Changes in the Expression of Aquaporin-3 in the Gastrointestinal Tract Affect Drug Absorption

Abstract
:1. Introduction
2. Results
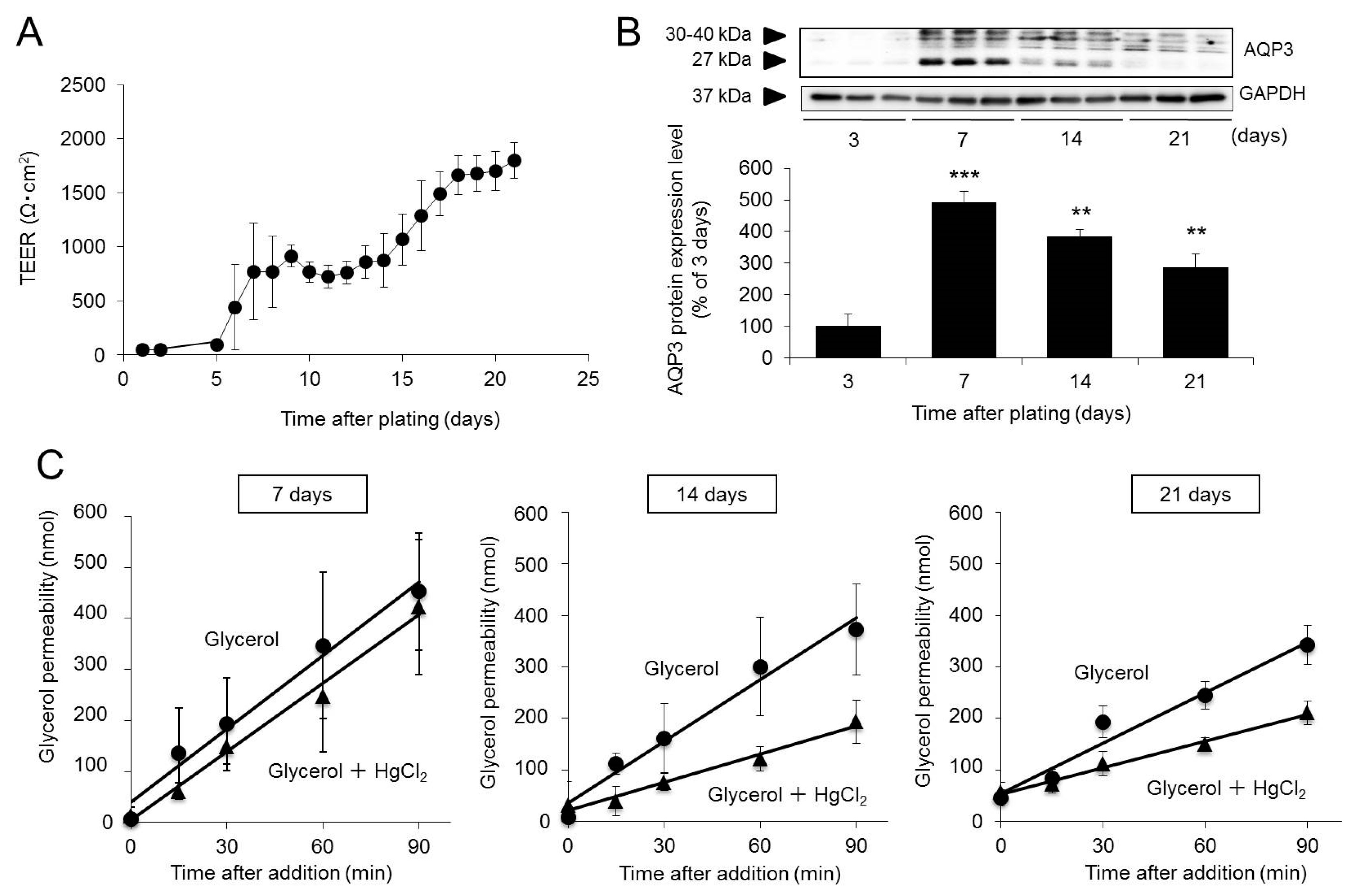
2.1. Changes in the Expression and Function of AQP3 during Caco-2 Cell Differentiation
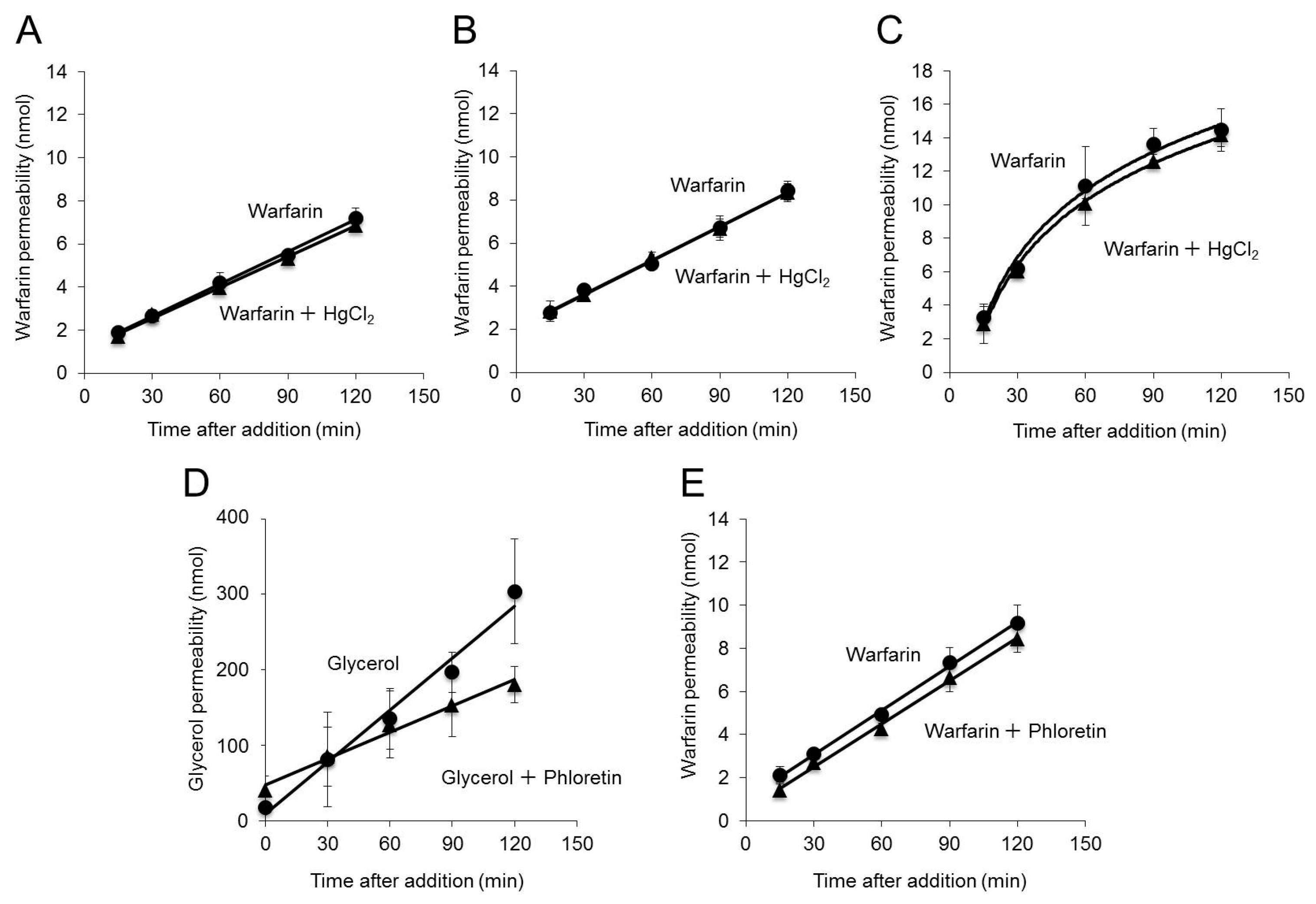
2.2. Effect of Inhibition of AQP3 Function on Warfarin Permeability
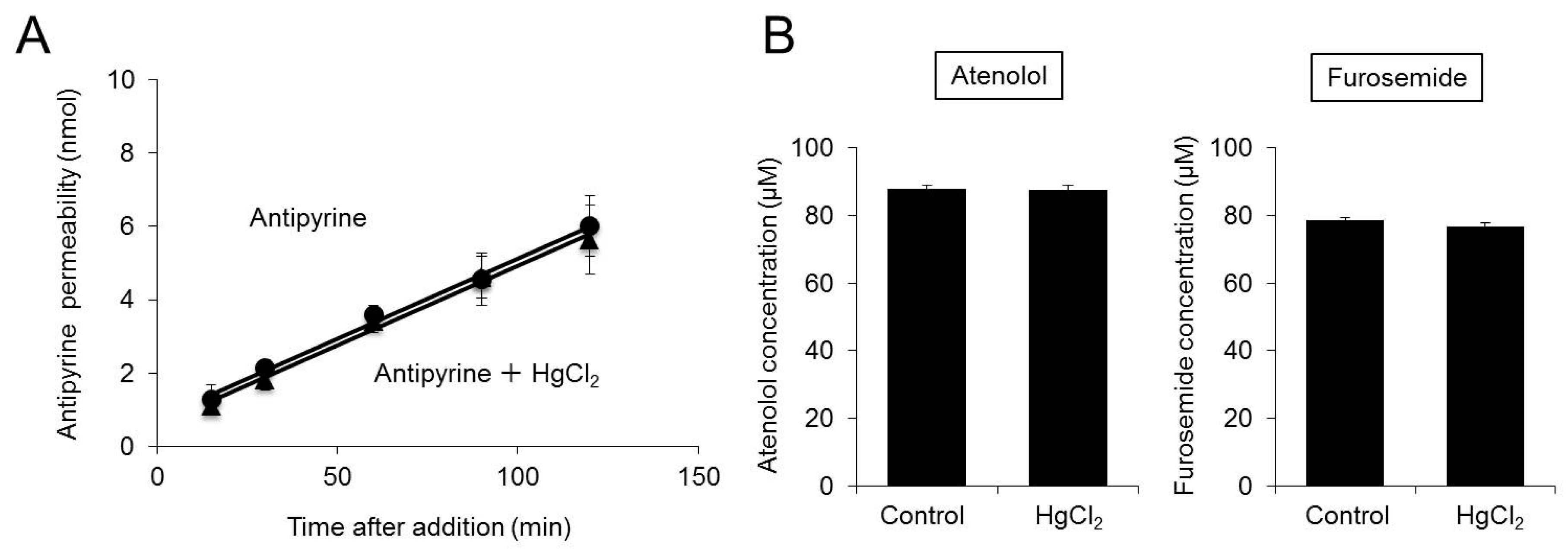
2.3. Effect of Inhibition of AQP3 Function on Permeability to Various Drugs
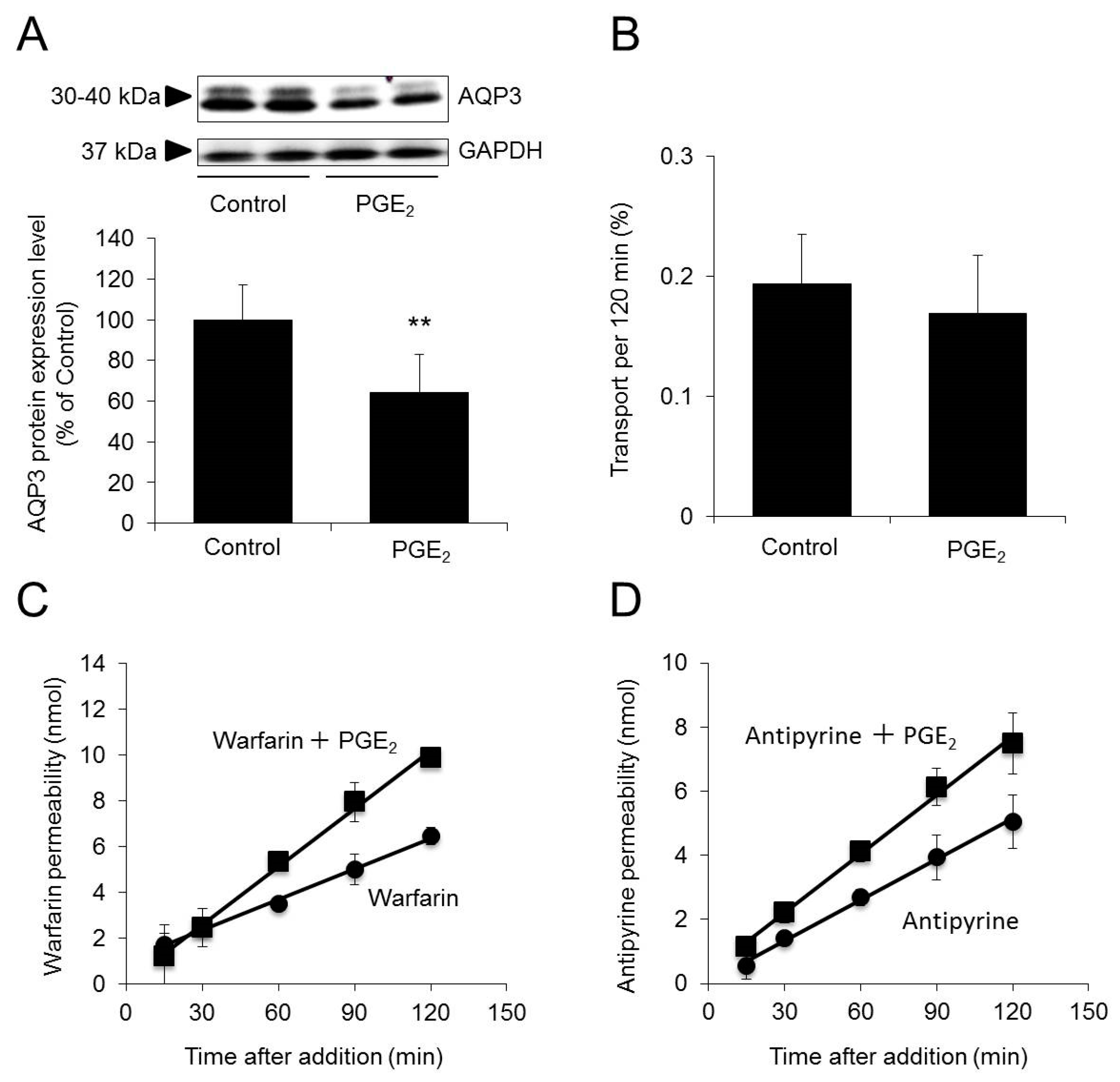
2.4. Effect of Decreased AQP3 Expression on Permeability to Warfarin/Antipyrine
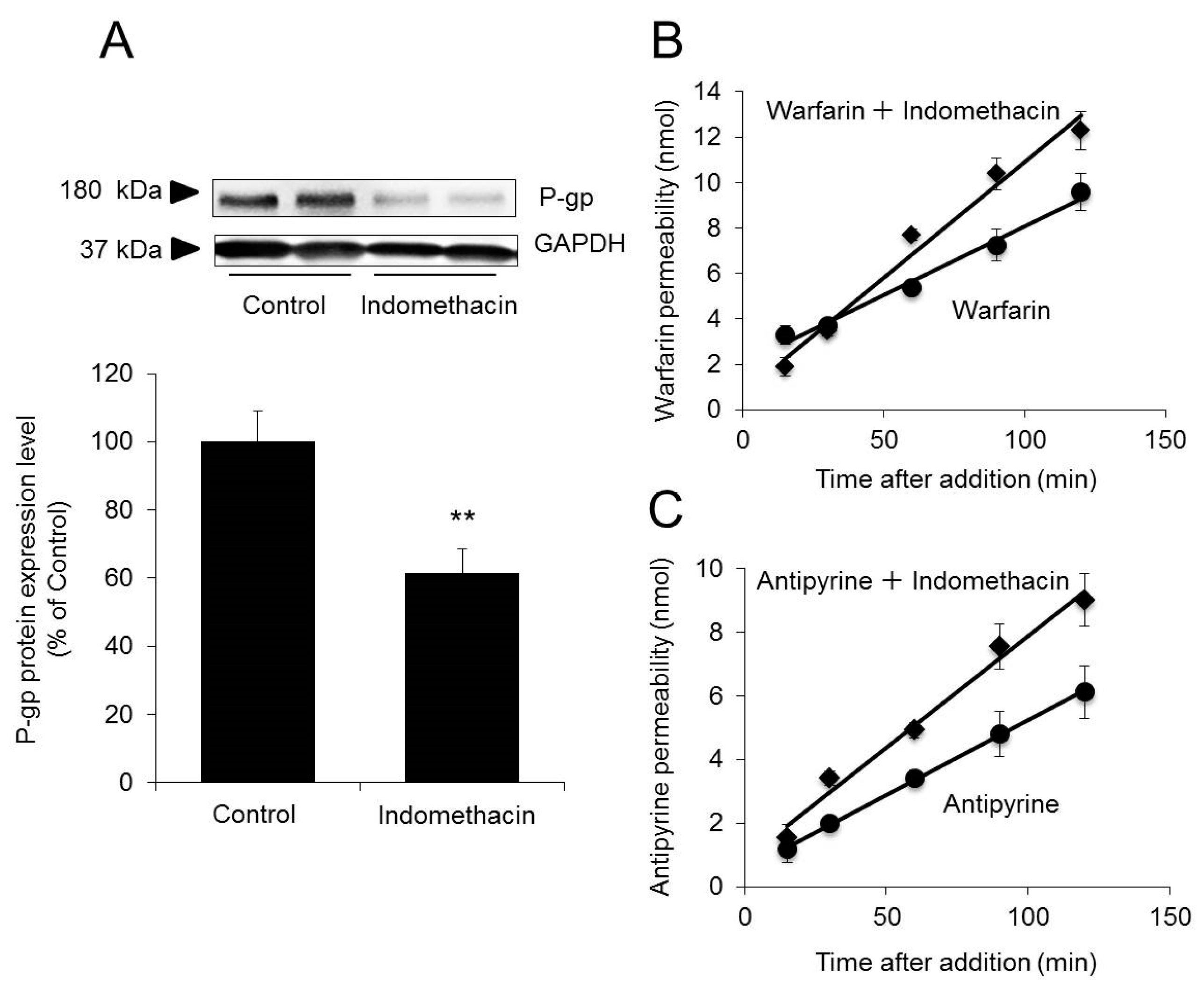
2.5. Effect of Decreased P-gp Expression on Permeability to Warfarin/Antipyrine
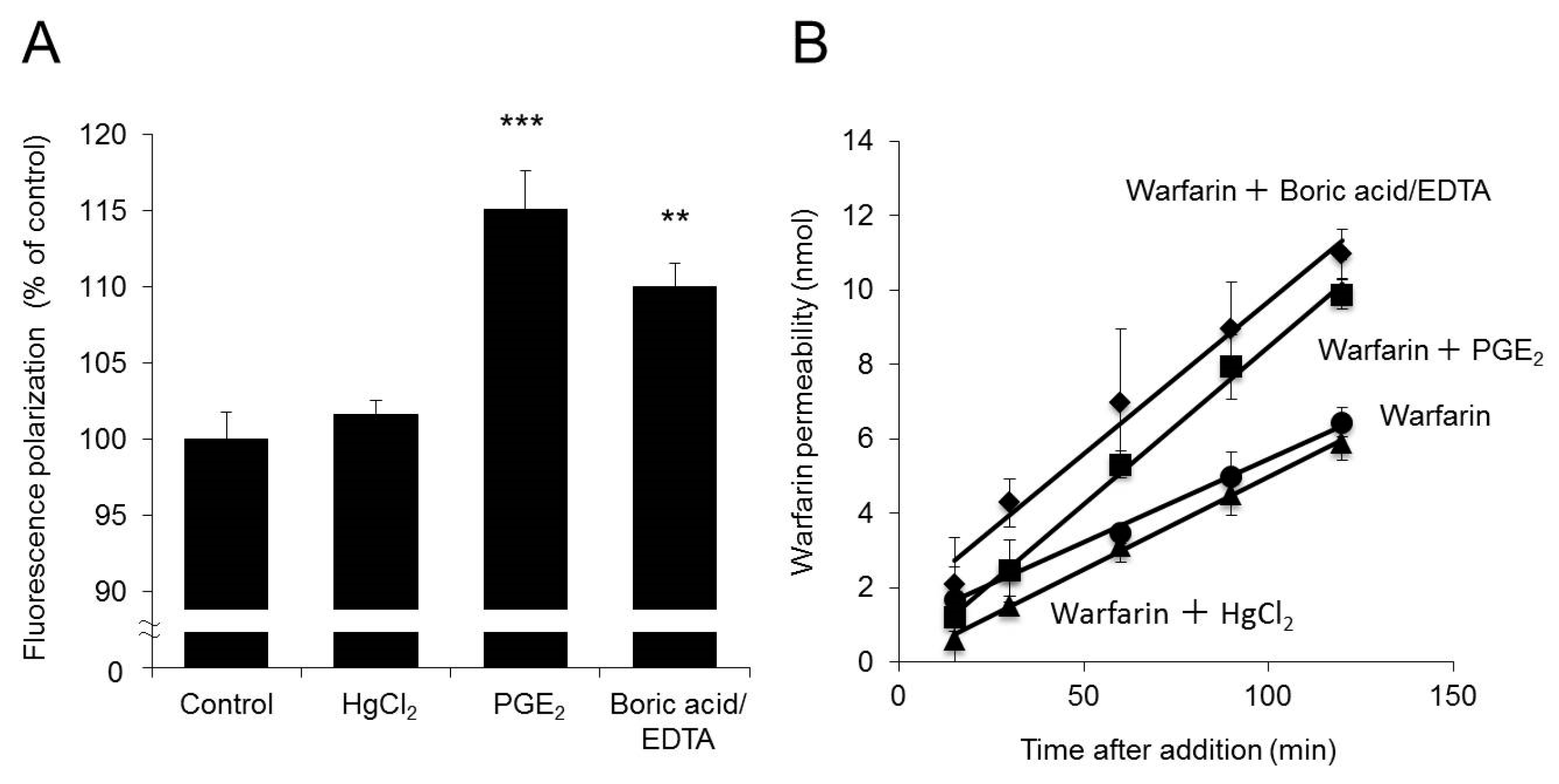
2.6. Effects of Increased Membrane Fluidity on Warfarin Permeability
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of Samples for Western Blotting
4.3. Western Blotting
4.4. Glycerol Permeability Assay
4.5. Drug Permeability Assay
4.6. HPLC
4.7. Lucifer Yellow Permeability Assay
4.8. Measurement of Cell Membrane Fluidity
r = (I// − I)/(I// + 2I⊥)
4.9. Statistical Analyses
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AQPs | Aquaporins |
| GI | Gastrointestinal |
| PGE2 | Prostaglandin E2 |
| P-gp | P-glycoprotein |
| CYP | Cytochrome P450 |
| TEER | Transepithelial Electrical Resistance |
| Papp | Apparent Permeability Coefficient |
| GAPDH | Glyceraldehyde-3-Phosphate Dehydrogenase |
| BCS | Biopharmaceutics Classification System |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| SD | Standard Deviation |
References
- Loo, D.D.; Wright, E.M.; Zeuthen, T. Water pumps. J. Physiol. 2002, 542, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwahara, M.; Shinbo, I.; Sato, K.; Terada, Y.; Marumo, F.; Sasaki, S. Transmembrane helix 5 is critical for the high water permeability of aquaporin. Biochemistry 1999, 38, 16340–16346. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.S.; Bhat, R.V.; Preston, G.M.; Guggino, W.B.; Baraban, J.M.; Agre, P. Molecular characterization of an aquaporin cDNA from brain: Candidate osmoreceptor and regulator of water balance. Proc. Natl. Acad. Sci. USA 1994, 91, 13052–13056. [Google Scholar] [CrossRef] [PubMed]
