Grain Legumes and Fear of Salt Stress: Focus on Mechanisms and Management Strategies

Abstract
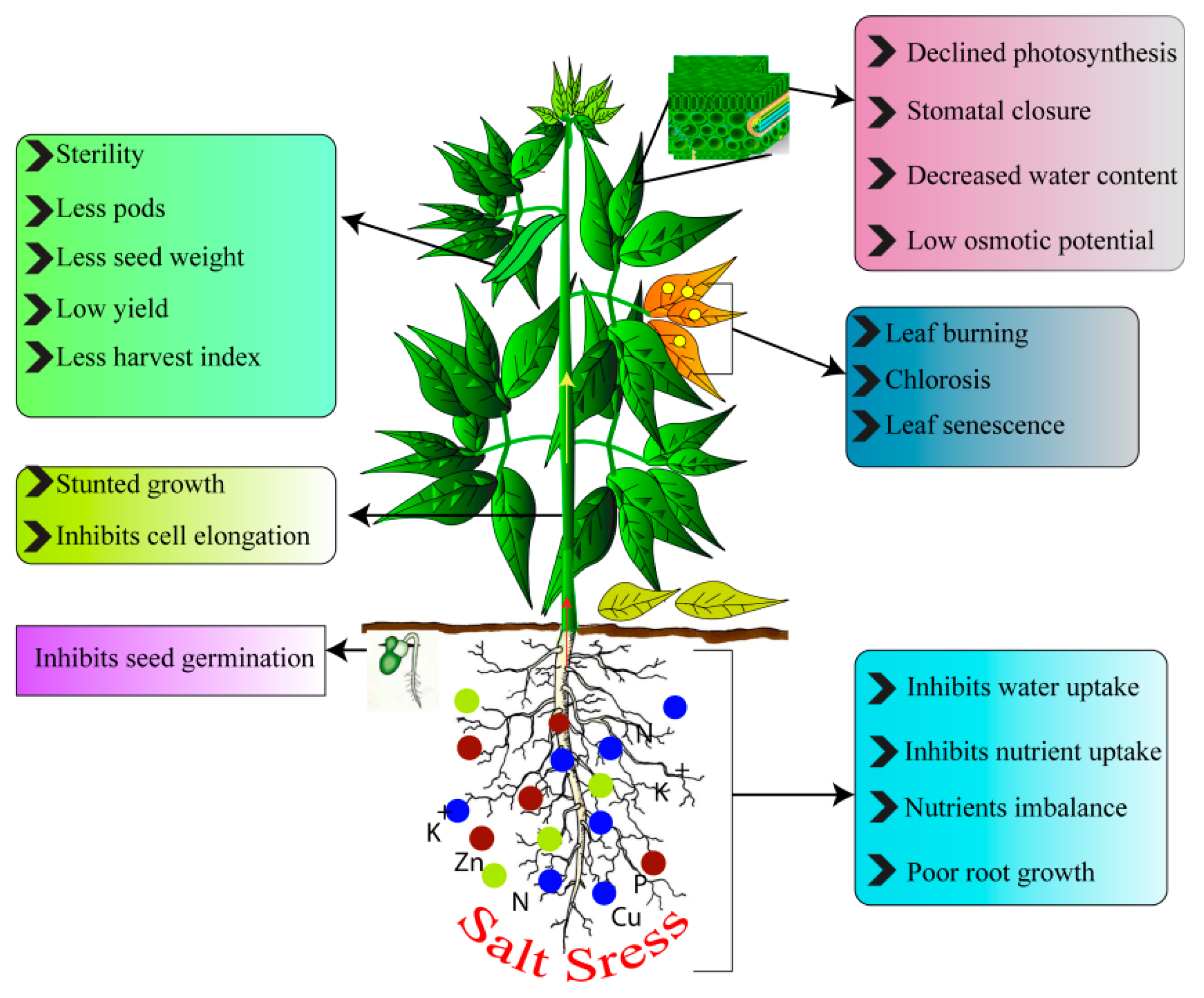
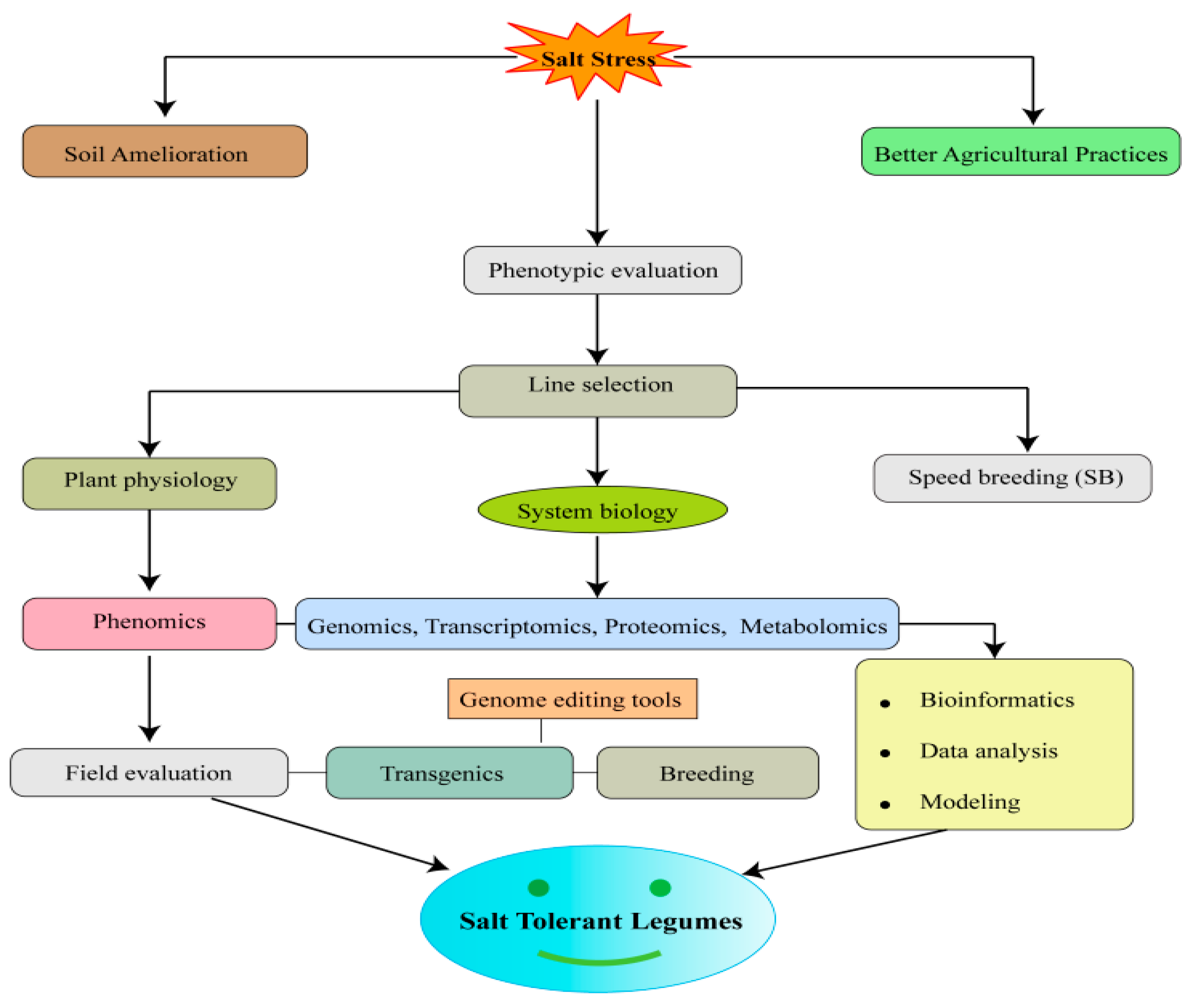
:1. Introduction
2. Legumes Sensitivity to Salt Stress (SS)
3. Tolerance Mechanisms
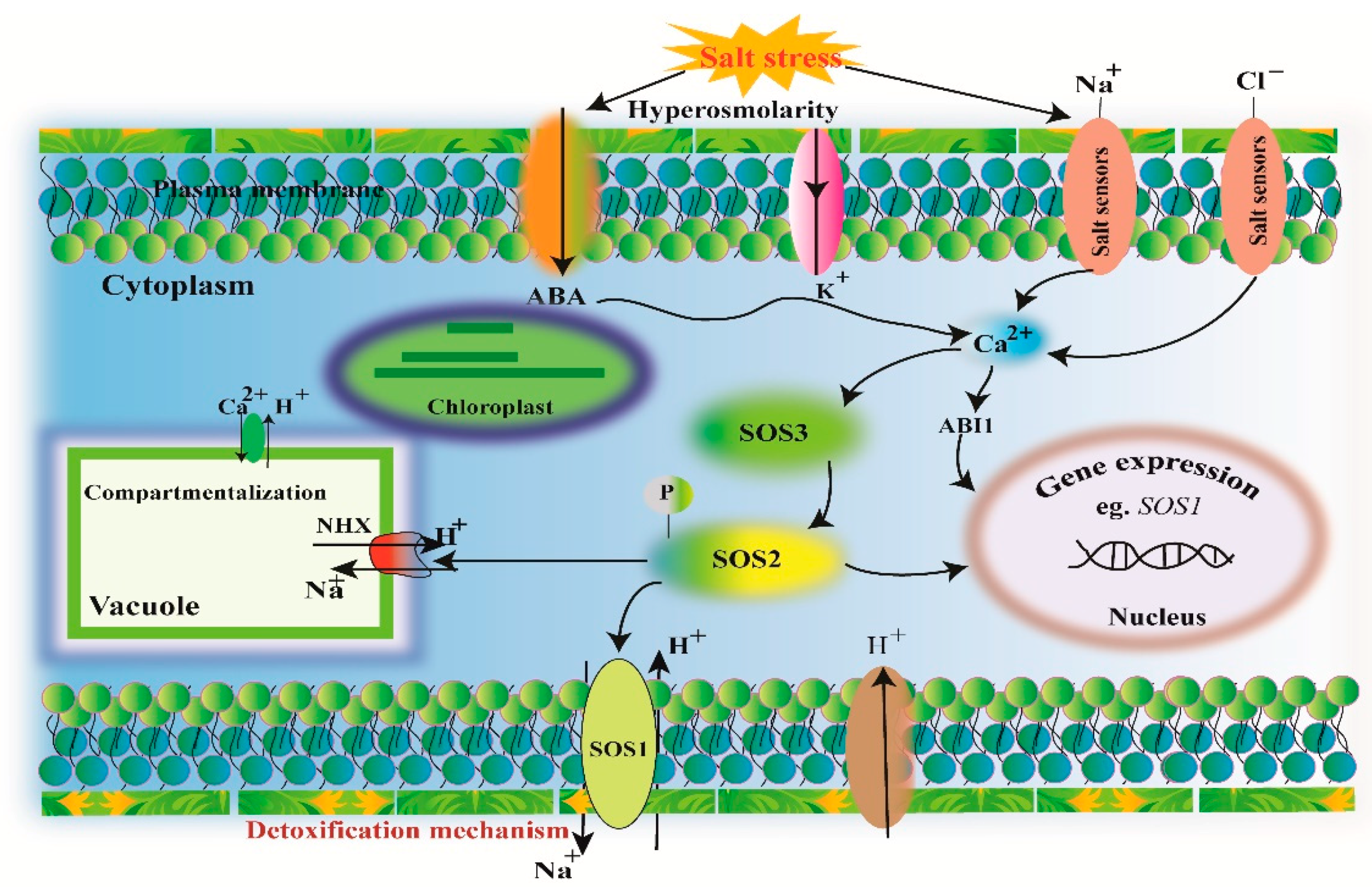
3.1. Ion Homeostasis and Salt Tolerance
3.2. Compatible Solute Accumulation and Osmotic Protection
3.3. Antioxidant Regulation of Salinity Tolerance
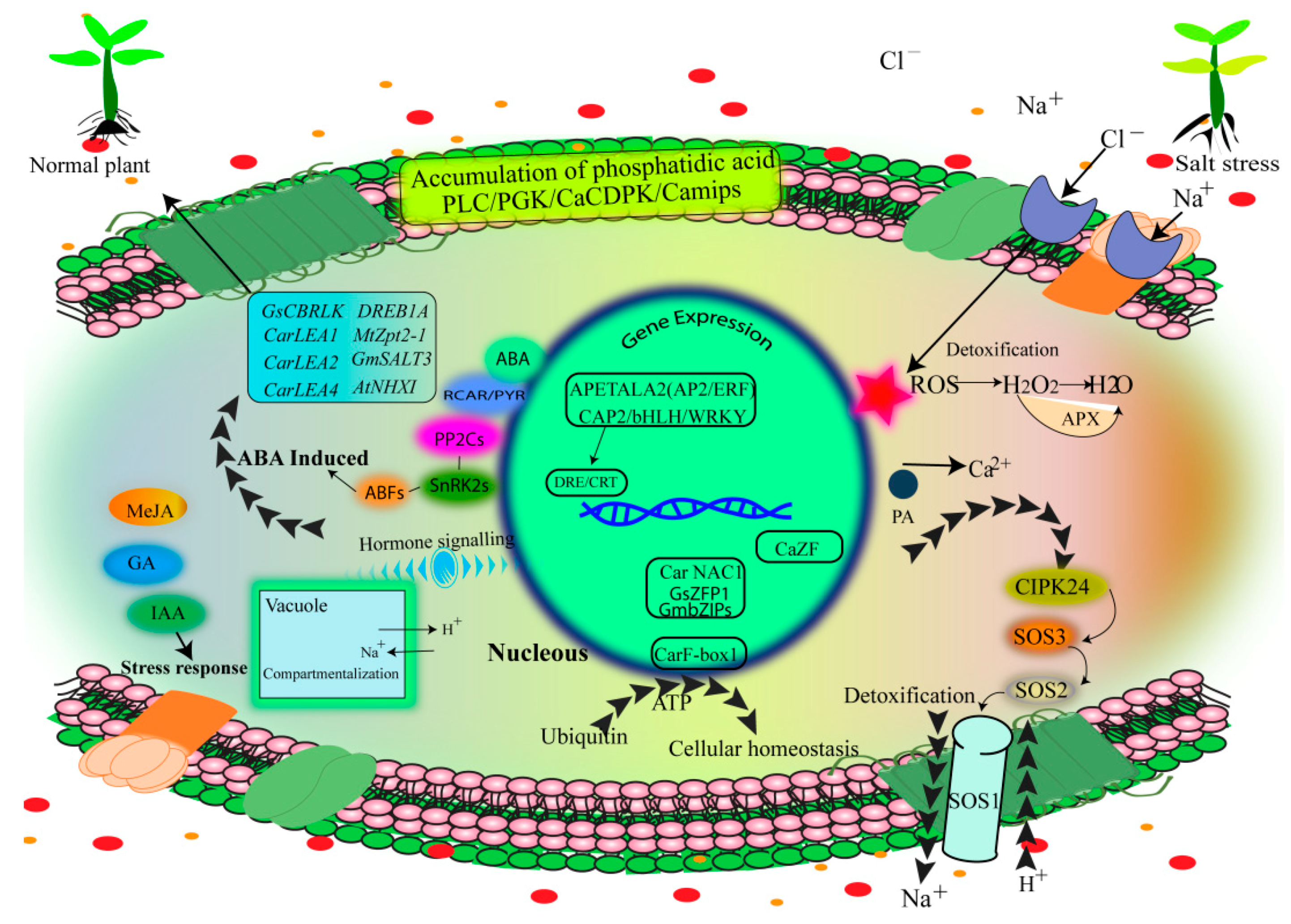
3.4. Hormone Regulation of Salinity Tolerance
4. Management Strategies to Improve Salt Tolerance in Legumes
4.1. Agronomic Strategies to Reduce SS
4.1.1. Reclamation of Salt-Affected Soils by Nutrients Management
4.1.2. Reclamation of Salt-Affected Soils by Water Management
4.2. Plant Growth-Promoting Rhizobacteria
4.3. Seed Priming
4.4. Role of Polyamines in Salinity Tolerance
4.5. Selection and Conventional Breeding Approaches
4.6. Bioengineering and Functional Genomics
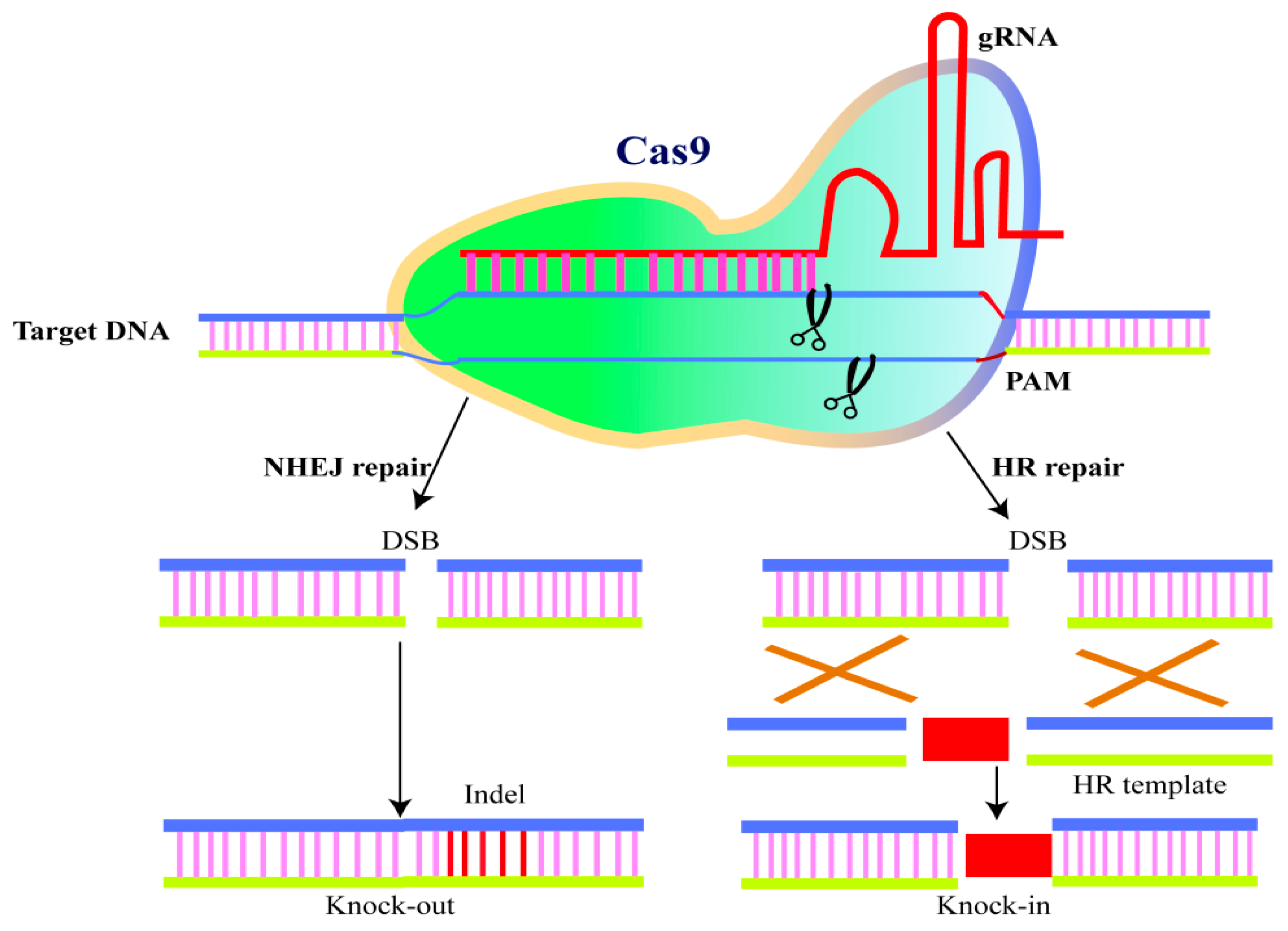
4.7. CRISPR-Cas9: Master Player for Genome Editing (GE)
5. Conclusions and Future Research Perspectives
Funding
Conflicts of Interest
Abbreviations
| GA | Gibberellin |
| ABA | Abscisic acid |
| SOS | Salt overly sensitive |
| GB | Glycine betaine |
| ROS | Reactive oxygen species |
| GR | Glutathione reductase |
| GPX | Glutathione peroxidases |
| APX | Ascorbate peroxidase |
| SOD | Superoxide dismutase |
| GST | Glutathione S-transferases |
| MDHAR | Monodehydroascorbate reductase |
| CAT | Catalase |
| PGPR | Plant growth promoting rhizobacteria |
| WGS | Whole genome sequencing |
| SNP | Single-nucleotide polymorphism |
| SSRs | Simple sequence repeats |
| NGS | Next-generation sequencing |
| MDA | Malondialdehyde |
| GE | Genome editing |
| TALENs | Transcriptional activator-like effector nuclease |
| ZFNs | Zinc finger nuclease |
References
- Kromdijk, J.; Long, S.P. One crop breeding cycle from starvation? How engineering crop photosynthesis for rising CO2 and temperature could be one important route to alleviation. Proc. R. Soc. B Biol. Sci. 2016, 283. [Google Scholar] [CrossRef]
- Pareek, A.; Sopory, S.K.; Bohnert, H.J. Govindjee Abiotic stress adaptation in plants: Physiological, molecular and genomic foundation. Abiotic Stress Adapt. Plants Physiol. Mol. Genomic Found. 2010, 99, 1–526. [Google Scholar] [CrossRef]
- Kaashyap, M.; Ford, R.; Bohra, A.; Kuvalekar, A.; Mantri, N. Improving salt tolerance of chickpea using modern genomics tools and molecular breeding. Curr. Genom. 2017, 18, 557–567. [Google Scholar] [CrossRef]
- Nadeem, M.; Li, J.; Wang, M.; Shah, L.; Lu, S.; Wang, X.; Ma, C. Unraveling field crops sensitivity to heat stress: Mechanisms, approaches, and future prospects. Agronomy 2018, 8, 128. [Google Scholar] [CrossRef]
- El-nasharty, A.B.; El-nwehy, S.S.; Rezk, A.I.; El-Motaleb Aly Abou El-Nour, E.-Z.A. Role of kinetin in improving salt tolerance of two wheat cultivars. Biosci. Res. 2017, 2, 193–200. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Status World’s Soil Resources; FAO: Rome, Italy, 2015; 650p, ISBN 978-92-5-109004-6. [Google Scholar]
- Flowers, T.J.; Flowers, S.A. Why does salinity pose such a difficult problem for plant breeders? Agric. Water Manag. 2005, 78, 15–24. [Google Scholar] [CrossRef]
- Qureshi, M.I.; Muneer, S.; Bashir, H.; Ahmad, J.; Iqbal, M. Nodule physiology and proteomics of stressed legumes. Adv. Bot. Res. 2010, 56, 1–48. [Google Scholar]
- Mishra, S.; Panda, S.K.; Sahoo, L. Transgenic asiatic grain legumes for salt tolerance and functional genomics. Rev. Agric. Sci. 2014, 2, 21–36. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef]
- Murillo-Amador, B.; Yamada, S.; Yamaguchi, T.; Rueda-Puente, E.; Ávila-Serrano, N.; García-Hernández, J.L.; López-Aguilar, R.; Troyo-Diéguez, E.; Nieto-Garibay, A. Influence of calcium silicate on growth, physiological parameters and mineral nutrition in two legume species under salt stress. J. Agron. Crop Sci. 2007, 193, 413–421. [Google Scholar] [CrossRef]
- He, Y.; Fu, J.; Yu, C.; Wang, X.; Jiang, Q.; Hong, J.; Lu, K.; Xue, G.; Yan, C.; James, A.; et al. Increasing cyclic electron flow is related to Na+ sequestration into vacuoles for salt tolerance in soybean. J. Exp. Bot. 2015, 66, 6877–6889. [Google Scholar] [CrossRef] [PubMed]
- Patil, G.; Do, T.; Vuong, T.D.; Valliyodan, B.; Lee, J.D.; Chaudhary, J.; Shannon, J.G.; Nguyen, H.T. Genomic-assisted haplotype analysis and the development of high-throughput SNP markers for salinity tolerance in soybean. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- López-Aguilar, R.; Orduño-Cruz, A.; Lucero-Arce, A.; Murillo-Amador, B.; Troyo-Diéguez, E. Response to salinity of three grain legumes for potential cultivation in arid areas. Soil Sci. Plant Nutr. 2003, 49, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Manchanda, G.; Garg, N. Salinity and its effects on the functional biology of legumes. Acta Physiol. Plant. 2008, 30, 595–618. [Google Scholar] [CrossRef]
- Sehrawat, N.; Bhat, K.V.; Sairam, R.K.; Jaiwal, P.K. Identification of salt resistant wild relatives of mungbean (Vigna radiata L. Wilczek). Asian J. Plant Sci. Res. 2013, 3, 41–49. [Google Scholar]
- Khan, H.A.; Siddique, K.H.M.; Colmer, T.D. Vegetative and reproductive growth of salt-stressed chickpea are carbon-limited: Sucrose infusion at the reproductive stage improves salt tolerance. J. Exp. Bot. 2017, 68, 2001–2011. [Google Scholar] [CrossRef]
- Flowers, T.J.; Gaur, P.M.; Gowda, C.L.L.; Krishnamurthy, L.; Samineni, S.; Siddique, K.H.M.; Turner, N.C.; Vadez, V.; Varshney, R.K.; Colmer, T.D. Salt sensitivity in chickpea. Plant Cell Environ. 2010, 33, 490–509. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Van Hoorn, J.W.; Katerji, N.; Hamdy, A.; Mastrorilli, M. Effect of salinity on yield and nitrogen uptake of four grain legumes and on biological nitrogen contribution from the soil. Agric. Water Manag. 2001, 51, 87–98. [Google Scholar] [CrossRef]
- Yadav, H.D.; Yadav, O.P.; Dhankar, O.P.; Oswal, M.C. Effect of chloride salintiy and gernmination, growth and mineral composition of chickpea. Ann. Arid Zone 1989, 28, 63–67. [Google Scholar]
- El Sayed, H.E.S. Influence of NaCl and Na2SO4 treatments on growth development of broad bean (Vicia faba L.) plant. J. Life Sci. 2011, 5, 513–523. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and salt tolerance: Bringing them togethe. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Shu, K.; Qi, Y.; Chen, F.; Meng, Y.; Luo, X.; Shuai, H.; Zhou, W.; Ding, J.; Du, J.; Liu, J.; et al. Salt stress represses soybean seed germination by negatively regulating GA biosynthesis while positively mediating ABA biosynthesis. Front. Plant Sci. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A. Germination behaviour of soybean varieties under different salinity stress. Int. J. Appl. Agric. Res. 2017, 12, 69–76. [Google Scholar]
- Ma, Q.; Kang, J.; Long, R.; Zhang, T.; Xiong, J.; Zhang, K.; Wang, T.; Yang, Q.; Sun, Y. Comparative proteomic analysis of alfalfa revealed new salt and drought stress-related factors involved in seed germination. Mol. Biol. Rep. 2017, 44, 261–272. [Google Scholar] [CrossRef]
