Transcriptomic Analysis of Porcine Endometrium during Implantation after In Vitro Stimulation by Adiponectin

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Microarray Data Analysis
2.1.1. Identification of Differentially Expressed (DE) Genes
2.1.2. Gene Ontology Analysis
2.1.3. Biological Pathway Analysis
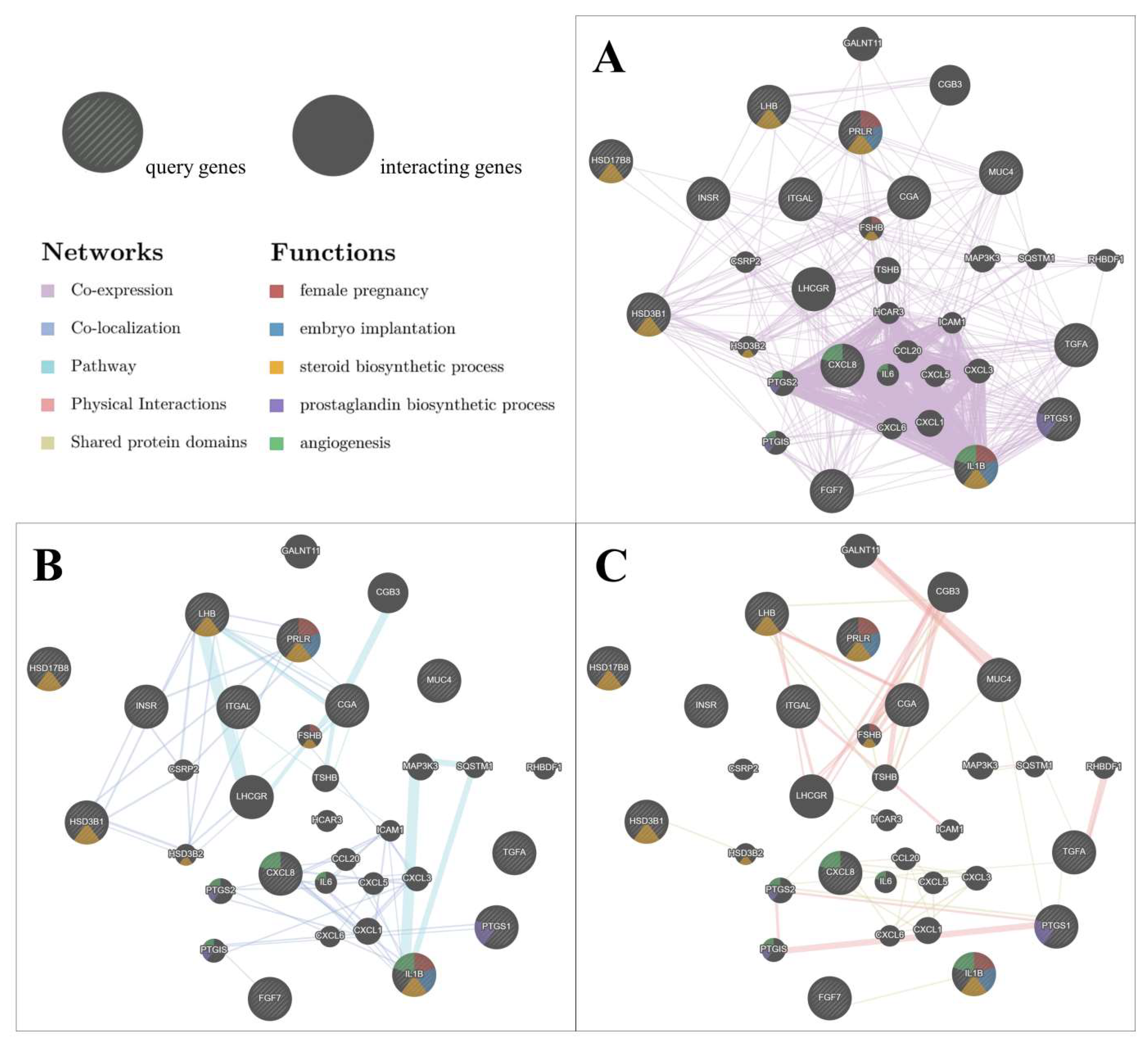
2.1.4. Network between Differentially Expressed Genes
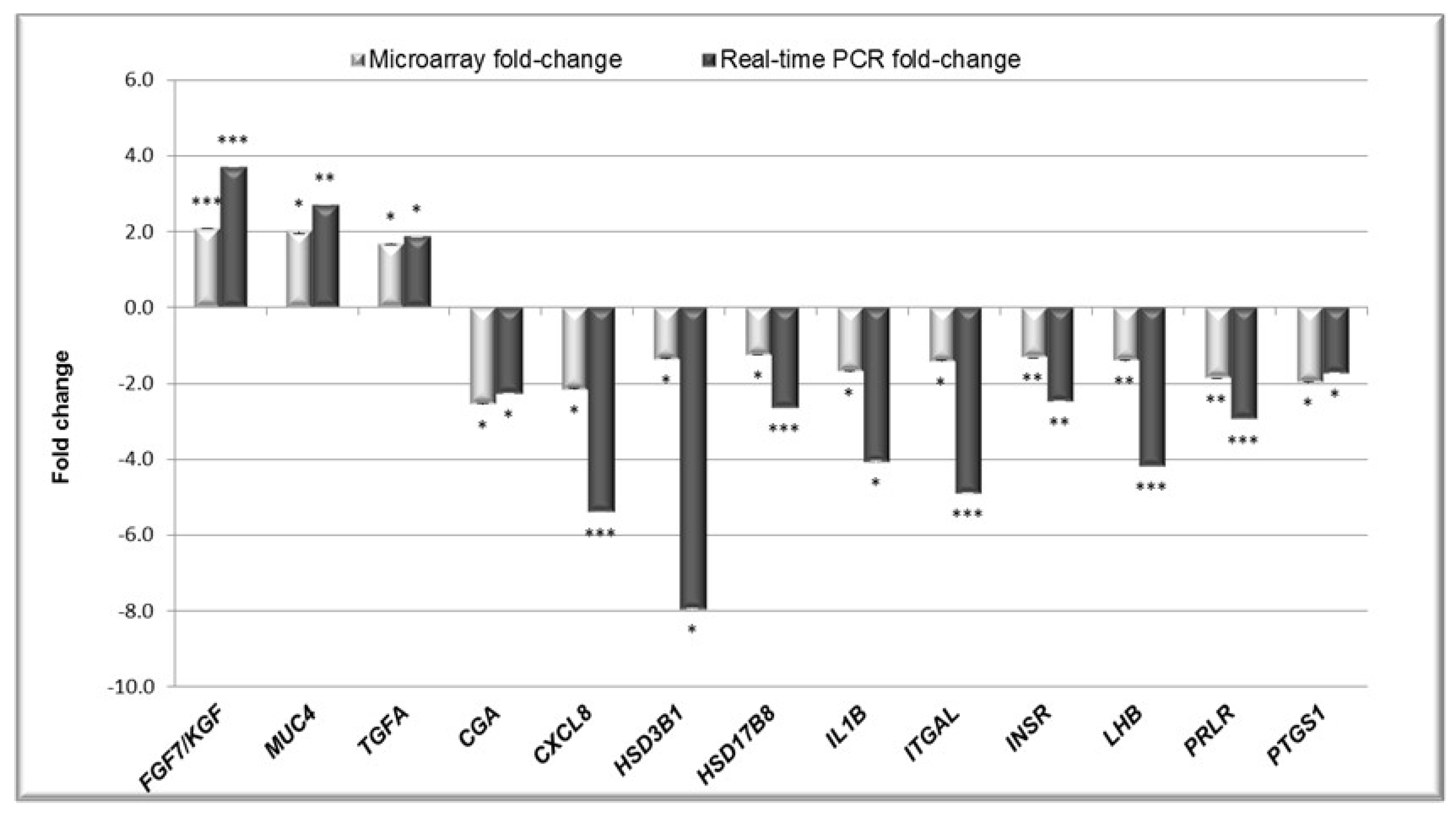
2.2. Validation of the Microarray Results by Real-Time PCR (qPCR).
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Tissue Collection
4.2. Endometrial Explant Culture
4.3. Total RNA Isolation and Quality Control
4.4. Microarray Hybridization
4.5. Bioinformatic Analysis
4.5.1. Gene Ontology Analysis
4.5.2. Biological Pathways Analysis
4.5.3. Interaction Network of Differentially Expressed Genes
4.6. Real-Time PCR Validations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Geisert, R.D.; Schmitt, R.A.M. Early embryonic survival in the pig: Can it be improved? J. Anim. Sci. 2002, 80, E54–E65. [Google Scholar] [CrossRef]
- Bazer, F.W.; Johnson, G.A. Pig blastocyst-uterine interactions. Differentiation 2014, 87, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Viganò, P.; Mangioni, S.; Pompei, F.; Chiodo, I. Maternal-conceptus cross talk-a review. Placenta 2003, 24, S56–S61. [Google Scholar] [CrossRef]
- Ziecik, A.J.; Waclawik, A.; Kaczmarek, M.M.; Blitek, A.; Jalali, B.M.; Andronowska, A. Mechanisms for the establishment of pregnancy in the pig. Reprod. Domest. Anim. 2011, 46 (Suppl. 3), 31–41. [Google Scholar] [CrossRef] [PubMed]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 1995, 270, 11259–11263. [Google Scholar] [CrossRef]
- Maeda, K.; Okubo, K.; Shimomura, I.; Funahashi, T.; Matsuzawa, Y.; Matsubara, K. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem. Biophys. Res. Commun. 1996, 221, 286–289. [Google Scholar] [CrossRef]
- Nakano, Y.; Tobe, T.; Choi-Miura, N.H.; Mazda, T.; Tomita, M. Isolation and characterization of GBP28, a novel gelatin-binding protein purified from human plasma. J. Biochem. 1996, 120, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Hu, E.; Liang, P.; Spiegelman, B.M. AdipoQ is a novel adipose-specific gene dysregulated in obesity. J. Biol. Chem. 1996, 271, 10697–10703. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T. Adiponectin and adiponectin receptors. Endocr. Rev. 2005, 26, 439–451. [Google Scholar] [CrossRef]
- Palin, M.F.; Bordignon, V.V.; Murphy, B.D. Adiponectin and the control of female reproductive functions. Vitam. Horm. 2012, 90, 239–287. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyn, K.; Smolinska, N.; Kiezun, M.; Szeszko, K.; Rytelewska, E.; Kisielewska, K.; Gudelska, M.; Kaminski, T. Adiponectin: A New Regulator of Female Reproductive System. Int. J. Endocrinol. 2018, 2018, 7965071. [Google Scholar] [CrossRef] [PubMed]