- Verbavatz, J.M.; Brown, D.; Sabolic, I.; Valenti, G.; Ausiello, D.A.; Van Hoek, A.N.; Ma, T.; Verkman, A.S. Tetrameric assembly of CHIP28 water channels in liposomes and cell membranes: A freeze-fracture study. J. Cell Biol. 1993, 123, 605–618. [Google Scholar] [CrossRef]
- Verkman, A.S. More than just water channels: Unexpected cellular roles of aquaporins. J. Cell Sci. 2005, 118, 3225–3232. [Google Scholar] [CrossRef]
- King, L.S.; Kozono, D.; Agre, P. From structure to disease: The evolving tale of aquaporin biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, P.; Cid, L.P.; Vio, C.P.; Sepulveda, F.V. Aquaporin-2, a regulated water channel, is expressed in apical membranes of rat distal colon epithelium. Am. J. Physiol. 2001, 281, G856–G863. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, N.; Kon, R.; Sugiyama, K. Aquaporins in the Colon as a New Therapeutic Target in Diarrhea and Constipation. Int. J. Mol. Sci. 2016, 17, 1172. [Google Scholar] [CrossRef]
- Koyama, Y.; Yamamoto, T.; Tani, T.; Nihei, K.; Kondo, D.; Funaki, H.; Yaoita, E.; Kawasaki, K.; Sato, N.; Hatakeyama, K.; et al. Expression and localization of aquaporins in rat gastrointestinal tract. Am. J. Physiol. 1999, 276, C621–C627. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, T.; Tajika, Y.; Ablimit, A.; Aoki, T.; Hagiwara, H.; Takata, K. Aquaporins in the digestive system. Med. Electron Microsc. 2004, 37, 71–80. [Google Scholar] [CrossRef]
- Silberstein, C.; Kierbel, A.; Amodeo, G.; Zotta, E.; Bigi, F.; Berkowski, D.; Ibarra, C. Functional characterization and localization of AQP3 in the human colon. Braz. J. Med. Biol. Res. 1999, 32, 1303–1313. [Google Scholar] [CrossRef] [Green Version]
- Zahn, A.; Moehle, C.; Langmann, T.; Ehehalt, R.; Autschbach, F.; Stremmel, W.; Schmitz, G. Aquaporin-8 expression is reduced in ileum and induced in colon of patients with ulcerative colitis. World, J. Gastroenterol. 2007, 13, 1687–1695. [Google Scholar] [CrossRef] [Green Version]
