Unique Regulation of Enterocyte Brush Border Membrane Na-Glutamine and Na-Alanine Co-Transport by Peroxynitrite during Chronic Intestinal Inflammation

 , and
, and
Abstract
:1. Introduction
2. Results
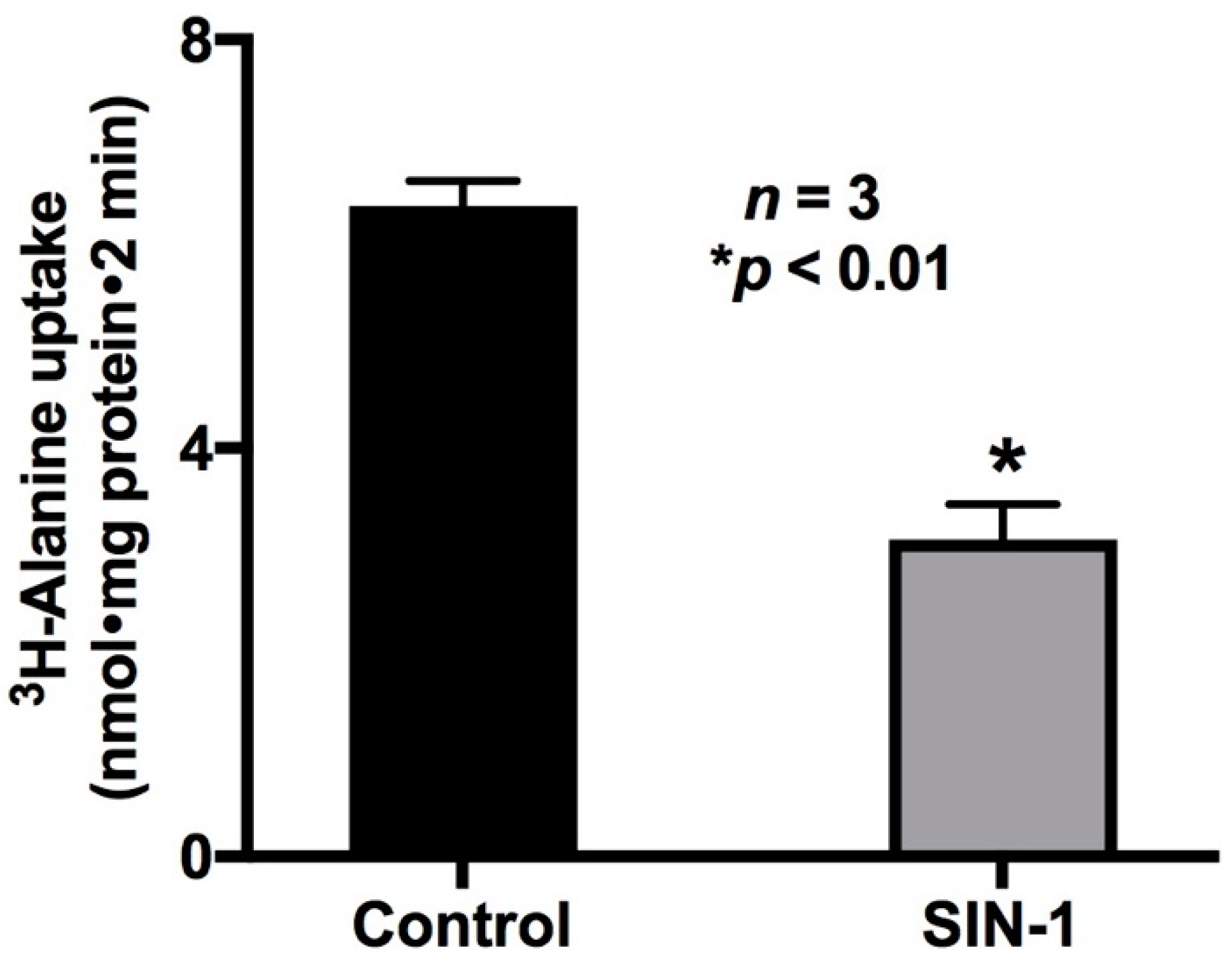
2.1. Effect of OONO on Na-Alanine Co-Transport Activity in IEC-18 Cells
2.2. Effect of OONO on Na/K-ATPase Activity in IEC-18 Cells
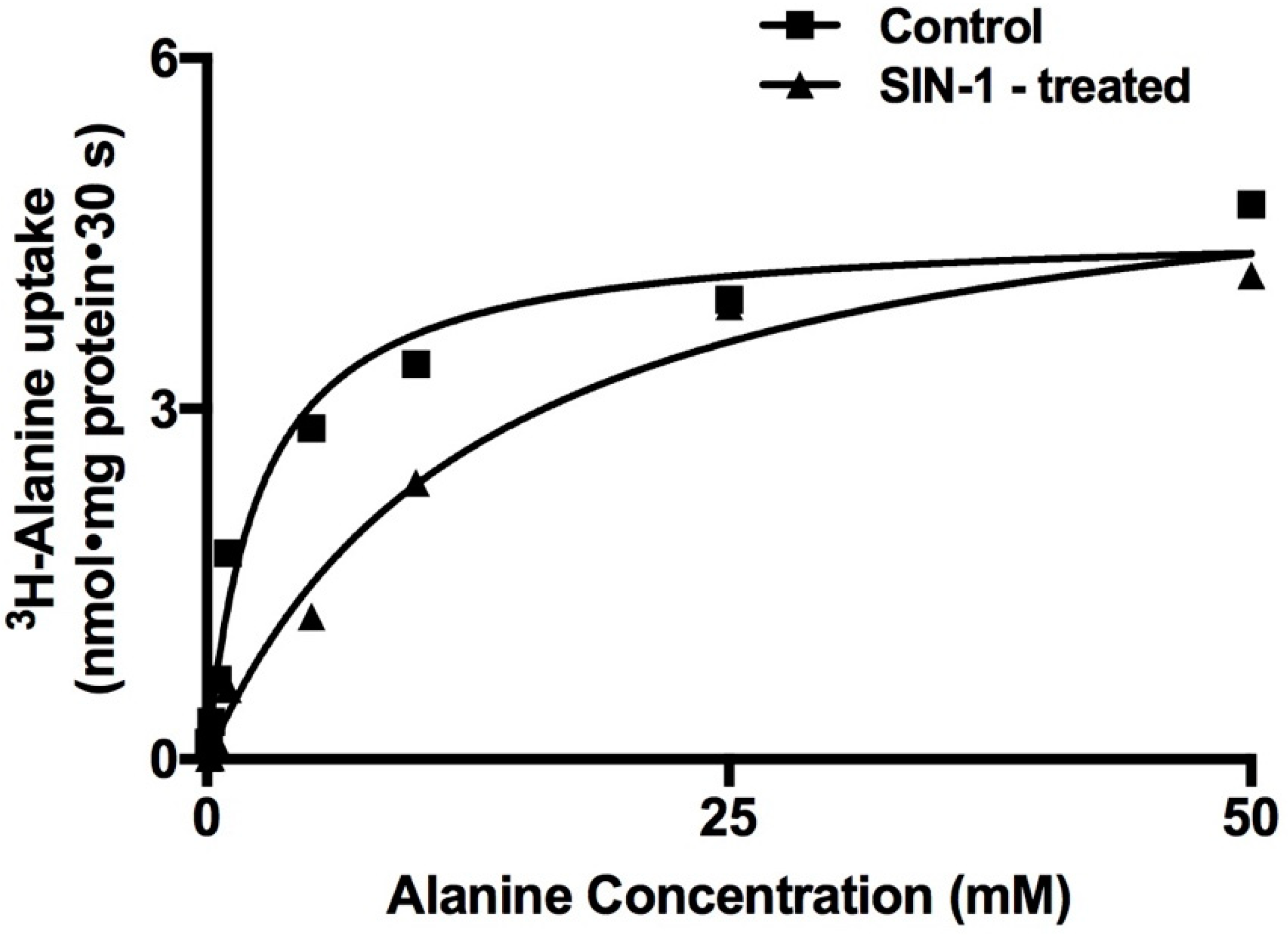
2.3. Kinetic Studies for the Inhibition of Na-Alanine Co-Transport Activity
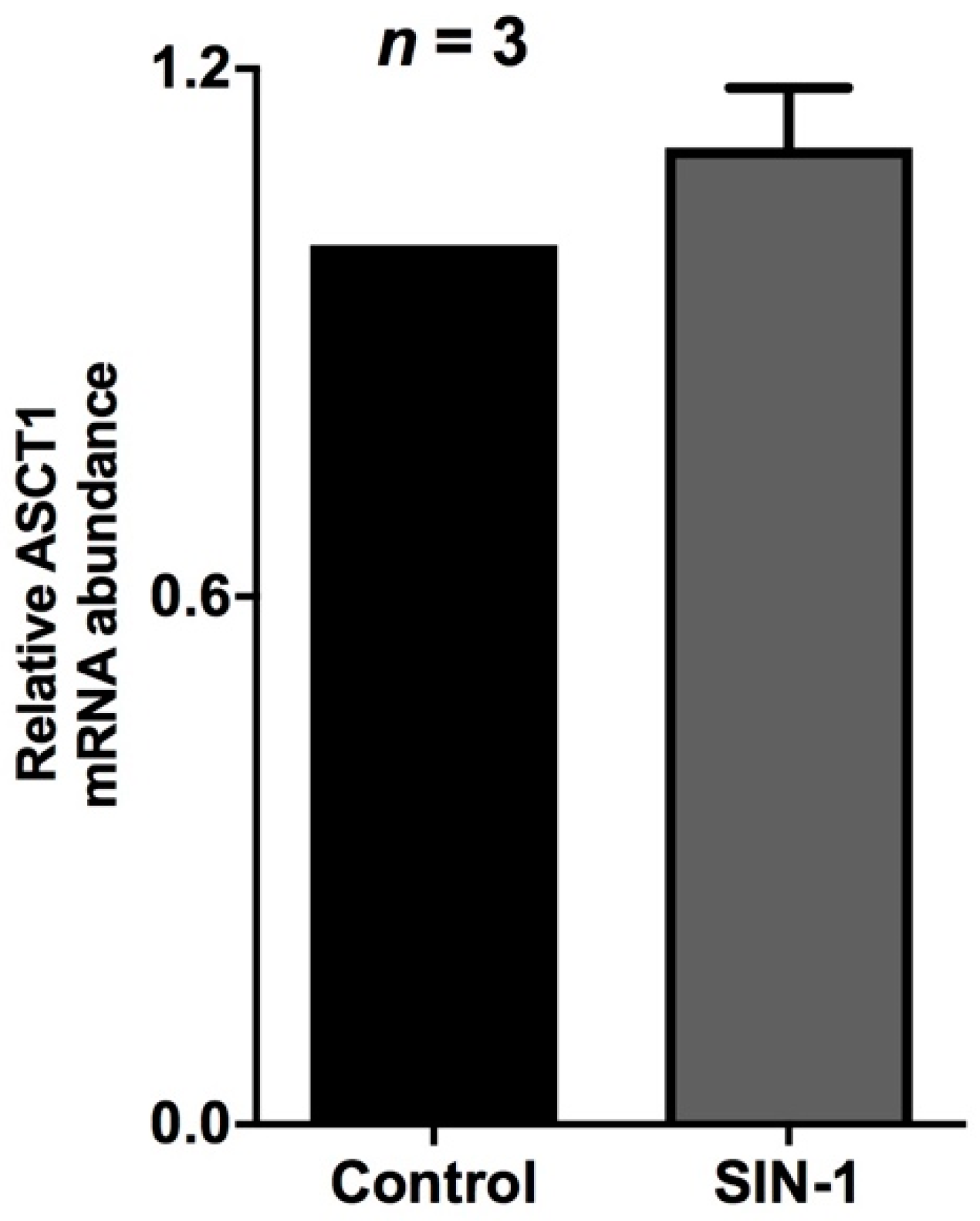
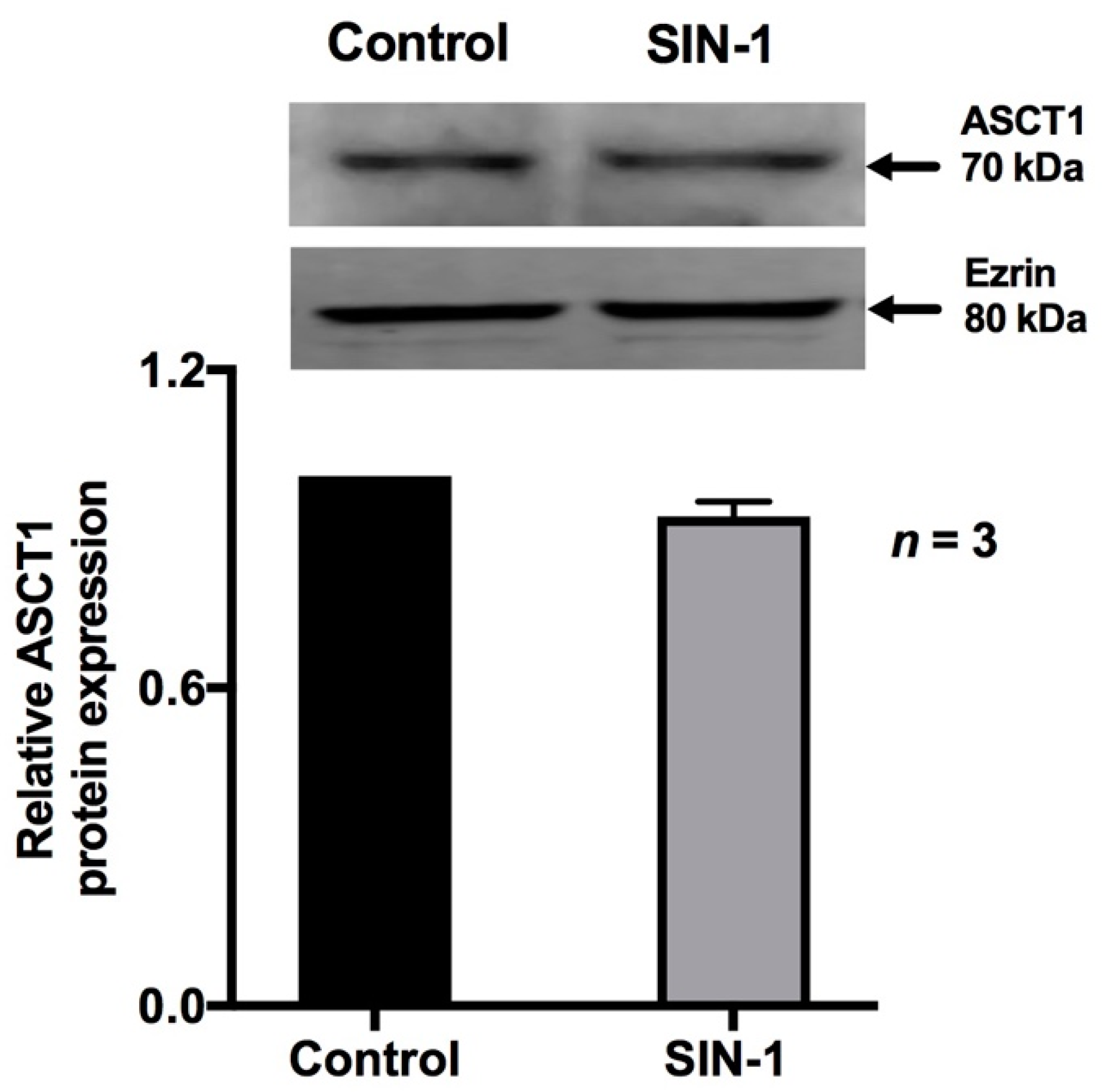
2.4. Effect of OONO on ASCT1 Expression
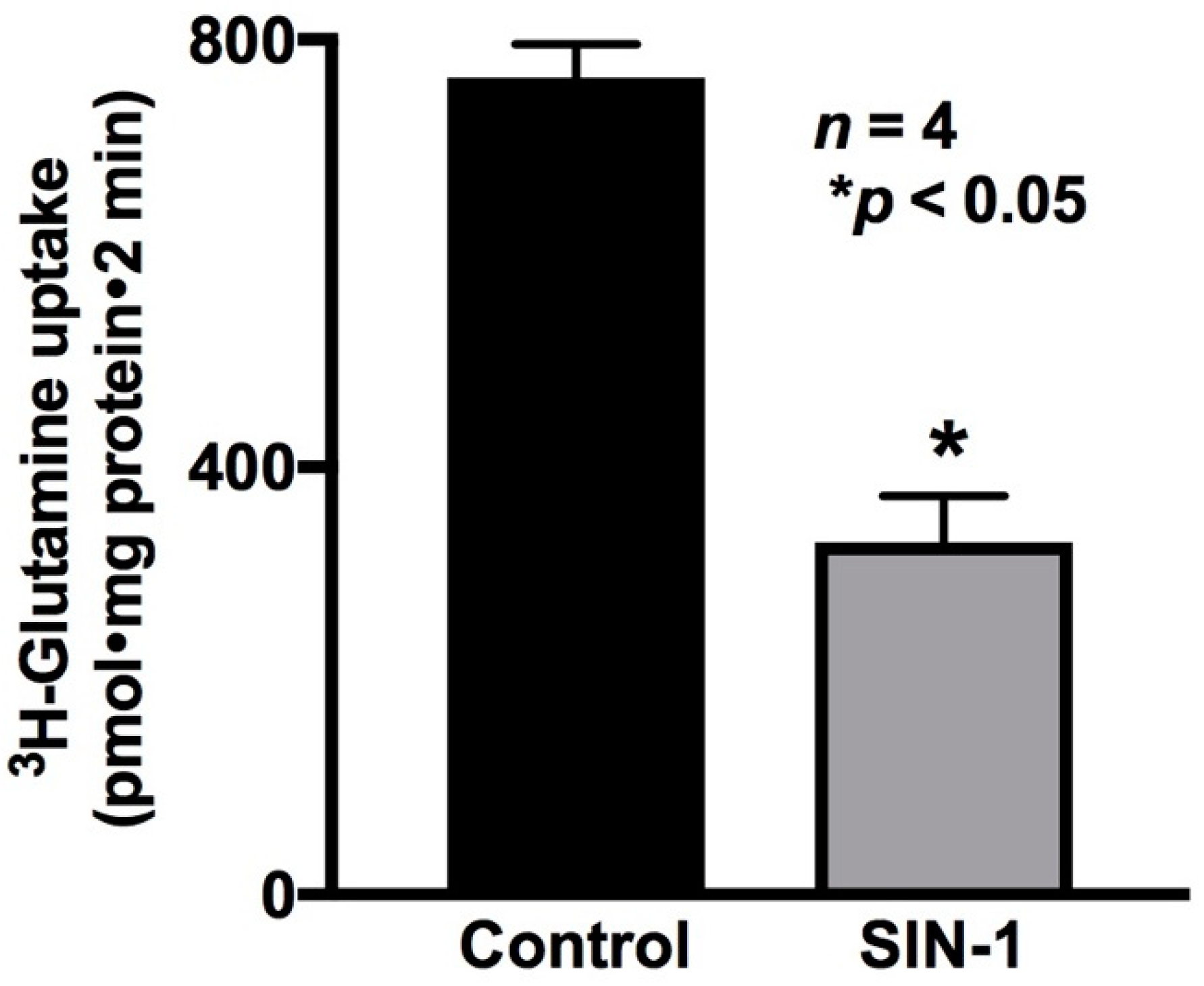
2.5. Effect of OONO on Na-Glutamine Co-Transport Activity in IEC-18 Cells
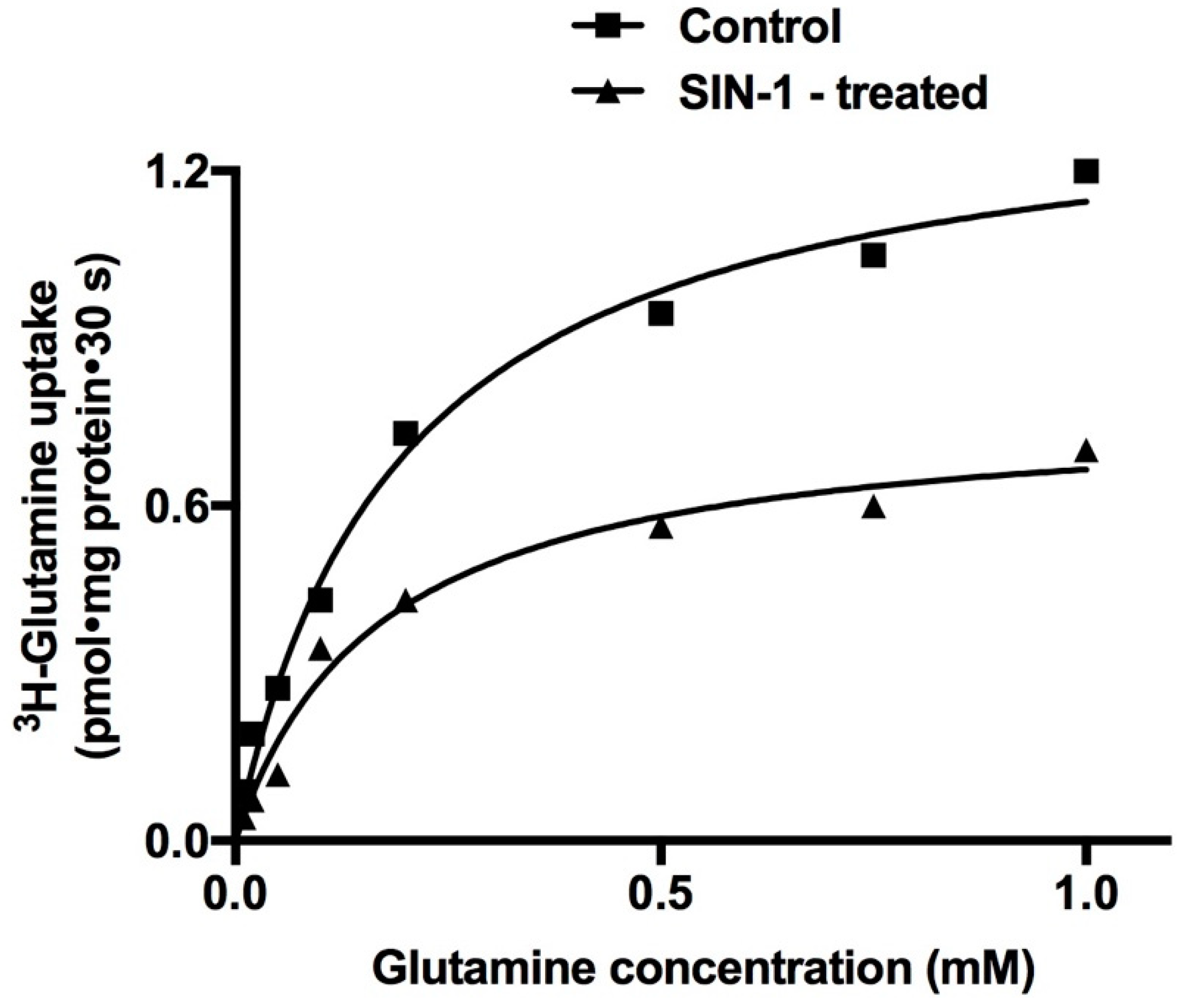
2.6. Kinetic Studies for the Inhibition of Na-Glutamine Co-Transport Activity
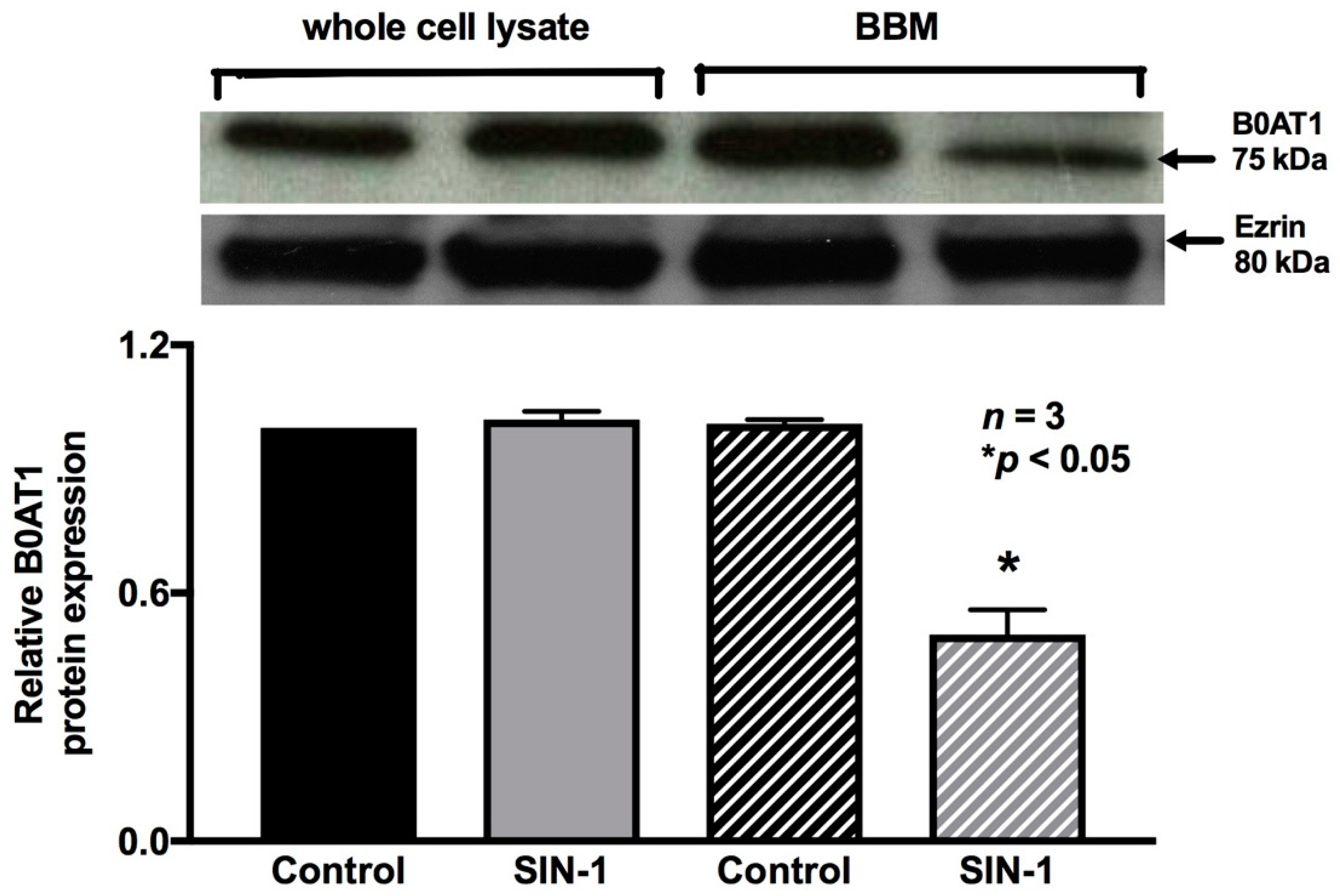
2.7. Effect of OONO on B0AT1 Expression
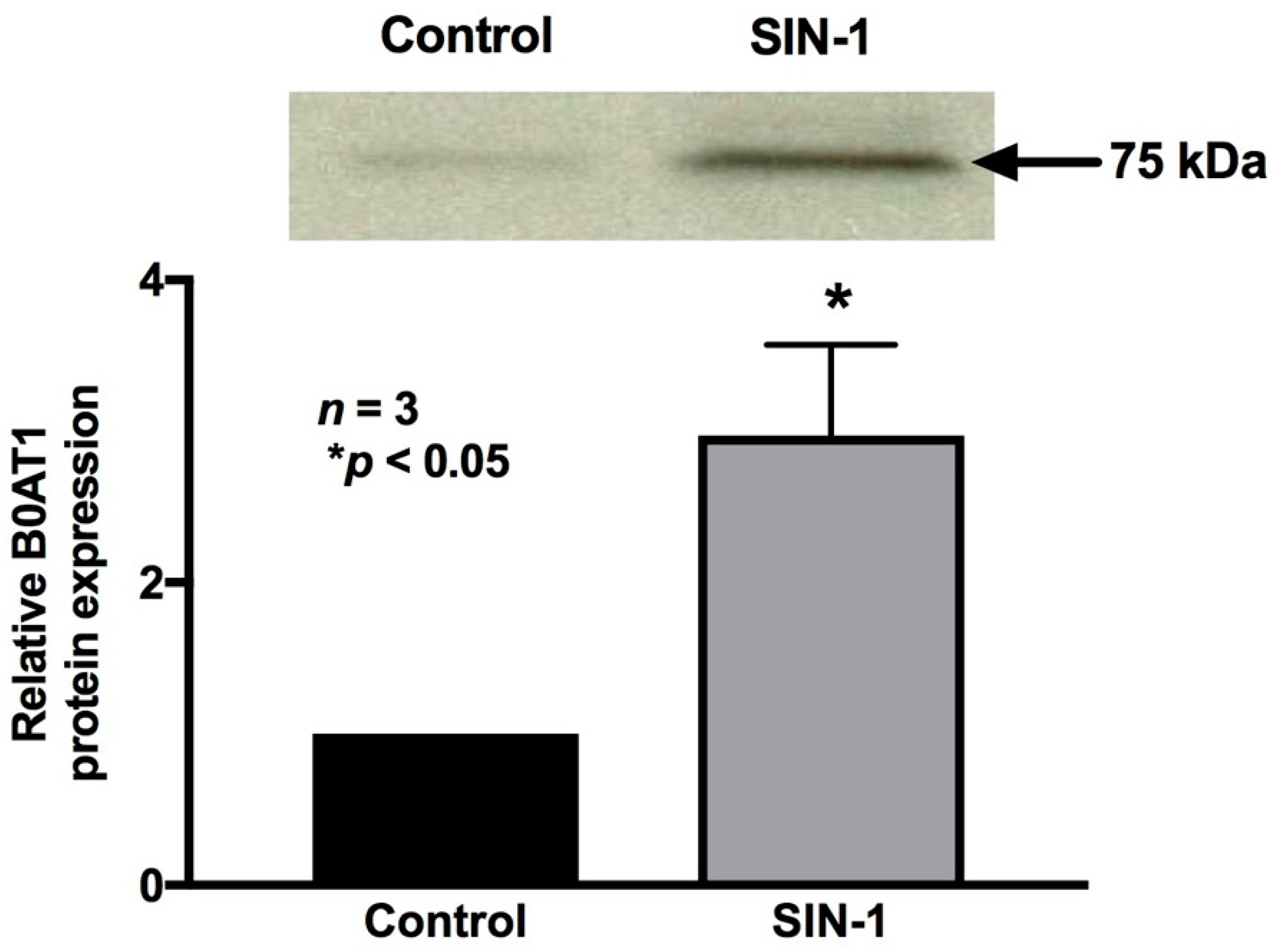
2.8. 3-Nitrotyrosine Formation
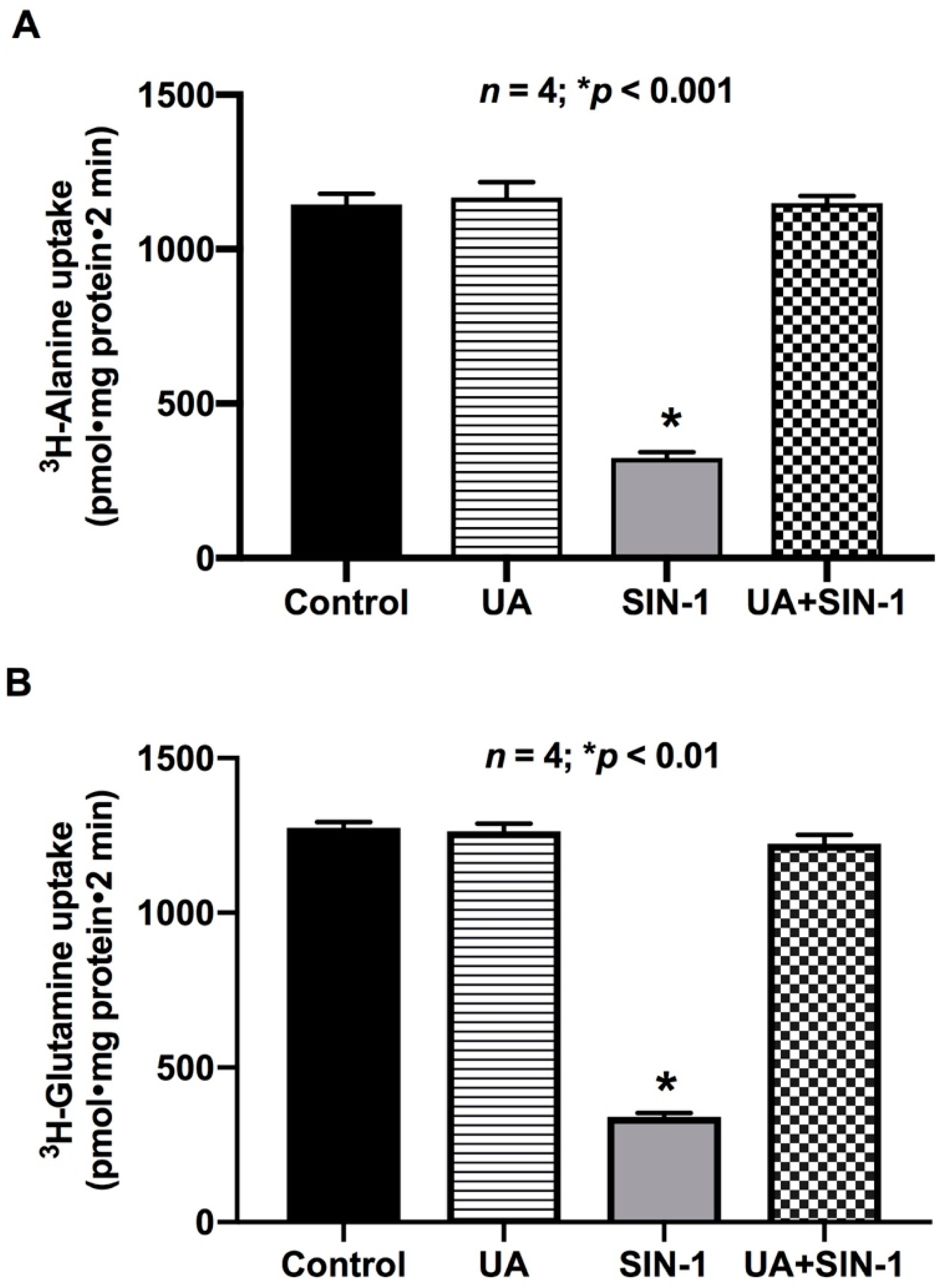
2.9. Uric Acid (UA) Reverses the Effect of OONO on ASCT1 and B0AT1 Activity
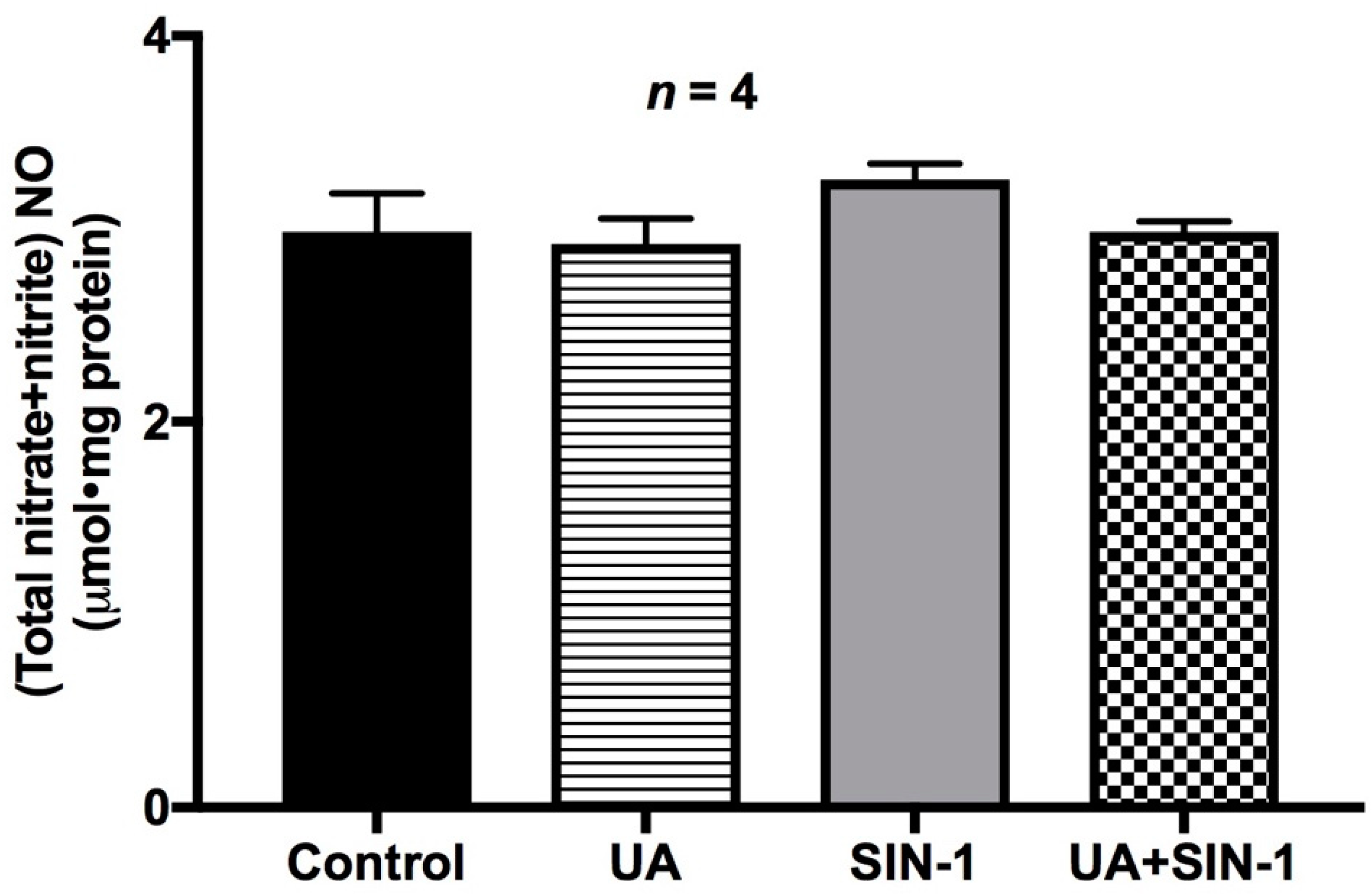
2.10. Nitric Oxide Levels in SIN-1-Treated Cells
3. Discussion
4. Materials and Methods
4.1. Tissue Culture and Drug Treatment
4.2. Na-Amino Acid Co-Transport in IEC-18 Cells
4.3. Na/K-ATPase Assay
4.4. Molecular Studies
4.5. Measurement of NO
4.6. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Binder, H.J.; Ptak, T. Jejunal absorption of water and electrolytes in inflammatory bowel disease. J. Lab. Clin. Med. 1970, 76, 915–924. [Google Scholar]