- Lord, E.; Ledoux, S.; Murphy, B.D.; Beaudry, D.; Palin, M.F. Expression of adiponectin and its receptors in swine. J. Anim. Sci. 2005, 83, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Takemura, Y.; Osuga, Y.; Yamauchi, T.; Kobayashi, M.; Harada, M.; Hirata, T.; Morimoto, C.; Hirota, Y.; Yoshino, O.; Koga, K.; et al. Expression of Adipoq receptors and its possible implication in the human endometrium. Endocrinology 2006, 147, 3203–3210. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.T.; Marquard, K.; Stephens, S.; Louden, E.; Allsworth, J.; Moley, K.H. Adipoq and Adipoq receptors in the mouse preimplantation embryo and uterus. Hum. Reprod. 2011, 26, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Chabrolle, C.; Tosca, L.; Dupont, J. Regulation of Adipoq and its receptors in rat ovary by human chorionic gonadotrophin treatment and potential involvement of Adipoq in granulosa cell steroidogenesis. Reproduction 2007, 133, 719–731. [Google Scholar] [CrossRef]
- Maleszka, A.; Smolinska, N.; Nitkiewicz, A.; Kiezun, M.; Dobrzyn, K.; Czerwinska, J.; Szeszko, K.; Kaminski, T. Expression of Adipoq receptors 1 and 2 in the ovary and concentration of plasma Adipoq during the oestrous cycle of the pig. Acta Vet. Hung. 2014, 62, 386–396. [Google Scholar] [CrossRef]
- Maleszka, A.; Smolinska, N.; Nitkiewicz, A.; Kiezun, M.; Chojnowska, K.; Dobrzyn, K.; Szwaczek, H.; Kaminski, T. Adipoq Expression in the Porcine Ovary during the Oestrous Cycle and Its Effect on Ovarian Steroidogenesis. Int. J. Endocrinol. 2014, 2014, 9570–9576. [Google Scholar] [CrossRef]
- Smolinska, N.; Maleszka, A.; Dobrzyn, K.; Kiezun, M.; Szeszko, K.; Kaminski, T. Expression of Adipoq and Adipoq receptors 1 and 2 in the porcine uterus, conceptus, and trophoblast during early pregnancy. Theriogenology 2014, 82, 951–965. [Google Scholar] [CrossRef]
- Smolinska, N.; Dobrzyn, K.; Maleszka, A.; Kiezun, M.; Szeszko, K.; Kaminski, T. Expression of Adipoq and Adipoq receptors 1 (AdipoR1) and 2 (AdipoR2) in the porcine uterus during the oestrous cycle. Anim. Reprod. Sci. 2014, 146, 42–54. [Google Scholar] [CrossRef]
- Smolinska, N.; Dobrzyn, K.; Kiezun, M.; Szeszko, K.; Maleszka, A.; Kaminski, T. Effect of adiponectin on the steroidogenic acute regulatory protein, P450 side chain cleavage enzyme and 3β-hydroxysteroid dehydrogenase genes expression, progesterone and androstenedione production by the porcine uterus during early pregnancy. J. Physiol. Pharmacol. 2016, 67, 443–456. [Google Scholar]
- Gamundi-Segura, S.; Serna, J.; Oehninger, S.; Horcajadas, J.A.; Arbones-Mainar, J.M. Effects of adipocyte-secreted factors on decidualized endometrial cells: Modulation of endometrial receptivity in vitro. J. Physiol. Biochem. 2015, 71, 537–546. [Google Scholar] [CrossRef]
- Smolinska, N.; Kiezun, M.; Dobrzyn, K.; Szeszko, K.; Maleszka, A.; Kaminski, T. Adiponectin, orexin A and orexin B concentrations in the serum and uterine luminal fluid during early pregnancy of pigs. Anim. Reprod. Sci. 2017, 178, 1–8. [Google Scholar] [CrossRef]
- Brochu-Gaudreau, K.; Beaudry, D.; Blouin, R.; Bordignon, V.; Murphy, B.D.; Palin, M.F. Adiponectin Regulates Gene Expression in the Porcine Uterus. Biol. Reprod. 2008, 78, 210–211. [Google Scholar] [CrossRef]
- Demetrius, L. Of mice and men. When it comes to studying ageing and the means to slow it down, mice are not just small humans. EMBO Rep. 2005, 6, S39–S44. [Google Scholar] [CrossRef]
- Vinogradov, A.E.; Anatskaya, O.V. Gene Golden Age paradox and its partial solution. Genomics 2018. [Google Scholar] [CrossRef]
- Geisert, R.; Fazleabas, A.; Lucy, M.; Mathew, D. Interaction of the conceptus and endometrium to establish pregnancy in mammals: Role of interleukin 1β. Cell Tissue Res. 2012, 349, 825–838. [Google Scholar] [CrossRef]
