Genome-Wide Characterization and Identification of Trihelix Transcription Factor and Expression Profiling in Response to Abiotic Stresses in Rice (Oryza sativa L.)


Abstract
:1. Introduction
2. Results
2.1. Identification of Trihelix Genes in Rice
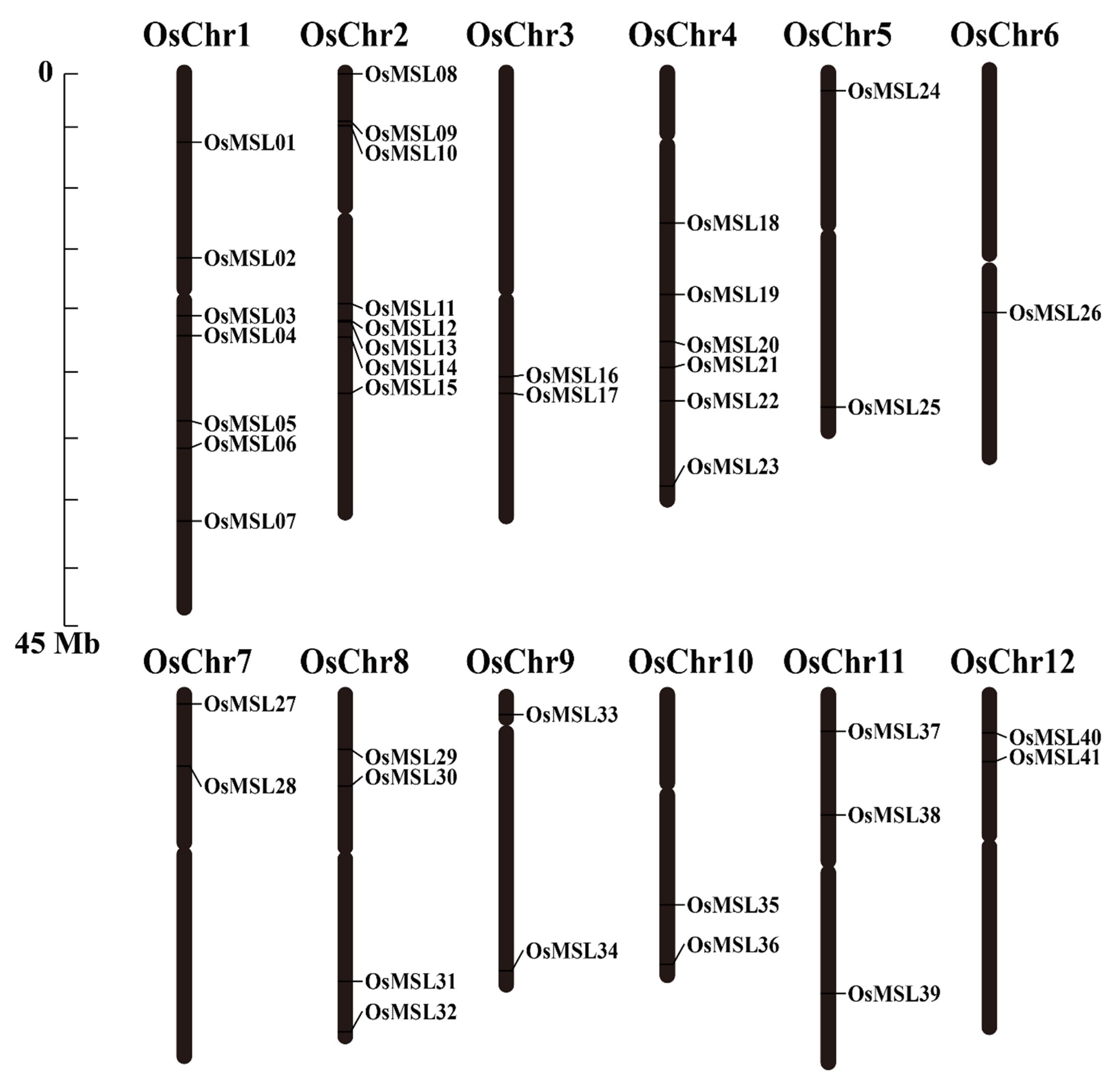
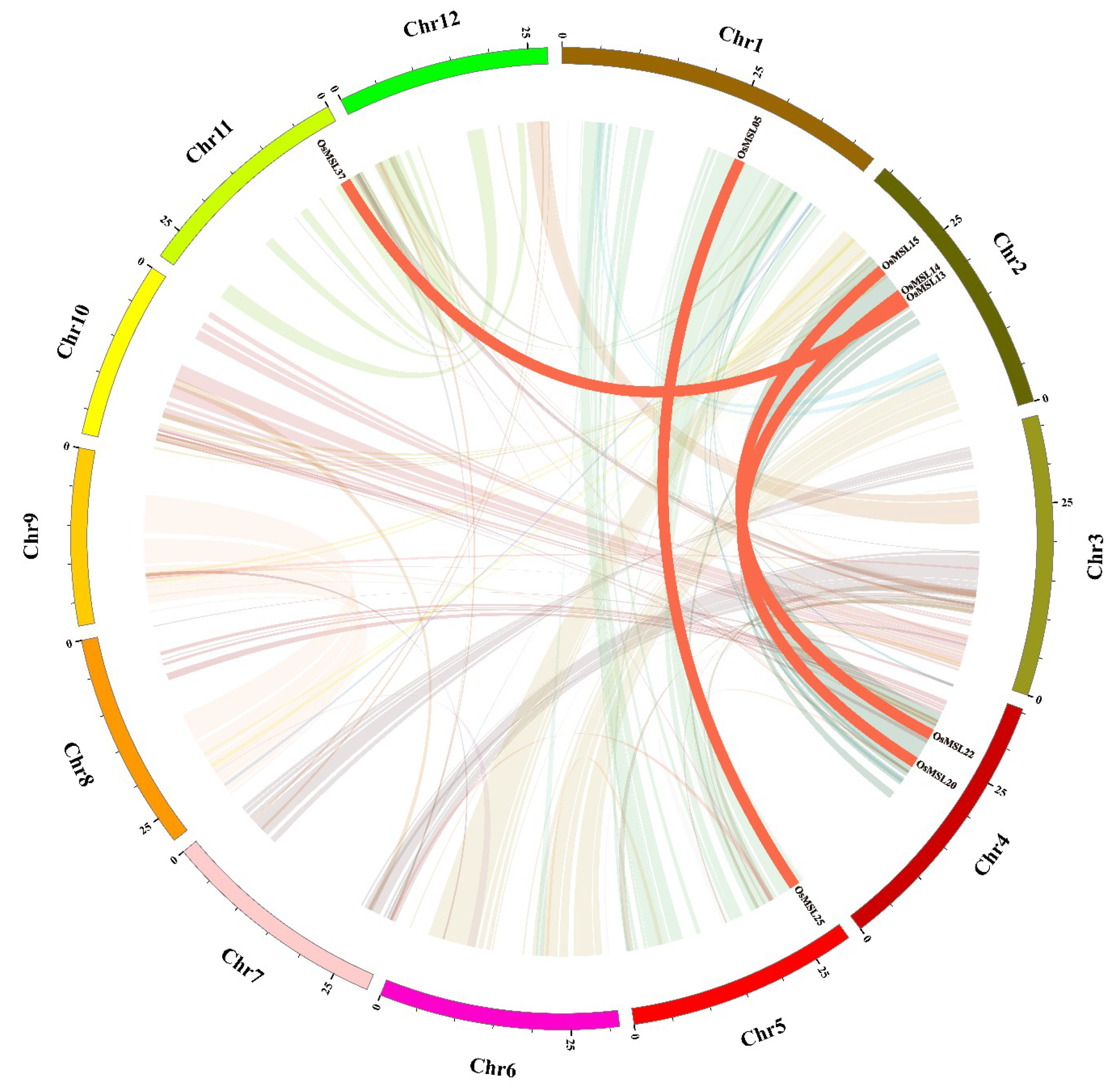
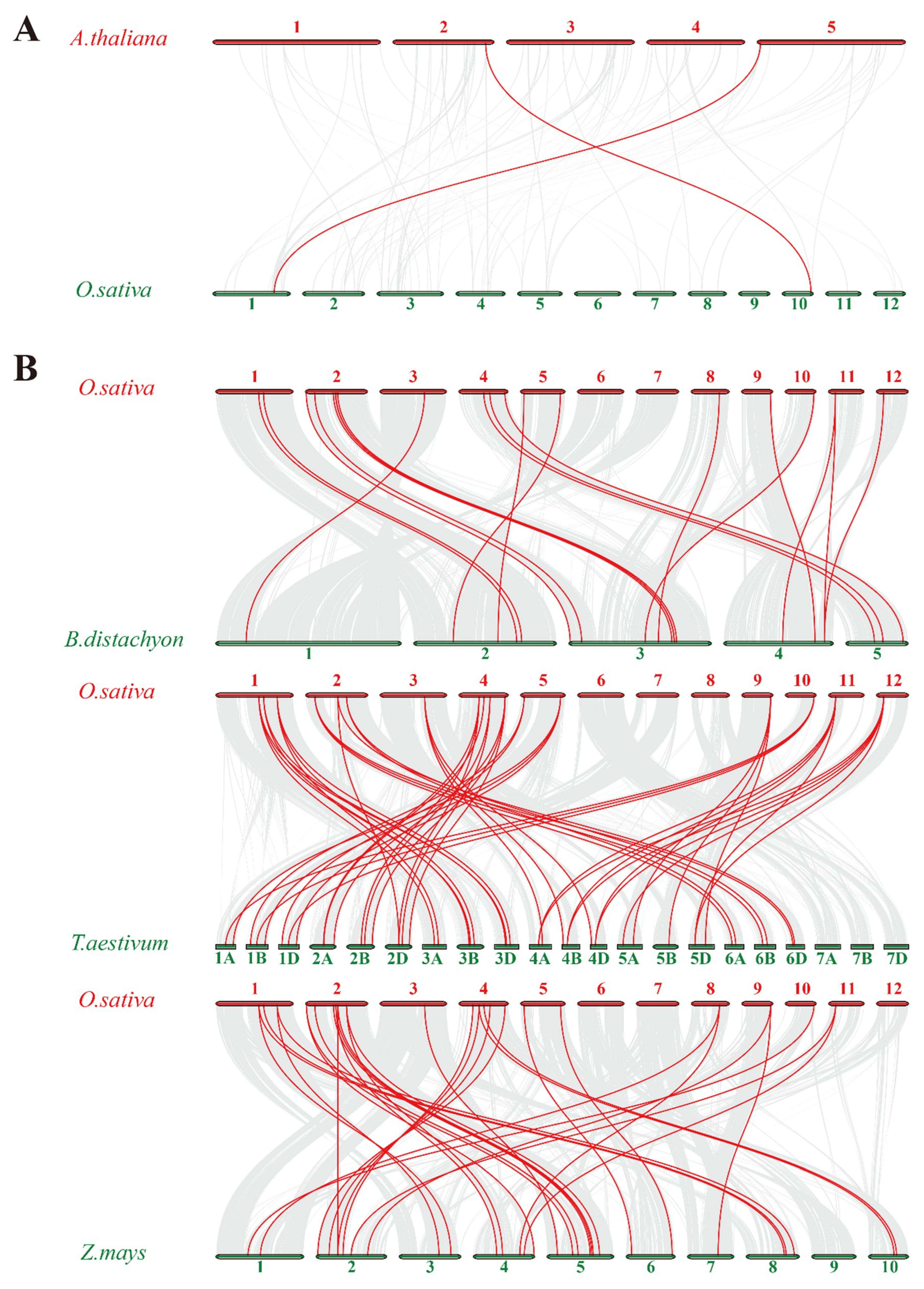
2.2. Chromosomal Distributions and Synteny Analysis of Rice Trihelix Genes
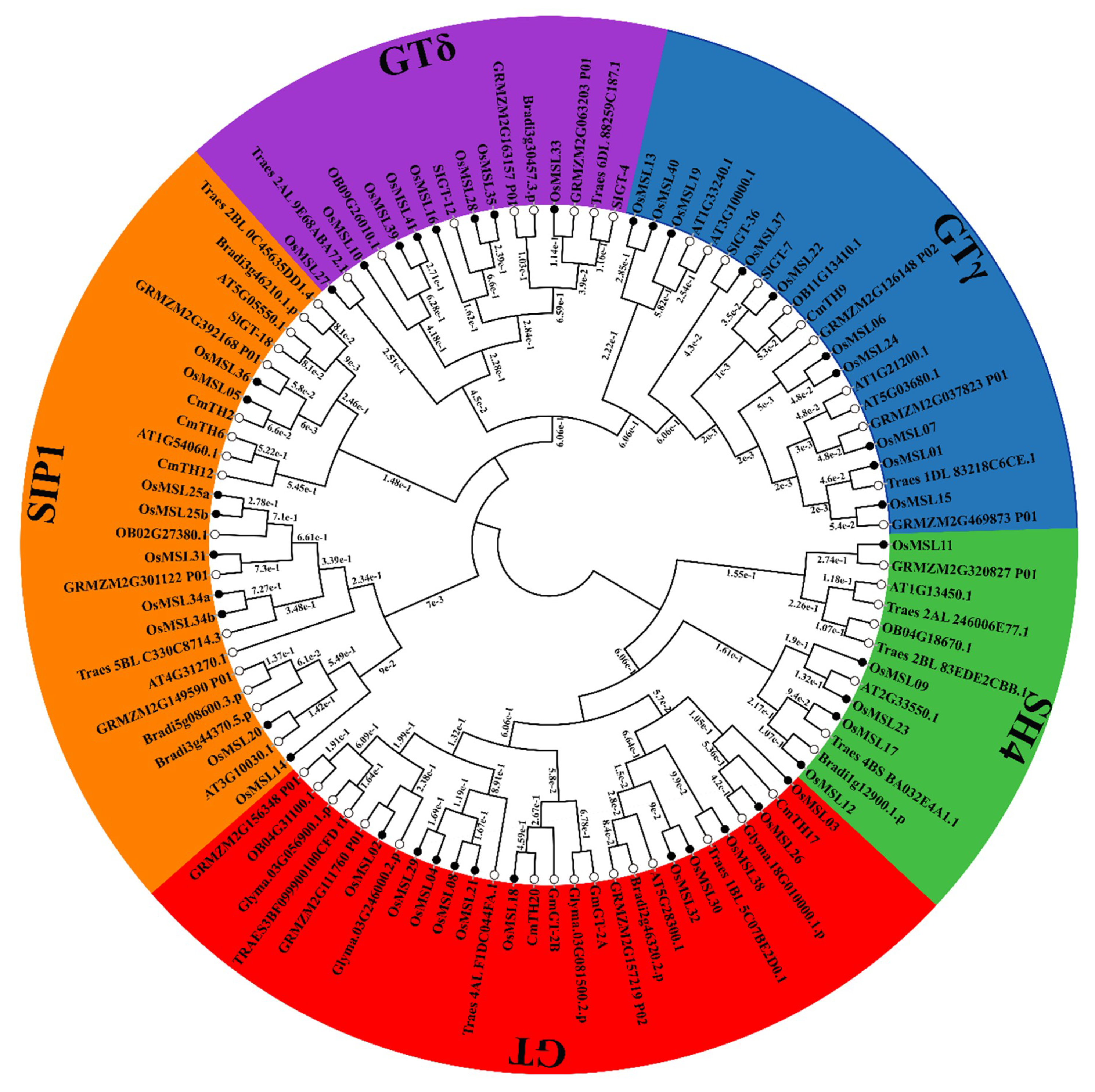
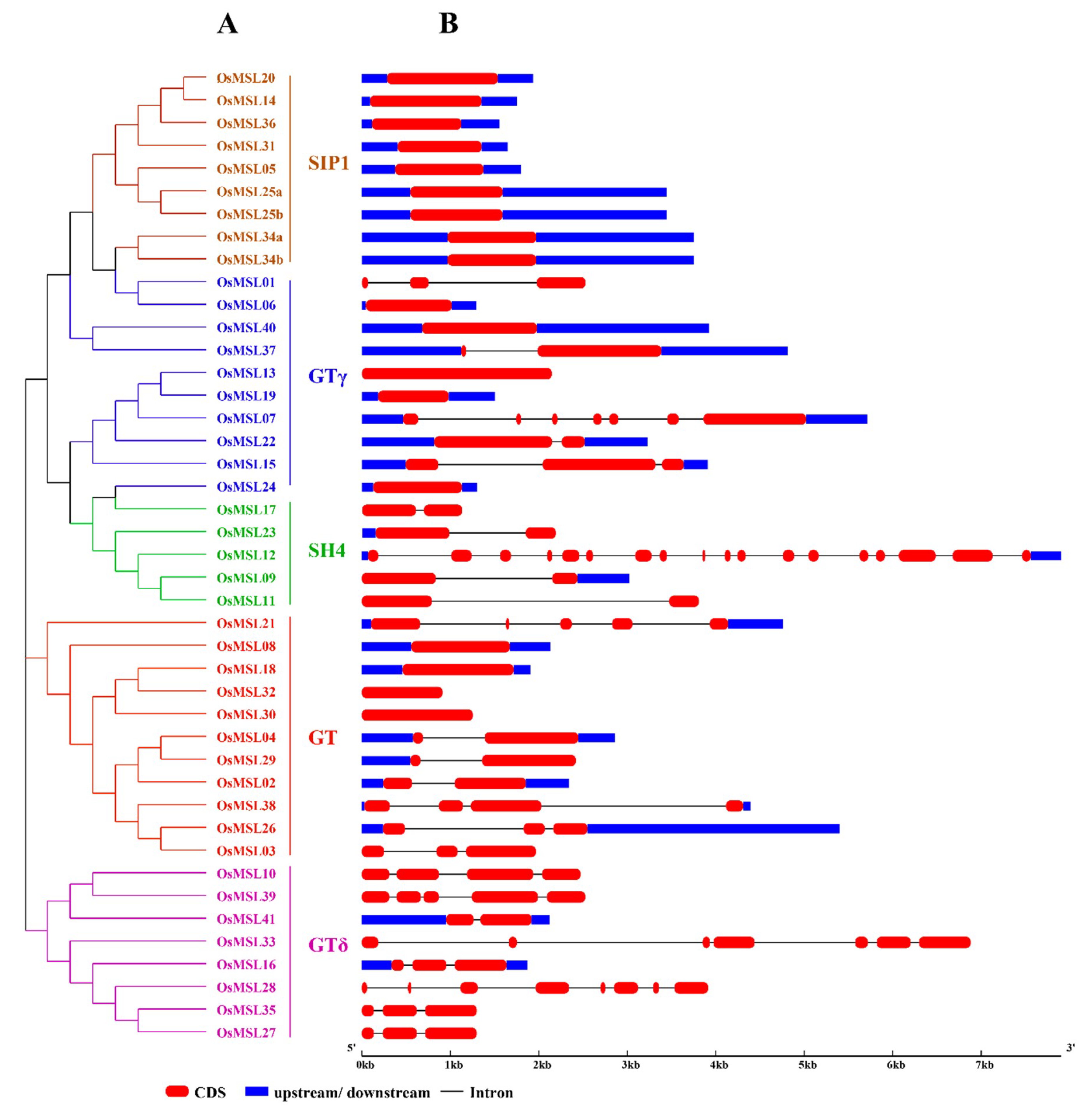
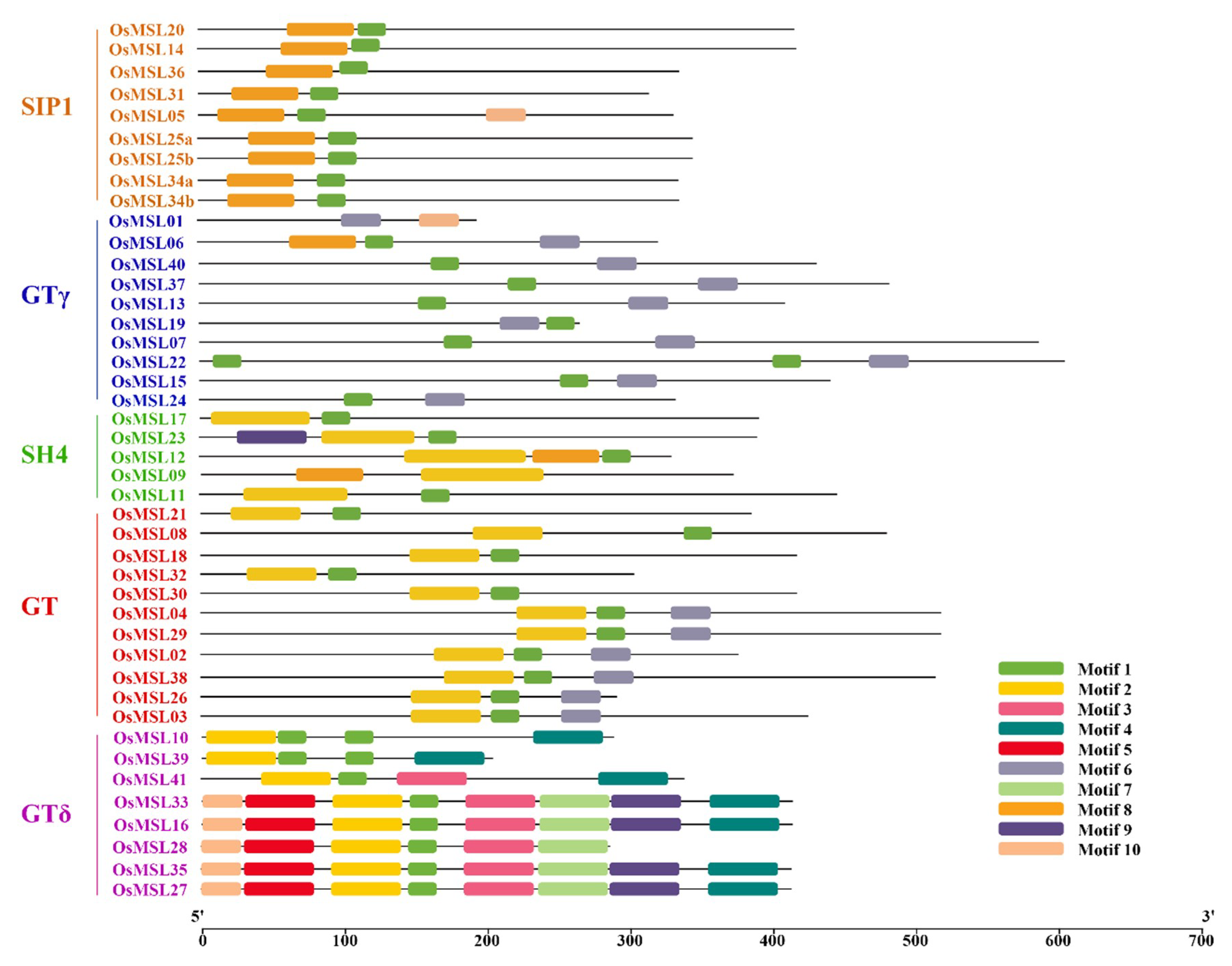
2.3. Phylogenetic Analysis, Gene Structure, and Motif Composition of Trihelix Genes
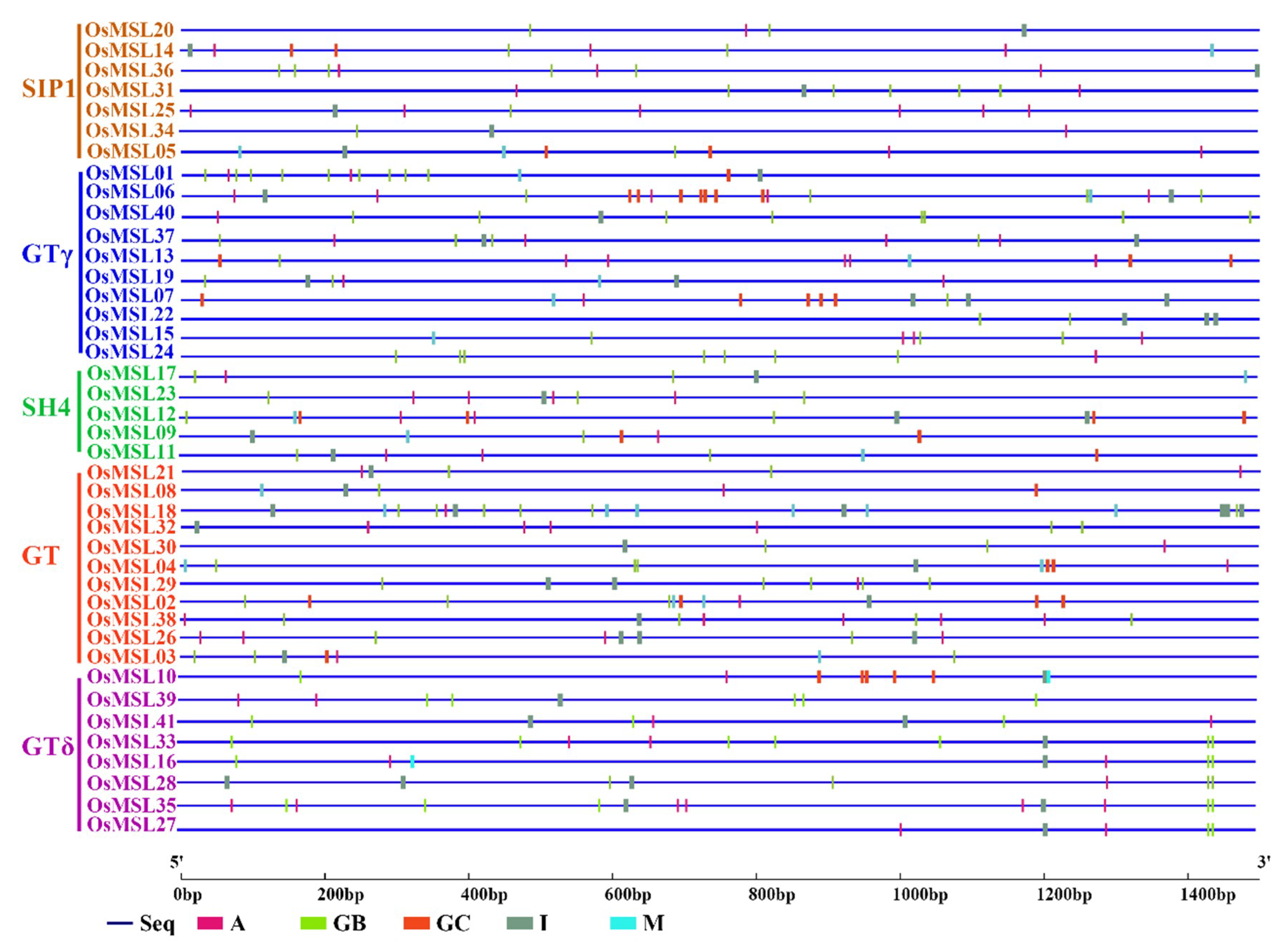
2.4. Cis-Element Analysis of Rice Trihelix Genes
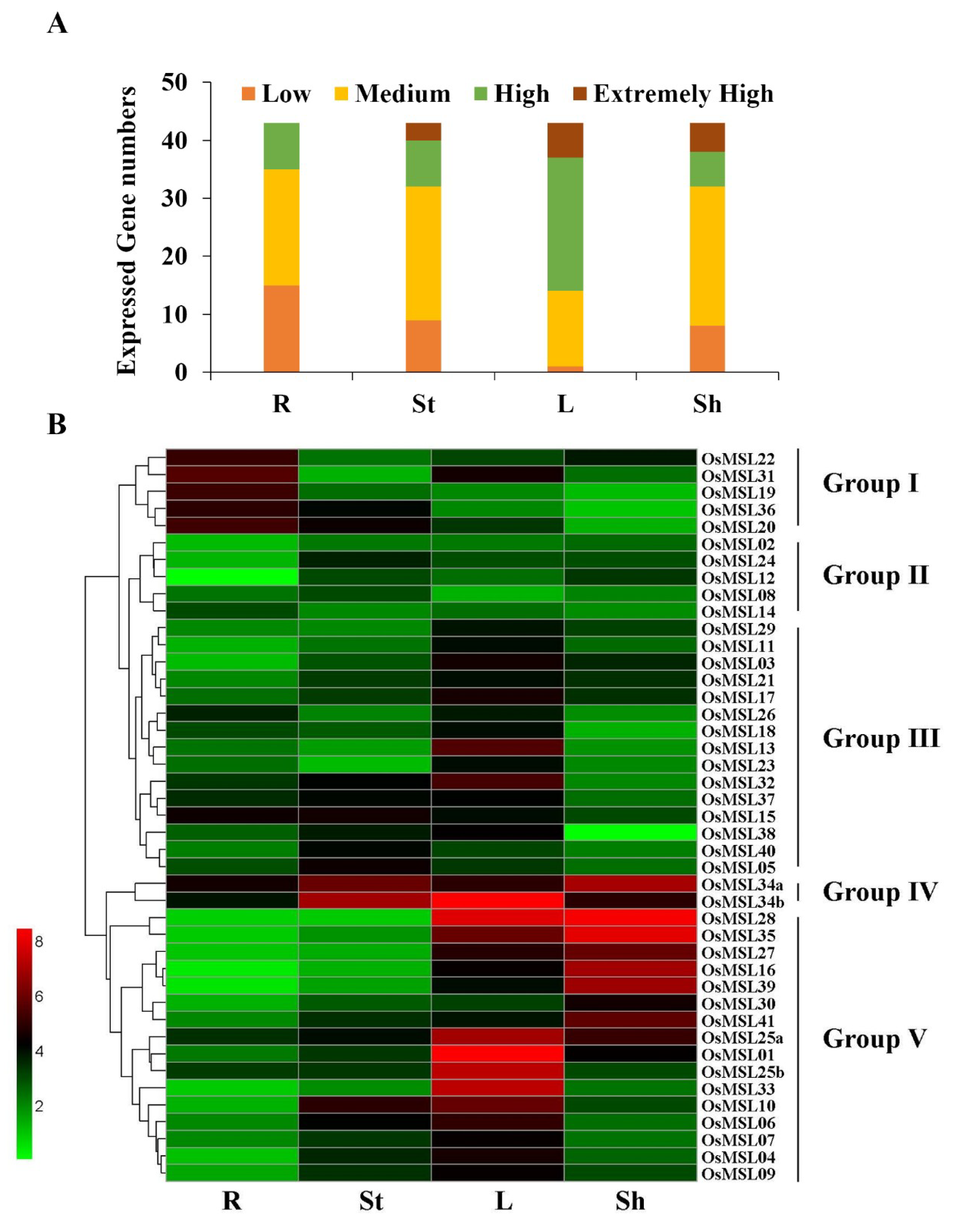
2.5. Expression Profiles of Trihelix Genes in Rice Tissues and Developmental Stages
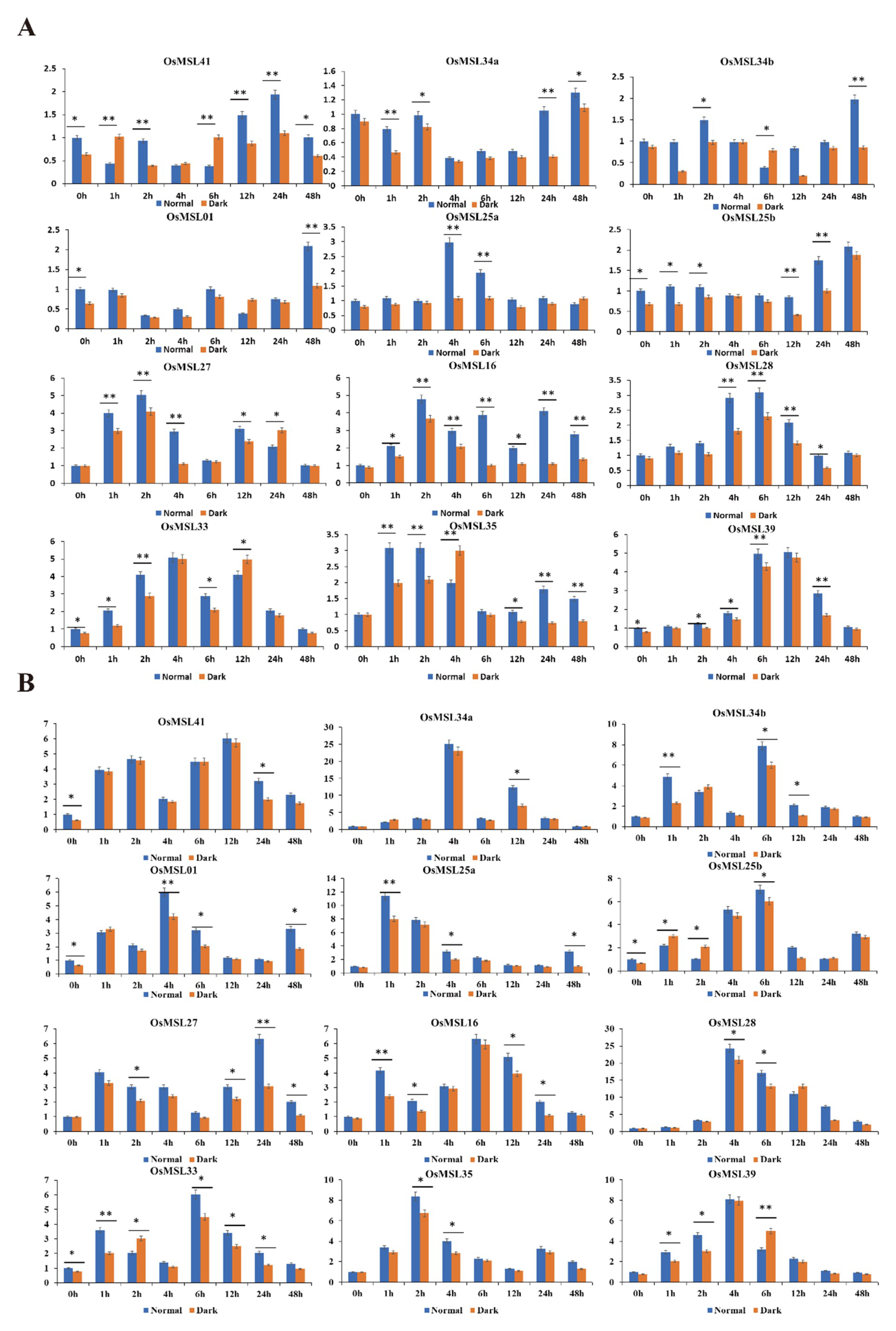
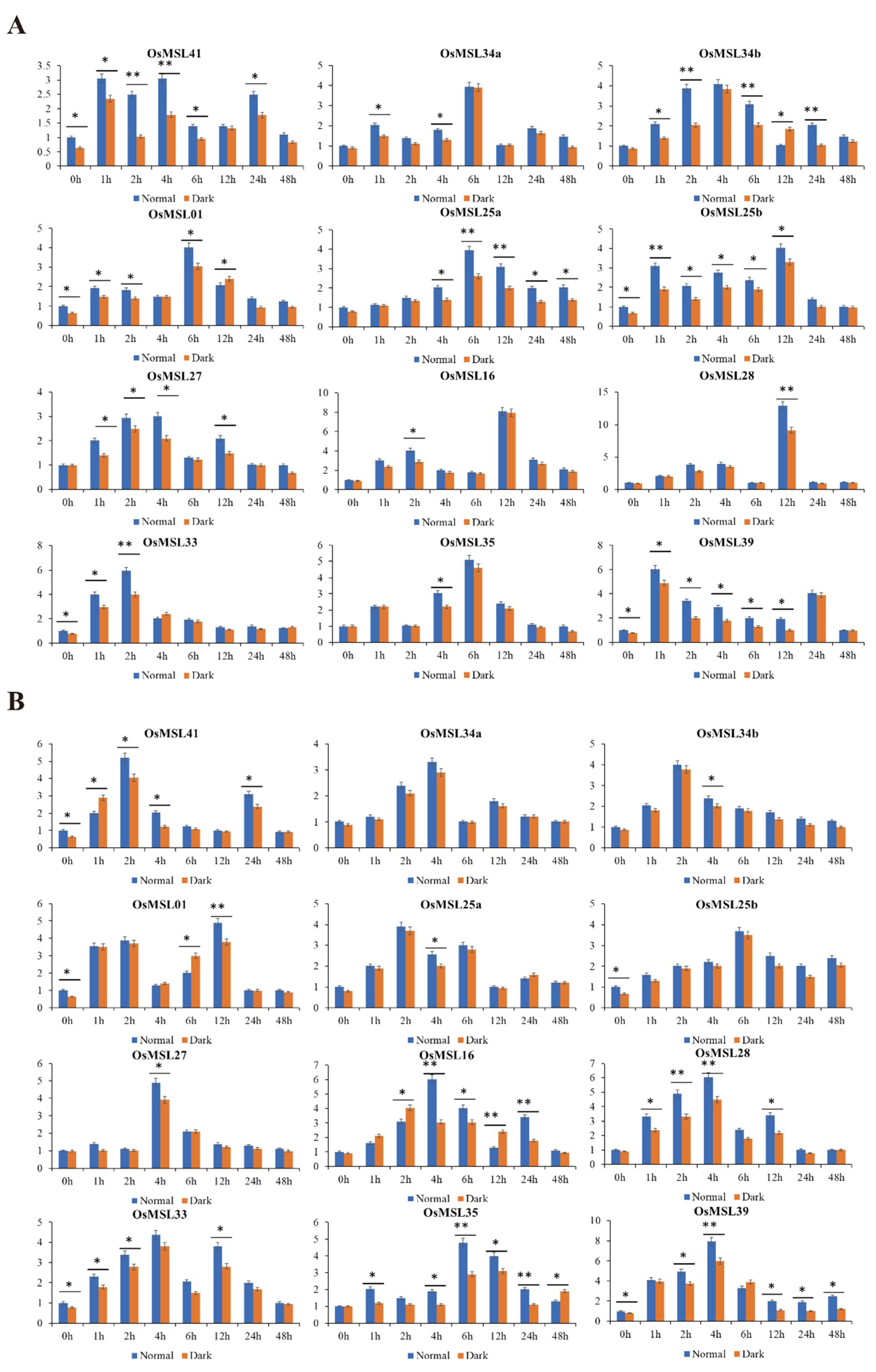
2.6. Quantitative Real-Time PCR Analysis of Rice Trihelix Genes in Responses to Different Treatments under Normal and Dark Conditions
3. Discussion
4. Materials and Methods
4.1. Identification and Sequence Analysis of Trihelix Transcription Factor Family in Rice
4.2. Phylogenetic Analysis
4.3. Gene Duplication and Ka/Ks Analysis
4.4. Cis-Element Analysis of Trihelix Transcription Factor Family
4.5. Plant Growth Conditions and Treatments
4.6. Expression Analysis of Trihelix Transcription Factor Family
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindemose, S.; O’Shea, C.; Jensen, M.K.; Skriver, K. Structure, Function and Networks of Transcription Factors Involved in Abiotic Stress Responses. Int. J. Mol. Sci. 2013, 14, 5842–5878. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.P.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.C.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic. Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105. [Google Scholar] [CrossRef] [PubMed]
