In Vitro Induction of Tendon-Specific Markers in Tendon Cells, Adipose- and Bone Marrow-Derived Stem Cells is Dependent on TGFβ3, BMP-12 and Ascorbic Acid Stimulation

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
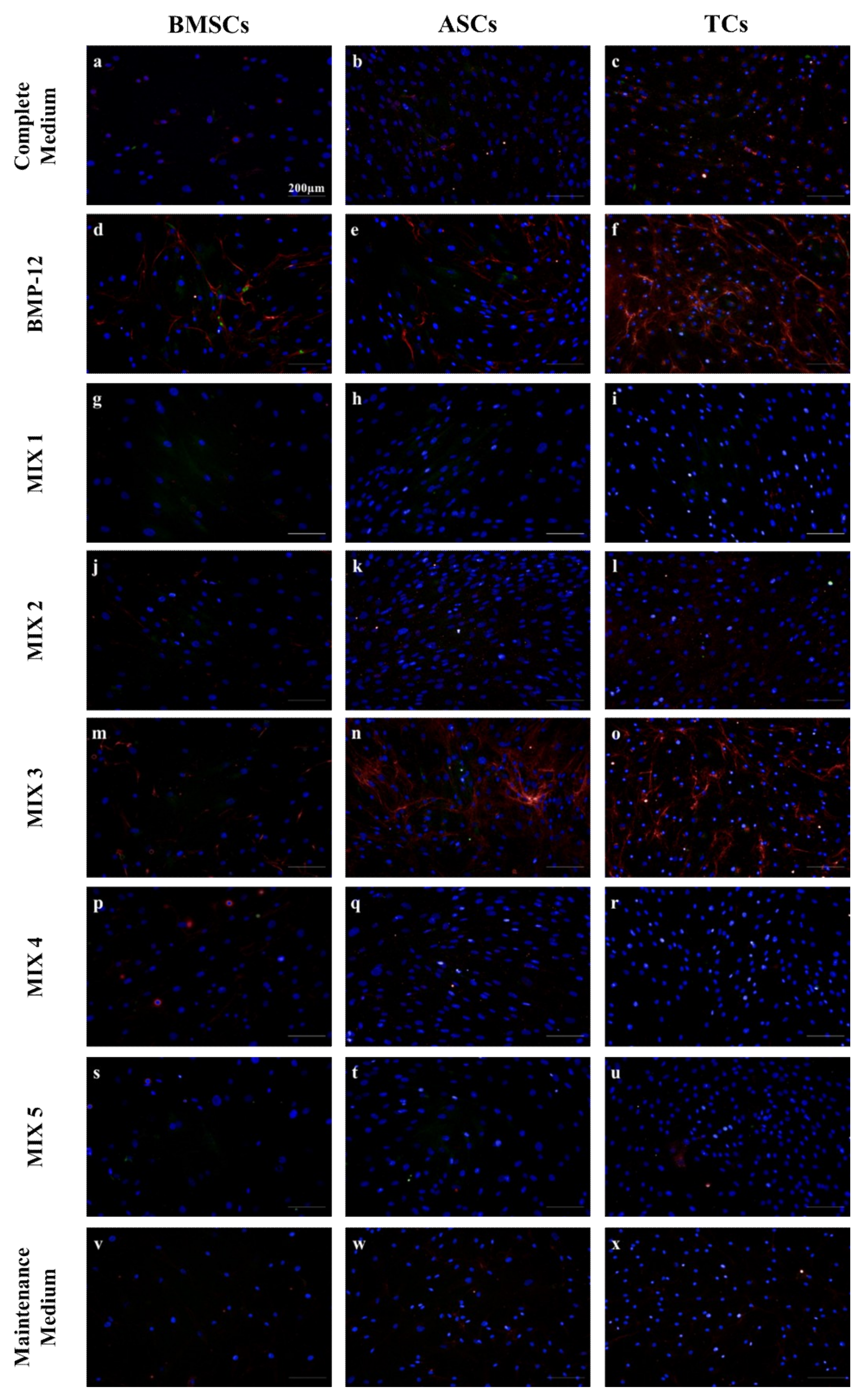
2.1. Scleraxis and Decorin Expression-Based Screen for Different Combinations of Candidate Tenogenic Media
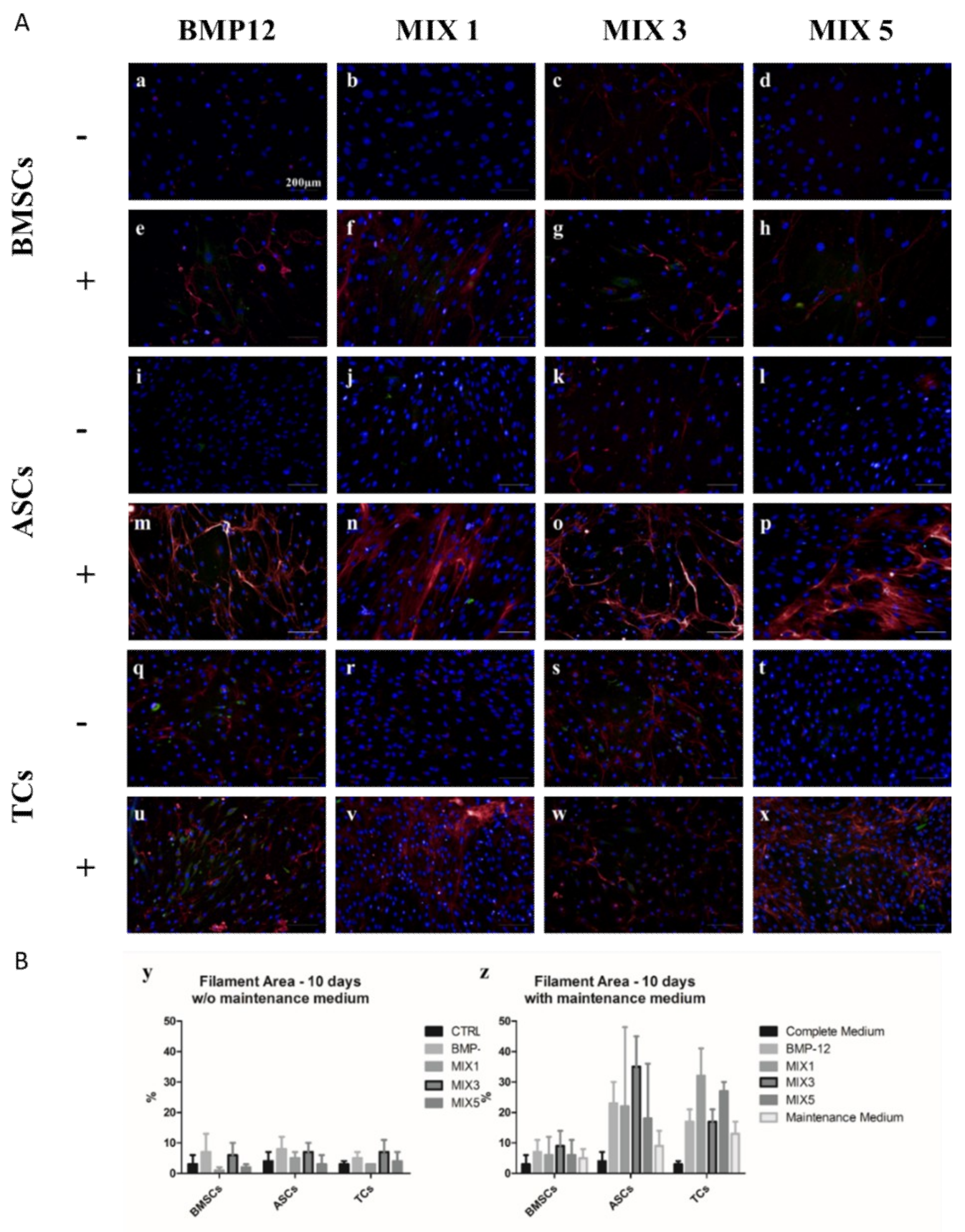
2.2. Role of b-FGF and Ascorbic Acid in the Maintenance of DCN Expression
2.3. Basal Levels of Tendon Specific Markers in Different Cell Types
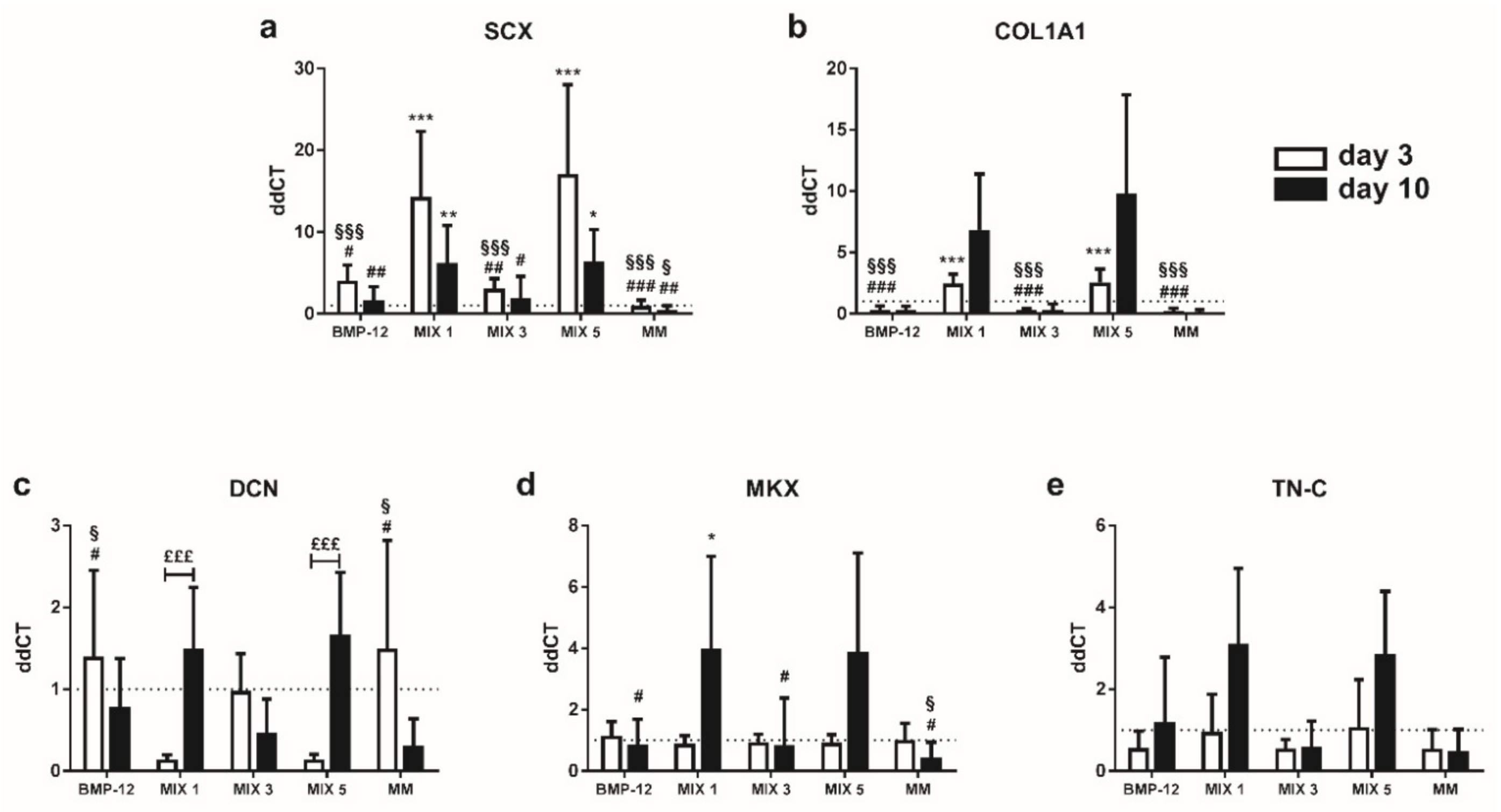
2.4. TGF-β3 Containing Media Induce the Expression of Tenogenic Markers in TCs
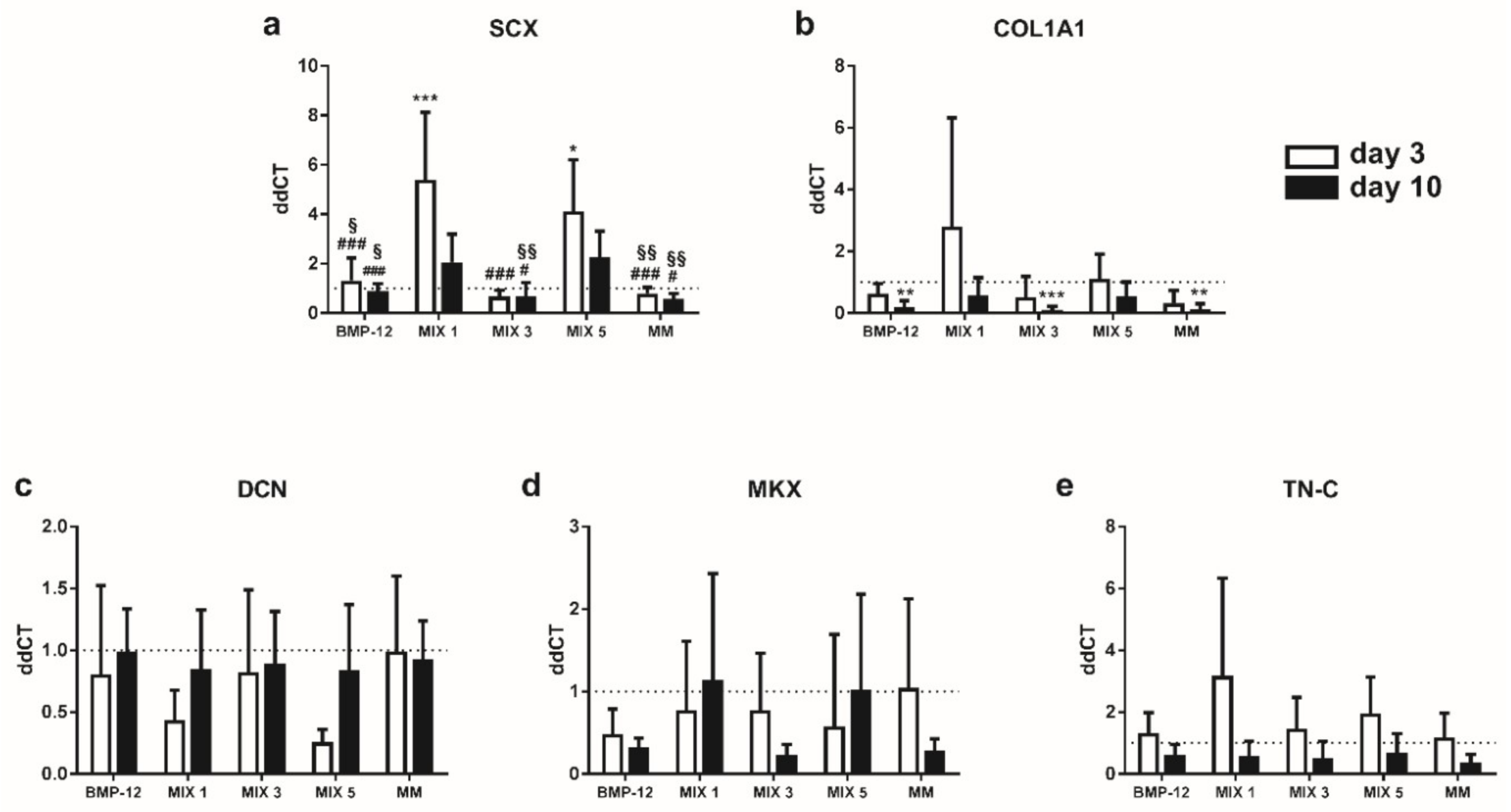
2.5. TGFβ3-Containing Media Induce the Expression of SCX in BMSCs
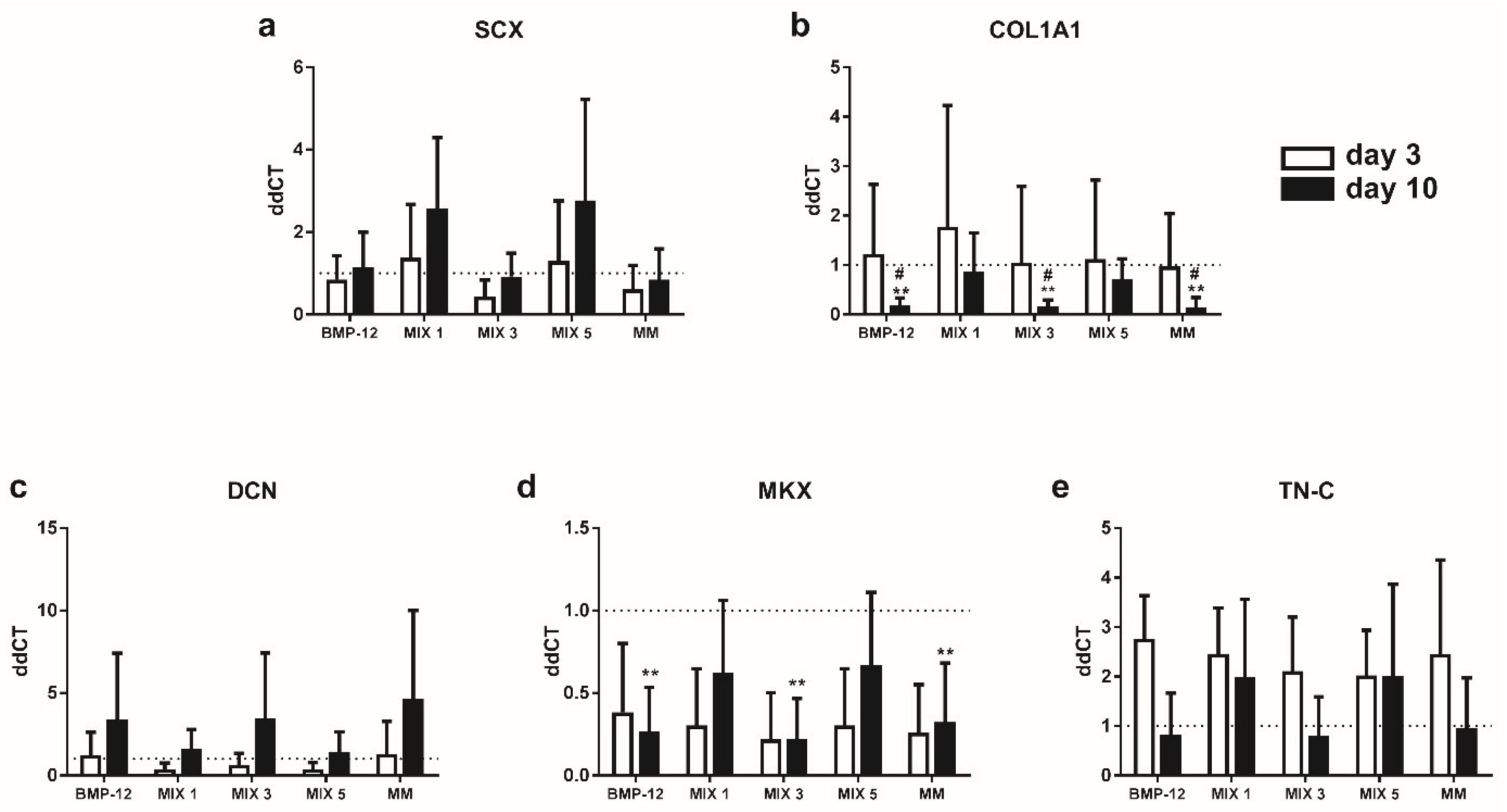
2.6. TGFβ3-Free Inductive Media Reduce the Expression of COL1A1 and MKX in ASCs
3. Discussion
4. Materials and Methods
4.1. MSCs Isolation and Culture
4.2. Growth Factors-Mediated Tenogenic Induction
4.3. Immunofluorescence Analysis
4.4. RNA Extraction and Gene Expression Analysis
4.5. Statistical Analysis
4.6. Data Availability
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yamamoto, A.; Takagishi, K.; Osawa, T.; Yanagawa, T.; Nakajima, D.; Shitara, H.; Kobayashi, T. Prevalence and risk factors of a rotator cuff tear in the general population. J. Shoulder Elbow Surg. 2010, 19, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Raikin, S.M.; Garras, D.N.; Krapchev, P.V. Achilles tendon injuries in a United States population. Foot Ankle Int. 2013, 34, 475–480. [Google Scholar] [CrossRef]
- Maffulli, N.; Testa, V.; Capasso, G.; Oliva, F.; Sullo, A.; Benazzo, F.; Regine, R.; King, J.B. Surgery for chronic Achilles tendinopathy yields worse results in nonathletic patients. Clin. J. Sport Med. 2006, 16, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Zhou, Z.; Taub, P.J.; Ramcharan, M.; Li, Y.; Akinbiyi, T.; Maharam, E.R.; Leong, D.J.; Laudier, D.M.; Ruike, T.; et al. BMP-12 treatment of adult mesenchymal stem cells in vitro augments tendon-like tissue formation and defect repair in vivo. PLoS ONE 2011, 6, e17531. [Google Scholar] [CrossRef] [PubMed]