- Tuo, W.; Harney, J.P.; Bazer, F.W. Developmentally regulated expression of interleukin-1β by periimplantation conceptuses in swine. J. Reprod. Immunol. 1996, 31, 185–198. [Google Scholar] [CrossRef]
- Ross, J.W.; Malayer, J.R.; Ritchey, J.W.; Geisert, R.D. Characterization of the interleukin-1beta system during porcine trophoblastic elongation and early placental attachment. Biol. Reprod. 2003, 69, 1251–1259. [Google Scholar] [CrossRef]
- Bohlouli, S.; Khazaei, M.; Teshfam, M.; Hassanpour, H. Adiponectin effect on the viability of human endometrial stromal cells and mRNA expression of adiponectin receptors. Int. J. Fertil. Steril. 2013, 7, 43–48. [Google Scholar]
- Bohlouli, S.; Rabzia, A.; Sadeghi, E.; Chobsaz, F.; Khazaei, M. In vitro Anti-Proliferative Effect of Adiponectin on Human Endometriotic Stromal Cells through AdipoR1 and AdipoR2 Gene Receptor Expression. Iran. Biomed. J. 2016, 20, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Bråkenhielm, E.; Veitonmäki, N.; Cao, R.; Kihara, S.; Matsuzawa, Y.; Zhivotovsky, B.; Funahashi, T.; Cao, Y. Adiponectin-induced antiangiogenesis and antitumor activity involve caspase-mediated endothelial cell apoptosis. Proc. Natl. Acad. Sci. USA 2004, 101, 2476–2481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, K.W.; Shin, J.H.; Park, H.T.; Kim, T.; Kim, S.H.; Hur, J.Y. Resistin concentration is increased in the peritoneal fluid of women with endometriosis. Am. J. Reprod. Immunol. 2010, 64, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Soliman, P.T.; Wu, D.; Tortolero-Luna, G.; Schmeler, K.M.; Slomovitz, B.M.; Bray, M.S.; Gershenson, D.M.; Lu, K.H. Association between adiponectin, insulin resistance, and endometrial cancer. Cancer 2006, 106, 2376–2381. [Google Scholar] [CrossRef] [Green Version]
- Dal Maso, L.; Augustin, L.S.; Karalis, A.; Talamini, R.; Franceschi, S.; Trichopoulos, D.; Mantzoros, C.S.; La Vecchia, C. Circulating adiponectin and endometrial cancer risk. J. Clin. Endocrinol. Metab. 2004, 89, 1160–1163. [Google Scholar] [CrossRef]
- Cust, A.E.; Kaaks, R.; Friedenreich, C.; Bonnet, F.; Laville, M.; Lukanova, A.; Rinaldi, S.; Dossus, L.; Slimani, N.; Lundin, E.; et al. Plasma adiponectin levels and endometrial cancer risk in pre- and postmenopausal women. J. Clin. Endocrinol. Metab. 2007, 92, 255–263. [Google Scholar] [CrossRef]
- Cong, L.; Gasser, J.; Zhao, J.; Yang, B.; Li, F.; Zhao, A.Z. Human adiponectin inhibits cell growth and induces apoptosis in human endometrial carcinoma cells, HEC-1-A and RL95 2. Endocr. Relat. Cancer 2007, 14, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.; Choi, M.J.; Bae, H.; Bazer, F.W.; Song, G. A critical role for adiponectin-mediated development of endometrial luminal epithelial cells during the peri-implantation period of pregnancy. J. Cell. Physiol. 2017, 232, 3146–3157. [Google Scholar] [CrossRef]
- Audie, J.P.; Tetaert, D.; Pigny, P.; Buisine, M.P.; Janin, A.; Aubert, J.P.; Porchet, N.; Boersma, A. Mucin gene expression in the human endocervix. Hum. Reprod. 1995, 10, 98–102. [Google Scholar] [CrossRef]
- Gipson, I.K.; Ho, S.B.; Spurr-Michaud, S.J.; Tisdale, A.S.; Zhan, Q.; Torlakovic, E.; Pudney, J.; Anderson, D.J.; Toribara, N.W.; Hill, J.A., 3rd. Mucin genes expressed by human female reproductive tract epithelia. Biol. Reprod. 1997, 56, 999–1011. [Google Scholar] [CrossRef] [Green Version]