- Kon, R.; Yamamura, M.; Matsunaga, Y.; Kimura, H.; Minami, M.; Kato, S.; Ikarashi, N.; Sugiyama, K. Laxative effect of repeated Daiokanzoto is attributable to decrease in aquaporin-3 expression in the colon. J. Nat. Med. 2018, 72, 493–502. [Google Scholar] [CrossRef]
- Kon, R.; Tsubota, Y.; Minami, M.; Kato, S.; Matsunaga, Y.; Kimura, H.; Murakami, Y.; Fujikawa, T.; Sakurai, R.; Tomimoto, R.; et al. CPT-11-Induced Delayed Diarrhea Develops via Reduced Aquaporin-3 Expression in the Colon. Int. J. Mol. Sci. 2018, 19, 170. [Google Scholar] [CrossRef]
- Kon, R.; Ikarashi, N.; Hayakawa, A.; Haga, Y.; Fueki, A.; Kusunoki, Y.; Tajima, M.; Ochiai, W.; Machida, Y.; Sugiyama, K. Morphine-Induced Constipation Develops With Increased Aquaporin-3 Expression in the Colon via Increased Serotonin Secretion. Toxicol. Sci. 2015, 145, 337–347. [Google Scholar] [CrossRef] [Green Version]
- Kon, R.; Ikarashi, N.; Nagoya, C.; Takayama, T.; Kusunoki, Y.; Ishii, M.; Ueda, H.; Ochiai, W.; Machida, Y.; Sugita, K.; et al. Rheinanthrone, a metabolite of sennoside A, triggers macrophage activation to decrease aquaporin-3 expression in the colon, causing the laxative effect of rhubarb extract. J. Ethnopharmacol. 2014, 152, 190–200. [Google Scholar] [CrossRef]
- Ikarashi, N.; Kon, R.; Iizasa, T.; Suzuki, N.; Hiruma, R.; Suenaga, K.; Toda, T.; Ishii, M.; Hoshino, M.; Ochiai, W.; et al. Inhibition of aquaporin-3 water channel in the colon induces diarrhea. Biol. Pharm. Bull. 2012, 35, 957–962. [Google Scholar] [CrossRef]
- Ikarashi, N.; Baba, K.; Ushiki, T.; Kon, R.; Mimura, A.; Toda, T.; Ishii, M.; Ochiai, W.; Sugiyama, K. The laxative effect of bisacodyl is attributable to decreased aquaporin-3 expression in the colon induced by increased PGE2 secretion from macrophages. Am. J. Physiol. 2011, 301, G887–G895. [Google Scholar] [CrossRef]
- Suzuki, H.; Sugiyama, Y. Role of metabolic enzymes and efflux transporters in the absorption of drugs from the small intestine. Eur. J. Pharm. Sci. 2000, 12, 3–12. [Google Scholar] [CrossRef]
- Drescher, S.; Glaeser, H.; Murdter, T.; Hitzl, M.; Eichelbaum, M.; Fromm, M.F. P-glycoprotein-mediated intestinal and biliary digoxin transport in humans. Clin. Pharmacol. Ther. 2003, 73, 223–231. [Google Scholar] [CrossRef]
