Leishmania infantum β-Tubulin Identified by Reverse Engineering Technology through Phage Display Applied as Theranostic Marker for Human Visceral Leishmaniasis

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
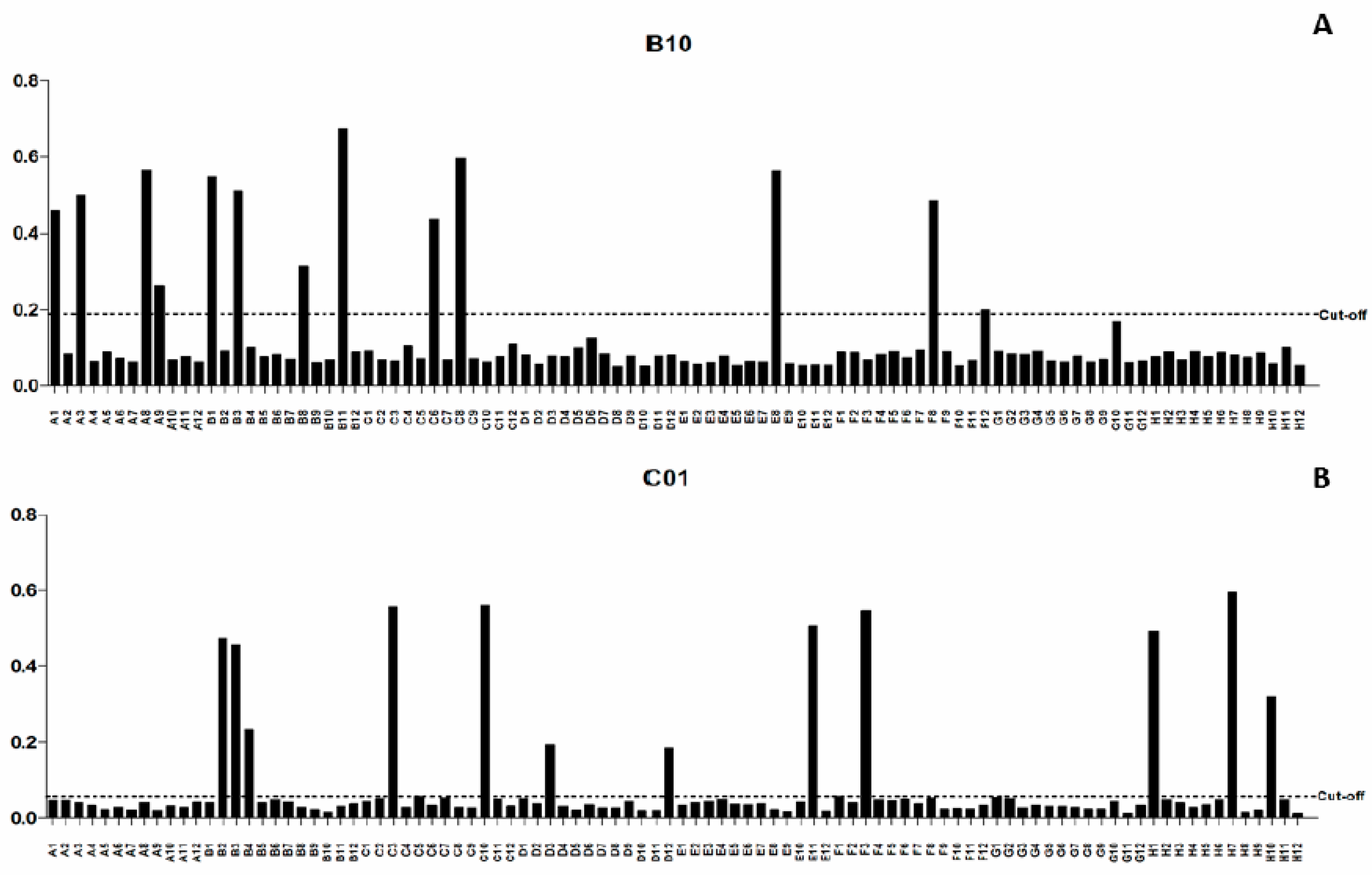
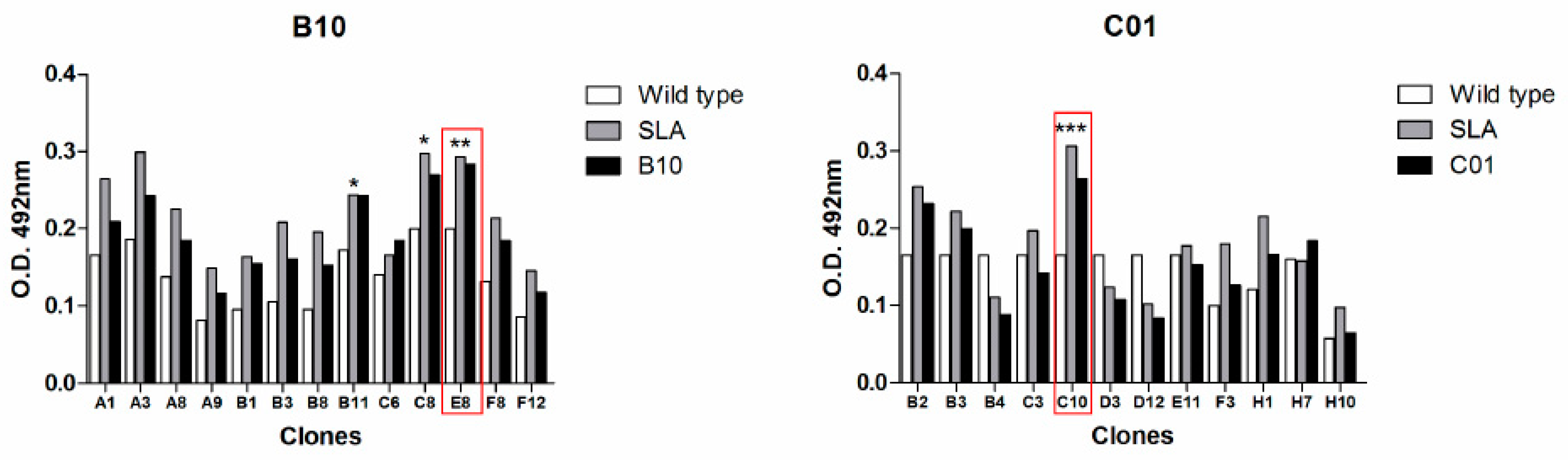
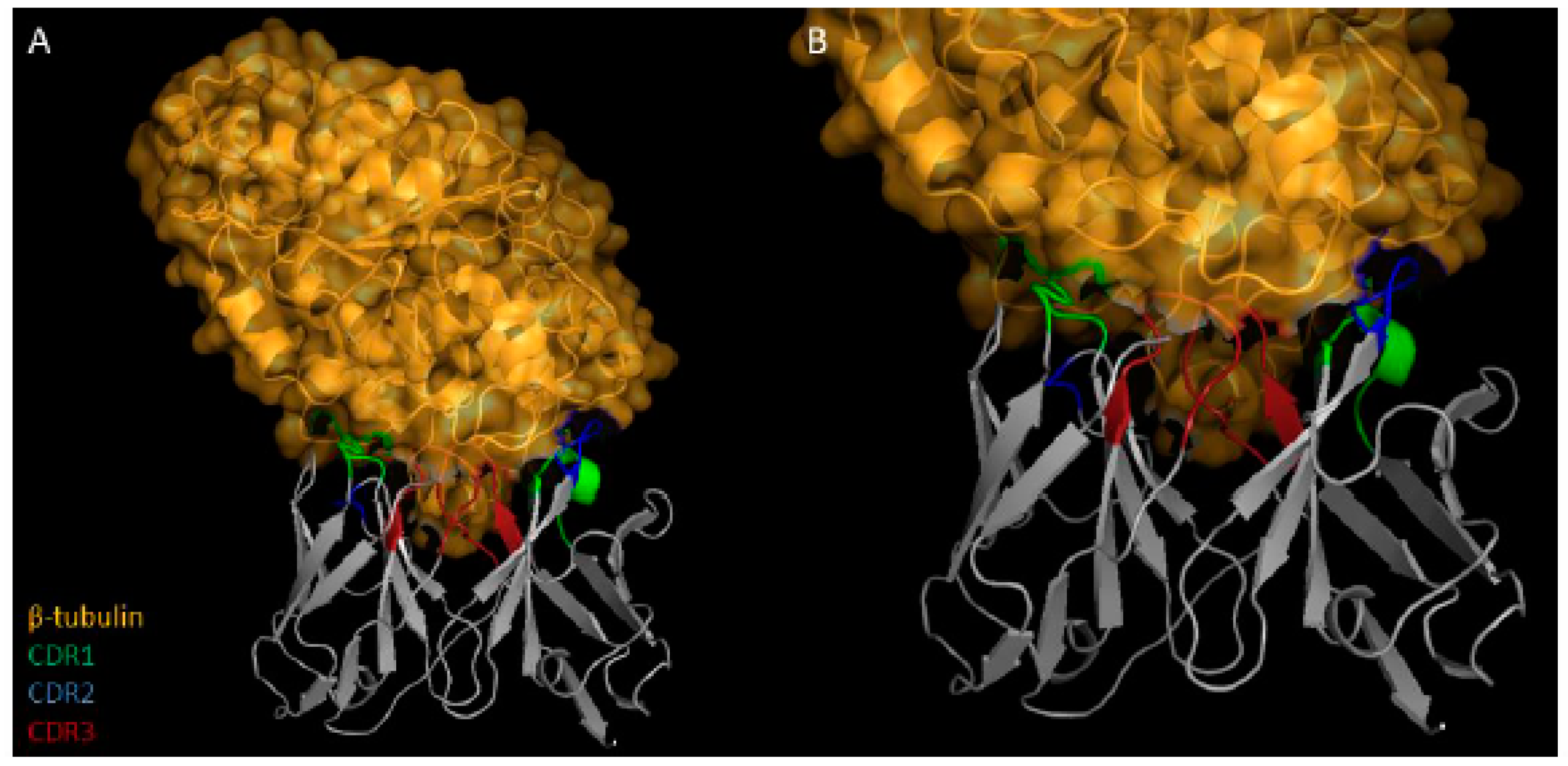
2.1. Characterization of scFv Antibody Clones
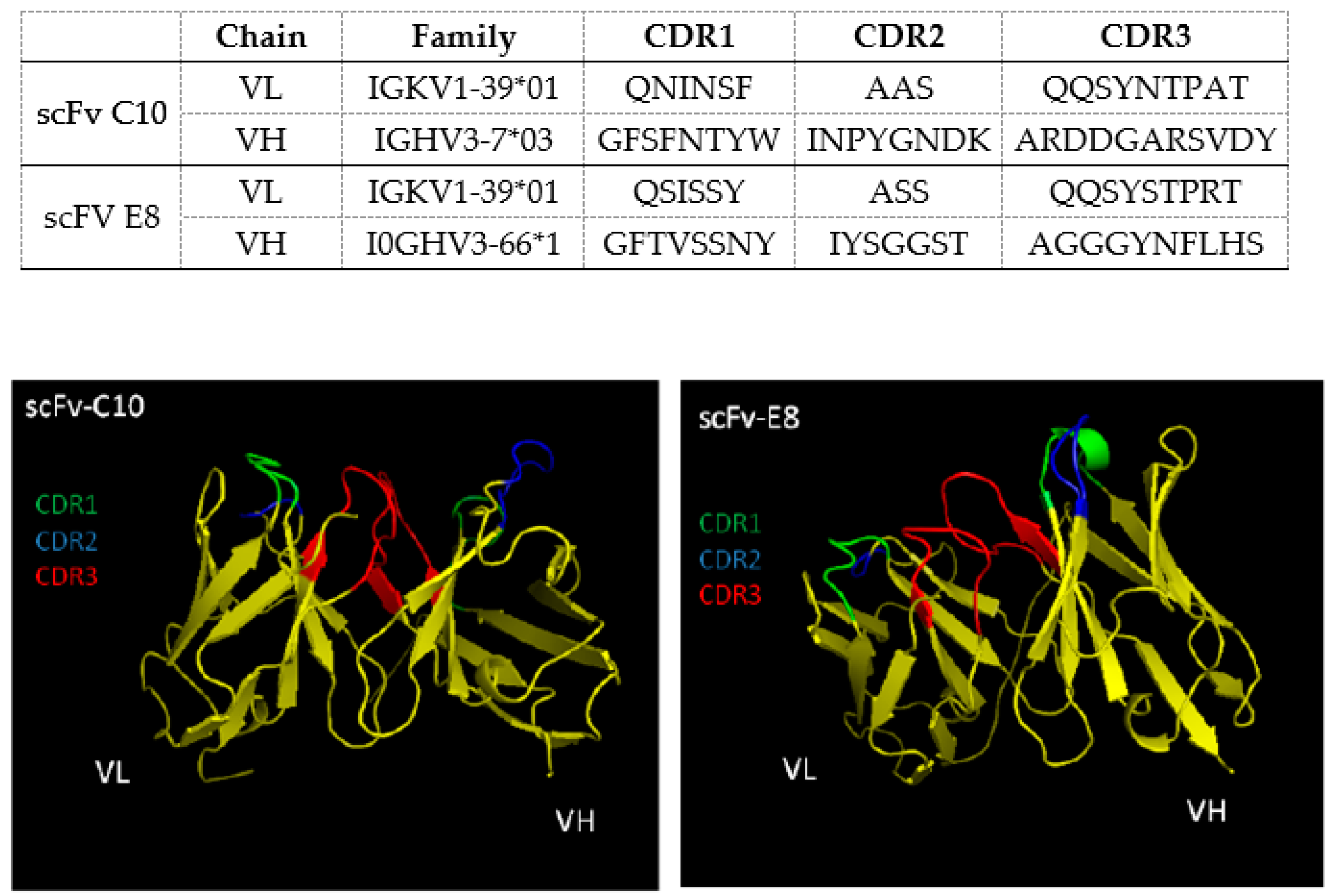
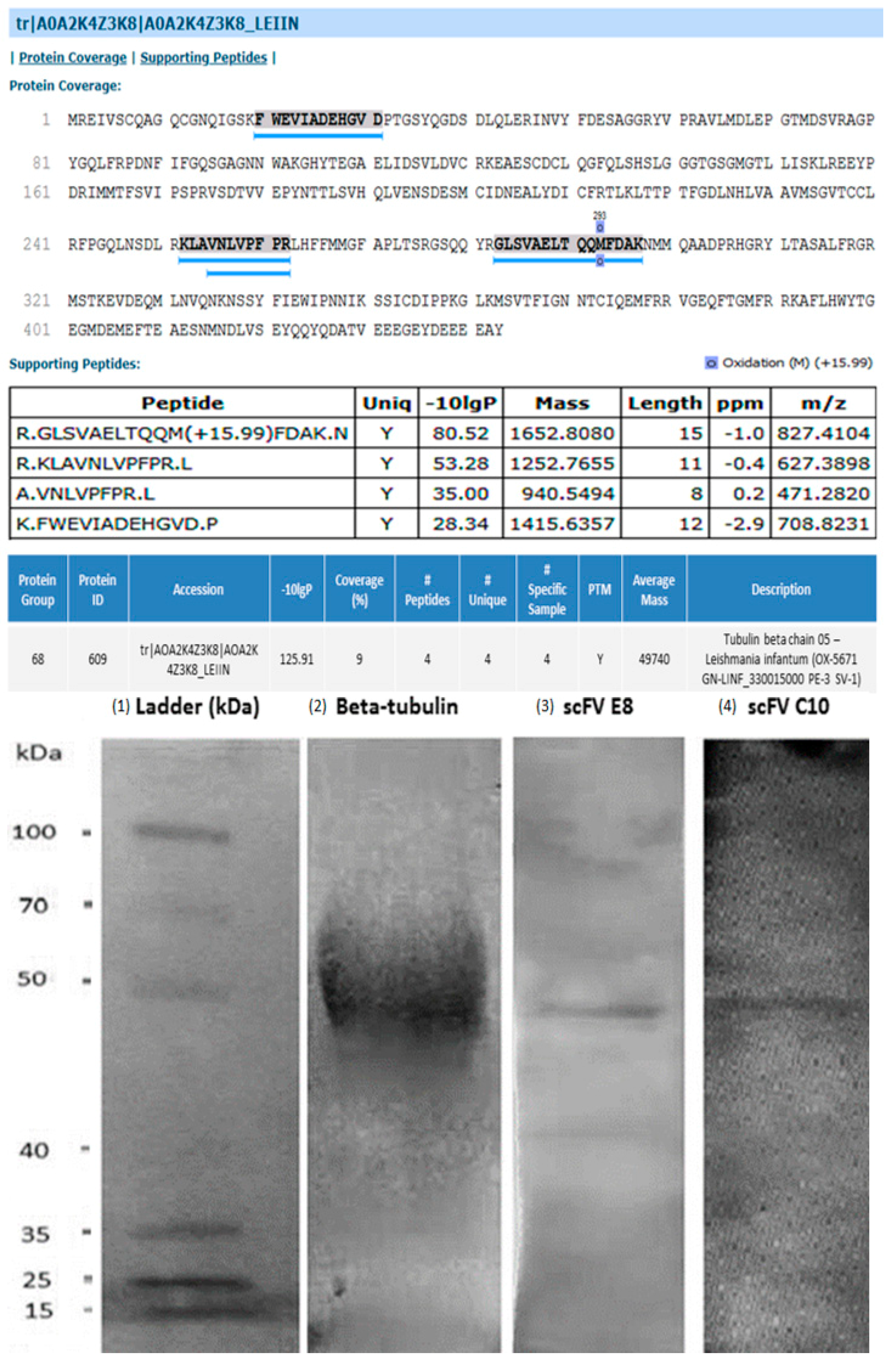
2.2. Identification of Target Protein in Leishmania Parasites
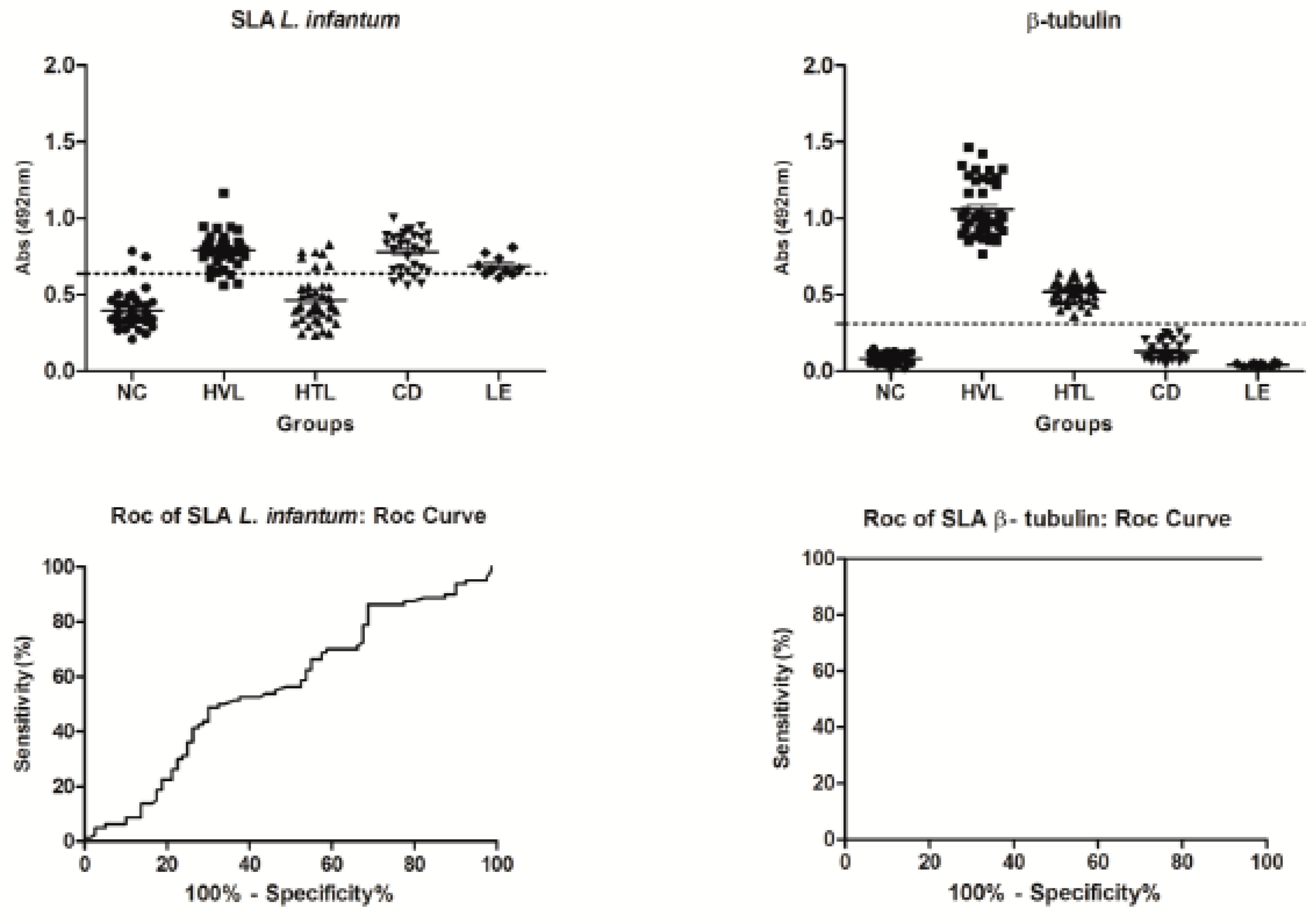
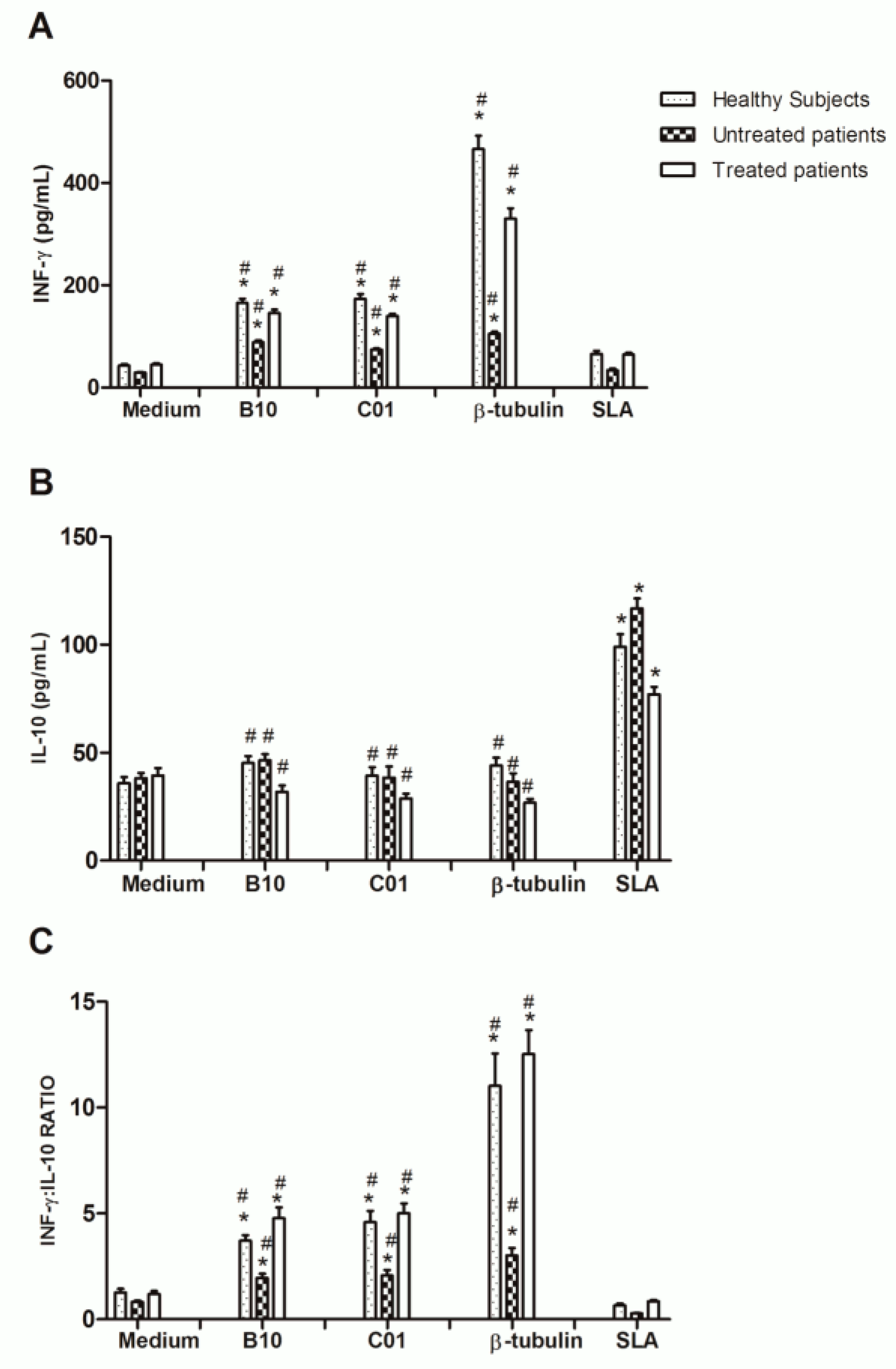
2.3. Diagnostic Potential and Immunogenicity of Recombinant β-Tubulin Protein
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Parasites
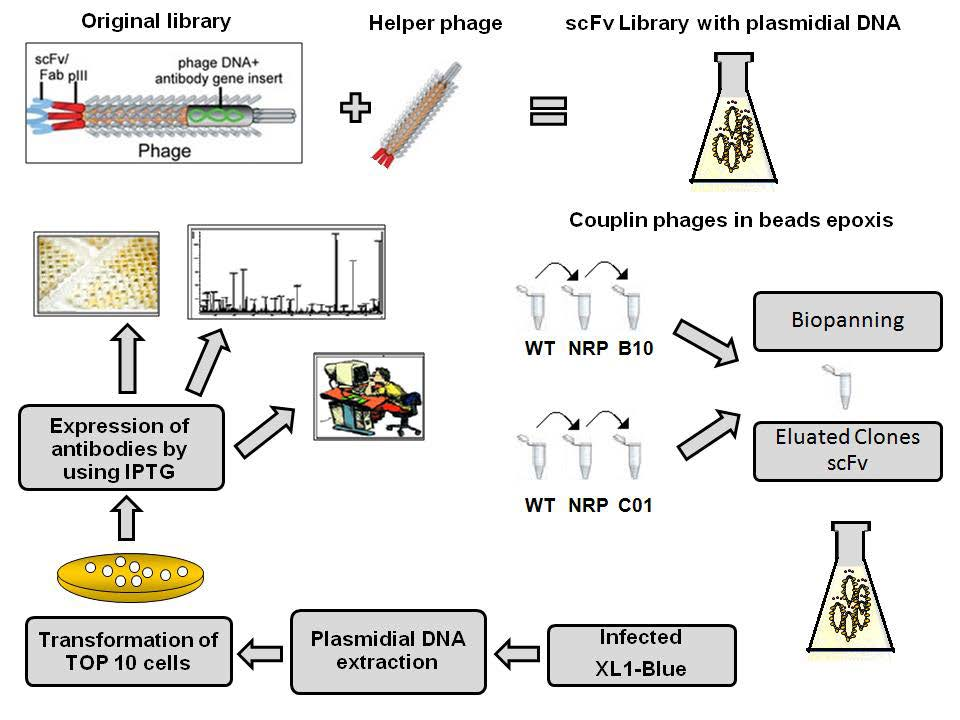
4.3. Phage Display for Selection of B10 and C01 Mimotopes-Specific scFv Antibodies
4.4. ELISA to Evaluate the scFv Expression
4.5. Analysis of Selected Clones’ Specificity
4.6. Purification of Selected scFv Clones and Identification of the Native Antigen
4.7. DNA Sequencing and Bioinformatics Assays
4.8. Cloning, Expression and Purification of Recombinant β-Tubulin Protein
4.9. Immunoblotting Assays
4.10. Sera and ELISA Assays
4.11. In Vitro PBMCs Culture and Cytokine Production
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schroeder, J.; Aebischer, T. Vaccines for leishmaniasis: From proteome to vaccine candidates. Hum. Vaccines 2011, 7, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Savoia, D. Recent updates and perspectives on leishmaniasis. J. Inf. Dev. Countr. 2015, 9, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; De-Boer, M. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, 35671. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.B.; Vilela, J.A.; Pires, M.S.; Santos, H.A.; Falqueto, A.; Peixoto, M.P.; Oliveira, T.A.; Santos, F.N.; Silva, V.L.; Massard, C.L.; et al. Seroepidemiological aspects of Leishmania spp. in dogs in the Itaguai micro-region, Rio de Janeiro. Brazil. Rev. Bras. Parasitol. Vet. 2013, 22, 39–45. [Google Scholar] [CrossRef]