- Green, P.J.; Kay, S.A.; Chua, N.H. Sequence-specific interactions of a pea nuclear factor with light-responsive elements upstream of the rbcS-3A gene. EMBO J. 1987, 6, 2543–2549. [Google Scholar] [CrossRef] [PubMed]
- Nagano, Y. Several Features of the GT-Factor Trihelix Domain Resemble Those of the Myb DNA-Binding Domain. Plant Physiol. 2000, 124, 491–493. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Ma, X.; Yu, G.H.; Wang, Q.; Wang, L.; Kong, L.R.; Kim, W.; Wang, H.W. Evolutionary History of Trihelix Family and Their Functional Diversification. DNA Res. 2014, 21, 499–510. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.J.; Lydiate, D.J.; Li, X.; Lui, H.; Gjetvaj, B.; Hegedus, D.D.; Rozwadowski, K. Repression of Seed Maturation Genes by a Trihelix Transcriptional Repressor in Arabidopsis Seedlings. Plant Cell 2009, 21, 54–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.Y.; Cal, X.F.; Ye, Z.B.; Li, H.X. Genome-wide identification and expression profiling analysis of trihelix gene family in tomato. Biochem. Biophys. Res. Commun. 2015, 468, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Murata, J.; Takase, H.; Hiratsuka, K. Characterization of a Novel GT-box Binding Protein from Arabidopsis. Plant Biotechnol. 2002, 19, 103–112. [Google Scholar] [CrossRef]
- Wang, R.H.G.; Han, B. Transcript abundance of rml1, encoding a putative GT1-like factor in rice, is up-regulated by Magnaporthe grisea and down-regulated by light. Gene 2004, 324, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.M.; Zou, H.F.; Lei, G.; Wei, W.; Zhou, Q.Y.; Niu, C.F.; Liao, Y.; Tian, A.G.; Ma, B.; Zhang, W.K. Soybean Trihelix Transcription Factors GmGT-2A and GmGT-2B Improve Plant Tolerance to Abiotic Stresses in Transgenic Arabidopsis. PLoS ONE 2009, 4, e6898. [Google Scholar] [CrossRef]