- Seidler, U.; Lenzen, H.; Cinar, A.; Tessema, T.; Bleich, A.; Riederer, B. Molecular mechanisms of disturbed electrolyte transport in intestinal inflammation. Ann. N. Y. Acad. Sci. 2006, 1072, 262–275. [Google Scholar] [CrossRef]
- Surawicz, C.M. Mechanisms of diarrhea. Curr. Gastroenterol. Rep. 2010, 12, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, U.; West, A.B. Effect of chronic inflammation on electrolyte transport in rabbit ileal villus and crypt cells. Am. J. Physiol. 1997, 272, G732–G741. [Google Scholar] [CrossRef]
- Field, M. Intestinal ion transport and the pathophysiology of diarrhea. J. Clin. Investig. 2003, 111, 931–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaram, U.; Wisel, S.; Rajendren, V.M.; West, A.B. Mechanism of inhibition of Na+-glucose cotransport in the chronically inflamed rabbit ileum. Am. J. Physiol. 1997, 273, G913–G919. [Google Scholar] [CrossRef] [PubMed]
- Saha, P.; Arthur, S.; Kekuda, R.; Sundaram, U. Na-glutamine co-transporters B(0)AT1 in villus and SN2 in crypts are differentially altered in chronically inflamed rabbit intestine. Biochim. Biophys. Acta 2012, 1818, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, U.; Wisel, S.; Coon, S. Neutral Na-amino acid cotransport is differentially regulated by glucocorticoids in the normal and chronically inflamed rabbit small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G467–G474. [Google Scholar] [CrossRef]
