Physiological Diversity of Spitsbergen Soil Microbial Communities Suggests Their Potential as Plant Growth-Promoting Bacteria

 ,
,
Abstract
:1. Introduction
2. Results
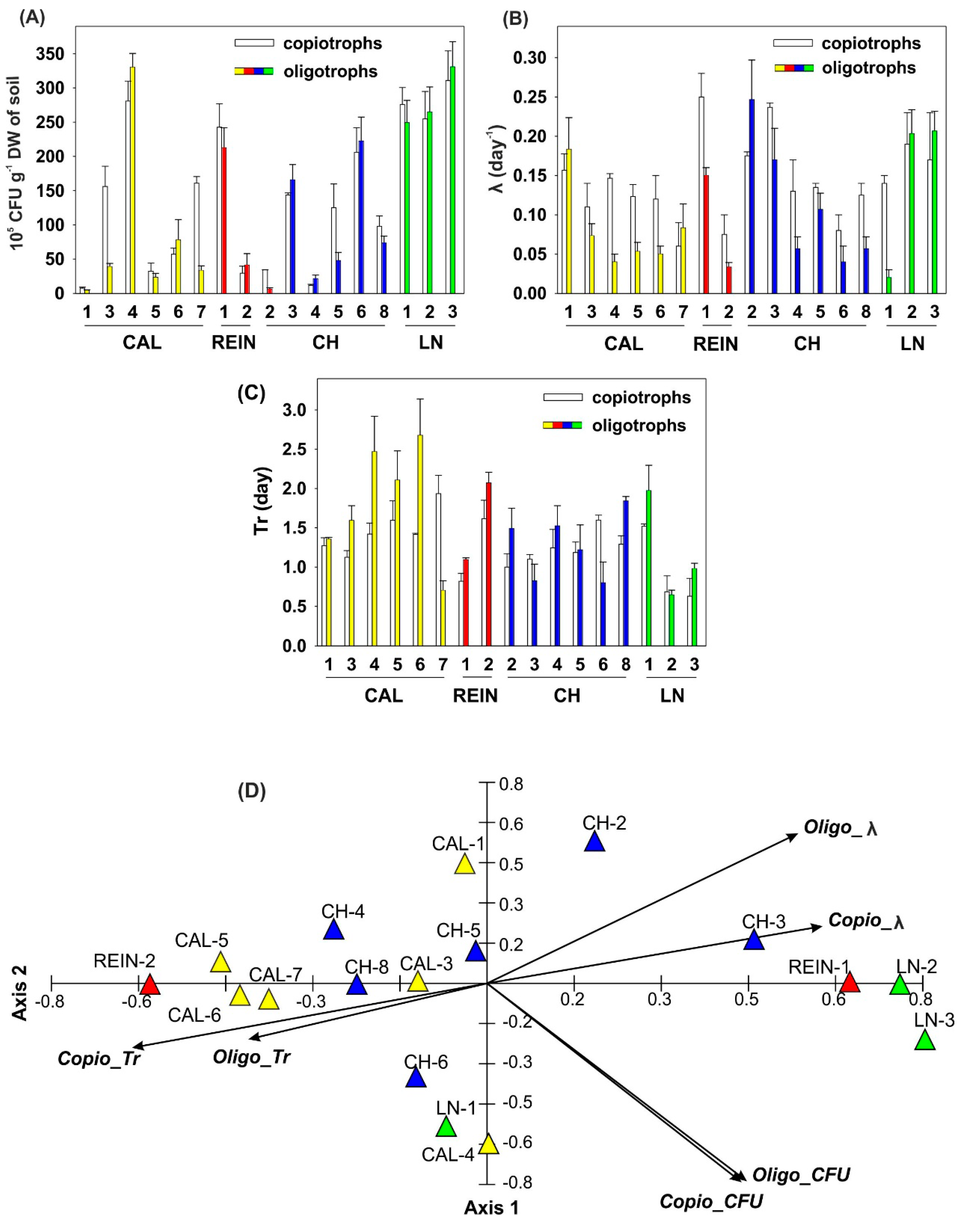
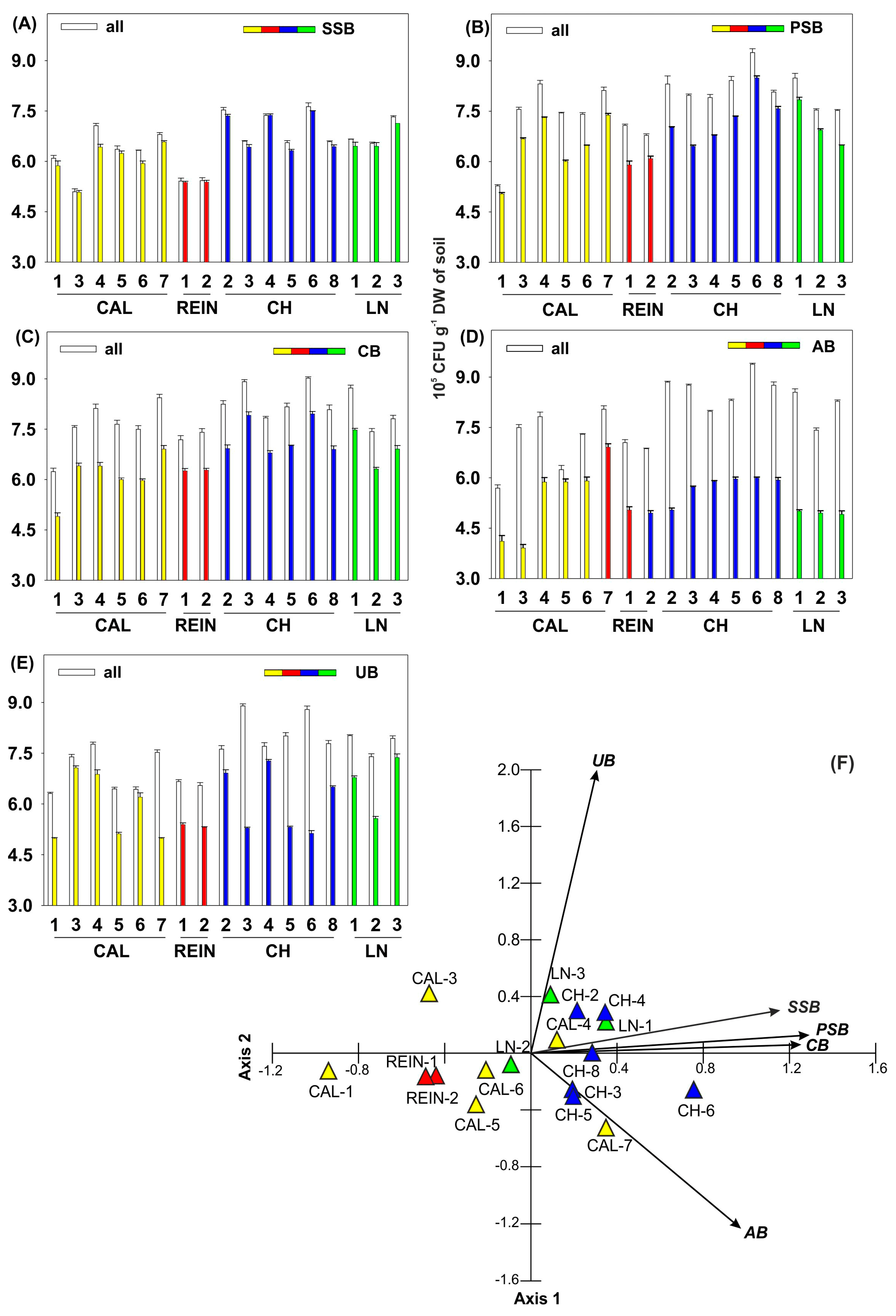
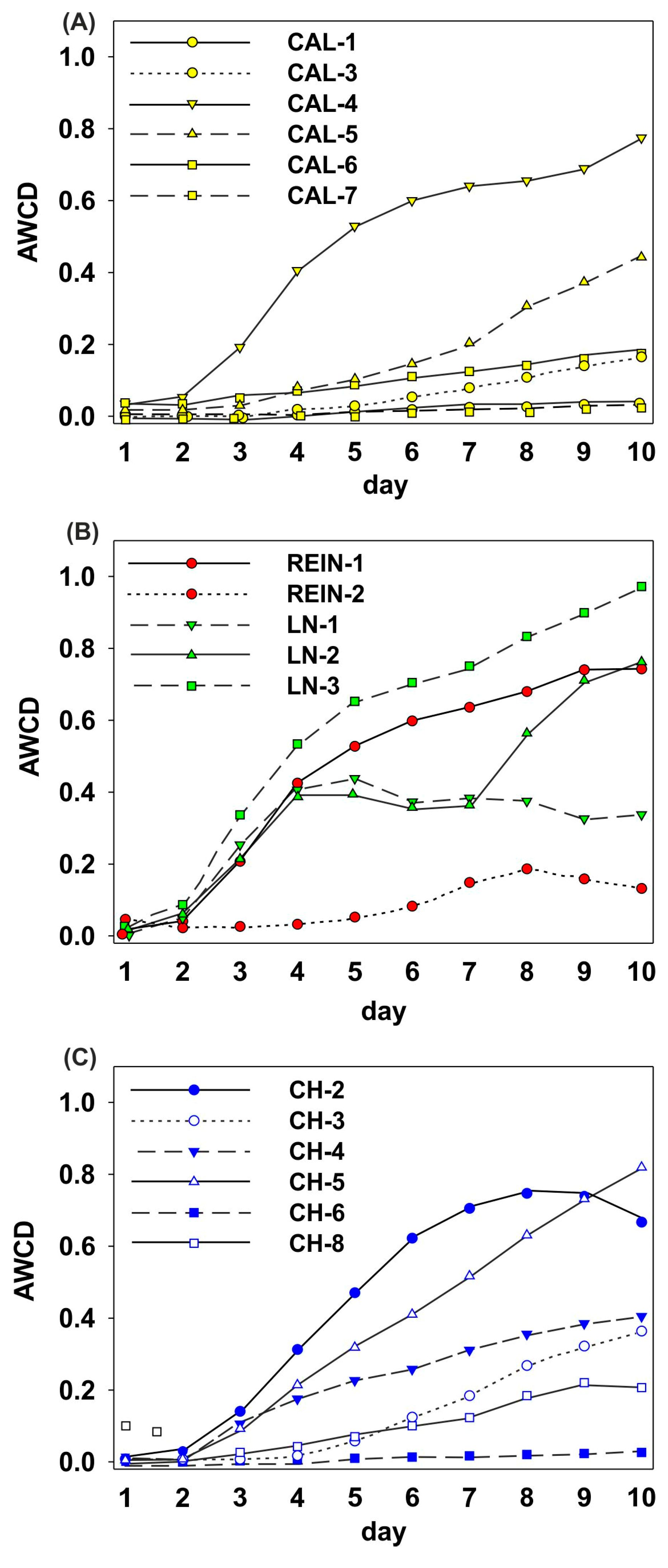
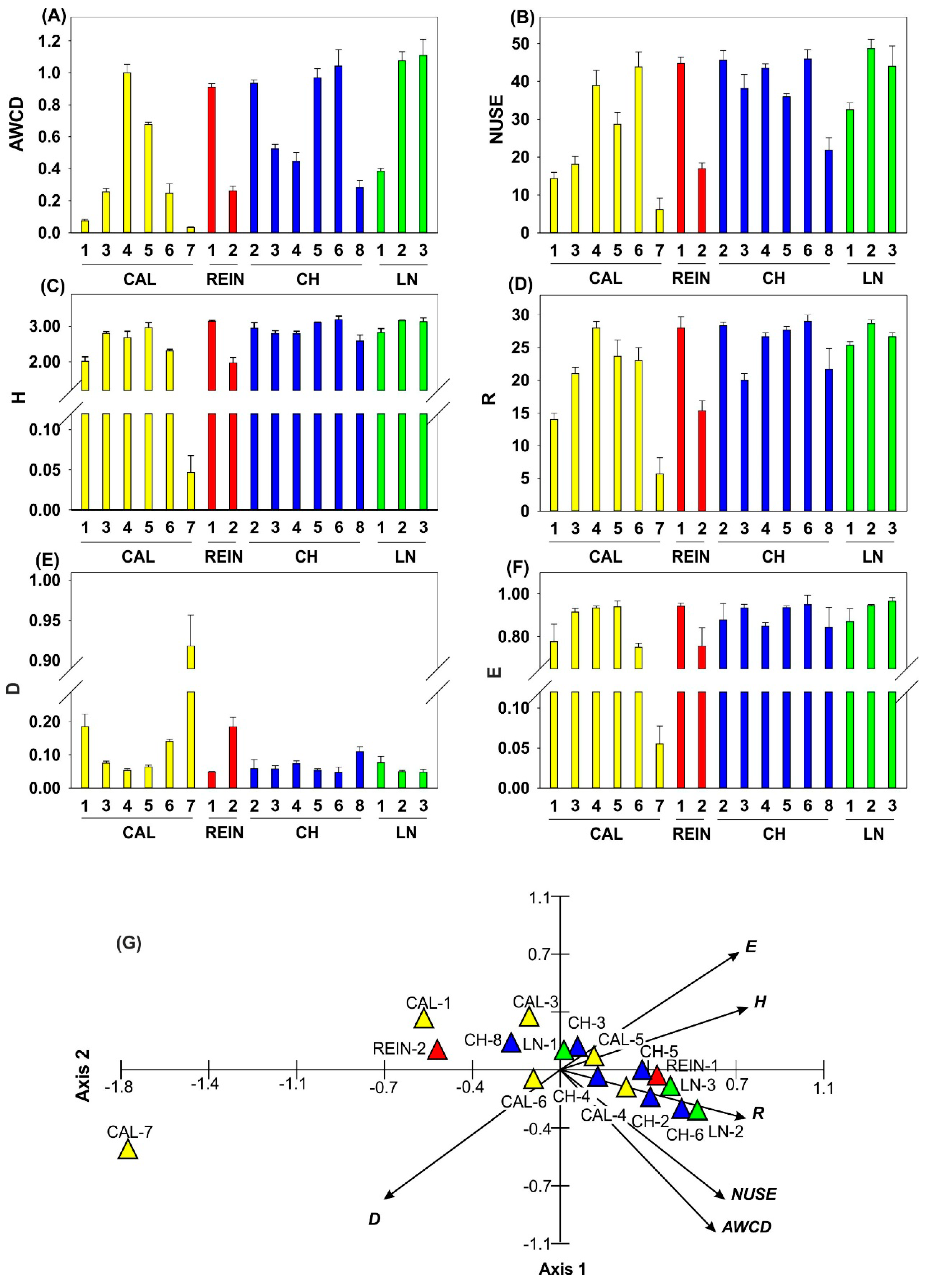
2.1. Microbiological Characteristics of the Soil Samples
2.1.1. Number of Heterotrophic Bacteria and Bacteria with Lytic Activity
2.1.2. Determination of Microbial Activity and Catabolic Diversity
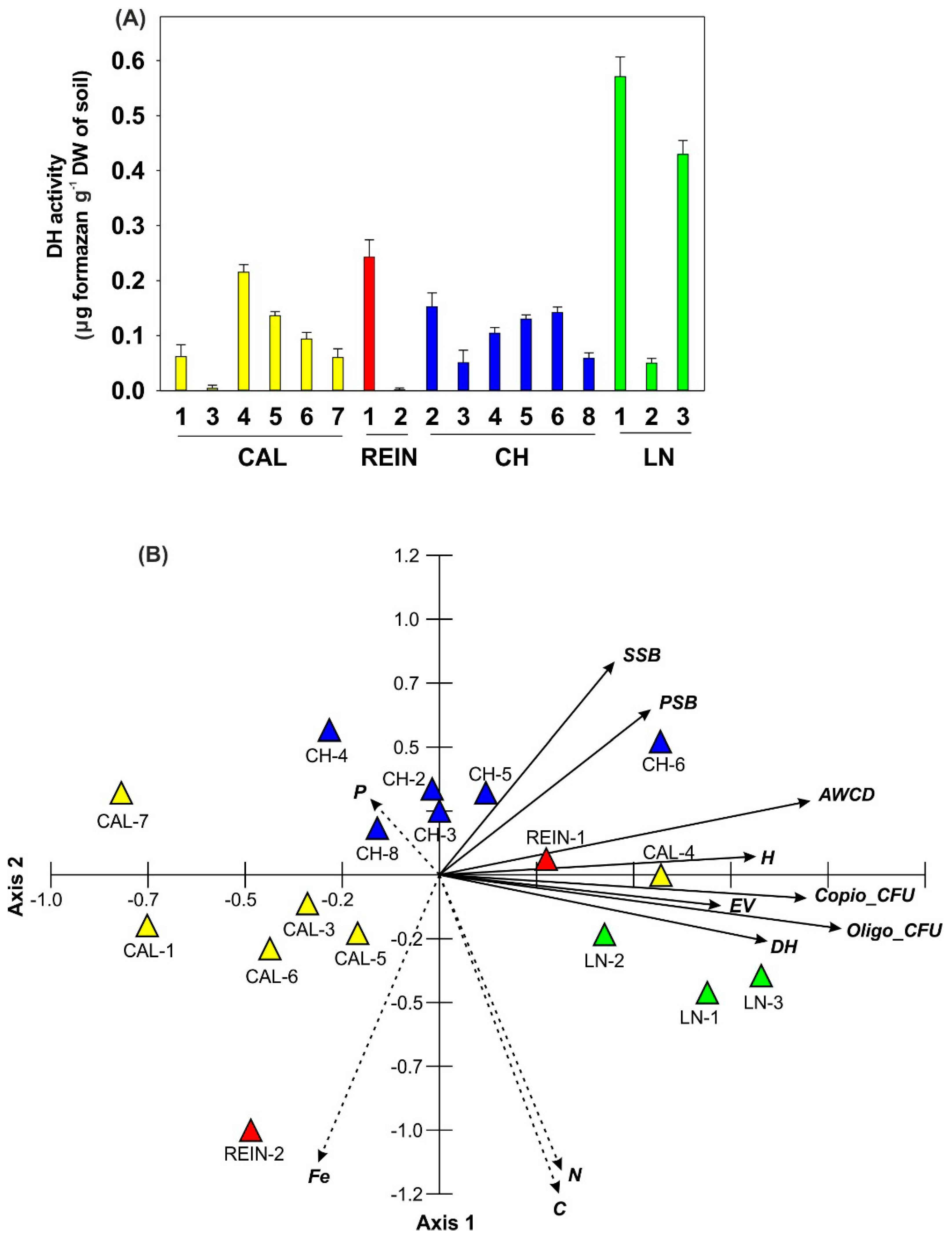
2.1.3. Enzymatic Activity
2.1.4. Relationship among Microbial Parameters Measured for Soil Samples
2.2. Characteristics of Bacterial Isolates
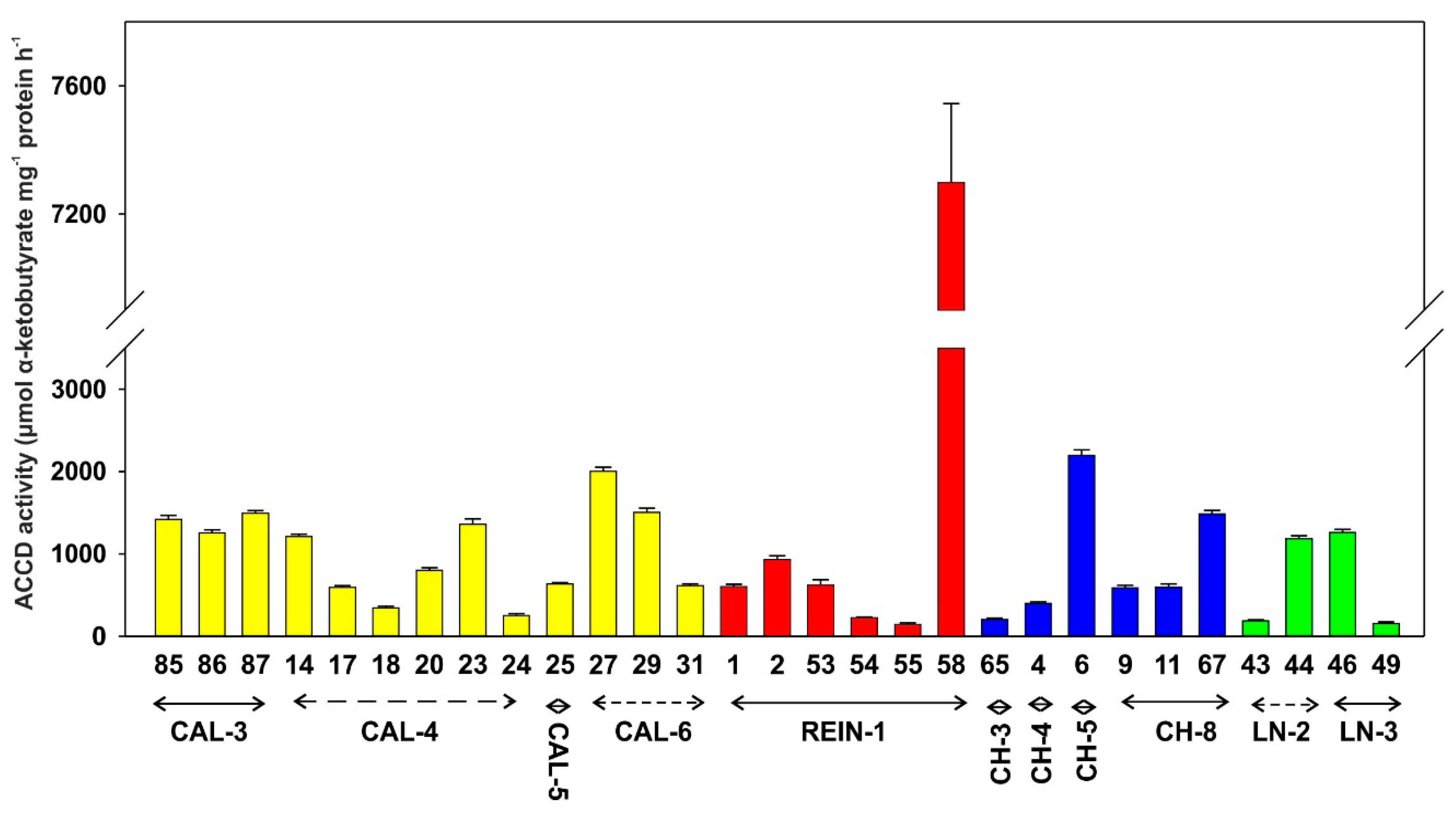
2.2.1. Enzymatic Activity
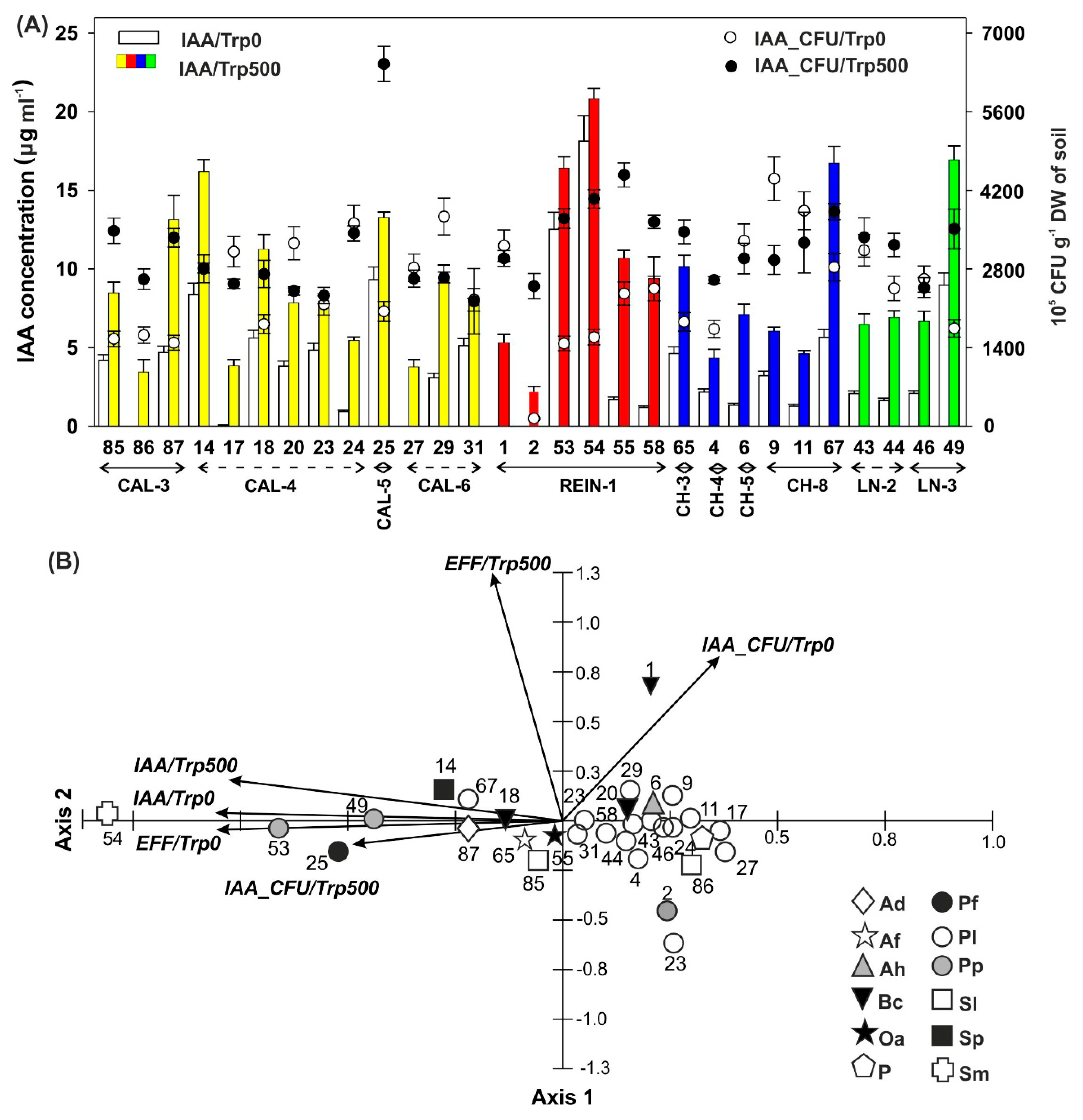
2.2.2. Synthesis of Growth Regulator
2.2.3. Minimal Inhibitory Concentration (MIC) for Cu
2.2.4. Biochemical Identification of Bacterial Species and Their Ability in Siderophore Synthesis
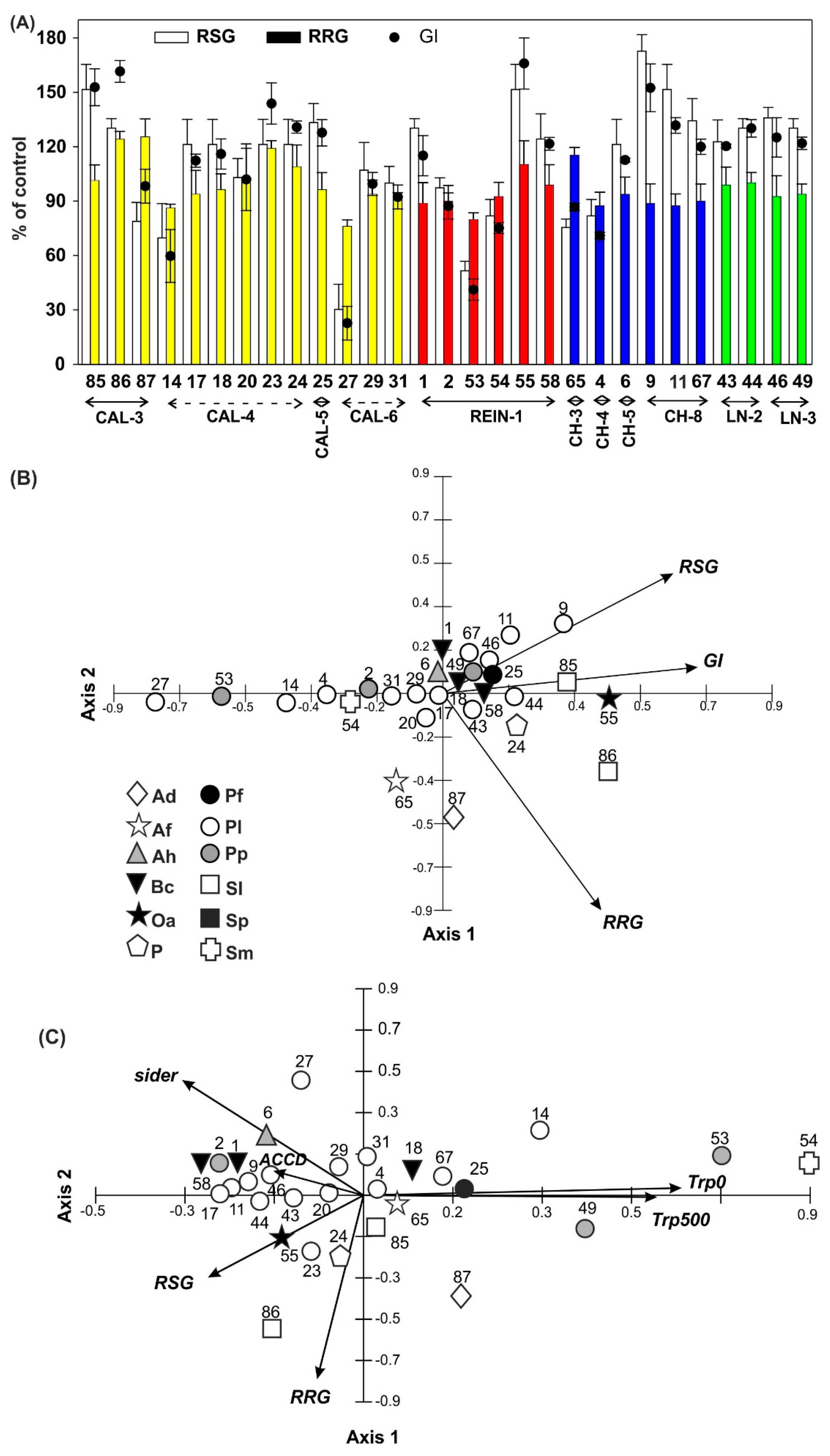
2.2.5. Germination of Seeds
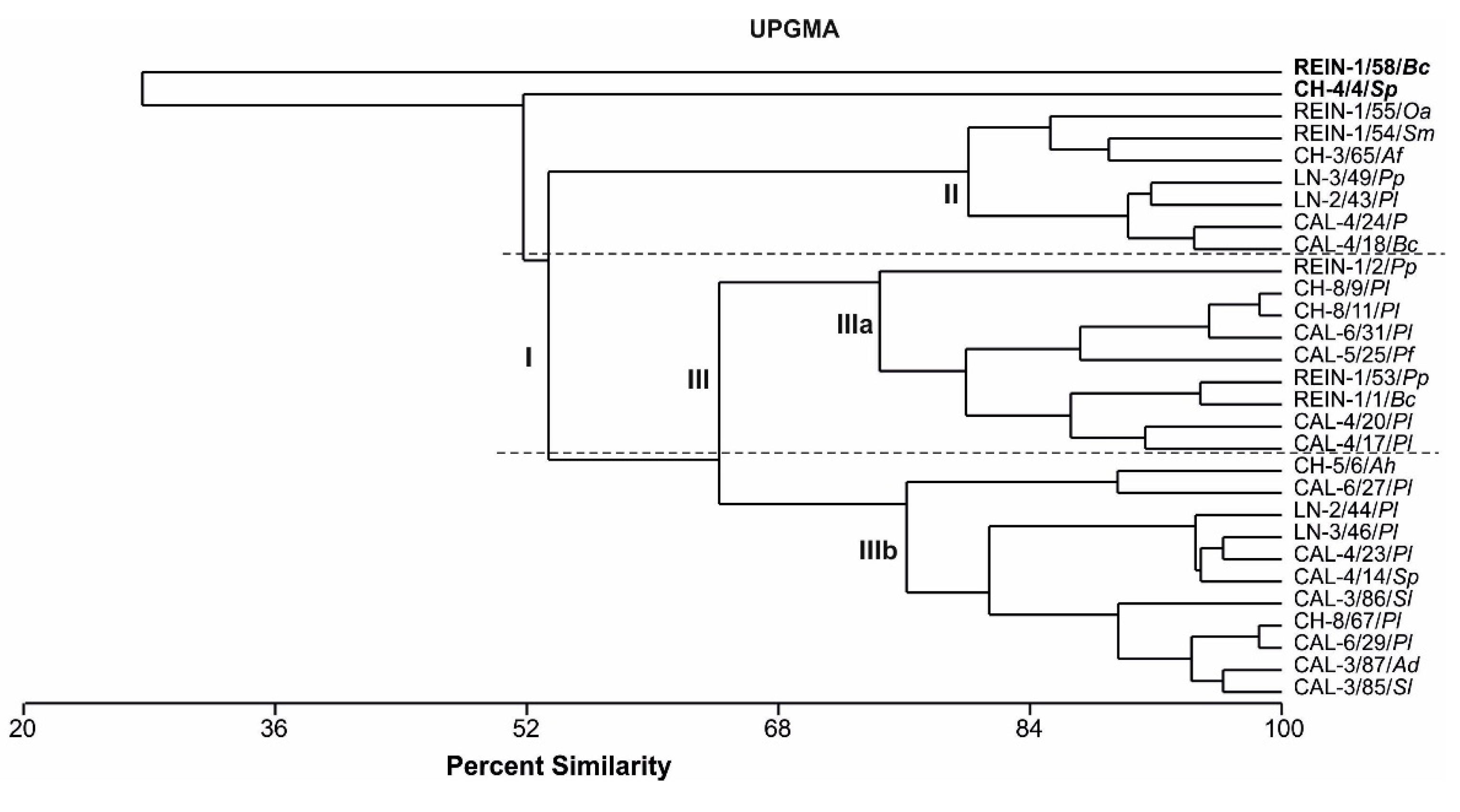
2.2.6. Relationship Among Bacterial Isolates
2.3. Relationship among Microbial Parameters Measured for Soil Samples and Bacterial Isolates
3. Discussion
4. Materials and Methods
4.1. Description of the Study Area and the Soil Samples
4.2. Soil Analysis
4.2.1. Copio- and Oligotrophic Bacteria Enumeration
4.2.2. Determination of Number of Specific Soil Bacteria
4.2.3. Analysis of the Community-Level Physiological Profiling
4.2.4. Determination of Soil Microbial Activity and Catabolic Diversity
4.2.5. Analysis of Dehydrogenase Activity
4.3. Bacterial Activity
4.3.1. ACC Deaminase Activity
4.3.2. Analysis of IAA Synthesis
4.3.3. MIC for Cu
4.3.4. Biochemical Identification of Bacteria
4.3.5. Seed Germination Test
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| PGPB | Plant growth promoting bacteria |
| CB | Cellulolytic bacteria |
| AB | Amylolytic bacteria |
| UB | Ureolytic bacteria |
| SSB | Siderophore synthesizing bacteria |
| PSB | Phosphate solubilizing bacteria |
| ACCD | ACC (1-aminocyclopropane-1-carboxylate) deaminase |
| IAA | Indole-3-acetic acid |
| AWCD | Average well color development |
| Trp | Tryptophan |
| CLPP | Community-level physiological profiling |
| H | Shannon’s diversity index |
| R | Substrate richness |
| E | Substrate evenness |
| D | Domination index |
| NUSE | Nitrogen use index |
| MIC | Minimal Inhibitory Concentration |
| CAL | Calypsostranda |
| REIN | Reinholmen |
| CH | Chamberlindalen |
| LN | Lӕgerneset |
References