- Carraway, K.L.; Price-Schiavi, S.A.; Komatsu, M.; Idris, N.; Perez, A.; Li, P.; Jepson, S.; Zhu, X.; Carvajal, M.E.; Carraway, C.A. Multiple facets of sialomucin complex/MUC4, a membrane mucin and erbb2 ligand, in tumors and tissues (Y2K update). Front. Biosci. 2000, 5, D95–D107. [Google Scholar]
- Idris, N.; Carraway, K.L. Sialomucin complex (Muc4) expression in the rat female reproductive tract. Biol. Reprod. 1999, 61, 1431–1438. [Google Scholar] [CrossRef]
- McNeer, R.R.; Carraway, C.A.; Fregien, N.L.; Carraway, K.L. Characterization of the expression and steroid hormone control of sialomucin complex in the rat uterus: Implications for uterine receptivity. J. Cell. Physiol. 1998, 176, 110–119. [Google Scholar] [CrossRef]
- Ferrell, A.D.; Malayer, J.R.; Carraway, K.L.; Geisert, R.D. Sialomucin complex (Muc4) expression in porcine endometrium during the oestrous cycle and early pregnancy. Reprod. Domest. Anim. 2003, 38, 63–65. [Google Scholar] [CrossRef]
- Dobrzyn, K.; Smolinska, N.; Szeszko, K.; Kiezun, M.; Maleszka, A.; Rytelewska, E.; Kaminski, T. Effect of progesterone on adiponectin system in the porcine uterus during early pregnancy. J. Anim. Sci. 2017, 95, 338–352. [Google Scholar] [CrossRef]
- Dobrzyn, K.; Smolinska, N.; Kiezun, M.; Szeszko, K.; Maleszka, A.; Kaminski, T. The effect of estrone and estradiol on the expression of the adiponectin system in the porcine uterus during early pregnancy. Theriogenology 2017, 88, 183–196. [Google Scholar] [CrossRef]
- Villar, J.; Celay, J.; Alonso, M.M.; Rotinen, M.; de Miguel, C.; Migliaccio, M.; Encío, I. Transcriptional regulation of the human type 8 17beta-hydroxysteroid dehydrogenase gene by C/EBPbeta. J. Steroid. Biochem. Mol. Biol. 2007, 105, 131–139. [Google Scholar] [CrossRef]
- Bazer, F.W.; Roberts, R.M. Biochemical aspects of conceptus—Endometrial interactions. J. Exp. Zool. 1983, 228, 373–383. [Google Scholar] [CrossRef]
- Bowen, J.A.; Burghardt, R.C. Cellular mechanisms of implantation in domestic farm animals. Semin. Cell Dev. Biol. 2000, 11, 93–104. [Google Scholar] [CrossRef]
- Hynes, R.O. Integrins: Versatility, modulation, and signaling in cell adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Burghardt, R.C.; Johnson, G.A.; Jaeger, L.A.; Ka, H.; Garlow, J.E.; Spencer, T.E.; Bazer, F.W. Integrins and extracellular matrix proteins at the maternal-fetal interface in domestic animals. Cells Tissues Organs 2002, 172, 202–217. [Google Scholar] [CrossRef]
- Lessey, B.A. Endometrial integrins and the establishment of uterine receptivity. Hum. Reprod. 1998, 13 (Suppl. 3), 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledoux, S.; Campos, D.B.; Lopes, F.L.; Dobias-Goff, M.; Palin, M.F.; Murphy, B.D. Adiponectin induces periovulatory changes in ovarian follicular cells. Endocrinology 2006, 147, 5178–5186. [Google Scholar] [CrossRef] [PubMed]
- Szeszko, K.; Smolinska, N.; Kiezun, M.; Dobrzyn, K.; Maleszka, A.; Kaminski, T. The influence of adiponectin on the transcriptomic profile of porcine luteal cells. Funct. Integr. Genom. 2016, 16, 101–114. [Google Scholar] [CrossRef]
- Dobrzyn, K.; Szeszko, K.; Kiezun, M.; Kisielewska, K.; Rytelewska, E.; Gudelska, M.; Wyrebek, J.; Bors, K.; Kaminski, T.; Smolinska, N. In vitro effect of orexin A on the transcriptomic profile of the endometrium during early pregnancy in pigs. Anim. Reprod. Sci. 2019, 200, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Pan, J.; Ma, J.; Zhang, Z.; Bai, Y. Microarray gene expression of periosteum in spontaneous bone regeneration of mandibular segmental defects. Sci. Rep. 2017, 7, 13535. [Google Scholar] [CrossRef] [PubMed]