- Lui, P.P.Y.; Wong, O.T.; Lee, Y.W. Application of tendon-derived stem cell sheet for the promotion of graft healing in anterior cruciate ligament reconstruction. Am. J. Sports Med. 2014, 42, 681–689. [Google Scholar] [CrossRef]
- Manning, C.N.; Martel, C.; Sakiyama-Elbert, S.E.; Silva, M.J.; Shah, S.; Gelberman, R.H.; Thomopoulos, S. Adipose-derived mesenchymal stromal cells modulate tendon fibroblast responses to macrophage-induced inflammation in vitro. Stem Cell Res. Ther. 2015, 6, 74. [Google Scholar] [CrossRef]
- Chen, H.-S.; Su, Y.-T.; Chan, T.-M.; Su, Y.-J.; Syu, W.-S.; Harn, H.-J.; Lin, S.-Z.; Chiu, S.-C. Human adipose-derived stem cells accelerate the restoration of tensile strength of tendon and alleviate the progression of rotator cuff injury in a rat model. Cell Transplant. 2015, 24, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.S.; Choi, Y.; Kim, H.-S.; Kim, H.O. Comparison of molecular profiles of human mesenchymal stem cells derived from bone marrow, umbilical cord blood, placenta and adipose tissue. Int. J. Mol. Med. 2016, 37, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Musina, R.A.; Bekchanova, E.S.; Belyavskii, A.V.; Sukhikh, G.T. Differentiation potential of mesenchymal stem cells of different origin. Bull. Exp. Biol. Med. 2006, 141, 147–151. [Google Scholar] [CrossRef]
- Guo, J.; Chan, K.-M.; Zhang, J.-F.; Li, G. Tendon-derived stem cells undergo spontaneous tenogenic differentiation. Exp. Cell Res. 2016, 341, 1–7. [Google Scholar] [CrossRef]
- Gaspar, D.; Spanoudes, K.; Holladay, C.; Pandit, A.; Zeugolis, D. Progress in cell-based therapies for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 240–256. [Google Scholar] [CrossRef] [PubMed]
- Docheva, D.; Müller, S.A.; Majewski, M.; Evans, C.H. Biologics for tendon repair. Adv. Drug Deliv. Rev. 2015, 84, 222–239. [Google Scholar] [CrossRef] [Green Version]