- Sabet, H.; Mortazaeinezhad, F. Yield, growth and Fe uptake of cumin (Cuminum cyminum L.) affected by Fe-nano, Fe-chelated and Fe-siderophore fertilization in the calcareous soils. J. Trace Elem. Med. Biol. 2018, 50, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Ernakovich, J.G.; Wallenstein, M.D. Permafrost microbial community traits and functional diversity indicate low activity at in situ thaw temperatures. Soil Biol. Biochem. 2015, 87, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Hayat, R.; Ali, A.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion, a review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Etesami, H. Bacterial mediated alleviation of heavy metal stress and decreased accumulation of metals in plant tissues. Mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 147, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Weber, N.F.; Herrmann, I.; Hochholdinger, F.; Ludewig, U.; Neumann, G. PDPR-induced growth stimulation and nutrient aquisition in maize, do root hairs matter? Sci. Agric. Bohem. 2018, 49, 164–172. [Google Scholar] [CrossRef]
- Tabassum, B.; Khan, A.; Tariq, M.; Ramzan, M.; Khan, M.S.I.; Shahid, N.; Aaliya, K. Bottlenecks in commercialisation and future prospects of PGPR. Appl. Soil Ecol. 2017, 121, 102–117. [Google Scholar] [CrossRef]
- Majeed, A.; Muhammad, Z.; Ahmad, H. Plant growth promoting bacteria, role in soil improvement, abiotic and biotic stress management of crops. Plant Cell Rep. 2018, 37, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Novo, L.A.; Castro, P.M.; Alvarenga, P.; da Silva, E.F. Plant Growth-Promoting Rhizobacteria—Assisted phytoremediation of mine soils. In Bio-Geotechnologies for Mine Site Rehabilitation; Prasad, M.N.V., de Campos Favas, P.J., Maiti, S.K., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 281–295. [Google Scholar]
- Souza, R.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef] [Green Version]
- Lobo, C.B.; Tomás, M.S.J.; Viruel, E.; Marcela-Ferrero, A.; Lucca, M.E. Development of low-cost formulations of plant growth-promoting bacteria to be used as inoculants in beneficial agricultural technologies. Microbiol. Res. 2019, 219, 12–25. [Google Scholar] [CrossRef]
- Ozimek, E.; Jaroszuk-Ściseł, J.; Bohacz, J.; Korniłłowicz-Kowalska, T.; Tyśkiewicz, R.; Słomka, A.; Nowak, A.; Hanaka, A. Synthesis of indoleacetic acid, gibberellic acid and ACC-deaminase by Mortierella strains promote winter wheat seedlings growth under different conditions. Int. J. Mol. Sci. 2018, 19, 3218. [Google Scholar] [CrossRef] [PubMed]
- Spaepen, S.; Vnderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaroszuk-Ściseł, J.; Kurek, E.; Trytek, M. Efficiency of indoleacetic acid, gibberellic acid and ethylene synthesized in vitro by Fusarium culmorum strains with different effects on cereal growth. Biologia 2014, 69, 281–292. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Ethylene and 1-aminocyclopropane-1-carboxylate (ACC) in plant-bacterial interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Shaharoona, B.; Bibi, R.M.; Arshad, Z.A.; Ul-Hassan, Z. 1-Aminocylopropane-1-carboxylate (ACC) deaminase-rhizobacteria extenuates ACC-induced classical triple response in etiolated pea seedlings. Pak. J. Bot. 2006, 38, 1491–1499. [Google Scholar]
- Robison, J.D.; Yamasaki, Y.; Randall, S.K. The ethylene signaling pathway negatively impacts CBF/DREB-regulated cold response in soybean (Glycine max). Front. Plant Sci. 2019, 10, 121. [Google Scholar] [CrossRef]