- Zglejc, K.; Martyniak, M.; Waszkiewicz, E.; Kotwica, G.; Franczak, A. Peri-conceptional under-nutrition alters transcriptomic profile in the endometrium during the peri-implantation period-The study in domestic pigs. Reprod. Domest. Animal. 2018, 53, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef]
- Quilter, C.R.; Gilbert, C.L.; Oliver, G.L.; Jafer, O.; Furlong, R.A.; Blott, S.C.; Wilson, A.E.; Sargent, C.A.; Mileham, A.; Affara, N.A. Gene expression profiling in porcine maternal infanticide: A model for puerperal psychosis. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2008, 47, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, T.G.; Caperna, T.J. Ontogeny of adipokine expression in neonatal pig adipose tissue. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 152, 72–78. [Google Scholar] [CrossRef]
- Wojciechowicz, B.; Kotwica, G.; Kolakowska, J.; Franczak, A. The Activity and Localization of 3β-hydroxysteroid Dehydrogenase/Δ5-Δ4 Isomerase and Release of Androstenedione and Progesterone by Uterine Tissues During Early Pregnancy and the Estrous Cycle in Pigs. J. Reprod. Dev. 2013, 59, 49–58. [Google Scholar] [CrossRef]
- Liu, G.M.; Wei, Y.; Wang, Z.S.; Wu, D.; Zhou, A.G.; Liu, G.L. Effects of herbal extract supplementation on growth performance and insulin-like growth factor (IGF)-I system in finishing pigs. J. Anim. Feed Sci. 2008, 17, 538–547. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.G.; O’Doherty, J.V.; Reilly, P.; Ryan, M.T.; Bahar, B.; Sweeney, T. The effects of laminarin derived from Laminaria digitata on measurements of gut health: Selected bacterial populations, intestinal fermentation, mucin gene expression and cytokine gene expression in the pig. Br. J. Nutr. 2011, 105, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Choi, Y.; Shim, J.; Choi, Y.; Ka, H. Regulatory mechanism for expression of IL1B receptors in the uterine endometrium and effects of IL1B on prostaglandin synthetic enzymes during the implantation period in pigs. Biol. Reprod. 2012, 87, 31. [Google Scholar] [CrossRef] [PubMed]
- Farmer, C.; Palin, M.F. Feeding flaxseed to sows during late-gestation and lactation affects mammary development but not mammary expression of selected genes in their offspring. Can. J. Anim. Sci. 2013, 93, 17. [Google Scholar] [CrossRef]
- Spagnuolo-Weaver, M.; Fuerst, R.; Campbell, S.T.; Meehan, B.M.; Mcneilly, F.; Adair, B.; Allan, G. A fluorimeter-Based RT-PCR method for the detection and quantitation of porcine cytokines. J. Immunol. Methods 1999, 230, 19–27. [Google Scholar] [CrossRef]
- Nitkiewicz, A.; Smolinska, N.; Przala, J.; Kaminski, T. Expression of orexin receptors 1 (OX1R) and 2 (OX2R) in the porcine ovary during the oestrous cycle. Regul. Pept. 2010, 165, 186–190. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| KEGG Pathway Analysis | |||
|---|---|---|---|
| Analysis Name | Gene Number | p-Value | Altered Genes |
| Pathways in cancer | 21 | 1.00 × 10−1 | CEBPA, STAT5A, CBL, CXCL8, NFKBIA, GNG12, APPL1, CBLB, CDKN1A, EP300, GNAQ, CXCR4, NCOA4, TGFA, PIK3R5, LAMC2, RARB, NOS2, CRK, CHUK, TRAF3 |