- Chan, Y.Y.; Pence, H.E.; Jing, B.J.; Miura, K.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. The Arabidopsis GTL1 Transcription Factor Regulates Water Use Efficiency and Drought Tolerance by Modulating Stomatal Density via Transrepression of SDD1. Plant Cell 2010, 22, 4128. [Google Scholar]
- Fang, Y.; Xie, K.; Xin, H.; Hu, H.; Xiong, L. Systematic analysis of GT factor family of rice reveals a novel subfamily involved in stress responses. Mol. Genet. Genom. 2010, 283, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Song, A.P.; Wu, D.; Fan, Q.Q.; Tian, C.; Chen, S.M.; Guan, Z.Y.; Xin, J.J.; Zhao, K.K.; Chen, F.D. Transcriptome-Wide Identification and Expression Profiling Analysis of Chrysanthemum Trihelix Transcription Factors. Int. J. Mol. Sci. 2016, 17, 198. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhou, A.; Sang, T. Rice Domestication by Reducing Shattering. Science 2006, 311, 1936–1939. [Google Scholar] [CrossRef] [PubMed]
- Holub, E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nat. Rev. Genet. 2001, 2, 516–527. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- KaplanLevy, R.N.; Brewer, P.B.; Quon, T.; Smyth, D.R. The trihelix family of transcription factors—Light, stress and development. Trends Plant Sci. 2012, 17, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Markus, S.; Davison, T.S.; Henz, S.R.; Pape, U.J.; Monika, D.; Martin, V.; Bernhard, S.L.; Detlef, W.; Lohmann, J.U. A gene expression map of Arabidopsis thaliana development. Nat. Genet. 2005, 37, 501–506. [Google Scholar]
- Nagata, T.; Niyada, E.; Fujimoto, N.; Nagasaki, Y.; Noto, K.; Miyanoiri, Y.; Murata, J.; Hiratsuka, K.; Katahira, M. Solution structures of the trihelix DNA-binding domains of the wild-type and a phosphomimetic mutant of Arabidopsis GT-1: Mechanism for an increase in DNA-binding affinity through phosphorylation. Proteins 2010, 78, 3033–3047. [Google Scholar] [CrossRef] [PubMed]
- Jianhui, J.; Yingjun, Z.; Hehe, W.; Liming, Y. Genome-wide analysis and functional prediction of the Trihelix transcription factor family in rice. Hereditas 2015, 37, 1228. (In Chinese) [Google Scholar]