- Arthur, S.; Saha, P.; Sundaram, S.; Kekuda, R.; Sundaram, U. Regulation of sodium-glutamine cotransport in villus and crypt cells by glucocorticoids during chronic enteritis. Inflamm. Bowel Dis. 2012, 18, 2149–2157. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, S.S.; Mastropaolo, L.A.; Murchie, R.; Griffiths, C.; Thoni, C.; Elkadri, A.; Xu, W.; Mack, A.; Walters, T.; Guo, C.; et al. Higher activity of the inducible nitric oxide synthase contributes to very early onset inflammatory bowel disease. Clin. Transl. Gastroenterol. 2014, 5, e46. [Google Scholar] [CrossRef]
- Banan, A.; Fields, J.Z.; Zhang, Y.; Keshavarzian, A. iNOS upregulation mediates oxidant-induced disruption of F-actin and barrier of intestinal monolayers. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G1234–G1246. [Google Scholar] [CrossRef]
- Beckman, J.S.; Koppenol, W.H. Nitric oxide, superoxide, and peroxynitrite: The good, the bad, and ugly. Am. J. Physiol. 1996, 271, C1424–C1437. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, D.M. Peroxynitrite and inflammatory bowel disease. Gut 2000, 46, 436–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckman, J.S.; Beckman, T.W.; Chen, J.; Marshall, P.A.; Freeman, B.A. Apparent hydroxyl radical production by peroxynitrite: Implications for endothelial injury from nitric oxide and superoxide. Proc. Natl. Acad. Sci. USA 1990, 87, 1620–1624. [Google Scholar] [CrossRef]