- Haileselasie, T.H.; Teferii, G. The effect of salinity stress on germination of chickpea (Cicer arietinum L.) Land Race of Tigray. Curr. Res. J. Biol. Sci. 2012, 4, 578–583. [Google Scholar]
- Bayuelo-Jiménez, J.S.; Craig, R.; Lynch, J.P. Salinity tolerance of Phaseolus species during germination and early seedling growth. Crop Sci. 2002, 42, 1584–1594. [Google Scholar] [CrossRef]
- Al-Mutawa, M.M. Effect of salinity on germination and seedling growth of chickpea (Cicer arietinum) genotypes. Int. J. Agric. Biol. 2003, 5, 226–229. [Google Scholar] [CrossRef]
- Sehrawat, N.; Yadav, M.; Bhat, K.; Sairam, R.; Jaiwal, P. Effect of salinity stress on mungbean [Vigna radiata (L.) Wilczek] during consecutive summer and spring seasons. J. Agric. Sci. Belgrade 2015, 60, 23–32. [Google Scholar] [CrossRef]
- Luo, G.Z.; Wang, Y.J.; Xie, Z.M.; Gai, J.Y.; Zhang, J.S.; Chen, S.Y. The putative ser/thr protein kinase gene CmAAPK from soybean is regulated by abiotic stress. J. Integr. Plant Biol. 2006, 48, 327–333. [Google Scholar] [CrossRef]
- Bandeoǧlu, E.; Eyidoǧan, F.; Yücel, M.; Öktem, H.A. Antioxidant responses of shoots and roots of lentil to NaCl-salinity stress. Plant Growth Regul. 2004, 42, 69–77. [Google Scholar] [CrossRef]
- Pitann, B.; Kranz, T.; Zörb, C.; Walter, A.; Schurr, U.; Mühling, K.H. Apoplastic pH and growth in expanding leaves of Vicia faba under salinity. Environ. Exp. Bot. 2011, 74, 31–36. [Google Scholar] [CrossRef]
- Garg, N.; Bhandari, P. Silicon nutrition and mycorrhizal inoculations improve growth, nutrient status, K+/Na+ ratio and yield of Cicer arietinum L. genotypes under salinity stress. Plant Growth Regul. 2016, 78, 371–387. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. Role of arbuscular mycorrhizae in the alleviation of ionic, osmotic and oxidative stresses induced by salinity in Cajanus cajan (L.) Millsp. (pigeonpea). J. Agron. Crop Sci. 2009, 195, 110–123. [Google Scholar] [CrossRef]
- Do, T.D.; Vuong, T.D.; Dunn, D.; Smothers, S.; Patil, G.; Yungbluth, D.C.; Chen, P.; Scaboo, A.; Xu, D.; Carter, T.E.; et al. Mapping and confirmation of loci for salt tolerance in a novel soybean germplasm, Fiskeby III. Theor. Appl. Genet. 2018, 131, 513–524. [Google Scholar] [CrossRef]
- Ning, L.; Kan, G.; Shao, H.; Yu, D. Physiological and transcriptional responses to salt stress in salt-tolerant and salt-sensitive soybean (Glycine max [L.] Merr.) seedlings. L. Degrad. Dev. 2018, 29, 2707–2719. [Google Scholar] [CrossRef]
- Eyidogan, F.; Öz, M.T. Effect of salinity on antioxidant responses of chickpea seedlings. Acta Physiol. Plant. 2007, 29, 485–493. [Google Scholar] [CrossRef]
- Khan, H.A.; Siddique, K.H.M.; Munir, R.; Colmer, T.D. Salt sensitivity in chickpea: Growth, photosynthesis, seed yield components and tissue ion regulation in contrasting genotypes. J. Plant Physiol. 2015, 182, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Lei, J.Q.; Zeng, F.J.; Zhang, B.; Liu, G.J.; Liu, B.; Li, X.Y. Effect of NaCl-induced changes in growth, photosynthetic characteristics, water status and enzymatic antioxidant system of Calligonum caput-medusae seedlings. Photosynthetica 2017, 55, 96–106. [Google Scholar] [CrossRef]
- Shanko, D.; Jateni, G.; Debela, A. Effects of salinity on chickpea (Cicer arietinum L.) landraces during germination stage. Biochem. Mol. Biol. J. 2017, 03, 214–219. [Google Scholar] [CrossRef]
- Kaya, M.D.; Ipek, A.; Öztürk, A. Effects of different soil salinity levels on germination and seedling growth of safflower (Carthamus tinctorius L.). Turk. J. Agric. For. 2003, 27, 221–227. [Google Scholar] [CrossRef]
- Talei, D.; Kadir, M.A.; Yusop, M.K.; Abdullah, M.P.; Valdiani, A. Salinity effects on macro and micronutrients uptake in medicinal plant King of Bitters (Andrographis paniculata Nees.). Plant Omics 2012, 5, 271–278. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Torabian, S.; Farhangi-Abriz, S.; Rathjen, J. Biochar and lignite affect H+-ATPase and H+-PPase activities in root tonoplast and nutrient contents of mungbean under salt stress. Plant Physiol. Biochem. 2018, 129, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Nandwal, A.S.; Godara, M.; Kamboj, D.V.; Kundu, B.S.; Mann, A.; Kumar, B.; Sharma, S.K. Nodule functioning in trifoliate and pentafoliate mungbean genotypes as influenced by salinity. Biol. Plant. 2000, 43, 459–462. [Google Scholar] [CrossRef]
- Gholipoor, M.; Ghasemi-Golezani, K.; Khooie, F.R.; Moghaddam, M. Effects of salinity on initial seedling growth of chickpea (Cicer Arietinum L.). Acta Agron. Hungarica 2001, 48, 337–343. [Google Scholar] [CrossRef]
- Essa, T.A. Effect of salinity stress on growth and nutrient composition of three soybean (Glycine max L. Merrill) Cultivars. J. Agron. Crop Sci. 2002, 93, 86–93. [Google Scholar] [CrossRef]
- Rabie, G.H.; Almadini, A.M. Role of bioinoculants in development of salt-tolerance of Vicia faba plants under salinity stress. Afr. J. Biotechnol. 2005, 4, 210–222. [Google Scholar] [CrossRef]
- Cordovilla, M.P.; Ocaña, A.; Ligero, F.; Lluch, C. Salinity effects on growth analysis and nutrient composition in four grain ulgumes-rhizobium symbiosis. J. Plant Nutr. 1995, 18, 1595–1609. [Google Scholar] [CrossRef]
- Sarath, G.; Wagner, F.W. Bacteroids Are Stable during Dark-Induced Senescence of Soybean Root Nodules. Plant Physiol. 1986, 1986. 82, 346–350. [Google Scholar] [CrossRef]
- Khan, M.S.A.; Karim, M.A.; Haque, M.M.; Islam, M.M.; Karim, A.J.M.S.; Mian, M.A.K. Influence of salt and water stress on growth and yield of soybean genotypes. Trop. Agric. Sci. 2016, 39, 167–180. [Google Scholar]
- Ghassemi-Golezani, K.; Taifeh-Noori, M.; Oustan, S. Oil and protein accumulation in soybean grains under salinity stress. Not. Sci. Biologicae 2010, 2, 64–67. [Google Scholar] [CrossRef]
- Kumar, S.; Promila, K. Effects of chloride and sulfate types of salinization and desalinization on nodulation and nitrogenfixation in chickpea. Indian J. Plant Physiol. 1983, 26, 396–401. [Google Scholar]
- Vadez, V.; Krishnamurthy, L.; Serraj, R.; Gaur, P.M.; Upadhyaya, H.D.; Hoisington, D.A.; Varshney, R.K.; Turner, N.C.; Siddique, K.H.M. Large variation in salinity tolerance in chickpea is explained by differences in sensitivity at the reproductive stage. Field Crop. Res. 2007, 104, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S. Effect of soil salinity on the yield and yield components of mungbean. Pak. J. Bot. 2009, 41, 263–268. [Google Scholar]
- Farooq, M.; Gogoi, N.; Hussain, M.; Barthakur, S.; Paul, S.; Bharadwaj, N.; Migdadi, H.M.; Alghamdi, S.S.; Siddique, K.H.M. Effects, tolerance mechanisms and management of salt stress in grain legumes. Plant Physiol. Biochem. 2017, 118, 199–217. [Google Scholar] [CrossRef]
- Qados, A.M.S.A. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15. [Google Scholar] [CrossRef]
- Qados, A.M.S.A. Effect of arginine on growth, nutrient composition, yield and nutritional value of mungbean plants grown under salinity stress. Nature. 2010, 8, 30–42. [Google Scholar]