- Geraldo, N.; Baurle, I.; Kidou, S.; Hu, X.; Dean, C. FRIGIDA Delays Flowering in Arabidopsis via a Cotranscriptional Mechanism Involving Direct Interaction with the Nuclear Cap-Binding Complex. Plant Physiol. 2009, 150, 1611. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Yasmeen, E.; Riaz, M.; Azeem, F.; Sultan, S.; Abbas, A.; Riaz, K. Genome-wide analysis of trihelix Transcription factor gene family in Arabidopsis thaliana. Pak. J. Agric. Sci. 2016, 53, 439–448. [Google Scholar]
- Lam, E. Domain analysis of the plant DNA-binding protein GT1a: Requirement of four putative alpha-helices for DNA binding and identification of a novel oligomerization region. Mol. Cell. Biol. 1995, 15, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Lau, O.S.; Deng, X.W. Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 2007, 8, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Toledoortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis Basic/Helix-Loop-Helix Transcription Factor Family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef] [Green Version]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Ballesteros, M.L.; Bolle, C.; Lois, L.M.; Moore, J.M.; Viellecalzada, J.P.; Grossniklaus, U.; Chua, N.H. LAF1, a MYB transcription activator for phytochrome A signaling. Gene Dev. 2001, 15, 2613–2625. [Google Scholar] [CrossRef] [Green Version]
- Hudson, M.; Ringli, C.; Boylan, M.T.; Quail, P.H. The FAR1 locus encodes a novel nuclear protein specific to phytochrome A signaling. Gene Dev. 1999, 13, 2017–2027. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Deng, X.W. Arabidopsis FHY3 defines a key phytochrome A signaling component directly interacting with its homologous partner FAR1. EMBO J. 2002, 21, 1339–1349. [Google Scholar] [CrossRef]
