bufalin-bound Na+,K+-atpase compared with the ouabain-bound complex. Proc. Natl. Acad. Sci. USA 2015, 112, 1755–1760. [Google Scholar] [CrossRef]
- Aizman, O.; Uhlen, P.; Lal, M.; Brismar, H.; Aperia, A. Ouabain, a steroid hormone that signals with slow calcium oscillations. Proc. Natl. Acad. Sci. USA 2001, 98, 13420–13424. [Google Scholar] [CrossRef] [Green Version]
- Haas, M.; Askari, A.; Xie, Z. Involvement of src and epidermal growth factor receptor in the signal-transducing function of Na+,K+-atpase. J. Biol. Chem. 2000, 275, 27832–27837. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zelenin, S.; Aperia, A.; Aizman, O. Low doses of ouabain protect from serum deprivation-triggered apoptosis and stimulate kidney cell proliferation via activation of nf-kappab. J. Am. Soc. Nephrol. 2006, 17, 1848–1857. [Google Scholar] [CrossRef]
- Liu, J.; Liang, M.; Liu, L.; Malhotra, D.; Xie, Z.; Shapiro, J.I. Ouabain-induced endocytosis of the plasmalemmal Na/K-atpase in llc-pk1 cells requires caveolin-1. Kidney Int. 2005, 67, 1844–1854. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, X.; Pierre, S.V.; Askari, A. Association of pi3k-akt signaling pathway with digitalis-induced hypertrophy of cardiac myocytes. Am. J. Physiol. Cell Physiol. 2007, 293, C1489–C1497. [Google Scholar] [CrossRef] [PubMed]
- Larre, I.; Cereijido, M. Na,K-atpase is the putative membrane receptor of hormone ouabain. Commun. Integr. Biol. 2010, 3, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Li, X.; Liang, M.; Liu, L.; Xie, J.X.; Ye, Q.; Kometiani, P.; Tillekeratne, M.; Jin, R.; Xie, Z. Changes in sodium pump expression dictate the effects of ouabain on cell growth. J. Biol. Chem. 2009, 284, 14921–14929. [Google Scholar] [CrossRef]
- Larre, I.; Lazaro, A.; Contreras, R.G.; Balda, M.S.; Matter, K.; Flores-Maldonado, C.; Ponce, A.; Flores-Benitez, D.; Rincon-Heredia, R.; Padilla-Benavides, T.; et al. Ouabain modulates epithelial cell tight junction. Proc. Natl. Acad. Sci. USA 2010, 107, 11387–11392. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.; Tian, J.; Liu, L.; Pierre, S.; Liu, J.; Shapiro, J.; Xie, Z.J. Identification of a pool of non-pumping Na/K-atpase. J. Biol. Chem. 2007, 282, 10585–10593. [Google Scholar] [CrossRef]
- Tian, J.; Cai, T.; Yuan, Z.; Wang, H.; Liu, L.; Haas, M.; Maksimova, E.; Huang, X.Y.; Xie, Z.J. Binding of src to Na+,K+-atpase forms a functional signaling complex. Mol. Biol. Cell 2006, 17, 317–326. [Google Scholar] [CrossRef]
- Simonini, M.; Casanova, P.; Citterio, L.; Messaggio, E.; Lanzani, C.; Manunta, P. Endogenous ouabain and related genes in the translation from hypertension to renal diseases. Int. J. Mol. Sci. 2018, 19, 1948. [Google Scholar] [CrossRef]
- Simonini, M.; Casanova, P.; Citterio, L.; Messaggio, E.; Lanzani, C.; Manunta, P. Reply: “Comment on: Endogenous ouabain and related genes in the translation from hypertension to renal diseases”. Int. J. Mol. Sci. 2019, 20, 542. [Google Scholar] [CrossRef]
- Furstenwerth, H. Comment on: Endogenous ouabain and related genes in the translation from hypertension to renal diseases, Int. J. Mol. Sci. 2018, 19, 1948. Int. J. Mol. Sci. 2019, 20, 505. [Google Scholar] [CrossRef]
- Mathews, W.R.; DuCharme, D.W.; Hamlyn, J.M.; Harris, D.W.; Mandel, F.; Clark, M.A.; Ludens, J.H. Mass spectral characterization of an endogenous digitalislike factor from human plasma. Hypertension 1991, 17, 930–935. [Google Scholar] [CrossRef]