- Fromm, M.F.; Kauffmann, H.M.; Fritz, P.; Burk, O.; Kroemer, H.K.; Warzok, R.W.; Eichelbaum, M.; Siegmund, W.; Schrenk, D. The effect of rifampin treatment on intestinal expression of human MRP transporters. Am. J. Pathol. 2000, 157, 1575–1580. [Google Scholar] [CrossRef]
- Greiner, B.; Eichelbaum, M.; Fritz, P.; Kreichgauer, H.P.; von Richter, O.; Zundler, J.; Kroemer, H.K. The role of intestinal P-glycoprotein in the interaction of digoxin and rifampin. J. Clin. Investig. 1999, 104, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Chan, E.; Pan, S.Q.; Huang, M.; Lee, E.J. Pharmacokinetic interactions of drugs with St John’s wort. J Psychopharmacol 2004, 18, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.Z.; Jang, G.R.; Tsunoda, S. Dietary effects on drug metabolism and transport. Clin. Pharmacokinet. 2003, 42, 1071–1088. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, I.J.; Raub, T.J.; Borchardt, R.T. Characterization of the human colon carcinoma cell line (Caco-2) as a model system for intestinal epithelial permeability. Gastroenterology 1989, 96, 736–749. [Google Scholar] [CrossRef]
- Fleet, J.C.; Wood, R.J. Specific 1,25(OH)2D3-mediated regulation of transcellular calcium transport in Caco-2 cells. Am. J. Physiol. 1999, 276, G958–G964. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.P.; Marrone, A.; Ciancetta, A.; Galan Cobo, A.; Echevarria, M.; Moura, T.F.; Re, N.; Casini, A.; Soveral, G. Targeting aquaporin function: Potent inhibition of aquaglyceroporin-3 by a gold-based compound. PLoS ONE 2012, 7, e37435. [Google Scholar] [CrossRef]
- Yang, B.; Verkman, A.S. Water and glycerol permeabilities of aquaporins 1-5 and MIP determined quantitatively by expression of epitope-tagged constructs in Xenopus oocytes. J. Biol. Chem. 1997, 272, 16140–16146. [Google Scholar] [CrossRef]
- Kuwahara, M.; Gu, Y.; Ishibashi, K.; Marumo, F.; Sasaki, S. Mercury-sensitive residues and pore site in AQP3 water channel. Biochemistry 1997, 36, 13973–13978. [Google Scholar] [CrossRef]