- Rezvan, H.; Moafi, M. An overview on Leishmania vaccines: A narrative review article. Vet. Res. Forum. 2015, 6, 1–7. [Google Scholar] [PubMed]
- Costa, L.E.; Goulart, L.R.; Pereira, N.C.; Lima, M.I.; Duarte, M.C.; Martins, V.T.; Lage, P.S.; Menezes-Souza, D.; Ribeiro, T.G.; Coelho, E.A.; et al. Mimotope-based vaccines of Leishmania infantum antigens and their protective efficacy against visceral leishmaniasis. PLoS ONE 2014, 9, e110014. [Google Scholar] [CrossRef]
- Costa, L.E.; Chavez-Fumagalli, M.A.; Martins, V.T.; Duarte, M.C.; Lage, D.P.; Lima, M.I.; Pereira, N.C.; Soto, M.; Tavares, C.A.; Coelho, E.A.; et al. Phage-fused epitopes from Leishmania infantum used as immunogenic vaccines confer partial protection against Leishmania amazonensis infection. Parasitology. 2015, 142, 1335–1347. [Google Scholar] [CrossRef]
- Costa, L.E.; Salles, B.C.S.; Santos, T.T.O.; Ramos, F.F.; Lima, M.P.; Lima, M.I.S.; Portela, Á.S.B.; Chávez-Fumagalli, M.A.; Duarte, M.C.; Coelho, E.A.F.; et al. Antigenicity of phage clones and their synthetic peptides for the serodiagnosis of canine and human visceral leishmaniasis. Microb. Pathog. 2017, 110, 14–22. [Google Scholar] [CrossRef]
- Chapman, L.A.C.; Morgan, A.L.K.; Adams, E.R.; Bern, C.; Medley, G.F.; Hollingsworth, T.D. Age trends in asymptomatic and symptomatic Leishmania donovani infection in the Indian subcontinent: A review and analysis of data from diagnostic and epidemiological studies. PLoS Negl. Trop. Dis. 2018, 12, e0006803. [Google Scholar] [CrossRef]
- Sarkari, B.; Rezaei, Z.; Mohebal, M. Immunodiagnosis of Visceral Leishmaniasis: Current Status and Challenges: A Review Article. Iran J. Parasitol. 2018, 13, 331–341. [Google Scholar]