- Ludens, J.H.; Clark, M.A.; DuCharme, D.W.; Harris, D.W.; Lutzke, B.S.; Mandel, F.; Mathews, W.R.; Sutter, D.M.; Hamlyn, J.M. Purification of an endogenous digitalislike factor from human plasma for structural analysis. Hypertension 1991, 17, 923–929. [Google Scholar] [CrossRef]
- el-Masri, M.A.; Clark, B.J.; Qazzaz, H.M.; Valdes, R., Jr. Human adrenal cells in culture produce both ouabain-like and dihydroouabain-like factors. Clin. Chem. 2002, 48, 1720–1730. [Google Scholar]
- Laredo, J.; Hamilton, B.P.; Hamlyn, J.M. Secretion of endogenous ouabain from bovine adrenocortical cells: Role of the zona glomerulosa and zona fasciculata. Biochem. Biophys. Res. Commun. 1995, 212, 487–493. [Google Scholar] [CrossRef]
- Doris, P.A.; Hayward-Lester, A.; Bourne, D.; Stocco, D.M. Ouabain production by cultured adrenal cells. Endocrinology 1996, 137, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Qazzaz, H.M.; Cao, Z.; Bolanowski, D.D.; Clark, B.J.; Valdes, R., Jr. De novo biosynthesis and radiolabeling of mammalian digitalis-like factors. Clin. Chem. 2004, 50, 612–620. [Google Scholar] [CrossRef]
- Bauer, N.; Muller-Ehmsen, J.; Kramer, U.; Hambarchian, N.; Zobel, C.; Schwinger, R.H.; Neu, H.; Kirch, U.; Grunbaum, E.G.; Schoner, W. Ouabain-like compound changes rapidly on physical exercise in humans and dogs: Effects of beta-blockade and angiotensin-converting enzyme inhibition. Hypertension 2005, 45, 1024–1028. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, O.V.; Lakatta, E.G.; Bagrov, A.Y. Endogenous Na,K pump ligands are differentially regulated during acute nacl loading of dahl rats. Circulation 2000, 102, 3009–3014. [Google Scholar] [CrossRef]
- Fedorova, O.V.; Kolodkin, N.I.; Agalakova, N.I.; Namikas, A.R.; Bzhelyansky, A.; St-Louis, J.; Lakatta, E.G.; Bagrov, A.Y. Antibody to marinobufagenin lowers blood pressure in pregnant rats on a high nacl intake. J. Hypertens. 2005, 23, 835–842. [Google Scholar] [CrossRef]
- Fedorova, O.V.; Agalakova, N.I.; Talan, M.I.; Lakatta, E.G.; Bagrov, A.Y. Brain ouabain stimulates peripheral marinobufagenin via angiotensin ii signalling in nacl-loaded dahl-s rats. J. Hypertens. 2005, 23, 1515–1523. [Google Scholar] [CrossRef]
- Gottlieb, S.S.; Rogowski, A.C.; Weinberg, M.; Krichten, C.M.; Hamilton, B.P.; Hamlyn, J.M. Elevated concentrations of endogenous ouabain in patients with congestive heart failure. Circulation 1992, 86, 420–425. [Google Scholar] [CrossRef]
- Manunta, P.; Ferrandi, M.; Bianchi, G.; Hamlyn, J.M. Endogenous ouabain in cardiovascular function and disease. J. Hypertens. 2009, 27, 9–18. [Google Scholar] [CrossRef]
- Rincon-Heredia, R.; Flores-Benitez, D.; Flores-Maldonado, C.; Bonilla-Delgado, J.; Garcia-Hernandez, V.; Verdejo-Torres, O.; Castillo, A.M.; Larre, I.; Poot-Hernandez, A.C.; Franco, M.; et al. Ouabain induces endocytosis and degradation of tight junction proteins through erk1/2-dependent pathways. Exp. Cell Res. 2014, 320, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Ponce, A.; Larre, I.; Castillo, A.; Garcia-Villegas, R.; Romero, A.; Flores-Maldonado, C.; Martinez-Rendon, J.; Contreras, R.G.; Cereijido, M. Ouabain increases gap junctional communication in epithelial cells. Cell. Physiol. Biochem. 2014, 34, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Vagin, O.; Turdikulova, S.; Sachs, G. Recombinant addition of n-glycosylation sites to the basolateral Na,K-atpase beta1 subunit results in its clustering in caveolae and apical sorting in hgt-1 cells. J. Biol. Chem. 2005, 280, 43159–43167. [Google Scholar] [CrossRef] [PubMed]