- Katerji, N.; Hamdy, A.; Mastrorilli, M. Salt tolerance classification of crops according to soil salinity and to water stress day index. Agric. Water Manag. 2000, 43, 99–109. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Y.F.; Wong, F.L.; Tsai, S.N.; Phang, T.H.; Shao, G.; Lam, H.M. Tonoplast-located GmCLC1 and GmNHX1 from soybean enhance NaCl tolerance in transgenic bright yellow (BY)-2 cells. Plant Cell Environ. 2006, 29, 1122–1137. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Johansen, C.; Jana, M.K.; Kumar Rao, J.V.D.K. Effects of the sodium/calcium ratio in modifying salinity response of pigeonpea (Cajanus cajan). J. Plant Physiol. 1990, 136, 439–443. [Google Scholar] [CrossRef]
- Waheed, A.; Hafiz, I.A.; Qadir, G.; Murtaza, G.; Mahmood, T.; Ashraf, M. Effect of salinity on germination, growth, yield, ionic balance and solute composition of Pigeon pea (Cajanus cajan (L.) Millsp). Pak. J. Bot. 2006, 38, 1103–1117. [Google Scholar] [CrossRef]
- Turner, N.C.; Colmer, T.D.; Quealy, J.; Pushpavalli, R.; Krishnamurthy, L.; Kaur, J.; Singh, G.; Siddique, K.H.M.; Vadez, V. Salinity tolerance and ion accumulation in chickpea (Cicer arietinum L.) subjected to salt stress. Plant Soil 2013, 365, 347–361. [Google Scholar] [CrossRef]
- Phang, T.H.; Shao, G.; Lam, H.M. Salt tolerance in soybean. J. Integr. Plant Biol. 2008, 50, 1196–1212. [Google Scholar] [CrossRef] [PubMed]
- Lacan, D.; Durand, M. Na+-K+ exchange at the xylem/symplast boundary (Its significance in the salt sensitivity of soybean). Plant Physiol. 1996, 110, 705–711. [Google Scholar] [CrossRef]
- Yu, B.J.; Lam, H.M.; Shao, G.H.; Liu, Y.L. Effects of salinity on activities of H+ -ATPase, H+-PPase and membrane lipid composition in plasma membrane and tonoplast vesicles isolated from soybean (Glycine max L.) seedlings. J. Environ. Sci. 2005, 17, 259–262. [Google Scholar]
- Sanders, D. Plant biology: The salty tale of Arabidopsis. Curr. Biol. 2000, 10, 486–488. [Google Scholar] [CrossRef]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef]
- Liu, J. The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc. Natl. Acad. Sci. USA 2000, 97, 3730–3734. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, M. SOS3 function in plant salt tolerance requires N-myristoylation and calcium binding. Plant Cell Online 2000, 12, 1667–1678. [Google Scholar] [CrossRef]
- Garg, N.; Noor, Z. Genotypic differences in plant growth, osmotic and antioxidative defence of Cajanus cajan (L.) Millsp. modulated by salt stress. Arch. Agron. Soil Sci. 2009, 55, 3–33. [Google Scholar] [CrossRef]
- Panda, M.D.K. Salt stress induced changes in growth and enzyme activities in germinating Phaseolus mungo seeds. Biol. Plant. 2001, 44, 587–589. [Google Scholar] [CrossRef]
- Sumithra, K.; Jutur, P.P.; Carmel, B.D.; Reddy, A.R. Salinity-induced changes in two cultivars of Vigna radiata: Responses of antioxidative and proline metabolism. Plant Growth Regul. 2006, 50, 11–22. [Google Scholar] [CrossRef]
- Qurashi, A.W.; Sabri, A.N. Osmolyte accumulation in moderately halophilic bacteria improves salt tolerance of chickpea. Pak. J. Bot. 2013, 45, 1011–1016. [Google Scholar]
- El Sabagh, A.; Sorour, S.; Omar, A.E.; Ragab, A.; Islam, M.S.; Ueda, A.; Saneoka, H. Alleviation of adverse effects of salt stress on soybean (Glycine max. L.) by using osmoprotectants and organic nutrients. Int. J. Innov. Res. Sci. Eng. 2015, 9, 921–925. [Google Scholar]
- Enrique Olmos, E.H. Mechanisms of salt tolerance in a cell line of Pisum sativum. Plant Sci. 1996, 120, 37–45. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Campillo, A.; Jimenez, A.; Alarcon, J.J.; Sevilla, F. Response of antioxidant systems and leaf water relations to NaCl stress in pea plants. New Phytol. 1999, 141, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Kukreja, S.; Nandwal, A.S.; Kumar, N.; Sharma, S.K.; Sharma, S.K.; Unvi, V.; Sharma, P.K. Plant water status, H2O2 scavenging enzymes, ethylene evolution and membrane integrity of Cicer arietinum roots as affected by salinity. Biol. Plant. 2005, 49, 305–308. [Google Scholar] [CrossRef]
- Hernández, J.A.; Jiménez, A.; Mullineaux, P.; Sevilla, F. Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ. 2000, 23, 853–862. [Google Scholar] [CrossRef]
- Yasar, F.; Ellialtioglu, S.; Yildiz, K. Effect of salt stress on antioxidant defense systems, lipid peroxidation, and chlorophyll content in green bean. Russ. J. Plant Physiol. 2008, 55, 782–786. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic acid signaling and abiotic stress tolerance in plants: A review on current knowledge and future prospects. Front. Plant Sci. 2017, 08, 1–12. [Google Scholar] [CrossRef]
- Singh, N.K.; Larosa, P.C.; Handa, A.K.; Hasegawa, P.M.; Bressan, R.A. Hormonal regulation of protein synthesis associated with salt tolerance in plant cells. Proc. Natl. Acad. Sci. USA 1987, 84, 739–743. [Google Scholar] [CrossRef] [Green Version]
- Wolf, O.; Jeschke, W.D. Long distance transport of abscisic acid in NaCl-treated intact plants of Lupinus albus. J. Exp. Bot. 1990, 5, 593–600. [Google Scholar] [CrossRef]
- Grattan, S.R.; Grieve, C.M. Salinity-mineral nutrient relations in horticultural crops. Sci. Hortic. (Amst.) 1998, 78, 127–157. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Pandey, S.K.; Pathak, L.P.; Pathak, R.K. Effect of some nutrients in rice plant under sodic soils. Int. J. Tech. Res. Appl. 2013, 1, 9–18. [Google Scholar]
- Ezeaku, P.; Ene, J.; Shehu, J. Application of different reclamation methods on salt affected soils for crop production. Am. J. Exp. Agric. 2015, 9, 1–11. [Google Scholar] [CrossRef]
- Hussain, Z.; Khattak, R.A.; Irshad, M.; Mahmood, Q.; An, P. Effect of saline irrigation water on the leachability of salts, growth and chemical composition of wheat (Triticum aestivum L.) in saline-sodic soil supplemented with phosphorus and potassium. J. Soil Sci. Plant Nutr. 2016, 16, 604–620. [Google Scholar] [CrossRef]
- Ma, T.; Zeng, W.; Li, Q.; Wu, J.; Huang, J. Effects of water, salt and nitrogen stress on sunflower (Helianthus annuus L.) at different growth stages. J. Soil Sci. Plant Nutr. 2016, 16, 1024–1037. [Google Scholar] [CrossRef]
- Hussain, N.; Hassan, G.; Arshadullah, M.; Mujeeb, F. Evaluation of amendments for the improvement of physical properties of sodic soil. Int. J. Agric. Biol. 2001, 3, 319–322. [Google Scholar]
- Manzoor, A.; Khattak, R.A.; Dost, M. Humic acid and micronutrient effects on wheat yield and nutrients uptake in salt affected soils. Int. J. Agric. Biol. 2014, 16, 991–995. [Google Scholar]
- Matuszak-Slamani, R.; Bejger, R.; Cieśla, J.; Bieganowski, A.; Koczańska, M.; Gawlik, A.; Kulpa, D.; Sienkiewicz, M.; Włodarczyk, M.; Gołębiowska, D. Influence of humic acid molecular fractions on growth and development of soybean seedlings under salt stress. Plant Growth Regul. 2017, 83, 465–477. [Google Scholar] [CrossRef]
- Lawson, I.Y.D.; Hayatsu, M.; Nioh, I. Effects of compost application on growth and nodulation of kidney bean, soybean and alfalfa under salt stress. West Afr. J. Appl. Ecol. 2004, 5, 1–9. [Google Scholar] [CrossRef]
- Sun, H.; Lu, H.; Chu, L.; Shao, H.; Shi, W. Biochar applied with appropriate rates can reduce N leaching, keep N retention and not increase NH3 volatilization in a coastal saline soil. Sci. Total Environ. 2017, 575, 820–825. [Google Scholar] [CrossRef]