- Martinello, T.; Bronzini, I.; Perazzi, A.; Testoni, S.; De Benedictis, G.M.; Negro, A.; Caporale, G.; Mascarello, F.; Iacopetti, I.; Patruno, M. Effects of in vivo applications of peripheral blood-derived mesenchymal stromal cells (PB-MSCs) and platlet-rich plasma (PRP) on experimentally injured deep digital flexor tendons of sheep. J. Orthop. Res. 2013, 31, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.K.W.; Werling, N.J.; Dakin, S.G.; Alam, R.; Goodship, A.E.; Dudhia, J. Beneficial effects of autologous bone marrow-derived mesenchymal stem cells in naturally occurring tendinopathy. PLoS ONE 2013, 8, e75697. [Google Scholar] [CrossRef]
- Harris, M.T.; Butler, D.L.; Boivin, G.P.; Florer, J.B.; Schantz, E.J.; Wenstrup, R.J. Mesenchymal stem cells used for rabbit tendon repair can form ectopic bone and express alkaline phosphatase activity in constructs. J. Orthop. Res. 2004, 22, 998–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, P.P.Y.; Cheuk, Y.C.; Lee, Y.W.; Chan, K.M. Ectopic chondro-ossification and erroneous extracellular matrix deposition in a tendon window injury model. J. Orthop. Res. 2012, 30, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T.; Fox, P.M.; Woon, C.Y.; Farnebo, S.J.; Bronstein, J.A.; Behn, A.; Pham, H.; Chang, J. Human flexor tendon tissue engineering: In vivo effects of stem cell reseeding. Plast. Reconstr. Surg. 2013, 132, 567e–576e. [Google Scholar] [CrossRef]
- Deng, D.; Wang, W.; Wang, B.; Zhang, P.; Zhou, G.; Zhang, W.J.; Cao, Y.; Liu, W. Repair of Achilles tendon defect with autologous ASCs engineered tendon in a rabbit model. Biomaterials 2014, 35, 8801–8809. [Google Scholar] [CrossRef]
- Chiou, G.J.; Crowe, C.; McGoldrick, R.; Hui, K.; Pham, H.; Chang, J. Optimization of an injectable tendon hydrogel: The effects of platelet-rich plasma and adipose-derived stem cells on tendon healing in vivo. Tissue Eng. Part A 2015, 21, 1579–1586. [Google Scholar] [CrossRef]
- Kang, K.-K.; Lee, E.-J.; Kim, Y.-D.; Chung, M.-J.; Kim, J.-Y.; Kim, S.-Y.; Hwang, S.-K.; Jeong, K.-S. Vitamin C Improves Therapeutic Effects of Adipose-derived Stem Cell Transplantation in Mouse Tendonitis Model. Vivo Athens Greece 2017, 31, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Uysal, A.C.; Mizuno, H. Tendon regeneration and repair with adipose derived stem cells. Curr. Stem Cell Res. Ther. 2010, 5, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tao, X.; Chen, L.; Han, W.; Zhou, Y.; Tang, K. CTGF positively regulates BMP12 induced tenogenic differentiation of tendon stem cells and signaling. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2015, 35, 1831–1845. [Google Scholar] [CrossRef] [PubMed]