- Tang, Y.; Qin, S.; Guo, Y.; Chen, Y.; Wu, P.; Chen, Y.; Li, M.; Jiang, H.; Wu, G. Genome-Wide Analysis of the AP2/ERF Gene Family in Physic Nut and Overexpression of the JcERF011 Gene in Rice Increased Its Sensitivity to Salinity Stress. PLoS ONE 2016, 11, e0150879. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Sun, S.; Xu, D.; Lan, H.; Sun, H.; Wang, Z.; Bao, Y.; Wang, J.; Tang, H.; Zhang, H. A TFIIIA-type zinc finger protein confers multiple abiotic stress tolerances in transgenic rice (Oryza sativa L.). Plant Mol. Biol. 2012, 80, 337. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Cheng, X.; Gu, Z.; Huang, W.; Li, S.; Wang, L.; Wang, Y.F.; Xu, P.; Ma, H.; Ge, X. The AWPM-19 Family Protein OsPM1 Mediates Abscisic Acid Influx and Drought Response in Rice. Plant Cell 2018, 30, 1258–1276. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Gilmartin, P.M.; Memelink, J.; Hiratsuka, K.; Kay, S.A.; Chua, N.H. Characterization of a gene encoding a DNA binding protein with specificity for a light-responsive element. Plant Cell 1992, 4, 839–849. [Google Scholar] [CrossRef]
- Lozano, R.; Hamblin, M.T.; Prochnik, S.; Jannink, J.L. Identification and distribution of the NBS-LRR gene family in the Cassava genome. BMC Genom. 2015, 16, 360. [Google Scholar] [CrossRef] [PubMed]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A. The Pfam protein families database in 2019. Nucleic Acids Res. 2018, 28, 263–266. [Google Scholar] [CrossRef]
- Kersey, P.J.; Allen, J.E.; Allot, A.; Barba, M.; Boddu, S.; Bolt, B.J.; Carvalho-Silva, D.; Christensen, M.; Davis, P.; Grabmueller, C. Ensembl Genomes 2018: An integrated omics infrastructure for non-vertebrate species. Nucleic Acids Res. 2018, 46, D802–D808. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.-Y.; El-Gebali, S.; Fraser, M.I. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296. [Google Scholar] [CrossRef] [PubMed]