- EL Sabagh, A.; Sobhy, S.; Akram, M.; Mohammad, S.I.; Akihiro, U.; Celaleddin, B.; Halis, A.; Hirofumi, S. Role of osmoprotectants and compost application in improving water stress tolerance insoybean (Glycine max L.). Int. J. Curr. Res. 2016, 8, 25949–25954. [Google Scholar]
- Matijeví, L.; Romí, D.; Romí, M.; Mauroví, N.; Kondres, N. Faba bean (Vicia faba L.) salt stress response under different soil organic matter content. Agric. Conspec. Sci. 2014, 79, 13–18. [Google Scholar]
- Garg, N.; Chandel, S. Effect of mycorrhizal inoculation on growth, nitrogen fixation, and nutrient uptake in Cicer arietinum (L.) under salt stress. Turk. J. Agric. For. 2011, 35, 205–214. [Google Scholar] [CrossRef]
- Kabir, M.E.; Karim, M.A.; Azad, M.A.K. Effect of potassium on salinity tolerance of mungbean (Vigana radiata L. Wilczek). J. Biol. Sci. 2004, 4, 103–110. [Google Scholar]
- Guo, J.S.; Zhou, Q.; Li, X.J.; Yu, B.J.; Luo, Q.Y. Enhancing NO3- supply confers NaCl tolerance by adjusting Cl-uptake and transport in G. max & G. soja. J. Soil Sci. Plant Nutr. 2017, 17, 194–204. [Google Scholar]
- El-Nour, A.A.A. Can supplemented potassium foliar feeding reduce the recommended soil potassium. Pak. J. Biol. Sci. 2002, 5, 259–262. [Google Scholar] [CrossRef]
- Abou-El-Nour, E.Z.A.; Aly, E.M.; El-Fouly, M.M.; Salama, Z.A.E.R. Chelated Fe and Zn foliar spray improve the tolerance of kidney bean (var. nebraska) plants in salinized media. Biosci. Res. 2017, 14, 525–531. [Google Scholar]
- Zuccarini, P. Effects of silicon on photosynthesis, water relations and nutrient uptake of Phaseolus vulgaris under NaCl stress. Biol. Plant. 2008, 52, 157–160. [Google Scholar] [CrossRef]
- Kardoni, F.; Jalil, S.; Seyyed Mosavi, S.J.; Parande, S. Effect of salinity stress and silicon application on yield and component yield offaba bean (Vicia faba). Int. J. Agric. Crop Sci. 2013, 6, 814–818. [Google Scholar]
- Murtaza, G.; Ghafoor, A.; Qadir, M. Irrigation and soil management strategies for using saline-sodic water in a cotton-wheat rotation. Agric. Water Manag. 2006, 81, 98–114. [Google Scholar] [CrossRef]
- Yamika, W.S.D.; Aini, N.; Setiawan, A.; Purwaningrahayu, R.D. Effect of gypsum and cow manure on yield, proline content, and K/Na ratio of soybean genotypes under saline conditions. J. Degrad. Min. Lands Manag. 2018, 5, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.; Zhang, J.; Zhong, C.; Zhu, L.; Cao, X.; Yu, S.; Allen, B.J.; Hu, J.; Jin, Q. Effects of salt stress on rice growth, development characteristics, and the regulating ways: A review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Abdel-Fattah, M.K. Role of gypsum and compost in reclaiming saline-sodic soils. IOSR J. Agric. Vet. Sci. 2012, 1, 30–38. [Google Scholar] [CrossRef]
- Thakuria, D.; Hazarika, S.; Krishnappa, R. Soil acidity and management options. Indian J. Fertil. 2016, 12, 40–56. [Google Scholar]
- Han, H.; Lee, S. Physiological responses of soybean inoculation of Bradyrhizobium japonicum with PGPR in saline soil conditions. Res. J. Agric. Biol. Sci. 2005, 1, 216–221. [Google Scholar]
- Egamberdieva, D. Survival of Pseudomonas extremorientalis TSAU20 and P. chlororaphis TSAU13 in the rhizosphere of common bean (Phaseolus vulgaris) under saline conditions. Plant Soil Environ. 2011, 57, 122–127. [Google Scholar] [CrossRef]
- Egamberdiyeva, D.; Höflich, G. Root colonization and growth promotion of winter wheat and pea by Cellulomonas spp. at different temperatures. Plant Growth Regul. 2002, 38, 219–224. [Google Scholar] [CrossRef]
- Afolayan, E.T.; Eguavon, M.I. Morphological and yield assessment of soybean (Glycine max L.) as influenced by arbuscular mycorrhizal fungi and other soil amendments. J. Agric. Sci. Food Res. 2017, 8, 1–5. [Google Scholar]
- Metwali, E.M.R.; Abdelmoneim, T.S.; Bakheit, M.A.; Kadasa, N.M.S. Alleviation of salinity stress in faba bean (Vicia faba L.) plants by inoculation with plant growth promoting rhizobacteria (PGPR). Plant Omics 2015, 8, 449–460. [Google Scholar]
- Chaudhary, D.; Sindhu, S. Inducing salinity tolerance in chickpea (Cicer arietinum L.) by inoculation of 1-aminocyclopropane-1-carboxylic acid deaminase-containing Mesorhizobium strains. Afr. J. Microbiol. Res. 2015, 9, 117–124. [Google Scholar] [CrossRef]
- Valverde, A.; Velázquez, E.; Fernández-Santos, F.; Vizcaíno, N.; Mateos, P.F.; Martínez-Molina, E.; Igual, J.M.; Willems, A. Phyllobacterium trifolii sp. nov., nodulating Trifolium and Lupinas in Spanish soils. Int. J. Syst. Evol. Microbiol. 2005, 55, 1985–1989. [Google Scholar] [CrossRef]
- Yadegari, M.; Rahmani, H.A.; Noormohammadi, G.; Ayneband, A. Evaluation of bean (Phaseolus vulgaris) seeds inoculation with Rhizobium phaseoli and plant growth promoting rhizobacteria on yield and yield components. Pak. J. Biol. Sci. 2010, 5, 792–799. [Google Scholar] [CrossRef]
- Ahmad, M.; Zahir, Z.; Asghar, H.; Arshad, M. The combined application of rhizobial strains and plant growth promoting rhizobacteria improves growth and productivity of mung bean (Vigna radiata L.) under salt-stressed conditions. Ann. Microbiol. 2012, 62, 1321–1330. [Google Scholar] [CrossRef]
- Ahmad, M.; Zahir, Z.A.; Asghar, H.N.; Asghar, M. Inducing salt tolerance in mung bean through coinoculation with Rhizobia and plant-growth-promoting rhizobacteria containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2011, 57, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Zahir, Z.A.; Khalid, M.; Nazli, F.; Arshad, M. Efficacy of Rhizobium and Pseudomonas strains to improve physiology, ionic balance and quality of mungbean under salt-affected conditions on farmer’s fields. Plant Physiol. Biochem. 2013, 63, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Penrose, D.M.; Moffatt, B.A.; Glick, B.R. Determination of 1-aminocycopropane-1-carboxylic acid (ACC) to assess the effects of ACC deaminase-containing bacteria on roots of canola seedlings. Can. J. Microbiol. 2001, 47, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Pliego, C.; Kamilova, F.; Lugtenberg, B. Plant Growth-Promoting Bacteria: Fundamentals and Exploitation. In Bacteria in Agrobiology: Crop Ecosystems; Maheshwari, D.K., Ed.; Springer: Heidelberg/Berlin, Germany, 2011; ISBN 978-3-642-18356-0. [Google Scholar]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Khan, F.; Hussain, H.A.; Nie, L. Physiological and biochemical mechanisms of seed priming-induced chilling tolerance in rice cultivars. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Dai, L.-Y.; Zhu, H.-D.; Yin, K.-D.; Du, J.-D.; Zhang, Y.-X. Seed priming mitigates the effects of saline-alkali stress in soybean seedlings. Chil. J. Agric. Res. 2017. [Google Scholar] [CrossRef]
- Harris, D.; Rashid, A.; Miraj, G. ‘On-farm’ seed priming with zinc in chickpea and wheat in Pakistan ‘On-farm’ seed priming with zinc in chickpea and wheat in Pakistan. Plant Soil 2007. [Google Scholar] [CrossRef]
- Younesi, O.; Moradi, A. Effect of priming of seeds of m edicago sativa ‘bami’ with gibberellic acid on germination, seedlings growth and antioxidant enzymes activity under salinity stress. J. Hortic. Res. 2014, 22, 167–174. [Google Scholar] [CrossRef]