- Li, M.; Guo, R.; Yu, F.; Chen, X.; Zhao, H.; Li, H.; Wu, J. Indole-3-acetic acid biosynthesis pathways in the plant-beneficial bacterium Arthrobacter pascens ZZ21. Int. J. Mol. Sci. 2018, 19, 443. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Mohite, B. Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nutr. 2013, 13, 638–649. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef]
- Bashan, Y.; de Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth promoting bacterial inoculant technology, formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Wang, G.H.; Jin, J.; Chen, X.L.; Liu, J.D.; Liu, X.B.; Herbert, S.J. Biomass and catabolic diversity of microbial communities with long-term restoration, bare fallow and cropping history in Chinese Mollisols. Plant Soil Environ. 2007, 53, 177–185. [Google Scholar] [CrossRef]
- Wolińska, A.; Frąc, M.; Oszust, K.; Szafranek-Nakonieczna, A.; Zielenkiewicz, U.; Stępniewska, Z. Microbial biodiversity of meadows under different modes of land use: Catabolic and genetic fingerprinting. World J. Microbiol. Biotechnol. 2017, 33, 154. [Google Scholar] [CrossRef] [PubMed]
- Hanaka, A.; Plak, A.; Zagórski, P.; Ozimek, E.; Rysiak, A.; Majewska, M.; Jaroszuk-Ściseł, J. Relationships between the properties of Spitsbergen soil, number and biodiversity of rhizosphere microorganisms, and heavy metal concentration in selected plant species. Plant Soil 2019. [Google Scholar] [CrossRef]
- Kenarova, A.; Encheva, M.; Chipeva, V.; Chipev, N.; Hristova, P.; Moncheva, P. Physiological diversity of bacterial communities from different soil locations on Livingston Island, South Shetland archipelago, Antarctica. Polar Biol. 2013, 36, 223–233. [Google Scholar] [CrossRef]
- Kim, H.M.; Chae, N.; Jung, J.Y.; Lee, Y.K. Isolation of facultatively anaerobic soil bacteria from Ny-Ålesund, Svalbard. Polar Biol. 2013, 36, 787–796. [Google Scholar] [CrossRef]
- Delshadi, S.; Ebrahimi, M.; Shirmohammadi, E. Influence of plant-growth-promoting bacteria on germination, growth and nutrients’ uptake of Onobrychis sativa L. under drought stress. J. Plant Interact. 2017, 12, 200–208. [Google Scholar] [CrossRef]
- Hattori, T. Further analysis of plate data of bacteria count. J. Gen. Appl. Microbiol. 1983, 29, 9–16. [Google Scholar] [CrossRef]
- Hattori, T. Kinetics of colony formation of bacteria, an approach to the basis of the plate count method. In The Reports of the Institute for Agricultural Research Tohoku University; Tohoku University: Sendai, Japan, 1985; Volume 34, pp. 1–36. [Google Scholar]
- López, D.; Vlamakis, H.; Losick, R.; Kolter, R. Cannibalism enhances biofilm development in Bacillus subtilis. Mol. Microbiol. 2009, 74, 609–618. [Google Scholar] [CrossRef]
- Awan, K.; Jabeen, F.; Manzoor, M.; Qazi, J.I. Potential of thermophilic amylolytic bacteria for growth in unconventional media, Potato peels. J. Food Process Eng. 2017, 41, e12635. [Google Scholar] [CrossRef]