- Rachmilewitz, D.; Stamler, J.S.; Bachwich, D.; Karmeli, F.; Ackerman, Z.; Podolsky, D.K. Enhanced colonic nitric oxide generation and nitric oxide synthase activity in ulcerative colitis and Crohn’s disease. Gut 1995, 36, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Crow, J.P. Dichlorodihydrofluorescein and dihydrorhodamine 123 are sensitive indicators of peroxynitrite in vitro: Implications for intracellular measurement of reactive nitrogen and oxygen species. Nitric Oxide Biol. Chem. 1997, 1, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Martin-Romero, F.J.; Gutierrez-Martin, Y.; Henao, F.; Gutierrez-Merino, C. Fluorescence measurements of steady state peroxynitrite production upon SIN-1 decomposition: NADH versus dihydrodichlorofluorescein and dihydrorhodamine 123. J. Fluoresc. 2004, 14, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, G.; Moshage, H.; van Dullemen, H.M.; de Jager-Krikken, A.; Tiebosch, A.T.; Kleibeuker, J.H.; Jansen, P.L.; van Goor, H. Expression of nitric oxide synthases and formation of nitrotyrosine and reactive oxygen species in inflammatory bowel disease. J. Pathol. 1998, 186, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H.; Hokari, R.; Miura, S.; Shigematsu, T.; Hirokawa, M.; Akiba, Y.; Kurose, I.; Higuchi, H.; Fujimori, H.; Tsuzuki, Y.; et al. Increased expression of an inducible isoform of nitric oxide synthase and the formation of peroxynitrite in colonic mucosa of patients with active ulcerative colitis. Gut 1998, 42, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.J.; Thompson, J.H.; Zhang, X.J.; Sadowska-Krowicka, H.; Kakkis, J.L.; Munshi, U.K.; Sandoval, M.; Rossi, J.L.; Eloby-Childress, S.; Beckman, J.S.; et