- Azooz, M.M. Salt Stress Mitigation by seed priming with salicylic acid in two faba bean salt stress mitigation by seed priming with salicylic acid in two faba bean genotypes differing in salt tolerance. Int. J. Agric. Biol. 2009, 11, 343–350. [Google Scholar]
- Jisha, K.C.; Puthur, J.T. Halopriming of seeds imparts tolerance to NaCl and PEG induced stress in (Vigna radiata L.) Wilczek varieties. Physiol. Mol. Biol. Plants 2014, 20, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, N.; Yousaf, S.; Jamil, F.F. Induction of salt tolerance in chickpea by using simple and safe chemicals. Pak. J. Bot. 2006, 38, 325–329. [Google Scholar]
- Ahmadvand, G.; Soleimani, F.; Saadatian, B.; Pouya, M. Effects of seed priming on germination and emergence traits of two soybean cultivars under salinity stress. J. Basic Appl. Sci. Res. 2012, 3, 234–241. [Google Scholar]
- Khomari, S.; Golshan-doust, S.; Seyed-sharifi, R.; Davari, M. Improvement of soybean seedling growth under salinity stress by biopriming of high-vigour seeds with salt-tolerant isolate of Trichoderma harzianum. N. Z. J. Crop Hortic. Sci. 2017, 00, 1–16. [Google Scholar] [CrossRef]
- Abdolahpour, M.; Lotfi, R. Seed priming affected physiology and grain yield of chickpea under salt stress. J. Biodivers. Environ. Sci. 2014, 5, 442–446. [Google Scholar]
- Hussain, S.S.; Ali, M.; Ahmad, M.; Siddique, K.H.M. Polyamines: Natural and engineered abiotic and biotic stress tolerance in plants. Biotechnol. Adv. 2011, 29, 300–311. [Google Scholar] [CrossRef]
- Berberich, T.K.T.; Takahashi, C.T.Y. Polyamines: Essential factors for growth and survival. Planta 2008, 228, 367–381. [Google Scholar] [CrossRef]
- Gupta, K.; Dey, A.; Gupta, B.; Bengal, W. Plant polyamines in abiotic stress responses. Acta Physiol. Plant. 2013, 35, 2015–2036. [Google Scholar] [CrossRef]
- Silveira, V.; Martins, A.; Ferreira, A.; Maria, M.; Ribeiro, F.; Iochevet, E.; Floh, S. Morphological and polyamine content changes in embryogenic and non-embryogenic callus of sugarcane. Plant Cell Tissue Organ Cult. 2013, 114, 351–364. [Google Scholar] [CrossRef] [Green Version]
- Gomez-jimenez, J.A.G.M.C. Regulation of polyamine metabolism and biosynthetic gene expression during Regulation of polyamine metabolism and biosynthetic gene expression during olive mature-fruit abscission. Planta 2012, 235, 1221–1237. [Google Scholar] [CrossRef]
- Kim, N.H.; Kim, B.S.; Hwang, B.K. Pepper arginine decarboxylase is required for polyamine and g-Aminobutyric acid signaling in cell death and defense response. Plant Physiol. 2013, 162, 2067–2083. [Google Scholar] [CrossRef] [PubMed]
- Tisi, A.; Federico, R.; Moreno, S.; Lucretti, S.; Moschou, P.N.; Roubelakis-Angelakis, K.A.; Angelini, R.; Cona, A. Perturbation of polyamine catabolism can strongly affect root development and xylem differentiation. Plant Physiol. 2011, 157, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-W.; Xu, S.-C.; Hu, Q.-Z.; Mao, W.-H.; Gong, Y.-M. Putrescine plays a positive role in salt-tolerance mechanisms by reducing oxidative damage in roots of vegetable soybean. J. Integr. Agric. 2014, 13, 349–357. [Google Scholar] [CrossRef]
- Sheokand, S.; Kumari, A.; Sawhney, V. Effect of nitric oxide and putrescine on antioxidative responses under NaCl stress in chickpea plants. Physiol. Mol. Biol. Plants 2008, 14, 355–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suleiman, S. Effects of exogenous application of diamine (Putrescine) on growth and mineral elements distribution in faba bean plants under saline conditions. Biol. Sci. Ser. 2008, 2008. 30, 258–265. [Google Scholar]
- Mahdi, A.H.A. Improvement of salt tolerance in Vicia faba (L.) plants by exogenous application of polyamines. Egypt. J. Agron. 2016, 38, 1–21. [Google Scholar]
- Radhakrishnan, R.; Lee, I.-J. Spermine promotes acclimation to osmotic stress by modifying antioxidant, abscisic acid, and jasmonic acid signals in soybean. J. Plant Growth Regul. 2012, 32, 22–30. [Google Scholar] [CrossRef]
- Lou, Y.; Guan, R.; Sun, M.; Han, F.; He, W.; Wang, H.; Song, F.; Cui, X.; Zhuge, Y. Spermidine application alleviates salinity damage to antioxidant enzyme activity and gene expression in alfalfa. Ecotoxicology 2018. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Rahman, A.; Alam, M. Polyamines confer salt tolerance in mungbean (Vigna radiata L.) by reducing sodium uptake, improving nutrient homeostasis, antioxidant defense, and methylglyoxal detoxification systems. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Ghosh, S.; Mitra, S.; Paul, A. Physiochemical studies of sodium chloride on mungbean (Vigna radiata L. Wilczek) and its possible recovery with spermine and gibberellic acid. Sci. World J. 2015, 2015. [Google Scholar] [CrossRef]
- Smýkal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Berger, J.; Greene, S.L.; Nelson, M.N.; Besharat, N.; et al. Legume crops phylogeny and genetic diversity for science and breeding. Crit. Rev. Plant Sci. 2015, 34, 43–104. [Google Scholar] [CrossRef]
- Duc, G.; Agrama, H.; Bao, S.; Berger, J.; Bourion, V.; De Ron, A.M.; Gowda, C.L.L.; Mikic, A.; Millot, D.; Singh, K.B.; et al. Breeding annual grain legumes for sustainable agriculture: New methods to approach complex traits and target new cultivar ideotypes. Crit. Rev. Plant Sci. 2015, 34, 381–411. [Google Scholar] [CrossRef]
- Sehrawat, N.; Bhat, K.V.; Kaga, A.; Tomooka, N.; Yadav, M.; Jaiwal, P.K. Development of new gene-specific markers associated with salt tolerance for mungbean (Vigna radiata L. Wilczek). Span. J. Agric. Res. 2014, 12, 732–741. [Google Scholar] [CrossRef]
- Sharma, S.; Upadhyaya, H.D.; Varshney, R.K.; Gowda, C.L.L. Pre-breeding for diversification of primary gene pool and genetic enhancement of grain legumes. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foolad, M.R. Recent advances in genetics of salt tolerance in tomato. Plant Cell Tissue Organ Cult. 2004, 76, 101–119. [Google Scholar] [CrossRef]
- Dita, M.A.; Rispail, N.; Prats, E.; Rubiales, D.; Singh, K.B. Biotechnology approaches to overcome biotic and abiotic stress constraints in legumes. Euphytica 2006, 147, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.J.; Boerma, H.R.; Villagarcia, M.R.; Zhou, X.; Carter, T.E.; Li, Z.; Gibbs, M.O. A major QTL conditioning salt tolerance in S-100 soybean and descendent cultivars. Theor. Appl. Genet. 2004, 109, 1610–1619. [Google Scholar] [CrossRef]
- Vadez, V.; Krishnamurthy, L.; Thudi, M.; Anuradha, C.; Timothy, D.; Turner, N.C.; Siddique, K.H.M.; Gaur, P.M.; Varshney, R.K.; Vadez, V.; et al. Assessment of ICCV 2 × JG 62 chickpea progenies shows sensitivity of reproduction to salt stress and reveals QTL for seed yield and yield components. Mol. Breed. 2012, 30, 9–21. [Google Scholar] [CrossRef]
- Leonforte, A.; Sudheesh, S.; Cogan, N.O.I.; Salisbury, P.A.; Nicolas, M.E.; Materne, M.; Forster, J.W.; Kaur, S. SNP marker discovery, linkage map construction and identification of QTL for enhanced salinity tolerance in field pea (Pisum sativum L.). BMC Plant Biol. 2013, 13, 161. [Google Scholar] [CrossRef]