- Kurata, N.; Yamazaki, Y. Oryzabase. An integrated biological and genome information database for rice. Plant Physiol. 2006, 140, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; De Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Tang, H.; Wang, X.; Paterson, A.H. PGDD: A database of gene and genome duplication in plants. Nucleic Acids Res. 2013, 41, 1152–1158. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2011, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Gao, F.; Wu, X.; Lu, X.; Zeng, L.; Lv, J.; Su, X.; Luo, H.; Ren, G. Bph32, a novel gene encoding an unknown SCR domain-containing protein, confers resistance against the brown planthopper in rice. Sci. Rep. 2016, 6, 37645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coordinators, N.R. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2017, 45, D12. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Papatheodorou, I.; Fonseca, N.A.; Keays, M.; Tang, Y.A.; Barrera, E.; Bazant, W.; Burke, M.; Füllgrabe, A.; Fuentes, A.M.-P.; George, N. Expression Atlas: Gene and protein expression across multiple studies and organisms. Nucleic Acids Res. 2017, 46, D246–D251. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, Y.; Liu, Z.; Cheng, H.; Xue, Y. HemI: A Toolkit for Illustrating Heatmaps. PLoS ONE 2014, 9, e111988. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Locus | Chr | ORF (bp) | Exon No. | Length (aa) | MW (kD) | pI | Localization |
|---|---|---|---|---|---|---|---|---|
| OsMSL01 | LOC_Os01g11200.1 | Chr1 | 828 | 3 | 275 | 31.8 | 8.57 | Nucleus |
| OsMSL02 | LOC_Os01g27590.1 | Chr1 | 1131 | 2 | 376 | 41.08 | 8.6 | Nucleus |
| OsMSL03 | LOC_Os01g34400.1 | Chr1 | 1227 | 3 | 408 | 46.06 | 9.22 | Chloroplast |
| OsMSL04 | LOC_Os01g36850.1 | Chr1 | 1170 | 2 | 389 | 57.61 | 9.46 | Peroxisome |
| OsMSL05 | LOC_Os01g48320.1 | Chr1 | 999 | 1 | 332 | 35.92 | 9.62 | Nucleus |
| OsMSL06 | LOC_Os01g52090.1 | Chr1 | 969 | 1 | 322 | 35.76 | 5.54 | Nucleus |
| OsMSL07 | LOC_Os01g62410.1 | Chr1 | 1764 | 7 | 587 | 64.1 | 8.5 | Nucleus |