al. Role of inducible nitric oxide synthase expression and peroxynitrite formation in guinea pig ileitis. Gastroenterology 1995, 109, 1475–1483. [Google Scholar] [CrossRef]
- Nishikawa, M.; Kanamori, M.; Hidaka, H. Inhibition of platelet aggregation and stimulation of guanylate cyclase by an antianginal agent molsidomine and its metabolites. J. Pharmacol. Exp. Ther. 1982, 220, 183–190. [Google Scholar]
- Arthur, S.; Sundaram, U. Inducible nitric oxide regulates intestinal glutamine assimilation during chronic intestinal inflammation. Nitric Oxide Biol. Chem. 2015, 44, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Coon, S.; Kim, J.; Shao, G.; Sundaram, U. Na-glucose and Na-neutral amino acid cotransport are uniquely regulated by constitutive nitric oxide in rabbit small intestinal villus cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G1030–G1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coon, S.; Kekuda, R.; Saha, P.; Sundaram, U. Reciprocal regulation of the primary sodium absorptive pathways in rat intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2011, 300, C496–C505. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, P.; Sundaram, S.; Singh, S.; Sundaram, U. Inducible Nitric Oxide Regulates Brush Border Membrane Na-Glucose Co-transport, but Not Na:H Exchange via p38 MAP Kinase in Intestinal Epithelial Cells. Cells 