- Samineni, S. Genetics and QTL Mapping of Salt Tolerance in Chickpea (Cicer arietinum L.). Ph.D. Thesis, University of Western Australia, Perth, Australia, 2010. [Google Scholar]
- Shi, X.; Yan, L.; Yang, C.; Yan, W.; Moseley, D.O.; Wang, T.; Liu, B.; Di, R.; Chen, P.; Zhang, M. Identification of a major quantitative trait locus underlying salt tolerance in ‘Jidou 12′ soybean cultivar. BMC Res. Notes 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chankaew, S.; Isemura, T.; Naito, K.; Kaga, A.; Vaughan, D.A.; Srinives, P. QTL mapping for salt tolerance and domestication related traits in Vigna marina subsp. oblonga, a halophytic species. Theor. Appl. Genet. 2014, 127, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Crop breeding for salt tolerance in the era of molecular markers and marker-assisted selection. Plant Breed. 2012, 132, 10–20. [Google Scholar] [CrossRef]
- Guan, R.; Chen, J.; Jiang, J.; Liu, G.; Liu, Y.; Tian, L.; Yu, L.; Chang, R.; Qiu, L.-J. Mapping and validation of a dominant salt tolerance gene in the cultivated soybean (Glycine max L.) variety Tiefeng 8. Crop J. 2014, 2, 358–365. [Google Scholar] [CrossRef]
- Hamwieh, A.; Tuyen, D.D.; Cong, H.; Benitez, E.R.; Takahashi, R.; Xu, D.H. Identification and validation of a major QTL for salt tolerance in soybean. Euphytica 2011, 179, 451–459. [Google Scholar] [CrossRef]
- Kumar, J.; Choudhary, A.K.; Solanki, R.K.; Pratap, A. Towards marker-assisted selection in pulses: A review. Plant Breed. 2011, 130, 297–313. [Google Scholar] [CrossRef]
- Pandiyan, M.; Senthil, N.; Packiaraj, D.; Gupta, S.; Nadarajan, N.; Pandian, R.T.; Suresh, R.; Jagadeesh, S. Characterisation and evaluation of 646 greengram (Vigna radiata) genotypes for constituting core collection. Wudpecker J. Agric. Res. 2012, 1, 294–301. [Google Scholar]
- Nirmala, S.; Mukesh, Y.; Venkataraman, B.K.; Kumar, S.R.; Kumar, J.P. Hybridization between salt resistant and salt susceptible genotypes of mungbean (Vigna radiata L. Wilczek) and purity testing of the hybrids using SSRs markers. J. Integr. Agric. 2016, 15, 521–527. [Google Scholar] [CrossRef]
- Covarrubias, A.A.; Reyes, J.L. Post-transcriptional gene regulation of salinity and drought responses by plant microRNAs. Plant Cell Environ. 2010, 33, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Shi, L.; Wang, Y.; Chen, L.; Cai, Z.; Wang, Y.; Jin, J.; Li, X. Identification and dynamic regulation of microRNAs involved in salt stress responses in functional soybean nodules by high-throughput sequencing. Int. J. Mol. Sci. 2013, 14, 2717–2738. [Google Scholar] [CrossRef]
- Liao, Y.; Zou, H.F.; Wang, H.W.; Zhang, W.K.; Ma, B.; Zhang, J.S.; Chen, S.Y. Soybean GmMYB76, GmMYB92, and GmMYB177 genes confer stress tolerance in transgenic Arabidopsis plants. Cell Res. 2008, 18, 1047–1060. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, L.; Qu, Y.; Chen, J.; Liu, X.; Hong, H.; Liu, Z.; Chang, R.; Gilliham, M.; Qiu, L.; et al. GmSALT3, which confers improved soybean salt tolerance in the field, increases leaf Cl- exclusion prior to Na+ exclusion but does not improve early vigor under salinity. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Guan, R.; Qu, Y.; Guo, Y.; Yu, L.; Liu, Y.; Jiang, J.; Chen, J.; Ren, Y.; Liu, G.; Tian, L.; et al. Salinity tolerance in soybean is modulated by natural variation in GmSALT3. Plant J. 2014, 80, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.C.; Xu, J.Y.; Wang, C.; Guo, N.; Hou, J.F.; Xue, D.; Zhao, J.M.; Xing, H. Molecular cloning and functional characterization of a soybean GmGMP1 gene reveals its involvement in ascorbic acid biosynthesis and multiple abiotic stress tolerance in transgenic plants. J. Integr. Agric. 2018, 17, 539–553. [Google Scholar] [CrossRef]
- Chen, Y.; Chi, Y.; Meng, Q.; Wang, X.; Yu, D. GmSK1, an SKP1 homologue in soybean, is involved in the tolerance to salt and drought. Plant Physiol. Biochem. 2018, 127, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hu, Z.; Jiang, Q.Y.; Sun, X.J.; Guo, Y.; Qi, J.C.; Zhang, H. GmNAC15 overexpression in hairy roots enhances salt tolerance in soybean. J. Integr. Agric. 2018, 17, 530–538. [Google Scholar] [CrossRef]
- An, J.; Cheng, C.; Hu, Z.; Chen, H.; Cai, W.; Yu, B. The Panax ginseng PgTIP1 gene confers enhanced salt and drought tolerance to transgenic soybean plants by maintaining homeostasis of water, salt ions and ROS. Environ. Exp. Bot. 2018, 155, 45–55. [Google Scholar] [CrossRef]
- Wang, L.S.; Chen, Q.S.; Xin, D.W.; Qi, Z.M.; Zhang, C.; Li, S.N.; Jin, Y.M.; Li, M.; Mei, H.Y.; Su, A.Y.; et al. Overexpression of GmBIN2, a soybean glycogen synthase kinase 3 gene, enhances tolerance to salt and drought in transgenic Arabidopsis and soybean hairy roots. J. Integr. Agric. 2018, 17, 1959–1971. [Google Scholar] [CrossRef]
- Pruthvi, V.; Narasimhan, R.; Nataraja, K.N. Simultaneous expression of abiotic stress responsive transcription factors, AtDREB2A, AtHB7 and AtABF3 improves salinity and drought tolerance in peanut (Arachis hypogaea L.). PLoS ONE 2014, 9, 1–21. [Google Scholar] [CrossRef]
- Lopez, C.M.L.; Takahashi, H.; Yamazaki, S. Plant-water relations of kidney bean plants treated with NaCl and foliarly applied glycinebetaine. J. Agron. Crop Sci. 2002, 188, 73–80. [Google Scholar] [CrossRef]
- Frugier, F.; Poirier, S.; Satiat-Jeunemaître, B.; Kondorosi, A.; Crespi, M. A Kruppel-like zinc finger protein is involved in nitrogen-fixing root nodule organogenesis. Genes Dev. 2000, 14, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Hanafy, M.S.; El-Banna, A.; Schumacher, H.M.; Jacobsen, H.J.; Hassan, F.S. Enhanced tolerance to drought and salt stresses in transgenic faba bean (Vicia faba L.) plants by heterologous expression of the PR10a gene from potato. Plant Cell Rep. 2013, 32, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Liu, J.; Tang, L.; Cai, H.; Chen, M.; Ji, W.; Liu, Y.; Zhu, Y. Overexpression of GsCBRLK from Glycine soja enhances tolerance to salt stress in transgenic alfalfa (Medicago sativa). Funct. Plant Biol. 2013, 40, 1048–1056. [Google Scholar] [CrossRef]
- Ji, W.; Koh, J.; Li, S.; Zhu, N.; Dufresne, C.P.; Zhao, X.; Chen, S.; Li, J. Quantitative proteomics reveals an important role of GsCBRLK in salt stress response of soybean. Plant Soil 2016, 402, 159–178. [Google Scholar] [CrossRef]
- Cao, D.; Hou, W.; Liu, W.; Yao, W.; Wu, C.; Liu, X.; Han, T. Overexpression of TaNHX2 enhances salt tolerance of “composite” and whole transgenic soybean plants. Plant Cell. Tissue Organ Cult. 2011, 107, 541–552. [Google Scholar] [CrossRef]
- Bao, A.K.; Wang, S.M.; Wu, G.Q.; Xi, J.J.; Zhang, J.L.; Wang, C.M. Overexpression of the Arabidopsis H+-PPase enhanced resistance to salt and drought stress in transgenic alfalfa (Medicago sativa L.). Plant Sci. 2009, 176, 232–240. [Google Scholar] [CrossRef]
- De Zélicourt, A.; Diet, A.; Marion, J.; Laffont, C.; Ariel, F.; Moison, M.; Zahaf, O.; Crespi, M.; Gruber, V.; Frugier, F. Dual involvement of a Medicago truncatula NAC transcription factor in root abiotic stress response and symbiotic nodule senescence. Plant J. 2012, 70, 220–230. [Google Scholar] [CrossRef]
- Tripathi, P.; Rabara, R.C.; Rushton, P.J. A systems biology perspective on the role of WRKY transcription factors in drought responses in plants. Planta 2014, 239, 255–266. [Google Scholar] [CrossRef]