- Juliano, R.L. Adhesion and detachment characteristics of chinese hamster cell membrane mutants. J. Cell Biol. 1978, 76, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Harper, P.A.; Juliano, R.L. Isolation and characterization of chinese hamster ovary cell variants defective in adhesion to fibronectin-coated collagen. J. Cell Biol. 1980, 87, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Tokhtaeva, E.; Sun, H.; Deiss-Yehiely, N.; Wen, Y.; Soni, P.N.; Gabrielli, N.M.; Marcus, E.A.; Ridge, K.M.; Sachs, G.; Vazquez-Levin, M.; et al. The o-glycosylated ectodomain of fxyd5 impairs adhesion by disrupting cell-cell trans-dimerization of Na,K-atpase beta1 subunits. J. Cell Sci. 2016, 129, 2394–2406. [Google Scholar] [CrossRef]
- Wang, H.; Haas, M.; Liang, M.; Cai, T.; Tian, J.; Li, S.; Xie, Z. Ouabain assembles signaling cascades through the caveolar Na+/K+-atpase. J. Biol. Chem. 2004, 279, 17250–17259. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.C.; Pessoa, M.T.; Neves, L.D.; Alves, S.L.; Silva, L.M.; Santos, H.L.; Oliveira, S.M.; Taranto, A.G.; Comar, M.; Gomes, I.V.; et al. 21-benzylidene digoxin: A proapoptotic cardenolide of cancer cells that up-regulates Na,K-atpase and epithelial tight junctions. PLoS ONE 2014, 9, e108776. [Google Scholar] [CrossRef] [PubMed]
- Contreras, R.G.; Shoshani, L.; Flores-Maldonado, C.; Lazaro, A.; Cereijido, M. Relationship between Na+,K+-atpase and cell attachment. J. Cell Sci. 1999, 112, 4223–4232. [Google Scholar]
- Xie, Z. Molecular mechanisms of Na/K-atpase-mediated signal transduction. Ann. N. Y. Acad. Sci. 2003, 986, 497–503. [Google Scholar] [CrossRef]
- Ponce, A.; Larre, I.; Castillo, A.; Flores-Maldonado, C.; Verdejo-Torres, O.; Contreras, R.G.; Cereijido, M. Ouabain modulates the distribution of connexin 43 in epithelial cells. Cell. Physiol. Biochem. 2016, 39, 1329–1338. [Google Scholar] [CrossRef]
- Zhang, B.; Luo, Q.; Sun, J.; Xu, B.; Ju, Y.; Yang, L.; Song, G. Mgf enhances tenocyte invasion through mmp-2 activity via the fak-erk1/2 pathway. Wound Repair Regen. 2015, 23, 394–402. [Google Scholar] [CrossRef]
- Li, W.; Liu, Z.; Zhao, C.; Zhai, L. Binding of mmp-9-degraded fibronectin to beta6 integrin promotes invasion via the fak-src-related erk1/2 and pi3k/akt/smad-1/5/8 pathways in breast cancer. Oncol. Rep. 2015, 34, 1345–1352. [Google Scholar] [CrossRef]
- Chang, H.; Dong, T.; Ma, X.; Zhang, T.; Chen, Z.; Yang, Z.; Zhang, Y. Spondin 1 promotes metastatic progression through fak and src dependent pathway in human osteosarcoma. Biochem. Biophys. Res. Commun. 2015, 464, 45–50. [Google Scholar] [CrossRef]
- Patel, A.; Sabbineni, H.; Clarke, A.; Somanath, P.R. Novel roles of src in cancer cell epithelial-to-mesenchymal transition, vascular permeability, microinvasion and metastasis. Life Sci. 2016, 157, 52–61. [Google Scholar] [CrossRef]
- Flores-Benitez, D.; Rincon-Heredia, R.; Razgado, L.F.; Larre, I.; Cereijido, M.; Contreras, R.G. Control of tight junctional sealing: Roles of epidermal growth factor and prostaglandin e2. Am. J. Physiol. Cell Physiol. 2009, 297, C611–C620. [Google Scholar] [CrossRef]
- Garcia-Hernandez, V.; Flores-Maldonado, C.; Rincon-Heredia, R.; Verdejo-Torres, O.; Bonilla-Delgado, J.; Meneses-Morales, I.; Gariglio, P.; Contreras, R.G. Egf regulates claudin-2 and -4 expression through src and stat3 in mdck cells. J. Cell Physiol. 2015, 230, 105–115. [Google Scholar] [CrossRef]