2018, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Singer, I.I.; Kawka, D.W.; Scott, S.; Weidner, J.R.; Mumford, R.A.; Riehl, T.E.; Stenson, W.F. Expression of inducible nitric oxide synthase and nitrotyrosine in colonic epithelium in inflammatory bowel disease. Gastroenterology 1996, 111, 871–885. [Google Scholar] [CrossRef]
- Zingarelli, B.; Szabo, C.; Salzman, A.L. Reduced oxidative and nitrosative damage in murine experimental colitis in the absence of inducible nitric oxide synthase. Gut 1999, 45, 199–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundaram, U.; Wisel, S.; Fromkes, J.J. Unique mechanism of inhibition of Na+-amino acid cotransport during chronic ileal inflammation. Am. J. Physiol. 1998, 275, G483–G489. [Google Scholar] [CrossRef]
- Becker, B.F. Towards the physiological function of uric acid. Free Radic. Biol. Med. 1993, 14, 615–631. [Google Scholar] [CrossRef]
- Hooper, D.C.; Spitsin, S.; Kean, R.B.; Champion, J.M.; Dickson, G.M.; Chaudhry, I.; Koprowski, H. Uric acid, a natural scavenger of peroxynitrite, in experimental allergic encephalomyelitis and multiple sclerosis. Proc. Natl. Acad. Sci. USA 1998, 95, 675–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, D.C.; Scott, G.S.; Zborek, A.; Mikheeva, T.; Kean, R.B.; Koprowski, H.; Spitsin, S.V. Uric acid, a peroxynitrite scavenger, inhibits CNS inflammation, blood-CNS barrier permeability changes, and tissue damage in a mouse model of multiple sclerosis. FASEB J. 2000, 14, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Skinner, K.A.; White, C.R.; Patel, R.; Tan, S.; Barnes, S.; Kirk, M.; Darley-Usmar, V.; Parks, D.A. Nitrosation of uric acid by peroxynitrite. Formation of a vasoactive nitric oxide donor. J. Biol. Chem. 1998, 273, 24491–24497. [Google Scholar] [CrossRef]