| OsMSL08 | LOC_Os02g01380.1 | Chr2 | 1113 | 1 | 370 | 40.5 | 5.33 | Nucleus |
| OsMSL09 | LOC_Os02g07800.1 | Chr2 | 1308 | 2 | 435 | 46.24 | 4.45 | Chloroplast |
| OsMSL10 | LOC_Os02g08450.1 | Chr2 | 1968 | 4 | 655 | 74.35 | 7.09 | Chloroplast |
| OsMSL11 | LOC_Os02g31160.1 | Chr2 | 1125 | 2 | 374 | 39.55 | 5.17 | Nucleus |
| OsMSL12 | LOC_Os02g33610.1 | Chr2 | 2649 | 18 | 882 | 97.37 | 8.97 | Chloroplast |
| OsMSL13 | LOC_Os02g33770.1 | Chr2 | 1233 | 1 | 410 | 46.67 | 6.28 | Nucleus |
| OsMSL14 | LOC_Os02g35690.1 | Chr2 | 1260 | 1 | 419 | 44.44 | 6.11 | Nucleus |
| OsMSL15 | LOC_Os02g43300.1 | Chr2 | 1887 | 3 | 628 | 67.74 | 4.87 | Nucleus |
| OsMSL16 | LOC_Os03g44130.1 | Chr3 | 1104 | 3 | 367 | 41.74 | 6.61 | Chloroplast |
| OsMSL17 | LOC_Os03g46350.1 | Chr3 | 1038 | 2 | 345 | 42.49 | 11.38 | Nucleus |
| OsMSL18 | LOC_Os04g21860.1 | Chr4 | 1254 | 1 | 417 | 47.15 | 9.1 | Nucleus |
| OsMSL19 | LOC_Os04g30890.1 | Chr4 | 801 | 1 | 266 | 28.62 | 9.64 | Nucleus |
| OsMSL20 | LOC_Os04g36790.1 | Chr4 | 1254 | 1 | 417 | 43.93 | 6.53 | Nucleus |
| OsMSL21 | LOC_Os04g40930.1 | Chr4 | 1158 | 5 | 385 | 41.93 | 5.82 | Nucleus |
| OsMSL22 | LOC_Os04g45750.1 | Chr4 | 1587 | 2 | 528 | 57.46 | 5.74 | Nucleus |
| OsMSL23 | LOC_Os04g57530.1 | Chr4 | 1173 | 2 | 390 | 41.44 | 9.01 | Nucleus |
| OsMSL24 | LOC_Os05g03740.1 | Chr5 | 1002 | 1 | 333 | 36.95 | 5.85 | Nucleus |
| OsMSL25 | LOC_Os05g48690.1 | Chr5 | 1041 | 1 | 346 | 37.44 | 9.92 | Nucleus |
| OsMSL26 | LOC_Os06g32944.1 | Chr6 | 876 | 3 | 291 | 32.82 | 8.22 | Nucleus |
| OsMSL27 | LOC_Os07g02500.1 | Chr7 | 1104 | 3 | 367 | 41.74 | 6.61 | Chloroplast |
| OsMSL28 | LOC_Os07g10950.1 | Chr7 | 1437 | 8 | 478 | 54.23 | 9.06 | Chloroplast |
| OsMSL29 | LOC_Os08g08130.1 | Chr8 | 1170 | 2 | 389 | 44.28 | 6.2 | Peroxisome |
| OsMSL30 | LOC_Os08g12950.1 | Chr8 | 1254 | 1 | 417 | 47.15 | 8.92 | Nucleus |
| OsMSL31 | LOC_Os08g37810.1 | Chr8 | 948 | 1 | 315 | 35.06 | 7.09 | Nucleus |