- Weber, K.P.; Legge, R.L. Community-level physiological profiling. Methods Mol. Biol. 2010, 599, 63–81. [Google Scholar] [CrossRef]
- Campbell, C.D.; Grayston, S.J.; Hirst, D.J. Use of rhizosphere carbon sources in sole carbon source tests to discriminate soil microbial communities. J. Microbiol. Meth. 1997, 30, 33–41. [Google Scholar] [CrossRef]
- Xie, W.; Zhou, J.; Wang, H.; Chen, X.; Lu, Z.; Yu, J.; Chen, X. Short-term effects of copper, cadmium and cypermethrin on dehydrogenase activity and microbial functional diversity in soils after long-term mineral or organic fertilization. Agric. Ecosyst. Environ. 2009, 129, 450–456. [Google Scholar] [CrossRef]
- Grządziel, J.; Gałązka, A. Microplot long-term experiment reveals strong soil type influence on bacteria composition and its functional diversity. Appl. Soil Ecol. 2018, 124, 117–123. [Google Scholar] [CrossRef]
- Wolińska, A.; Rekosz-Burlaga, H.; Goryluk-Salmonowicz, A.; Błaszczyk, M.; Stępniewska, Z. Bacterial abundance and dehydrogenase activity in selected agricultural soils from Lublin region. Pol. J. Environ. Stud. 2015, 24, 2677–2682. [Google Scholar] [CrossRef]
- He, L.; Ye, J.; Wu, B.; Huang, L.; Ren, J.; Wu, X. Effects of genetically modified Burkholderia pyrrocinia JK-SH007E1 on soil microbial community in polar rhizosphere. For. Pathol. 2016, 48, e12430. [Google Scholar] [CrossRef]
- Kurek, E.; Jaroszuk-Ściseł, J. Rye (Secale cereale) growth promotion by Pseudomonas fluorescens strains and their interactions with Fusarium culmorum under various soil conditions. Biol. Control 2003, 26, 48–56. [Google Scholar] [CrossRef]
- Pathak, D.V.; Kumar, M.; Rani, K. Biofertilizer application in horticultural crops. In Microorganisms for Green Revolution; Panpatte, D.G., Jhala, Y.K., Vyas, R.V., Shelat, H.N., Eds.; Springer: Singapore, 2017; pp. 215–227. [Google Scholar]
- Hanif, K.; Hameed, S.; Imran, A.; Naqqash, T.; Shahid, M.; Van Elsas, J.D. Isolation and characterization of a β-propeller gene containing phosphobacterium Bacillus subtilis strain KPS-11 for growth promotion of potato (Solanum tuberosum L.). Front. Microbiol. 2015, 6, 583. [Google Scholar] [CrossRef]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase, a weapon to “stress ethylene” produced in plants. Front. Microbiol. 2015, 6, 937. [Google Scholar] [CrossRef]
- Croes, S.; Weyens, N.; Janssen, J.; Vercampt, H.; Colpaert, J.V.; Carleer, R.; Vangronsveld, J. Bacterial communities associated with Brassica napus L. grown on trace element-contaminated and non-contaminated fields, a genotypic and phenotypic comparison. Microb. Biotechnol. 2013, 6, 371–384. [Google Scholar] [CrossRef]