- Moreno, J.J.; Pryor, W.A. Inactivation of alpha 1-proteinase inhibitor by peroxynitrite. Chem. Res. Toxicol. 1992, 5, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Rubbo, H.; Radi, R.; Trujillo, M.; Telleri, R.; Kalyanaraman, B.; Barnes, S.; Kirk, M.; Freeman, B.A. Nitric oxide regulation of superoxide and peroxynitrite-dependent lipid peroxidation. Formation of novel nitrogen-containing oxidized lipid derivatives. J. Biol. Chem. 1994, 269, 26066–26075. [Google Scholar] [PubMed]
- Schopfer, F.J.; Baker, P.R.; Freeman, B.A. NO-dependent protein nitration: A cell signaling event or an oxidative inflammatory response? Trends Biochem. Sci. 2003, 28, 646–654. [Google Scholar] [CrossRef]
- Forbush, B., 3rd. Assay of Na,K-ATPase in plasma membrane preparations: Increasing the permeability of membrane vesicles using sodium dodecyl sulfate buffered with bovine serum albumin. Anal. Biochem. 1983, 128, 159–163. [Google Scholar] [CrossRef]
- Knickelbein, R.; Aronson, P.S.; Schron, C.M.; Seifter, J.; Dobbins, J.W. Sodium and chloride transport across rabbit ileal brush border. II. Evidence for Cl-HCO3 exchange and mechanism of coupling. Am. J. Physiol. 1985, 249, G236–G245. [Google Scholar] [CrossRef]