- Karlsson, B.C.; Olsson, G.D.; Friedman, R.; Rosengren, A.M.; Henschel, H.; Nicholls, I.A. How warfarin’s structural diversity influences its phospholipid bilayer membrane permeation. J. Phys. Chem. B 2013, 117, 2384–2395. [Google Scholar] [CrossRef]
- Smetanova, L.; Stetinova, V.; Svoboda, Z.; Kvetina, J. Caco-2 cells, biopharmaceutics classification system (BCS) and biowaiver. Acta Med. 2011, 54, 3–8. [Google Scholar]
- Zrieki, A.; Farinotti, R.; Buyse, M. Cyclooxygenase inhibitors down regulate P-glycoprotein in human colorectal Caco-2 cell line. Pharm. Res. 2008, 25, 1991–2001. [Google Scholar] [CrossRef]
- Barta, C.A.; Sachs-Barrable, K.; Feng, F.; Wasan, K.M. Effects of monoglycerides on P-glycoprotein: Modulation of the activity and expression in Caco-2 cell monolayers. Mol. Pharm. 2008, 5, 863–875. [Google Scholar] [CrossRef]
- Kikuchi, T.; Suzuki, M.; Kusai, A.; Iseki, K.; Sasaki, H.; Nakashima, K. Mechanism of permeability-enhancing effect of EDTA and boric acid on the corneal penetration of 4-[1-hydroxy-1-methylethyl]-2-propyl-1-[4-[2-[tetrazole-5-yl]phenyl]phenyl] methylimidazole-5-carboxylic acid monohydrate (CS-088). Int. J. Pharm. 2005, 299, 107–114. [Google Scholar] [CrossRef]
- Yasui, H.; Kubota, M.; Iguchi, K.; Usui, S.; Kiho, T.; Hirano, K. Membrane trafficking of aquaporin 3 induced by epinephrine. Biochem. Biophys. Res. Commun. 2008, 373, 613–617. [Google Scholar] [CrossRef]
- Higuchi, S.; Kubota, M.; Iguchi, K.; Usui, S.; Kiho, T.; Hirano, K. Transcriptional regulation of aquaporin 3 by insulin. J. Cell. Biochem. 2007, 102, 1051–1058. [Google Scholar] [CrossRef]
- Campos, E.; Moura, T.F.; Oliva, A.; Leandro, P.; Soveral, G. Lack of Aquaporin 3 in bovine erythrocyte membranes correlates with low glycerol permeation. Biochem. Biophys. Res. Commun. 2011, 408, 477–481. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Lu, D.; Zhang, Y.; Li, J.; Fang, Y.; Li, F.; Sun, J. Critical role of aquaporin-3 in epidermal growth factor-induced migration of colorectal carcinoma cells and its clinical significance. Oncol. Rep. 2013, 29, 535–540. [Google Scholar] [CrossRef]
- Alleva, K.; Chara, O.; Amodeo, G. Aquaporins: Another piece in the osmotic puzzle. FEBS Lett. 2012, 586, 2991–2999. [Google Scholar] [CrossRef] [Green Version]