- Stanco, D.; Viganò, M.; Perucca Orfei, C.; Di Giancamillo, A.; Peretti, G.M.; Lanfranchi, L.; de Girolamo, L. Multidifferentiation potential of human mesenchymal stem cells from adipose tissue and hamstring tendons for musculoskeletal cell-based therapy. Regen. Med. 2015, 10, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Lui, P.P.Y.; Wong, O.T.; Lee, Y.W. Transplantation of tendon-derived stem cells pre-treated with connective tissue growth factor and ascorbic acid in vitro promoted better tendon repair in a patellar tendon window injury rat model. Cytotherapy 2016, 18, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Dale, T.P.; Mazher, S.; Webb, W.R.; Zhou, J.; Maffulli, N.; Chen, G.-Q.; El Haj, A.J.; Forsyth, N.R. Tenogenic Differentiation of Human Embryonic Stem Cells. Tissue Eng. Part A 2018, 24, 361–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarychta-Wiśniewska, W.; Burdzinska, A.; Kulesza, A.; Gala, K.; Kaleta, B.; Zielniok, K.; Siennicka, K.; Sabat, M.; Paczek, L. Bmp-12 activates tenogenic pathway in human adipose stem cells and affects their immunomodulatory and secretory properties. BMC Cell Biol. 2017, 18, 13. [Google Scholar] [CrossRef] [PubMed]
- Barsby, T.; Guest, D. Transforming growth factor beta3 promotes tendon differentiation of equine embryo-derived stem cells. Tissue Eng. Part A 2013, 19, 2156–2165. [Google Scholar] [CrossRef] [PubMed]
- Havis, E.; Bonnin, M.-A.; Esteves de Lima, J.; Charvet, B.; Milet, C.; Duprez, D. TGFβ and FGF promote tendon progenitor fate and act downstream of muscle contraction to regulate tendon differentiation during chick limb development. Dev. Camb. Engl. 2016, 143, 3839–3851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Wang, X.; Zhang, Y.; Carr, A.J.; Zhu, L.; Xia, Z.; Sabokbar, A. In vitro two-dimensional and three-dimensional tenocyte culture for tendon tissue engineering. J. Tissue Eng. Regen. Med. 2016, 10, E216–E226. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Wang, X.; Zhang, Y.; Rout, R.; Carr, A.J.; Zhu, L.; Xia, Z.; Sabokbar, A. Development of a refined tenocyte differentiation culture technique for tendon tissue engineering. Cells Tissues Organs 2013, 197, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Lee, F.Y.; Tarafder, S.; Kao, K.; Jun, Y.; Yang, G.; Mao, J.J. Harnessing endogenous stem/progenitor cells for tendon regeneration. J. Clin. Investig. 2015, 125, 2690–2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Ramcharan, M.; Zhou, Z.; Leong, D.J.; Akinbiyi, T.; Majeska, R.J.; Sun, H.B. The Role of Scleraxis in Fate Determination of Mesenchymal Stem Cells for Tenocyte Differentiation. Sci. Rep. 2015, 5, 13149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otabe, K.; Nakahara, H.; Hasegawa, A.; Matsukawa, T.; Ayabe, F.; Onizuka, N.; Inui, M.; Takada, S.; Ito, Y.; Sekiya, I.; et al. Transcription factor Mohawk controls tenogenic differentiation of bone marrow mesenchymal stem cells in vitro and in vivo. J. Orthop. Res. 2015, 33, 1–8. [Google Scholar] [CrossRef]
- Carlberg, A.L.; Tuan, R.S.; Hall, D.J. Regulation of scleraxis function by interaction with the bHLH protein E47. Mol. Cell Biol. Res. Commun. MCBRC 2000, 3, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, R.; Chyung, J.H.; Murtaugh, L.C.; Brent, A.E.; Rosen, V.; Olson, E.N.; Lassar, A.; Tabin, C.J. Analysis of the tendon cell fate using Scleraxis, a specific marker for tendons and ligaments. Dev. Camb. Engl. 2001, 128, 3855–3866. [Google Scholar]
- Shukunami, C.; Takimoto, A.; Oro, M.; Hiraki, Y. Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev. Biol. 2006, 298, 234–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogány, G.; Vogel, K.G. The interaction of decorin core protein fragments with type I collagen. Biochem. Biophys. Res. Commun. 1992, 189, 165–172. [Google Scholar] [CrossRef]
- Kalamajski, S.; Aspberg, A.; Oldberg, A. The decorin sequence SYIRIADTNIT binds collagen type I. J. Biol. Chem. 2007, 282, 16062–16067. [Google Scholar] [CrossRef]
- Orgel, J.P.; Eid, A.; Antipova, O.; Bella, J.; Scott, J.E. Decorin core protein (decoron) shape complements collagen fibril surface structure and mediates its binding. PLoS ONE 2009, 4, e7028. [Google Scholar] [CrossRef]