- Palaniappan, B.; Sundaram, U. Direct and specific inhibition of constitutive nitric oxide synthase uniquely regulates brush border membrane Na-absorptive pathways in intestinal epithelial cells. Nitric Oxide Biol. Chem. 2018, 79, 8–13. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vmax (µmol/mg Protein·30 s) | Km (mM) | |
|---|---|---|
| Control | 4.5 ± 0.3 | 2.5 ± 0.3 |
| SIN-1 | 5.4 ± 0.8 | 13.3 ± 0.5 * |
| Vmax (pmol/mg Protein·30 s) | Km (mM) | |
|---|---|---|
| Control | 1.48 ± 0.06 | 0.18 ± 0.01 |
| SIN-1 | 0.76 ± 0.01 * | 0.17 ± 0.004 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arthur, S.; Manoharan, P.; Sundaram, S.; Rahman, M.M.; Palaniappan, B.; Sundaram, U. Unique Regulation of Enterocyte Brush Border Membrane Na-Glutamine and Na-Alanine Co-Transport by Peroxynitrite during Chronic Intestinal Inflammation. Int. J. Mol. Sci. 2019, 20, 1504. https://doi.org/10.3390/ijms20061504
Arthur S, Manoharan P, Sundaram S, Rahman MM, Palaniappan B, Sundaram U. Unique Regulation of Enterocyte Brush Border Membrane Na-Glutamine and Na-Alanine Co-Transport by Peroxynitrite during Chronic Intestinal Inflammation. International Journal of Molecular Sciences. 2019; 20(6):1504. https://doi.org/10.3390/ijms20061504
Chicago/Turabian StyleArthur, Subha, Palanikumar Manoharan, Shanmuga Sundaram, M Motiur Rahman, Balasubramanian Palaniappan, and Uma Sundaram. 2019. "Unique Regulation of Enterocyte Brush Border Membrane Na-Glutamine and Na-Alanine Co-Transport by Peroxynitrite during Chronic Intestinal Inflammation" International Journal of Molecular Sciences 20, no. 6: 1504. https://doi.org/10.3390/ijms20061504