- Georgiadou, S.P.; Makaritsis, K.P.; Dalekos, G.N. Leishmaniasis revisited: Current aspects on epidemiology, diagnosis and treatment. J. Transl. Int. Med. 2015, 3, 43–50. [Google Scholar] [CrossRef]
- Lima, M.P.; Costa, L.E.; Duarte, M.C.; Menezes-Souza, D.; Salles, B.C.S.; De-Oliveira-Santos, T.T.; Ramos, F.F.; Chávez-Fumagalli, M.A.; Kursancew, A.C.S.; Ambrósio, R.P.; Coelho, E.A.F.; et al. Evaluation of a hypothetical protein for serodiagnosis and as a potential marker for post-treatment serological evaluation of tegumentary leishmaniasis patients. Parasitol. Res. 2017, 116, 1197–1206. [Google Scholar] [CrossRef]
- Mohapatra, T.M.; Singh, D.P.; Sen, M.R.; Bharti, K.; Sundar, S. Compararative evaluation of rK9, rK26 and rK39 antigens in the serodiagnosis of Indian visceral leishmaniasis. J. Infect. Dev. Ctries 2010, 4, 114–117. [Google Scholar] [CrossRef]
- Gadisa, E.; Custodio, E.; Cañavate, C.; Sordo, L.; Abebe, Z.; Nieto, J.; Chicharro, C.; Aseffa, A.; Yamuah, L.; Cruz, I. Usefulness of the rK39-immunochromatographic test, direct agglutination test, and leishmanin skin test for detecting asymptomatic Leishmania infection in children in a new visceral leishmaniasis focus in Amhara State, Ethiopia, Am. J. Trop. Med. Hyg. 2012, 86, 792–798. [Google Scholar] [CrossRef]
- Salles, B.C.S.; Dias, D.S.; Steiner, B.T.; Lage, D.P.; Ramos, F.F.; Ribeiro, P.A.F.; Santos, T.T.O.; Lima, M.P.; Costa, L.E.; Coelho, E.A.F.; et al. Potential application of small myristoylated protein-3 evaluated as recombinant antigen and a synthetic peptide containing its linear B-cell epitope for the serodiagnosis of canine visceral and human tegumentary leishmaniasis. Immunobiology 2018, 1, 163–171. [Google Scholar] [CrossRef]
- Costa, M.M.; Penido, M.; Dos-Santos, M.S.; Doro, D.; De-Freitas, E.; Michalick, M.S.; Grimaldi, G.; Gazzinelli, R.T.; Fernandes, A.P. Improved canine and human visceral leishmaniasis immunodiagnosis using combinations of synthetic peptides in enzyme-linked immunosorbent assay. PLoS Negl. Trop. Dis. 2012, 6, e1622. [Google Scholar] [CrossRef]