- Thao, N.P.; Khan, M.I.R.; Thu, N.B.A.; Hoang, X.L.T.; Asgher, M.; Khan, N.A.; Tran, L.S.P. Role of ethylene and its cross talk with other signaling molecules in plant responses to heavy metal stress. Plant Physiol. 2015, 169, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Acuña, J.J.; Campos, M.; de la Luz Mora, M.; Jaisi, D.P.; Jorquera, M.A. ACCD-producing rhizobacteria from an Andean Altiplano native plant (Parastrephia quadrangularis) and their potential to alleviate salt stress in wheat seedlings. Appl. Soil Ecol. 2019. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, K.; Li, K.; Du, W.; He, S.; Liu, H.W. Reaction of 1-amino-2-methylenecyclopropane-1-carboxylate with 1-aminocyclopropane-1-carboxylate deaminase: Analysis and mechanistic implications. Biochemistry 2003, 42, 2089–2103. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A renaissance of elicitors, perception of microbe associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Koevoets, I.T.; Venema, J.H.; Elzenga, J.T.M.; Testerink, C. Roots withstanding their environment, exploiting root system architecture responses to abiotic stress to improve crop tolerance. Front. Plant Sci. 2016, 7, 1335. [Google Scholar] [CrossRef]
- Kumar, K.V.; Singh, N.; Behl, H.M.; Srivastava, S. Influence of plant growth promoting bacteria and its mutant on heavy metal toxicity in Brassica juncea grown in fly ash amended soil. Chemosphere 2008, 72, 678–683. [Google Scholar] [CrossRef]
- Gosh, S.; Basu, P.S. Production and metabolism of indole acetic acid in roots and root nodule of Phaseolus mungo. Microbiol. Res. 2006, 161, 362–366. [Google Scholar] [CrossRef]
- Hinsinger, P.; Plassard, C.; Tang, C.X.; Jaillard, B. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints, a review. Plant Soil 2003, 248, 43–59. [Google Scholar] [CrossRef]
- Leveau, J.H.; Lindow, S.E. Utilization of the plant hormone indole-3-acetic acid for growth by Pseudomonas putida strain 1290. Appl. Environ. Microbiol. 2005, 71, 2365–2371. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef]
- Ponmurugan, P.; Gopi, C. In vitro production of growth regulators and phosphatase activity by phosphate solubilizing bacteria. Afr. J. Biotechnol. 2006, 5, 348–350. [Google Scholar]
- Kurek, E.; Ozimek, E.; Sobiczewski, P.; Słomka, A.; Jaroszuk-Ściseł, J. Effect of Pseudomonas luteola on mobilization of phosphorusand growth of young apple trees (Ligol)—Pot experiment. Sci. Hortic. 2013, 164, 270–276. [Google Scholar] [CrossRef]
- van Noorden, G.E.; Ross, J.J.; Reid, J.B.; Rolfe, B.G.; Mathesius, U. Defective long-distance auxin transport regulation in the Medicago truncatula super numeric nodules mutant. Plant Physiol. 2006, 140, 1494–1506. [Google Scholar] [CrossRef] [PubMed]
- Persello-Cartieaux, F.; David, P.; Sarrobert, C.; Thibaud, M.C.; Achouak, W.; Robaglia, C.; Nussaume, L. Utilization of mutants to analyze the interaction between Arabidopsis thaliana and its naturally root-associated Pseudomonas. Planta 2001, 212, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Pasternak, J.J.; Glick, B.R. Isolation and characterization of mutants of the plant growth-promoting rhizobacterium Pseudomonas putida CR12-2 that overproduce indoleacetic acid. Curr. Microbiol. 1996, 32, 67–71. [Google Scholar] [CrossRef]