- Reese, S.P.; Underwood, C.J.; Weiss, J.A. Effects of decorin proteoglycan on fibrillogenesis, ultrastructure, and mechanics of type I collagen gels. Matrix Biol. J. Int. Soc. Matrix Biol. 2013, 32, 414–423. [Google Scholar] [CrossRef] [Green Version]
- Pryce, B.A.; Watson, S.S.; Murchison, N.D.; Staverosky, J.A.; Dünker, N.; Schweitzer, R. Recruitment and maintenance of tendon progenitors by TGFbeta signaling are essential for tendon formation. Dev. Camb. Engl. 2009, 136, 1351–1361. [Google Scholar] [CrossRef]
- Barsby, T.; Bavin, E.P.; Guest, D.J. Three-dimensional culture and transforming growth factor beta3 synergistically promote tenogenic differentiation of equine embryo-derived stem cells. Tissue Eng. Part A 2014, 20, 2604–2613. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Guo, J.; Wu, T.-Y.; Chen, X.; Xu, L.-L.; Lin, S.-E.; Sun, Y.-X.; Chan, K.-M.; Ouyang, H.; Li, G. Stepwise Differentiation of Mesenchymal Stem Cells Augments Tendon-Like Tissue Formation and Defect Repair In Vivo. Stem Cells Transl. Med. 2016, 5, 1106–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrand, A.; Romarís, M.; Rasmussen, L.M.; Heinegård, D.; Twardzik, D.R.; Border, W.A.; Ruoslahti, E. Interaction of the small interstitial proteoglycans biglycan, decorin and fibromodulin with transforming growth factor beta. Biochem. J. 1994, 302, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.-C.; Wong, Y.-P.; Cheuk, Y.-C.; Lee, K.-M.; Chan, K.-M. TGF-beta1 reverses the effects of matrix anchorage on the gene expression of decorin and procollagen type I in tendon fibroblasts. Clin. Orthop. 2005, 226–232. [Google Scholar] [CrossRef]
- Ferdous, Z.; Wei, V.M.; Iozzo, R.; Höök, M.; Grande-Allen, K.J. Decorin-transforming growth factor- interaction regulates matrix organization and mechanical characteristics of three-dimensional collagen matrices. J. Biol. Chem. 2007, 282, 35887–35898. [Google Scholar] [CrossRef]
- Lee, C.H.; Moioli, E.K.; Mao, J.J. Fibroblastic differentiation of human mesenchymal stem cells using connective tissue growth factor. IEEE Eng. Med. Biol. Soc. 2006, 1, 775–778. [Google Scholar] [CrossRef]