- Menezes-Souza, D.; Mendes, T.A.; Gomes, M.S.; Bartholomeu, D.C.; Fujiwara, R.T. Improving serodiagnosis of human and canine leishmaniasis with recombinant Leishmania braziliensis cathepsin l-like protein and a synthetic peptide containing its linear B-cell epitope. PLoS Negl. Trop. Dis. 2015, 9, e3426. [Google Scholar] [CrossRef]
- Magalhães, F.B.; Castro Neto, A.L.; Nascimento, M.B.; Santos, W.J.T.; Medeiros, Z.M.; Lima Neto, A.S.; Costa, D.L.; Costa, C.H.N.; Dos-Santos, W.L.C.; De-Melo Neto, O.P. Evaluation of a new set of recombinant antigens for the serological diagnosis of human and canine visceral leishmaniasis. PLoS ONE 2017, 12, e0184867. [Google Scholar] [CrossRef]
- Carvalho, A.M.R.S.; Mendes, T.A.O.; Coelho, E.A.F.; Duarte, M.C.; Menezes-Souza, D. New antigens for the serological diagnosis of human visceral leishmaniasis identified by immunogenomic screening. PLoS ONE 2018, 13, e0209599. [Google Scholar] [CrossRef]
- Salles, B.C.; Costa, L.E.; Alves, P.T.; Dias, A.C.; Vaz, E.R.; Menezes-Souza, D.; Ramos, F.F.; Duarte, M.C.; Roatt, B.M.; Coelho, E.A.; et al. Leishmania infantum mimotopes and a phage-ELISA assay as tools for a sensitive and specific serodiagnosis of human visceral leishmaniasis. Diagn. Microbiol. Infect. Dis. 2017, 87, 219–225. [Google Scholar] [CrossRef]
- Townsend, C.L.; Laffy, J.M.; Wu, Y.B.; Silva O’Hare, J.; Martin, V.; Kipling, D.; Fraternali, F.; Dunn-Walters, D.K. Significant Differences in Physicochemical Properties of Human Immunoglobulin Kappa and Lambda CDR3 Regions. Front. Immunol. 2016, 7, 388. [Google Scholar] [CrossRef]
- McKean, P.G.; Vaughan, S.; Gull, K. The extended tubulin superfamily. J. Cell Sci. 2001, 114, 2723–2733. [Google Scholar]