- Yu, X.; Liu, Y.; Wang, S.; Tao, Y.; Wang, Z.; Shu, Y.; Peng, H.; Mijiti, A.; Wang, Z.; Zhang, H.; et al. CarNAC4, a NAC-type chickpea transcription factor conferring enhanced drought and salt stress tolerances in Arabidopsis. Plant Cell Rep. 2016, 35, 613–627. [Google Scholar] [CrossRef]
- Sarkar, T.; Thankappan, R.; Kumar, A.; Mishra, G.P. Heterologous expression of the AtDREB1A gene in transgenic peanut-conferred tolerance to drought and salinity stresses. PLoS ONE 2014, 9, 1–25. [Google Scholar] [CrossRef]
- Khatib, F.; Makris, A.; Yamaguchi-Shinozaki, K.; Kumar, S.; Sarker, A.; Erskine, W.; Baum, M. Expression of the DREB1A gene in lentil (Lens culinaris Medik. subsp. culinaris) transformed with the Agrobacterium system. Crop Pasture Sci. 2011, 62, 488–495. [Google Scholar] [CrossRef]
- Singh, R.J.; Chung, G.H.; Nelson, R.L. Landmark research in legumes. Genome 2007, 50, 525–537. [Google Scholar] [CrossRef]
- Zuryn, S.; Le Gras, S.; Jamet, K.; Jarriault, S. A strategy for direct mapping and identification of mutations by whole-genome sequencing. Genetics 2010, 186, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Kudapa, H.; Ramalingam, A.; Nayakoti, S.; Chen, X.; Zhuang, W.J.; Liang, X.; Kahl, G.; Edwards, D.; Varshney, R.K. Functional genomics to study stress responses in crop legumes: Progress and prospects. Funct. Plant Biol. 2013, 40, 1221–1233. [Google Scholar] [CrossRef]
- O’Rourke, J.A.; Bolon, Y.T.; Bucciarelli, B.; Vance, C.P. Legume genomics: Understanding biology through DNA and RNA sequencing. Ann. Bot. 2014, 113, 1107–1120. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.K.; Roorkiwal, M.; Singh, V.K.; Ramalingam, A.; Kudapa, H.; Thudi, M.; Chitikineni, A.; Rathore, A.; Varshney, R.K. Emerging genomic tools for legume breeding: Current status and future prospects. Front. Plant Sci. 2016, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.C.; Zhu, W.L.; Gai, J.Y.; Zhu, Y.L.; Yang, L.F. Enhanced salt tolerance of transgenic vegetable soybeans resulting from overexpression of a novel Δ1-pyrroline-5-carboxylate synthetase gene from Solanum torvum Swartz. Hortic. Environ. Biotechnol. 2015, 56, 94–104. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, L.; Chen, J.; Tao, L.; An, Y.; Cai, H.; Guo, C. Overexpression of the alfalfa WRKY11 gene enhances salt tolerance in soybean. PLoS ONE 2018, 13, 1–16. [Google Scholar] [CrossRef]
- Ali, Z.; Ullah, N.; Naseem, S.; Inam-Ul-Haq, M.; Jacobsen, H.J. Soil bacteria conferred a positive relationship and improved salt stress tolerance in transgenic pea (Pisum sativum L.) harboring Na+/H+ antiporter. Turk. J. Bot. 2015, 39, 962–972. [Google Scholar] [CrossRef]
- Najafi, F.; Rastgar-jazii, F.; Khavari-Nejad, R.A.; Sticklen, M. Salt tolerance in transgenic pea (Pisum sativum L.) plants by P5CS gene transfer. J. Plant Biotechnol. 2005, 7, 233–240. [Google Scholar]
- Kiran Kumar Ghanti, S.; Sujata, K.G.; Vijay Kumar, B.M.; Nataraja Karba, N.; Janardhan Reddy, K.; Srinath Rao, M.; Kavi Kishor, P.B. Heterologous expression of P5CS gene in chickpea enhances salt tolerance without affecting yield. Biol. Plant. 2011, 55, 634–640. [Google Scholar] [CrossRef]
- Bhomkar, P.; Upadhyay, C.P.; Saxena, M.; Muthusamy, A.; Shiva Prakash, N.; Pooggin, M.; Hohn, T.; Sarin, N.B. Salt stress alleviation in transgenic Vigna mungo L. Hepper (blackgram) by overexpression of the glyoxalase I gene using a novel Cestrum yellow leaf curling virus (CmYLCV) promoter. Mol. Breed. 2008, 22, 169–181. [Google Scholar] [CrossRef]
- Surekha, C.; Kumari, K.N.; Aruna, L.V.; Suneetha, G.; Arundhati, A.; Kavi Kishor, P.B. Expression of the Vigna aconitifolia P5CSF129A gene in transgenic pigeonpea enhances proline accumulation and salt tolerance. Plant Cell. Tissue Organ Cult. 2014, 116, 27–36. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Vadez, V.; Jyostna Devi, M.; Lavanya, M.; Vani, G.; Sharma, K.K. Genetic engineering of chickpea (Cicer arietinum L.) with the P5CSF129A gene for osmoregulation with implications on drought tolerance. Mol. Breed. 2009, 23, 591–606. [Google Scholar] [CrossRef]
- Banjara, M.; Zhu, L.; Shen, G.; Payton, P.; Zhang, H. Expression of an Arabidopsis sodium/proton antiporter gene (AtNHX1) in peanut to improve salt tolerance. Plant Biotechnol. Rep. 2012, 6, 59–67. [Google Scholar] [CrossRef]
- Banavath, J.N.; Chakradhar, T.; Pandit, V.; Konduru, S.; Guduru, K.K.; Akila, C.S.; Podha, S.; Puli, C.O.R. Stress Inducible Overexpression of AtHDG11 leads to improved drought and salt stress tolerance in peanut (Arachis hypogaea L.). Front. Chem. 2018, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, D.; Jin, T.; Chang, Q.; Yin, D.; Xu, S.; Liu, B.; Liu, L. The Vacuolar Na+/H+ antiporter gene SsNHX1 from the halophyte salsola soda confers salt tolerance in transgenic alfalfa (Medicago sativa L.). Plant Mol. Biol. Report. 2011, 29, 278–290. [Google Scholar] [CrossRef]
- Duan, Z.; Zhang, D.; Zhang, J.; Di, H.; Wu, F.; Hu, X.; Meng, X.; Luo, K.; Zhang, J.; Wang, Y. Co-transforming bar and CsALDH genes enhanced resistance to herbicide and drought and salt stress in transgenic alfalfa (Medicago sativa L.). Front. Plant Sci. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Chang, Q.; Li, W. Stress-inducible expression of GmDREB1 conferred salt tolerance in transgenic alfalfa. Plant Cell Tissue Organ Cult. 2010, 100, 219–227. [Google Scholar] [CrossRef]
- Wang, Z.; Ke, Q.; Kim, M.D.; Kim, S.H.; Ji, C.Y. Transgenic alfalfa plants expressing the sweetpotato orange gene exhibit enhanced abiotic stress tolerance. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Liu, Z.H.; Wen, Z.Y.; Zhang, H.M.; Yang, F.; Guo, X.L. The vacuolar Na+/H+ antiport gene TaNHX2 confers salt tolerance on transgenic alfalfa (Medicago sativa L.). Funct. Plant Biol. 2012, 39, 708–716. [Google Scholar] [CrossRef]
- Liu, L.; Fan, X.D.; Wang, F.W.; Wang, N.; Dong, Y.Y.; Liu, X.M.; Yang, J.; Wang, Y.F.; Li, H.Y. Coexpression of ScNHX1 and ScVP in transgenic hybrids improves salt and saline-alkali tolerance in alfalfa (Medicago sativa L.). J. Plant Growth Regul. 2013, 32, 1–8. [Google Scholar] [CrossRef]
- Tang, L.; Cai, H.; Ji, W.; Luo, X.; Wang, Z.; Wu, J.; Wang, X.; Cui, L.; Wang, Y.; Zhu, Y.; et al. Overexpression of GsZFP1 enhances salt and drought tolerance in transgenic alfalfa (Medicago sativa L.). Plant Physiol. Biochem. 2013, 71, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Jain, M. Function genomics of abiotic stress tolerance in plants: A CRISPR approach. Front. Plant Sci. 2015, 6, 2011–2014. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Jain, M. The CRISPR-Cas system for plant genome editing: Advances and opportunities. J. Exp. Bot. 2015, 66, 47–57. [Google Scholar] [CrossRef]
- Jaganathan, D.; Ramasamy, K.; Sellamuthu, G.; Jayabalan, S.; Venkataraman, G. CRISPR for Crop Improvement: An Update Review. Front. Plant Sci. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, R.; Huang, G.; Li, Y.; Melaku, G.; Zhang, S.; Chen, H.; Zhao, Y.; Zhang, J.; Zhang, Y.; et al. Developing superior alleles of yield genes in rice by artificial mutagenesis using the CRISPR/Cas9 system. Crop J. 2018, 6, 475–481. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Sánchez-León, S.; Gil-Humanes, J.; Ozuna, C.V.; Giménez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018, 16, 902–910. [Google Scholar] [CrossRef]