- Zeuthen, T. Water-transporting proteins. J. Membr. Biol. 2010, 234, 57–73. [Google Scholar] [CrossRef]
- Karlsson, J.; Artursson, P. A new diffusion chamber system for the determination of drug permeability coefficients across the human intestinal epithelium that are independent of the unstirred water layer. Biochim. Biophys. Acta 1992, 1111, 204–210. [Google Scholar] [CrossRef]
- Velicky, M.; Tam, K.Y.; Dryfe, R.A. In situ artificial membrane permeation assay under hydrodynamic control: Correlation between drug in vitro permeability and fraction absorbed in humans. Eur. J. Pharm. Sci. 2011, 44, 299–309. [Google Scholar] [CrossRef]
- Prachayasittikul, V.; Isarankura-Na-Ayudhya, C.; Tantimongcolwat, T.; Nantasenamat, C.; Galla, H.J. EDTA-induced membrane fluidization and destabilization: Biophysical studies on artificial lipid membranes. Acta Biochim. Biophys. Sin. 2007, 39, 901–913. [Google Scholar] [CrossRef]
- Wilson, J.L.; Miranda, C.A.; Knepper, M.A. Vasopressin and the regulation of aquaporin-2. Clin. Exp. Nephrol. 2013, 17, 751–764. [Google Scholar] [CrossRef]
- Lebeck, J.; Gena, P.; O’Neill, H.; Skowronski, M.T.; Lund, S.; Calamita, G.; Praetorius, J. Estrogen prevents increased hepatic aquaporin-9 expression and glycerol uptake during starvation. Am. J. Physiol. 2012, 302, G365–G374. [Google Scholar] [CrossRef] [Green Version]
- Horie, I.; Maeda, M.; Yokoyama, S.; Hisatsune, A.; Katsuki, H.; Miyata, T.; Isohama, Y. Tumor necrosis factor-alpha decreases aquaporin-3 expression in DJM-1 keratinocytes. Biochem. Biophys. Res. Commun. 2009, 387, 564–568. [Google Scholar] [CrossRef]
- Ito, H.; Yamamoto, N.; Arima, H.; Hirate, H.; Morishima, T.; Umenishi, F.; Tada, T.; Asai, K.; Katsuya, H.; Sobue, K. Interleukin-1beta induces the expression of aquaporin-4 through a nuclear factor-κB pathway in rat astrocytes. J. Neurochem. 2006, 99, 107–118. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Sun, J.Y.; Wang, J.Y.; Bai, C.X.; He, B.M. Terlipressin resolves ascites of cirrhotic rats through downregulation of aquaporin 2. J. Int. Med. Res. 2012, 40, 1735–1744. [Google Scholar] [CrossRef]