- Haas, M.; Wang, H.; Tian, J.; Xie, Z. Src-mediated inter-receptor cross-talk between the Na+/K+-atpase and the epidermal growth factor receptor relays the signal from ouabain to mitogen-activated protein kinases. J. Biol. Chem. 2002, 277, 18694–18702. [Google Scholar] [CrossRef]
- Li, Z.; Cai, T.; Tian, J.; Xie, J.X.; Zhao, X.; Liu, L.; Shapiro, J.I.; Xie, Z. Naktide, a Na/K-atpase-derived peptide src inhibitor, antagonizes ouabain-activated signal transduction in cultured cells. J. Biol. Chem. 2009, 284, 21066–21076. [Google Scholar] [CrossRef]
- McNeill, H.; Ozawa, M.; Kemler, R.; Nelson, W.J. Novel function of the cell adhesion molecule uvomorulin as an inducer of cell surface polarity. Cell 1990, 62, 309–316. [Google Scholar] [CrossRef]
- Cereijido, M.; Stefani, E.; Palomo, A.M. Occluding junctions in a cultured transporting epithelium: Structural and functional heterogeneity. J. Membr. Biol. 1980, 53, 19–32. [Google Scholar] [CrossRef]
- Antolović, R. Low nanomolar concentrations of ouabain may induce higher activity of the Na+/K+-atpase in human erythrocytes. Veterinarski Arhiv 2006, 76, 489–495. [Google Scholar]
- Oselkin, M.; Tian, D.; Bergold, P.J. Low-dose cardiotonic steroids increase sodium-potassium atpase activity that protects hippocampal slice cultures from experimental ischemia. Neurosci. Lett. 2010, 473, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Klimanova, E.A.; Petrushanko, I.Y.; Mitkevich, V.A.; Anashkina, A.A.; Orlov, S.N.; Makarov, A.A.; Lopina, O.D. Binding of ouabain and marinobufagenin leads to different structural changes in Na,K-atpase and depends on the enzyme conformation. FEBS Lett. 2015, 589, 2668–2674. [Google Scholar] [CrossRef]
- Dempski, R.E.; Hartung, K.; Friedrich, T.; Bamberg, E. Fluorometric measurements of intermolecular distances between the alpha- and beta-subunits of the Na+/K+-atpase. J. Biol. Chem. 2006, 281, 36338–36346. [Google Scholar] [CrossRef] [PubMed]
- Aperia, A.; Akkuratov, E.E.; Fontana, J.M.; Brismar, H. Na+-K+-atpase, a new class of plasma membrane receptors. Am. J. Physiol. Cell Physiol. 2016, 310, C491–C495. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, F.K.; Dube, P.; Mohamed, A.; Tian, J.; Malhotra, D.; Haller, S.T.; Kennedy, D.J. Cardiotonic steroids and the sodium trade balance: New insights into trade-off mechanisms mediated by the Na+/K+-atpase. Int. J. Mol. Sci. 2018, 19, 2576. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, S.A.; Gopal, J.; Willis, D.; Espineda, C.; Twiss, J.L.; Rajasekaran, A.K. Na,K-atpase beta1-subunit increases the translation efficiency of the alpha1-subunit in msv-mdck cells. Mol. Biol. Cell 2004, 15, 3224–3232. [Google Scholar] [CrossRef] [PubMed]
- Caplan, M.J.; Anderson, H.C.; Palade, G.E.; Jamieson, J.D. Intracellular sorting and polarized cell surface delivery of (Na+,K+)atpase, an endogenous component of mdck cell basolateral plasma membranes. Cell 1986, 46, 623–631. [Google Scholar] [CrossRef]
- Zulian, A.; Linde, C.I.; Pulina, M.V.; Baryshnikov, S.G.; Papparella, I.; Hamlyn, J.M.; Golovina, V.A. Activation of c-src underlies the differential effects of ouabain and digoxin on Ca2+ signaling in arterial smooth muscle cells. Am. J. Physiol. Cell Physiol. 2013, 304, C324–C333. [Google Scholar] [CrossRef]
- Wu, J.; Akkuratov, E.E.; Bai, Y.; Gaskill, C.M.; Askari, A.; Liu, L. Cell signaling associated with Na+/K+-atpase: Activation of phosphatidylinositide 3-kinase ia/akt by ouabain is independent of src. Biochemistry 2013, 52, 9059–9067. [Google Scholar] [CrossRef]