- Staal, M.; De Cnodder, T.; Simon, D.; Vandenbussche, F.; Van der Straeten, D.; Verbelen, J.-P.; Elzenga, T.; Vissenberg, K. Apoplastic alkalinization is instrumental for the inhibition of cell elongation in the Arabidopsis root by the ethylene precursor 1-aminocyclopropane-1-carboxylic acid. Plant. Physiol. 2011, 155, 2049–2055. [Google Scholar] [CrossRef] [PubMed]
- Bosshard, P.P.; Zbinden, R.; Abels, S.; Böddinghaus, B.; Altwegg, M.; Böttger, E.C. 16S rRNA gene sequencing versus the API 20 NE system and the VITEK 2 ID-GNB card for identification of nonfermenting Gram-negative bacteria in the clinical laboratory. J. Clin. Microbiol. 2006, 44, 1359–1366. [Google Scholar] [CrossRef]
- Pampuro, N.; Bisaglia, C.; Romano, E.; Brambilla, M.; Pedretti, E.F.; Cavallo, E. Phytotoxicity and chemical characterization of compost derived from pig slurry solid fraction for organic pellet production. Agriculture 2017, 7, 94. [Google Scholar] [CrossRef]
- Mochizuki, M.; Hattori, T. Kinetics of microcolony formation of a soil oligotrophic bacterium, Agromonas sp. (Time-lapse microscopy: Cell kinetics; growth initiation). FEMS Microbiol. Ecol. 1986, 38, 51–55. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Microbiology 1948, 17, 362–370. [Google Scholar]
- Ugwuanyi, J.O.; Obeta, J.A.N. Pectynolytic and celluloplytic activities of heat resistant fungi and their macerating effects on mango and African mango. J. Sci. Agric. 1999, 79, 1054–1059. [Google Scholar] [CrossRef]
- Kasana, R.C.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A.A. Rapid and easy method for the detection of microbial cellulases on agar plates using gram’s iodine. Curr. Microbiol. 2008, 57, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M. Handbook of Media for Environmental Microbiology; CRC Press: Boca Raton, FL, USA, 1995; p. 495. [Google Scholar]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescein. J. Lab. Clin. Med. 1954, 59, 743–747. [Google Scholar]
- Feigl, V.; Ujaczki, E.; Vaszita, E.; Molnár, M. Influence of red mud on soil microbial communities. Application and comprehensive evaluation of the Biolog EcoPlate approach as a tool in soil microbiological studies. Sci. Total Environ. 2017, 595, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Vratama, C.J.; Bernas, S.M.; Amin, M. Water quality assessment based on biological and chemical analysis as a parameter for development of fresh water fishery in Lubuk Karet River of Banyuasin District. Ind. J. Environ. Manag. Sust. 2018, 2, 64–68. [Google Scholar] [CrossRef]
- Sala, M.M.; Pinhassi, J.; Gasol, J.M. Estimation of bacterial use of dissolved organic nitrogen compounds in aquatic ecosystems using Biolog plates. Aquat. Microb. Ecol. 2006, 42, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Tu, C.; Guan, F.; Sun, Y.; Guo, P.; Liu, Y.; Li, L.; Scheckel, K.G.; Luo, Y. Stabilizing effects on a Cd polluted coastal wetland soil using calcium polysulphide. Geoderma 2018, 332, 190–197. [Google Scholar] [CrossRef]