- Ni, M.; Rui, Y.F.; Tan, Q.; Liu, Y.; Xu, L.L.; Chan, K.M.; Wang, Y.; Li, G. Engineered scaffold-free tendon tissue produced by tendon-derived stem cells. Biomaterials 2013, 34, 2024–2037. [Google Scholar] [CrossRef]
- Moores, J. Vitamin C: A wound healing perspective. Br. J. Community Nurs. 2013, 18, S6–S11. [Google Scholar] [CrossRef]
- Piersma, B.; Wouters, O.Y.; de Rond, S.; Boersema, M.; Gjaltema, R.A.F.; Bank, R.A. Ascorbic acid promotes a TGFβ1-induced myofibroblast phenotype switch. Physiol. Rep. 2017, 5. [Google Scholar] [CrossRef]
- Gelberman, R.H.; Shen, H.; Kormpakis, I.; Rothrauff, B.; Yang, G.; Tuan, R.S.; Xia, Y.; Sakiyama-Elbert, S.; Silva, M.J.; Thomopoulos, S. Effect of adipose-derived stromal cells and BMP12 on intrasynovial tendon repair: A biomechanical, biochemical, and proteomics study. J. Orthop. Res. 2016, 34, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Hu, X.; Zhang, X.; Zhu, J.; Zhang, J.; Fu, X.; Duan, X.; Ao, Y.; Zhou, C. Different tenogenic differentiation capacities of different mesenchymal stem cells in the presence of BMP-12. J. Transl. Med. 2015, 13, 200. [Google Scholar] [CrossRef] [PubMed]
- Alberton, P.; Popov, C.; Prägert, M.; Kohler, J.; Shukunami, C.; Schieker, M.; Docheva, D. Conversion of human bone marrow-derived mesenchymal stem cells into tendon progenitor cells by ectopic expression of scleraxis. Stem Cells Dev. 2012, 21, 846–858. [Google Scholar] [CrossRef] [PubMed]
- De Girolamo, L.; Stanco, D.; Salvatori, L.; Coroniti, G.; Arrigoni, E.; Silecchia, G.; Russo, M.A.; Niada, S.; Petrangeli, E.; Brini, A.T. Stemness and osteogenic and adipogenic potential are differently impaired in subcutaneous and visceral adipose derived stem cells (ASCs) isolated from obese donors. Int. J. Immunopathol. Pharmacol. 2013, 26, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.-F.; Lui, P.P.Y.; Li, G.; Fu, S.C.; Lee, Y.W.; Chan, K.M. Isolation and characterization of multipotent rat tendon-derived stem cells. Tissue Eng. Part A 2010, 16, 1549–1558. [Google Scholar] [CrossRef] [PubMed]
- De Girolamo, L.; Stanco, D.; Galliera, E.; Viganò, M.; Colombini, A.; Setti, S.; Vianello, E.; Corsi Romanelli, M.M.; Sansone, V. Low frequency pulsed electromagnetic field affects proliferation, tissue-specific gene expression, and cytokines release of human tendon cells. Cell Biochem. Biophys. 2013, 66, 697–708. [Google Scholar] [CrossRef]