- Barrack, K.L.; Fyfe, P.K.; Finney, A.J.; Hunter, W.N. Crystal structure of the C-terminal domain of tubulin-binding cofactor C from Leishmania major. Mol. Biochem. Parasitol. 2015, 201, 26–30. [Google Scholar] [CrossRef]
- Escudero-Martínez, J.M.; Pérez-Pertejo, Y.; Reguera, R.M.; Castro, M.Á.; Rojo, M.V.; Santiago, C.; Abad, A.; García, P.A.; López-Pérez, J.L.; San Feliciano, A.; Balaña-Fouce, R. Antileishmanial activity and tubulin polymerization inhibition of podophyllotoxin derivatives on Leishmania infantum. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 272–285. [Google Scholar] [CrossRef]
- Probst, P.; Stromberg, E.; Ghalib, H.W.; Mozel, M.; Badaro, R.; Reed, S.G.; Webb, J.R. Identification and characterization of T cell-stimulating antigens from Leishmania by CD4 T cell expression cloning. J. Immunol. 2001, 166, 498–505. [Google Scholar] [CrossRef]
- Lima, M.P.; Costa, L.E.; Lage, D.P.; Dias, D.S.; Ribeiro, P.A.F.; Machado, A.S.; Ramos, F.F.; Salles, B.C.S.; Fagundes, M.I.; Coelho, E.A.F.; et al. Diagnostic application of recombinant Leishmania proteins and evaluation of their in vitro immunogenicity after stimulation of immune cells collected from tegumentary leishmaniasis patients and healthy individuals. Cell Immunol. 2018, 334, 61–69. [Google Scholar] [CrossRef]
- Vanloubbeeck, Y.; Jones, D.E. The immunology of Leishmania infection and the implications for vaccine development. Ann. N. Y. Acad. Sci. 2004, 1026, 267–272. [Google Scholar] [CrossRef]
- Rodrigues, V.; Cordeiro-da-Silva, A.; Laforge, M.; Silvestre, R.; Estaquier, J. Regulation of immunity during visceral Leishmania infection. Parasites Vectors 2016, 9, 118. [Google Scholar] [CrossRef]
- Srivastava, S.; Shankar, P.; Mishra, J.; Singh, S. Possibilities and challenges for developing a successful vaccine for leishmaniasis. Parasites Vectors 2016, 9, 277. [Google Scholar] [CrossRef] [Green Version]