- Dong, C.; Wang, G.; Li, B.; Xiao, K.; Ma, Z.; Huang, H.; Wang, X.; Bai, C. Anti-asthmatic agents alleviate pulmonary edema by upregulating AQP1 and AQP5 expression in the lungs of mice with OVA-induced asthma. Respir. Physiol. Neurobiol. 2012, 181, 21–28. [Google Scholar] [CrossRef]
- Volpe, D.A. Variability in Caco-2 and MDCK cell-based intestinal permeability assays. J. Pharm. Sci. 2008, 97, 712–725. [Google Scholar] [CrossRef]
- Markowska, M.; Oberle, R.; Juzwin, S.; Hsu, C.P.; Gryszkiewicz, M.; Streeter, A.J. Optimizing Caco-2 cell monolayers to increase throughput in drug intestinal absorption analysis. J. Pharmacol. Toxicol. Methods 2001, 46, 51–55. [Google Scholar] [CrossRef]
- Wang, C.; Chen, G.; Jiang, J.; Qiu, L.; Hosoi, K.; Yao, C. Aquaglyceroporins are involved in uptake of arsenite into murine gastrointestinal tissues. J. Med. Investig. 2009, 56, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Zelenina, M.; Tritto, S.; Bondar, A.A.; Zelenin, S.; Aperia, A. Copper inhibits the water and glycerol permeability of aquaporin-3. J. Biol. Chem. 2004, 279, 51939–51943. [Google Scholar] [CrossRef]
- Volpe, D.A. Permeability classification of representative fluoroquinolones by a cell culture method. AAPS J. 2004, 6, 1–6. [Google Scholar] [CrossRef]
- Ohkubo, R.; Tomita, M.; Hotta, Y.; Nagira, M.; Hayashi, M. Comparative study of flux of FITC-labeled Dextran 4000 on normal (iso)- and hyper-osmolarity in basal side in caco-2 cell monolayers. Drug Metab. Pharmacokinet. 2003, 18, 404–408. [Google Scholar] [CrossRef]
- Inokuchi, H.; Takei, T.; Aikawa, K.; Shimizu, M. The effect of hyperosmosis on paracellular permeability in Caco-2 cell monolayers. Biosci. Biotechnol. Biochem. 2009, 73, 328–334. [Google Scholar] [CrossRef]
- Korjamo, T.; Heikkinen, A.T.; Monkkonen, J. Analysis of unstirred water layer in in vitro permeability experiments. J. Pharm. Sci. 2009, 98, 4469–4479. [Google Scholar] [CrossRef]
- D’Souza, V.M.; Shertzer, H.G.; Menon, A.G.; Pauletti, G.M. High glucose concentration in isotonic media alters caco-2 cell permeability. AAPS PharmSci. 2003, 5, E24. [Google Scholar]