- Zhu, J.; Song, N.; Sun, S.; Yang, W.; Zhao, H.; Song, W.; Lai, J. Efficiency and inheritance of targeted mutagenesis in maize using CRISPR-Cas9. J. Genet. Genom. 2016, 43, 25–36. [Google Scholar] [CrossRef]
- Agarwal, A.; Yadava, P.; Kumar, K.; Singh, I.; Kaul, T.; Pattanayak, A.; Agrawal, P.K. Insights into maize genome editing via CRISPR/Cas9. Physiol. Mol. Biol. Plants 2018, 24, 175–183. [Google Scholar] [CrossRef]
- Lawrenson, T.; Shorinola, O.; Stacey, N.; Li, C.; Østergaard, L.; Patron, N.; Uauy, C.; Harwood, W. Induction of targeted, heritable mutations in barley and Brassica oleracea using RNA-guided Cas9 nuclease. Genome Biol. 2015, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kapusi, E.; Corcuera-Gómez, M.; Melnik, S.; Stoger, E. Heritable genomic fragment deletions and small indels in the Putative ENGase gene induced by CRISPR/Cas9 in barley. Front. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Nakayasu, M.; Akiyama, R.; Lee, H.J.; Osakabe, K.; Osakabe, Y.; Watanabe, B.; Sugimoto, Y.; Umemoto, N.; Saito, K.; Muranaka, T.; et al. Generation of α-solanine-free hairy roots of potato by CRISPR/Cas9 mediated genome editing of the St16DOX gene. Plant Physiol. Biochem. 2018, 131, 70–77. [Google Scholar] [CrossRef]
- Jia, H.; Nian, W. Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, T.B.; LaFayette, P.R.; Schmitz, R.J.; Parrott, W.A. Targeted genome modifications in soybean with CRISPR/Cas9. BMC Biotechnol. 2015, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Z.-B.; Xing, A.; Moon, B.P.; Koellhoffer, J.P.; Huang, L.; Ward, R.T.; Clifton, E.; Falco, S.C.; Cigan, A.M. Cas9-Guide RNA directed genome editing in soybean. Plant Physiol. 2015, 169, 960–970. [Google Scholar] [CrossRef]
- Fan, D.; Liu, T.; Li, C.; Jiao, B.; Li, S.; Hou, Y.; Luo, K. Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, X.; Yang, C.; Li, M.; Guo, Y. Exploiting the CRISPR/Cas9 system for targeted genome mutagenesis in Petunia. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Al-Sadi, A.M.; Pour-Aboughadareh, A.; Burritt, D.J.; Tran, L.S.P. Genome editing using CRISPR/Cas9–targeted mutagenesis: An opportunity for yield improvements of crop plants grown under environmental stresses. Plant Physiol. Biochem. 2018, 131, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, M.; Bhat, J.A.; Mir, Z.A.; Sakina, A.; Ali, S.; Singh, A.K.; Tyagi, A.; Salgotra, R.K.; Dar, A.A.; Bhat, R. CRISPR/Cas approach: A new way of looking at plant-abiotic interactions. J. Plant Physiol. 2018, 224–225, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, H.; Chen, L.-L.; Xie, K. Recent advances in genome editing using CRISPR/Cas9. Front. Plant Sci. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Watson, A.; Gonzalez-Navarro, O.E.; Ramirez-Gonzalez, R.H.; Yanes, L.; Mendoza-Suárez, M.; Simmonds, J.; Wells, R.; Rayner, T.; Green, P.; et al. Speed breeding in growth chambers and glasshouses for crop breeding and model plant research. Nat. Protoc. 2018, 13, 2944–2963. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Legume Crops | Salt Concentration | Yield Loss (%) | Reference |
|---|---|---|---|
| Soybean (Galarsum) | 14.4 dSm−1 | 50% | [54] |
| Soybean (Lee) | 8.5 dSm−1 | 53% | [50] |
| Soybean (loam soil) | 7 dSm−1 | 46% | [20] |
| Soybean (caly soil) | 6.3 dSm−1 | 46% | [62] |
| Mungbean (cv. Pusavishal) | 50 mM NaCl | 41% | [32] |
| Mungbean (var. 245/7) | 8 dSm−1 | 60% | [58] |
| Mungbean (var. NM-51) | 12 dSm−1 | 77% | [58] |
| Mungbean (var. NM-92) | 8 dSm−1 | 61% | [58] |
| Mungbean (var. 6601) | 12 dSm−1 | 72% | [58] |
| Chickpea (var. FLIP 87-59) | 3.8 dSm−1 | 69% | [62] |
| Chickpea (var. FLIP 87-59) | 2.5 dSm−1 | 43% | [20] |
| Chickpea (var. ILC 3279) | 3.8 dSm−1 | 72% | [20] |
| Fababean (loam soil) | 6.6 dSm−1 | 50% | [62] |
| Fababean (clay soil) | 5.6 dSm−1 | 52% | [62] |
| Fababean (loam soil) | 4.9 dSm−1 | 28% | [20] |
| Fababean (clay soil) | 4.3 dSm−1 | 19% | [20] |
| Lentil (cv. 6796) | 3.1 dSm−1 | 100% | [20] |
| Lentil (cv. 6796) | 2 dSm−1 | 14% | [20] |
| Lentil (cv. 5582) | 2 dSm−1 | 24% | [20] |
| Lentil (cv. 5582) | 3.1 dSm−1 | 88% | [20] |
| Transgenic Crop | Gene Transferred | Source | Function | Reference |
|---|---|---|---|---|
| Soybean | P5CS | Solanum torvum Sw. | Synthesis and accumulation of proline | [200] |
| TaNHX2 | T. aestivum L. | Sodium vacuolar sequestration | [188] | |
| WRKY11 | Medicago sativa L. | Improves salt tolerance | [201] | |
| Pea | Na+/H+ | Arabidopsis thaliana L. | Sodium vacuolar sequestration | [202] |
| P5CS | Arabidopsis thaliana L. | Synthesis and accumulation of proline | [203] | |
| Chickpea | P5CS | Vigna aconitifolia L. | Synthesis and accumulation of proline | [204] |
| Faba bean | PR10a | Solanum tuberosum L. | Synthesis and accumulation of osmolytes | [185] |
| Mashbean | gly I | Brassica juncea L. | Increase in antioxidant ability | [205] |
| Pigeon pea | VaP5CSF129A | Vigna aconitifolia L. | Synthesis and accumulation of proline | [206] |
| VaP5CSF129A | Vigna aconitifolia L. | Synthesis and accumulation of osmolytes | [207] | |
| Lentil | DREB1A | Arabidopsis thaliana L. | Synthesis and accumulation of osmolytes | [194] |
| Peanut | AtNHXI | Arabidopsis thaliana L. | Sodium vacuolar sequestration | [208] |
| AtDREB1A | Arabidopsis thaliana L. | Improves salt tolerance | [193] | |
| AtHDG11 | Arabidopsis thaliana L. | Improves salt tolerance | [209] | |
| Alfalfa | SsNHX1 | Suaeda salsa | Regulate plant Na+/H+ antiporters | [210] |
| CsALDH12A1 | Cleistogenes songorica L. | Improves salt tolerance | [211] | |
| GmDREB1 | Glycine max L. | Conferred salt tolerance | [212] | |
| IbOr | Ipomoea batatas L. | Increased tolerance to multiple abiotic stresses | [213] | |
| GsCBRLK | Glycine soja L. | Improves salt tolerance | [186] | |
| TaNHX2 | Triticum aestivum L. | Regulate plant Na+/H+ antiporters | [214] | |
| ScNHX1 and ScVP | Suaeda corniculata | Vacuolar membrane H+-pyrophosphatases and H+-ATPases | [215] | |
| GsZFP1 | Glycine soja L. | Improves salinity tolerance | [216] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadeem, M.; Li, J.; Yahya, M.; Wang, M.; Ali, A.; Cheng, A.; Wang, X.; Ma, C. Grain Legumes and Fear of Salt Stress: Focus on Mechanisms and Management Strategies. Int. J. Mol. Sci. 2019, 20, 799. https://doi.org/10.3390/ijms20040799
Nadeem M, Li J, Yahya M, Wang M, Ali A, Cheng A, Wang X, Ma C. Grain Legumes and Fear of Salt Stress: Focus on Mechanisms and Management Strategies. International Journal of Molecular Sciences. 2019; 20(4):799. https://doi.org/10.3390/ijms20040799
Chicago/Turabian StyleNadeem, Muhammad, Jiajia Li, Muhammad Yahya, Minghua Wang, Asif Ali, Andong Cheng, Xiaobo Wang, and Chuanxi Ma. 2019. "Grain Legumes and Fear of Salt Stress: Focus on Mechanisms and Management Strategies" International Journal of Molecular Sciences 20, no. 4: 799. https://doi.org/10.3390/ijms20040799