- Jamal, F.; Shivam, P.; Kumari, S.; Singh, M.K.; Sardar, A.H.; Pushpanjali, M.S.; Narayan, S.; Gupta, A.K.; Pandey, K.; Das, V.N.R.; et al. Identification of Leishmania donovani antigen in circulating immune complexes of visceral leishmaniasis subjects for diagnosis. PLoS ONE 2017, 12, e0182474. [Google Scholar] [CrossRef]
- Ruiter, C.M.; van der Veer, C.; Leeflang, M.M.; Deborggraeve, S.; Lucas, C.; Adams, E.R. Molecular tools for diagnosis of visceral leishmaniasis: Systematic review and meta-analysis of diagnostic test accuracy. J. Clin. Microbiol. 2014, 52, 3147–3155. [Google Scholar] [CrossRef]
- Anfossi, L.; Di Nardo, F.; Profiti, M.; Nogarol, C.; Cavalera, S.; Baggiani, C.; Giovannoli, C.; Spano, G.; Ferroglio, E.; Rosati, S.; et al. A versatile and sensitive lateral flow immunoassay for the rapid diagnosis of visceral leishmaniasis. Anal. Bioanal. Chem. 2018, 410, 4123–4134. [Google Scholar] [CrossRef]
- Duarte, M.C.; Pimenta, D.C.; Menezes-Souza, D.; Magalhães, R.D.; Diniz, J.L.; Costa, L.E.; Chávez-Fumagalli, M.A.; Lage, P.S.; Bartholomeu, D.C.; Alves, M.J.; Coelho, E.A.; et al. Proteins selected in Leishmania (Viannia) braziliensis by an immunoproteomic approach with potential serodiagnosis applications for tegumentary leishmaniasis. Clin. Vaccine Immunol. 2015, 22, 1187–1196. [Google Scholar] [CrossRef]
- Kiguchi, Y.; Oyama, H.; Morita, I.; Katayama, E.; Fujita, M.; Narasaki, M.; Yokoyama, A.; Kobayashi, N. Antibodies and engineered antibody fragments against M13 filamentous phage to facilitate phage-display-based molecular breeding. Biol. Pharm. Bull. 2018, 41, 1062–1070. [Google Scholar] [CrossRef]
- Rickert, K.W.; Grinberg, L.; Woods, R.M.; Wilson, S.; Bowen, M.A.; Baca, M. Combining phage display with de novo protein sequencing for reverse engineering of monoclonal antibodies. MAbs 2016, 8, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Coelho, E.A.F.; Tavares, C.A.P.; Carvalho, F.A.A.; Chaves, K.F.; Teixeira, K.N.; Rodrigues, R.C.; Charest, H.; Matlashewski, G.; Gazzinelli, R.T.; Fernandes, A.P. Immune responses induced by the Leishmania (Leishmania) donovani A2 antigen, but not by the LACK antigen, are protective against experimental Leishmania (Leishmania) amazonensis infection. Infect. Immun. 