- Zeng, N.; Gao, X.; Hu, Q.; Song, Q.; Xia, H.; Liu, Z.; Gu, G.; Jiang, M.; Pang, Z.; Chen, H.; et al. Lipid-based liquid crystalline nanoparticles as oral drug delivery vehicles for poorly water-soluble drugs: Cellular interaction and in vivo absorption. Int. J. Nanomed. 2012, 7, 3703–3718. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycerol (10−6 cm/s) | Glycerol + HgCl2 (10−6 cm/s) | |
|---|---|---|
| 7 days | 2380 ± 715 | 2172 ± 779 |
| 14 days | 1986 ± 1222 | 929 ± 101 * |
| 21 days | 1000 ± 148 | 415 ± 66 ** |
| Condition | Warfarin (10−6 cm/s) | Warfarin + HgCl2 (10−6 cm/s) |
|---|---|---|
| Normal condition | 25.2 ± 4.7 | 24.2 ± 1.9 |
| Osmotic pressure difference condition | 24.1 ± 2.2 | 24.2 ± 4.4 |
| Stirring condition | 54.6 ± 1.8 | 53.3 ± 5.4 |
| Drug | Mobile Phase | Wavelength (nm) | Flow Rate (mL/min) | Retention Time (min) |
|---|---|---|---|---|
| Warfarin | 20 mM phosphate buffer:methanol = 55:45 | 308 | 1.0 | 10.0 |
| Antipyrine | 20 mM phosphate buffer:methanol = 70:30 | 254 | 1.0 | 12.9 |
| Atenolol | 50 mM phosphate buffer:acetonitrile = 90:10 | 226 | 1.0 | 5.2 |
| Furosemide | 50 mM phosphate buffer:methanol = 60:40 | 280 | 1.2 | 6.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikarashi, N.; Nagoya, C.; Kon, R.; Kitaoka, S.; Kajiwara, S.; Saito, M.; Kawabata, A.; Ochiai, W.; Sugiyama, K. Changes in the Expression of Aquaporin-3 in the Gastrointestinal Tract Affect Drug Absorption. Int. J. Mol. Sci. 2019, 20, 1559. https://doi.org/10.3390/ijms20071559
Ikarashi N, Nagoya C, Kon R, Kitaoka S, Kajiwara S, Saito M, Kawabata A, Ochiai W, Sugiyama K. Changes in the Expression of Aquaporin-3 in the Gastrointestinal Tract Affect Drug Absorption. International Journal of Molecular Sciences. 2019; 20(7):1559. https://doi.org/10.3390/ijms20071559
Chicago/Turabian StyleIkarashi, Nobutomo, Chika Nagoya, Risako Kon, Satoshi Kitaoka, Sayuri Kajiwara, Masayo Saito, Akane Kawabata, Wataru Ochiai, and Kiyoshi Sugiyama. 2019. "Changes in the Expression of Aquaporin-3 in the Gastrointestinal Tract Affect Drug Absorption" International Journal of Molecular Sciences 20, no. 7: 1559. https://doi.org/10.3390/ijms20071559