| OsMSL32 | LOC_Os08g44690.1 | Chr8 | 912 | 1 | 303 | 34.83 | 9.29 | Nucleus |
| OsMSL33 | LOC_Os09g03570.1 | Chr9 | 1932 | 7 | 643 | 73.57 | 9.06 | Chloroplast |
| OsMSL34 | LOC_Os09g38570.1 | Chr9 | 1011 | 1 | 336 | 36.34 | 6.58 | Nucleus |
| OsMSL35 | LOC_Os10g33030.1 | Chr10 | 1104 | 3 | 367 | 41.7 | 6.61 | Chloroplast |
| OsMSL36 | LOC_Os10g41460.1 | Chr10 | 1011 | 1 | 336 | 35.64 | 8.9 | Nucleus |
| OsMSL37 | LOC_Os11g06410.1 | Chr11 | 1492 | 2 | 483 | 55.06 | 6.24 | Nucleus |
| OsMSL38 | LOC_Os11g17954.1 | Chr11 | 1545 | 4 | 514 | 58.1 | 8.3 | Nucleus |
| OsMSL39 | LOC_Os11g38660.1 | Chr11 | 1938 | 5 | 645 | 72.97 | 7.34 | Chloroplast |
| OsMSL40 | LOC_Os12g06640.1 | Chr12 | 1299 | 1 | 432 | 48.77 | 6.13 | Nucleus |
| OsMSL41 | LOC_Os12g10550.1 | Chr12 | 888 | 2 | 295 | 33.5 | 7.74 | Nucleus |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhang, M.; Sun, J.; Mao, X.; Wang, J.; Wang, J.; Liu, H.; Zheng, H.; Zhen, Z.; Zhao, H.; et al. Genome-Wide Characterization and Identification of Trihelix Transcription Factor and Expression Profiling in Response to Abiotic Stresses in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2019, 20, 251. https://doi.org/10.3390/ijms20020251
Li J, Zhang M, Sun J, Mao X, Wang J, Wang J, Liu H, Zheng H, Zhen Z, Zhao H, et al. Genome-Wide Characterization and Identification of Trihelix Transcription Factor and Expression Profiling in Response to Abiotic Stresses in Rice (Oryza sativa L.). International Journal of Molecular Sciences. 2019; 20(2):251. https://doi.org/10.3390/ijms20020251
Chicago/Turabian StyleLi, Jiaming, Minghui Zhang, Jian Sun, Xinrui Mao, Jing Wang, Jingguo Wang, Hualong Liu, Hongliang Zheng, Zhen Zhen, Hongwei Zhao, and et al. 2019. "Genome-Wide Characterization and Identification of Trihelix Transcription Factor and Expression Profiling in Response to Abiotic Stresses in Rice (Oryza sativa L.)" International Journal of Molecular Sciences 20, no. 2: 251. https://doi.org/10.3390/ijms20020251