| Cytokine–cytokine receptor interaction | 18 | 3.60 × 10−3 | CSF3, IL9, CXCL8, CCL28, TNFRSF4, CCL4, IFNAR1, TNFRSF1B, TNFRSF11B, PRLR, TNFSF13B, IL10RB, CXCR4, IL4R, IL13RA1, IFNGR2, BMPR1A |
| Jak–STAT signalling pathway | 16 | 1.60 × 10−4 | CSF3, STAT5A, SOCS1, IL9, SOCS4, IL24, IFNAR1, EP300, PRLR, IL10RB, IL4R, PIK3R5, JAK3, IL13RA1, IFNGR2 |
| Regulation of actin cytoskeleton | 15 | 1.40 × 10−2 | ITGAL, SSH1, DIAPH1, SSH2, ARPC5, GNG12, PPP1CC, NCKAP1, ARPC1B, ITGB7, PIK3R5, MSN, PIP4K2A, CRK, MYLK |
| HTLV-I infection | 15 | 8.30 × 10−2 | ITGAL, TLN1, KAT2B, STAT5A, NFKBIA, MYBL2, ATM, SLA-8, CDKN1A, EP300, ETS1, PIK3R5, JAK3, NFATC3, CHUK |
| Transcriptional misregulation in cancer | 14 | 3.80 × 10−3 | CEBPA, CEBPB, KMT2A, CEBPE, LDB1, CCNT1, ELANE, CXCL8, ATM, HHEX, CDKN1A, ITGB7, GOLPH3L |
| Herpes simplex infection | 14 | 1.60 × 10−2 | CSNK2A2, CFP, EP300, HNRNPK, TAF5, EIF2S1, NFKBIA, HCFC2, PPP1CC, IFNGR2, CHUK, IFNAR1, SLA-8, TRAF3 |
| Viral carcinogenesis | 14 | 1.90 × 10−2 | LOC100156127, KAT2B, UBE3A, STAT5A, NFKBIA, PMAIP1, SLA-8, CDKN1A, HNRNPK, EP300, PIK3R5, JAK3, LOC100621389, TRAF3 |
| Insulin signalling pathway | 13 | 2.40 × 10−3 | SOCS1, CBL, ACACA, FBP1, RPS6KB1, SOCS4, PPP1CC, CBLB, PYGM, PYGL, PIK3R5, CRK, INSR |
| Chemokine signalling pathway | 13 | 2.30 × 10−2 | AMCF-II, CXCR4, CXCL2, CXCL8, NFKBIA, PIK3R5, FOXO3, GNG12, JAK3, CCL4, CCL28, CRK, CHUK |
| Epstein–Barr virus infection | 12 | 7.90 × 10−2 | CSNK2A2, ITGAL, CDKN1A, EP300, IL10RB, NFKBIA, PIK3R5, JAK3, TNFAIP3, CHUK, SLA-8, TRAF3 |
| Measles | 11 | 1.80 × 10−2 | CSNK2A2, EIF2S1, STAT5A, NFKBIA, PIK3R5, JAK3, MSN, TNFAIP3, IFNGR2, CHUK, IFNAR1 |
| Ubiquitin-mediated proteolysis | 11 | 2.00 × 10−2 | CUL5, CBLB, UBE3A, UBR5, WWP1, SOCS1, CBL, RHOBTB1, KEAP1, LOC780419, TRIP12 |
| Prolactin signalling pathway | 10 | 2.90 × 10−4 | CGA, PRLR, STAT5A, SOCS1, IRF1, SOCS4, PIK3R5, FOXO3, LHB, CSN2 |
| Salmonella infection | 10 | 1.50 × 10−3 | ARPC1B, CXCL2, PKN2, CXCL8, ARPC5, NOS2, DYNC1H1, CASP1, CCL4, IFNGR2 |
| NF-kappa B signalling pathway | 10 | 3.10 × 10−3 | CSNK2A2, TNFSF13B, LY96, CXCL8, NFKBIA, TNFAIP3, CCL4, ATM, CHUK, TRAF3 |
| TNF signalling pathway | 10 | 1.20 × 10−2 | LOC100736836, TNFRSF1B, CEBPB, CXCL2, NFKBIA, PIK3R5, TNFAIP3, CHUK, TRAF3 |
| Hepatitis C | 10 | 3.30 × 10−2 | CDKN1A, EIF2S1, IRF1, CXCL8, NFKBIA, PIK3R5, CHUK, IFNAR1, PPP2R2A, TRAF3 |
| Osteoclast differentiation | 10 | 4.30 × 10−2 | CYLD, TNFRSF11B, CTSK, SOCS1, NFKBIA, PIK3R5, IFNGR2, SIRPA, CHUK, IFNAR1 |
| ErbB signalling pathway | 9 | 7.20 × 10−3 | CBLB, CDKN1A, STAT5A, CBL, TGFA, PIK3R5, RPS6KB1, ABL2, CRK |
| Toll-like receptor signalling pathway | 9 | 2.10 × 10−2 | CTSK, LY96, CXCL8, NFKBIA, PIK3R5, CCL4, CHUK, IFNAR1, TRAF3 |
| Chagas disease (American trypanosomiasis) | 9 | 2.90 × 10−2 | GNAQ, CD247, CXCL8, NFKBIA, PIK3R5, NOS2, IFNGR2, CHUK, PPP2R2A |
| Insulin resistance | 9 | 3.80 × 10−2 | RPS6KA3, PYGM, PYGL, NFKBIA, PIK3R5, RPS6KB1, PPP1CC, SLC27A2, INSR |
| Toxoplasmosis | 9 | 4.10 × 10−2 | LY96, IL10RB, SOCS1, NFKBIA, LAMC2, PIK3R5, NOS2, IFNGR2, CHUK |
| AMPK signalling pathway | 9 | 4.70 × 10−2 | EEF2K, ACACA, FBP1, RAB14, PIK3R5, RPS6KB1, FOXO3, INSR, PPP2R2A |
| Chronic myeloid leukemia | 8 | 6.50 × 10−3 | CBLB, CDKN1A, STAT5A, CBL, NFKBIA, PIK3R5, CRK, CHUK |
| Ribosome biogenesis in eukaryotes | 8 | 1.80 × 10−2 | CSNK2A2, RN18S, MPHOSPH10, NOP58, WDR3, NOP56, BMS1, GNL3 |