- Viganò, M.; Perucca Orfei, C.; de Girolamo, L.; Pearson, J.R.; Ragni, E.; De Luca, P.; Colombini, A. Housekeeping Gene Stability in Human Mesenchymal Stem and Tendon Cells Exposed to Tenogenic Factors. Tissue Eng. Part C Methods 2018, 24, 360–367. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Growth Factors and Supplements | ||||||
|---|---|---|---|---|---|---|---|
| FBS | BMP-12 (50 ng/mL) | IGF-1 (50 ng/mL) | CTGF (100 ng/mL) | TGF-β3 (20 ng/mL) | AA (25 µg/mL) | b-FGF (5 ng/mL) | |
| BMP-12 | 1% | + | - | - | - | + | + |
| MIX 1 | 1% | + | + | + | + | + | + |
| MIX 2 | 1% | - | + | + | + | + | + |
| MIX 3 | 1% | + | + | + | - | + | + |
| MIX 4 | 1% | + | - | + | + | + | + |
| MIX 5 | 1% | + | + | - | + | + | + |
| Maintenance Medium | 1% | - | - | - | - | + | + |
| Low Serum Medium | 1% | - | - | - | - | - | - |
| Complete Medium | 10% | - | - | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perucca Orfei, C.; Viganò, M.; Pearson, J.R.; Colombini, A.; De Luca, P.; Ragni, E.; Santos-Ruiz, L.; De Girolamo, L. In Vitro Induction of Tendon-Specific Markers in Tendon Cells, Adipose- and Bone Marrow-Derived Stem Cells is Dependent on TGFβ3, BMP-12 and Ascorbic Acid Stimulation. Int. J. Mol. Sci. 2019, 20, 149. https://doi.org/10.3390/ijms20010149
Perucca Orfei C, Viganò M, Pearson JR, Colombini A, De Luca P, Ragni E, Santos-Ruiz L, De Girolamo L. In Vitro Induction of Tendon-Specific Markers in Tendon Cells, Adipose- and Bone Marrow-Derived Stem Cells is Dependent on TGFβ3, BMP-12 and Ascorbic Acid Stimulation. International Journal of Molecular Sciences. 2019; 20(1):149. https://doi.org/10.3390/ijms20010149
Chicago/Turabian StylePerucca Orfei, Carlotta, Marco Viganò, John R. Pearson, Alessandra Colombini, Paola De Luca, Enrico Ragni, Leonor Santos-Ruiz, and Laura De Girolamo. 2019. "In Vitro Induction of Tendon-Specific Markers in Tendon Cells, Adipose- and Bone Marrow-Derived Stem Cells is Dependent on TGFβ3, BMP-12 and Ascorbic Acid Stimulation" International Journal of Molecular Sciences 20, no. 1: 149. https://doi.org/10.3390/ijms20010149