2003, 71, 3988–3994. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Carneiro, A.P.; Reis, C.F.; Morari, E.C.; Maia, Y.C.; Nascimento, R.; Bonatto, J.M.; Souza, M.A.; Goulart, L.R.; Ward, L.S. A putative OTU domain-containing protein 1 deubiquitinating enzyme is differentially expressed in thyroid cancer and identifies less-aggressive tumours. Br. J. Cancer 2014, 111, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Marks, J.D.; Hoogenboom, H.R.; Bonnert, T.P.; McCafferty, J.; Griffiths, A.D.; Winter, G. By-passing immunization. Human antibodies from V-gene libraries displayed on phage. J. Mol. Biol. 1991, 5, 581–597. [Google Scholar] [CrossRef]
- Andris-Widhopf, J.; Steinberger, P.; Fuller, R.; Rader, C.; Barbas, C.F. Generation of human Fab antibody libraries: PCR amplification and assembly of light- and heavy-chain coding sequences. Cold Spring Harb. Protoc. 2011, 2011. [Google Scholar] [CrossRef]
- Andris-Widhopf, J.; Rader, C.; Steinberger, P.; Fuller, R.; Barbas, C.F. Methods for the generation of chicken monoclonal antibody fragments by phage display. J. Immunol. Methods 2000, 242, 159–181. [Google Scholar] [CrossRef]
- Rader, C.; Barbas, C.F. Phage display of combinatorial antibody libraries. Curr. Opin. Biotechnol. 1997, 8, 503–508. [Google Scholar] [CrossRef]
- Ribeiro, V.S.; Araujo, T.G.; Gonzaga, H.T.; Nascimento, R.; Goulart, L.R.; Costa-Cruz, J.M. Development of specific scFv antibodies to detect neurocysticercosis antigens and potential applications in immunodiagnosis. Immunol. Lett. 2013, 156, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Barbas, C.F.; Burton, D.R.; Scott, J.K.; Silvermam, G.J. Phage Display: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–675. [Google Scholar]