- Casida, L.E.; Klein, D.A.; Santoro, T. Soil dehydrogenase activity. Soil Sci. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Alef, K. Enzyme activities. In Methods in Applied Soil Microbiology and Biochemistry; Alef, K., Nannipieri, P., Eds.; Academic Press: New York, NY, USA, 1995; pp. 123–191. [Google Scholar]
- Januszek, K.; Błońska, E.; Długa, J.; Socha, J. Dehydrogenase activity of forest soils depends on the assay used. Int. Agrophys. 2015, 29, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Belimov, A.A.; Dodd, I.C.; Safronova, V.I.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M. Rhizobacteria that produce auxins and contain 1-amino-cyclopropane-1-carboxylic acid deaminase decrease amino acid concentrations in the rhizosphere and improve growth and yield of well-watered and water-limited potato (Solanum tuberosum). Ann. Appl. Biol. 2015, 167, 11–25. [Google Scholar] [CrossRef]
- Bradford, M. Rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [PubMed]
- Kovach, W.L. MVSP–A Multi-Variate Statistical Package for Windows, Ver. 3.1; Kovach Computing Services: Pentraeth, Wales, UK, 1999; pp. 1–145. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Sample | Isolate No | Bacteria Genus/Species | Bacteria Abbreviation | ID [%] | Siderophore Synthesis |
|---|---|---|---|---|---|
| CAL-3 | 85 | Serratia liquefaciens | Sl | 96.3 | No |
| CAL-3 | 86 | Serratia liquefaciens | Sl | 95.9 | No |
| CAL-3 | 87 | Achromobacter denitrificans | Ad | 82.2 | No |
| CAL-4 | 14 | Serratia plymuthica | Sp | 99.9 | Yes |
| CAL-4 | 17 | Pseudomonas luteola | Pl | 92.5 | Yes |
| CAL-4 | 18 | Burkholderia cepacia | Bc | 67.8 | Yes |
| CAL-4 | 20 | Pseudomonas luteola | Pl | 92.5 | Yes |
| CAL-4 | 23 | Pseudomonas luteola | Pl | 92.5 | Yes |
| CAL-4 | 24 | Pantoea spp 1* | P | 82.3 | No |
| CAL-5 | 25 | Pseudomonas fluorescens | Pf | 99.9 | Yes |
| CAL-6 | 27 | Pseudomonas luteola | Pl | 92.5 | Yes |
| CAL-6 | 29 | Pseudomonas luteola | Pl | 99.8 | Yes |
| CAL-6 | 31 | Pseudomonas luteola | Pl | 99.8 | Yes |
| REIN-1 | 1 | Burkholderia cepacia | Bc | 67.8 | Yes |
| REIN-1 | 2 | Pseudomonas putida | Pp | 87.9 | Yes |
| REIN-1 | 53 | Pseudomonas putida | Pp | 89.1 | No |
| REIN-1 | 54 | Stenotrophomonas maltophilia | Sm | 99.9 | No |
| REIN-1 | 55 | Ochrobactrum anthropi | Oa | 95.9 | Yes |
| REIN-1 | 58 | Burkholderia cepacia | Bc | 67.8 | Yes |
| CH-3 | 65 | Alcaligenes faecalis 1 | Af | 90.1 | Yes |
| CH-4 | 4 | Serratia plymuthica | Sp | 99.9 | No |
| CH-5 | 6 | Aeromonas hydrophila | Ah | 99.7 | Yes |
| CH-8 | 9 | Pseudomonas luteola | Pl | 99.8 | Yes |
| CH-8 | 11 | Pseudomonas luteola | Pl | 99.5 | Yes |
| CH-8 | 67 | Pseudomonas luteola | Pl | 99.8 | Yes |
| LN-2 | 43 | Pseudomonas luteola | Pl | 99.8 | Yes |
| LN-2 | 44 | Pseudomonas luteola | Pl | 99.5 | Yes |
| LN-3 | 46 | Pseudomonas luteola | Pl | 99.8 | Yes |
| LN-3 | 49 | Pseudomonas putida | Pp | 99.1 | No |
| Soil Sample | Area | Coordinates | Dominant Plant Species | C 1 | N 1 | C/N 1 | P 1 | Fe 1 | |