| HIF-1 signalling pathway | 8 | 5.20 × 10−2 | PDK1, CDKN1A, EP300, PIK3R5, RPS6KB1, NOS2, IFNGR2, INSR |
| Bacterial invasion of epithelial cells | 7 | 3.40 × 10−2 | ARPC1B, CBLB, CBL, PIK3R5, ARPC5, CRK, ARHGAP10 |
| Peroxisome | 7 | 4.80 × 10−2 | ACSL1, NUDT12, NOS2, SLC27A2, ACSL3, CROT, SOD2 |
| Small cell lung cancer | 7 | 5.90 × 10−2 | NFKBIA, LAMC2, PIK3R5, NOS2, RARB, CHUK, TRAF3 |
| Phosphatidylinositol signalling system | 7 | 9.40 × 10−2 | MTMR14, PI4KA, PIK3R5, DGKH, INPP4A, PIP4K2A, INPP5B |
| Systemic lupus erythematosus | 7 | 9.80 × 10−2 | LOC100156127, LOC100157763, LOC100158121, ELANE, LOC100154071, LOC100153329, LOC100621389 |
| Pertussis | 6 | 7.50 × 10−2 | AMCF-II, LY96, IRF1, CXCL8, NOS2, CASP1 |
| NOD-like receptor signalling pathway | 5 | 5.10 × 10−2 | CXCL8, NFKBIA, TNFAIP3, CASP1, CHUK |
| Ovarian steroidogenesis | 5 | 5.40 × 10−2 | CGA, PLA2G4A, HSD3B1, LHB, INSR |
| Malaria | 5 | 5.80 × 10−2 | CSF3, ITGAL, SELP, CXCL8 |
| Acute myeloid leukemia | 5 | 6.70 × 10−2 | CEBPA, STAT5A, PIK3R5, RPS6KB1, CHUK |
| Pentose phosphate pathway | 4 | 1.10 × 10−2 | FBP1, TKT, RPIA, PRPS2 |
| Intestinal immune network for IgA production | 4 | 9.50 × 10−2 | TNFSF13B, CXCR4, ITGB7, CCL28 |
| Fatty acid biosynthesis | 3 | 1.40 × 10−2 | ACSL1, ACACA, ACSL3 |
| Glycosaminoglycan biosynthesis-chondroitin sulphate/dermatan sulphate | 3 | 5.10 × 10−2 | CSGALNACT2, CHST3, CHSY1 |
| Nicotinate and nicotinamide metabolism | 3 | 9.40 × 10−2 | ENPP1, NUDT12, NMRK1 |
| Dorso-ventral axis formation | 3 | 9.40 × 10−2 | CPEB2, ETS1, CPEB4 |
| Gene Symbol | Primers Sequences | Reaction Conditions | Primer (nM) | Target Sequence Accession Number | References | |
|---|---|---|---|---|---|---|
| CGA | F: 5′-CTCCAGAGCGTACCCAACTC-3′ R: 5′-ACTGTGGCCTTGGTAAATGC-3′ | Activation: 50 °C, 30 min; | 40 cycles | 500 nM | XM_005659277.1 | [60] |
| 95 °C 15 min, | ||||||
| 1. Denaturation: 94 °C, 15 s | ||||||
| 2. Annealing: 55 °C, 30 s | ||||||
| 3. Extension: 72 °C, 30 s | ||||||
| 77 °C, 15 s | ||||||
| CXCL8 | F: 5′-GGCAGTTTTCCTGCTTTCT-3′ R: 5′-CAGTGGGGTCCACTCTCAAT-3′ | Activation: 95 °C, 15 min | 40 cycles | 400 nM | X61151.1 | [61] |
| 1. Denaturation: 94 °C, 15 s | ||||||
| 2. Annealing: 58 °C, 30 s | ||||||
| 3. Extension: 72 °C, 30 s | ||||||
| FGF7/KGF | F: 5′-GCTTCCACATTATCTGTCTGGTG-3′ R: 5′-GTCCCTTTGACTTTGCCTCG-3′ | Activation: 95 °C, 10 min | 40 cycles | 500 nM | AF217463.1 | This study |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 60 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| HSD3B1 | F: 5′-AGGTTCGCCCGCTCATC-3′ R: 5′-CTGGGCACCGAGAAATACTTG-3′ | Activation: 95 °C, 10 min | 40 cycles | 300 nM | NM_001004049.1 | [62] |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 61 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| HSD17B8 | F: 5′-TTCTGCTCCGCATGTCTGAAG-3′ R: 5′-CCATGTTTCCCACCTTCCCTA-3′ | Activation: 95 °C, 10 min | 40 cycles | 500 nM | NM_001130730.1 | This study |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 60 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| IL1B | F: 5′-TGCCAACGTGCAGTCTATGG-3′ R: 5′-TGGGCCAGCCAGCACTAG-3′ | Activation: 95 °C, 10 min | 40 cycles | 100 nM | NM_214055 | [29] |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 60 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| INSR | F: 5′-AAACGCCAGGGACATCGTCAAGG-3′ R: 5′-CCGCAGGGAACGCAGGTAACTCT-3′ | Activation: 95 °C-10 min | 40 cycles | 200 nM | XM_005654749.1 | [63] |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 60 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| ITGAL | F: 5′-CTTGTCGAGCTGAAGGCTGA-3′ R: 5′-TTCCTGGTCCTTGGTGAGGA-3′ | Activation: 95 °C, 10 min | 40 cycles | 500 nM | NM_001044608.1 | This study |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 60 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| LHB | F: 5′-TTCACCACCAGCATCTGTGC-3′ R: 5′-AAGAGGAGGCCTGGGAGTAG-3′ | Activation: 95 °C, 10 min | 40 cycles | 500 nM | XM_005664700.1 | This study |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 60 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| MUC4 | F: 5′-GATGCCCTGGCCACAGAA-3′ R: 5′-TGATTCAAGGTAGCATTCATTTGC-3′ | Activation: 95 °C, 10 min | 40 cycles | 500 nM | NM_001206344.2 | [64] |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 60 °C, 1 min | ||||||
| PRLR | F: 5′-CCAGATACCTAATGACTTCTCAATG-3′ R: 5′-TCCAACAGATGGGTGTCAAA-3′ | Activation: 50 °C, 30 min; | 40 cycles | 500 nM | NM_214084 | [63] |
| 95 °C, 15 min | ||||||
| 1. Denaturation: 94 °C, 15 s | ||||||
| 2. Annealing: 55 °C, 30 s | ||||||
| 3. Extension: 72 °C, 30 s | ||||||
| 77 °C, 15 s | ||||||
| PTGS1 | F: 5′-CAACACTTCACCCACCAGTTCTTC-3′ R: 5′-TCCATAAATGTGGCCGAGGTCTAC-3′ | Activation: 95 °C, 10 min | 40 cycles | 500 nM | AF207823.1 | [65] |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 60 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| TGFA | F: 5′-CGCGCTGGGTATCTTGTTG-3′ R: 5′-GTGGGAATCTGGGCAGTCAT-3′ | Activation: 50 °C, 2 min; | 40 cycles | 200 nM | NM_214251.1 | [66] |
| 95 °C, 10 min | ||||||
| 1. Denaturation: 95 °C, 3 s | ||||||
| 2. Annealing: 60 °C, 30 s | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| ACTB | F: 5′-ACATCAAGGAGAAGCTCTGCTACG-3′ R: 5′-GAGGGGCGATGATCTTGATCTTCA-3′ | Activation: 95 °C, 10 min | 40 cycles | 500 nM | U07786 | [67] |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 61 °C, 1 min | ||||||
| 3. Extension: 72 °C, 1 min | ||||||
| GAPDH | F: 5′-CCTTCATTGACCTCCACTACATGG-3′ R: 5′-CCACAACATACGTAGCACCAGCATC-3′ | Activation: 95 °C, 10 min | 40 cycles | 500 nM | U48832 | [68] |
| 1. Denaturation: 95 °C, 15 s | ||||||
| 2. Annealing: 59 °C, 1 min |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smolinska, N.; Szeszko, K.; Dobrzyn, K.; Kiezun, M.; Rytelewska, E.; Kisielewska, K.; Gudelska, M.; Bors, K.; Wyrebek, J.; Kopij, G.; et al. Transcriptomic Analysis of Porcine Endometrium during Implantation after In Vitro Stimulation by Adiponectin. Int. J. Mol. Sci. 2019, 20, 1335. https://doi.org/10.3390/ijms20061335
Smolinska N, Szeszko K, Dobrzyn K, Kiezun M, Rytelewska E, Kisielewska K, Gudelska M, Bors K, Wyrebek J, Kopij G, et al. Transcriptomic Analysis of Porcine Endometrium during Implantation after In Vitro Stimulation by Adiponectin. International Journal of Molecular Sciences. 2019; 20(6):1335. https://doi.org/10.3390/ijms20061335
Chicago/Turabian StyleSmolinska, Nina, Karol Szeszko, Kamil Dobrzyn, Marta Kiezun, Edyta Rytelewska, Katarzyna Kisielewska, Marlena Gudelska, Kinga Bors, Joanna Wyrebek, Grzegorz Kopij, and et al. 2019. "Transcriptomic Analysis of Porcine Endometrium during Implantation after In Vitro Stimulation by Adiponectin" International Journal of Molecular Sciences 20, no. 6: 1335. https://doi.org/10.3390/ijms20061335