- Retter, I.; Althaus, H.H.; Munch, R.; Muller, W. VBASE2, an integrative V gene database. Nucleic Acids Res. 2005, 33, D671–D674. [Google Scholar] [CrossRef]
- Ramos, F.F.; Costa, L.E.; Dias, D.S.; Santos, T.T.O.; Rodrigues, M.R.; Lage, D.P.; Salles, B.C.S.; Martins, V.T.; Ribeiro, P.A.F.; Chávez-Fumagalli, M.A.; et al. Selection strategy of phage-displayed immunogens based on an in vitro evaluation of the Th1 response of PBMCs and their potential use as a vaccine against Leishmania infantum infection. Parasites Vectors 2017, 10, 617. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, L.E.; Alves, P.T.; Carneiro, A.P.; Dias, A.C.S.; Fujimura, P.T.; Araujo, G.R.; Tavares, G.S.V.; Ramos, F.F.; Duarte, M.C.; Menezes-Souza, D.; et al. Leishmania infantum β-Tubulin Identified by Reverse Engineering Technology through Phage Display Applied as Theranostic Marker for Human Visceral Leishmaniasis. Int. J. Mol. Sci. 2019, 20, 1812. https://doi.org/10.3390/ijms20081812
Costa LE, Alves PT, Carneiro AP, Dias ACS, Fujimura PT, Araujo GR, Tavares GSV, Ramos FF, Duarte MC, Menezes-Souza D, et al. Leishmania infantum β-Tubulin Identified by Reverse Engineering Technology through Phage Display Applied as Theranostic Marker for Human Visceral Leishmaniasis. International Journal of Molecular Sciences. 2019; 20(8):1812. https://doi.org/10.3390/ijms20081812
Chicago/Turabian StyleCosta, Lourena E., Patrícia T. Alves, Ana Paula Carneiro, Ana C. S. Dias, Patrícia T. Fujimura, Galber R. Araujo, Grasiele S. V. Tavares, Fernanda F. Ramos, Mariana C. Duarte, Daniel Menezes-Souza, and et al. 2019. "Leishmania infantum β-Tubulin Identified by Reverse Engineering Technology through Phage Display Applied as Theranostic Marker for Human Visceral Leishmaniasis" International Journal of Molecular Sciences 20, no. 8: 1812. https://doi.org/10.3390/ijms20081812