|---|---|---|---|---|---|---|---|---|---|
| N | E | % | mg kg−1 | g kg−1 | |||||
| CAL-1 | CAL | 77°34′10″ | 14°25′26″ | Sax | 4.37 ± 0.22 | 0.052 ± 0.003 | 84.04 | 3.44 ± 0.49 | 0.61 ± 0.08 |
| CAL-3 | 77°33′57″ | 14°28′32″ | Dra | 3.50 ± 0.17 | 0.105 ± 0.013 | 33.33 | 1.09 ± 0.11 | 0.52 ± 0.04 | |
| CAL-4 | 77°33′55″ | 14°28′57″ | Dry | 5.19 ± 0.12 | 0.394 ± 0.036 | 13.17 | 1.99 ± 0.16 | 0.40 ± 0.02 | |
| CAL-5 | 77°33′55″ | 14°28′57″ | Dry | 5.55 ± 0.04 | 0.381 ± 0.013 | 14.19 | 3.08 ± 0.15 | 0.65 ± 0.07 | |
| CAL-6 | 77°33′55″ | 14°28′57″ | Sax | 4.31 ± 0.01 | 0.310 ± 0.020 | 13.90 | 1.81 ± 0.14 | 0.74 ± 0.15 | |
| CAL-7 | 77°33′59″ | 14°29′41″ | Dra | 2.40 ± 0.19 | 0.075 ± 0.009 | 32.00 | 10.70 ± 1.05 | 0.34 ± 0.01 | |
| REIN-1 | REIN | 77°29′45″ | 14°33′17″ | Dry | 3.15 ± 0.20 | 0.202 ± 0.036 | 15.59 | 19.04 ± 1.95 | 0.29 ± 0.04 |
| REIN-2 | 77°29′45″ | 14°33′20″ | ns | 25.42 ± 1.49 | 1.253 ± 0.103 | 20.29 | 40.43 ± 2.11 | 0.70 ± 0.01 | |
| CH-2 | CH | 77°29′00″ | 14°31′48″ | Dry | 3.01 ± 0.13 | 0.218 ± 0.057 | 13.81 | 21.94 ± 1.40 | 0.42 ± 0.03 |
| CH-3 | 77°29′00″ | 14°31′51″ | ns | 0.88 ± 0.06 | 0.079 ± 0.005 | 11.14 | 4.89 ± 0.31 | 0.36 ± 0.02 | |
| CH-4 | 77°28′59″ | 14°31′53″ | Sal | 1.15 ± 0.12 | 0.116 ± 0.023 | 9.91 | 95.36 ± 4.23 | 0.26 ± 0.01 | |
| CH-5 | 77°28′59″ | 14°31′52″ | ns | 1.81 ± 0.14 | 0.148 ± 0.012 | 12.23 | 6.35 ± 0.51 | 0.31 ± 0.02 | |
| CH-6 | 77°28′52″ | 14°32′01″ | ns | 2.85 ± 0.29 | 0.235 ± 0.035 | 12.09 | 13.05 ± 1.27 | 0.27 ± 0.07 | |
| CH-7 | 77°34′10″ | 14°25′26″ | ns | 1.25 ± 0.41 | 0.145 ± 0.006 | 8.62 | 3.99 ± 0.40 | 0.39 ± 0.08 | |
| CH-8 | 77°34′04″ | 14°28′05″ | ns | 4.13 ± 0.26 | 0.399 ± 0.025 | 10.33 | 11.24 ± 0.72 | 0.31 ± 0.02 | |
| LN-1 | LN | 77°33′55″ | 14°28′57″ | Dry | 19.42 ± 0.24 | 0.883 ± 0.077 | 21.99 | 7.61 ± 0.69 | 0.37 ± 0.08 |
| LN-2 | 77°33′55″ | 14°28′57″ | ns | 12.62 ± 0.50 | 0.356 ± 0.032 | 35.45 | 4.71 ± 0.11 | 0.60 ± 0.01 | |
| LN-3 | 77°33′59″ | 14°29′41″ | ns | 15.23 ± 0.59 | 0.628 ± 0.055 | 24.25 | 13.96 ± 0.49 | 0.58 ± 0.08 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanaka, A.; Ozimek, E.; Majewska, M.; Rysiak, A.; Jaroszuk-Ściseł, J. Physiological Diversity of Spitsbergen Soil Microbial Communities Suggests Their Potential as Plant Growth-Promoting Bacteria. Int. J. Mol. Sci. 2019, 20, 1207. https://doi.org/10.3390/ijms20051207
Hanaka A, Ozimek E, Majewska M, Rysiak A, Jaroszuk-Ściseł J. Physiological Diversity of Spitsbergen Soil Microbial Communities Suggests Their Potential as Plant Growth-Promoting Bacteria. International Journal of Molecular Sciences. 2019; 20(5):1207. https://doi.org/10.3390/ijms20051207
Chicago/Turabian StyleHanaka, Agnieszka, Ewa Ozimek, Małgorzata Majewska, Anna Rysiak, and Jolanta Jaroszuk-Ściseł. 2019. "Physiological Diversity of Spitsbergen Soil Microbial Communities Suggests Their Potential as Plant Growth-Promoting Bacteria" International Journal of Molecular Sciences 20, no. 5: 1207. https://doi.org/10.3390/ijms20051207