- Feng, W.; Webb, P.; Nguyen, P.; Liu, X.; Li, J.; Karin, M.; Kushner, P.J. Potentiation of estrogen receptor activation function 1 (af-1) by src/jnk through a serine 118-independent pathway. Mol. Endocrinol. 2001, 15, 32–45. [Google Scholar] [CrossRef]
- Madan, N.; Xu, Y.; Duan, Q.; Banerjee, M.; Larre, I.; Pierre, S.V.; Xie, Z. Src-independent erk signaling through the rat alpha3 isoform of Na/K-atpase. Am. J. Physiol. Cell Physiol. 2017, 312, C222–C232. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Troyanovsky, R.B.; Troyanovsky, S.M. Spontaneous assembly and active disassembly balance adherens junction homeostasis. Proc. Natl. Acad. Sci. USA 2010, 107, 3528–3533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamrick, M.; Renaud, K.J.; Fambrough, D.M. Assembly of the extracellular domain of the Na,K-atpase beta subunit with the alpha subunit. Analysis of beta subunit chimeras and carboxyl-terminal deletions. J. Biol. Chem. 1993, 268, 24367–24373. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Ieggli, C.V.; Bohrer, D.; do Nascimento, P.C.; de Carvalho, L.M.; Garcia, S.C. Determination of sodium, potassium, calcium, magnesium, zinc, and iron in emulsified egg samples by flame atomic absorption spectrometry. Talanta 2010, 80, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Paskavitz, A.L.; Quintana, J.; Cangussu, D.; Tavera-Montanez, C.; Xiao, Y.; Ortiz-Miranda, S.; Navea, J.G.; Padilla-Benavides, T. Differential expression of zinc transporters accompanies the differentiation of c2c12 myoblasts. J. Trace Elem. Med. Biol. 2018, 49, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.J.; Xiao, Y.; Paskavitz, A.L.; Navarro-Tito, N.; Navea, J.G.; Padilla-Benavides, T. Atomic absorbance spectroscopy to measure intracellular zinc pools in mammalian cells. J. Vis. Exp. 2019, e59519, in press. [Google Scholar]
- Raimunda, D.; Padilla-Benavides, T.; Vogt, S.; Boutigny, S.; Tomkinson, K.N.; Finney, L.A.; Argüello, J.M. Periplasmic response upon disruption of transmembrane Cu transport in Pseudomonas aeruginosa. Metallomics 2013, 5, 144–151. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, F.; Shou, H.; Huang, F.; Zheng, L.; He, F.; Li, J.; Zhao, F.J.; Ueno, D.; Ma, J.F.; et al. Mutation in nicotianamine aminotransferase stimulated the Fe(ii) acquisition system and led to iron accumulation in rice. Plant Physiol. 2007, 145, 1647–1657. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilchis-Nestor, C.A.; Roldán, M.L.; Leonardi, A.; Navea, J.G.; Padilla-Benavides, T.; Shoshani, L. Ouabain Enhances Cell-Cell Adhesion Mediated by β1 Subunits of the Na+,K+-ATPase in CHO Fibroblasts. Int. J. Mol. Sci. 2019, 20, 2111. https://doi.org/10.3390/ijms20092111
Vilchis-Nestor CA, Roldán ML, Leonardi A, Navea JG, Padilla-Benavides T, Shoshani L. Ouabain Enhances Cell-Cell Adhesion Mediated by β1 Subunits of the Na+,K+-ATPase in CHO Fibroblasts. International Journal of Molecular Sciences. 2019; 20(9):2111. https://doi.org/10.3390/ijms20092111
Chicago/Turabian StyleVilchis-Nestor, Claudia Andrea, María Luisa Roldán, Angelina Leonardi, Juan G. Navea, Teresita Padilla-Benavides, and Liora Shoshani. 2019. "Ouabain Enhances Cell-Cell Adhesion Mediated by β1 Subunits of the Na+,K+-ATPase in CHO Fibroblasts" International Journal of Molecular Sciences 20, no. 9: 2111. https://doi.org/10.3390/ijms20092111