Idi1 and Hmgcs2 Are Affected by Stretch in HL-1 Atrial Myocytes

 ,
,
Abstract
:1. Introduction
2. Results
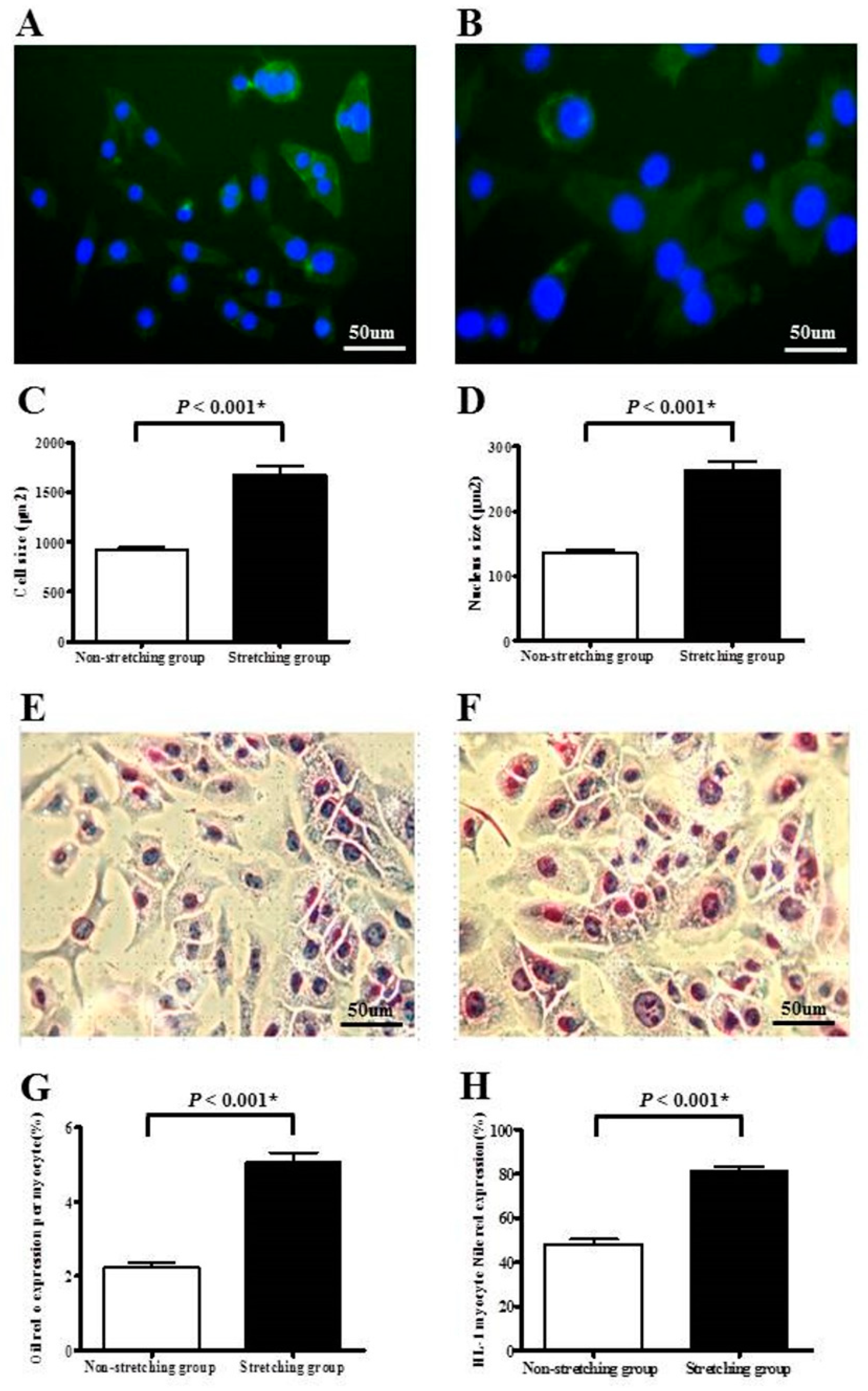
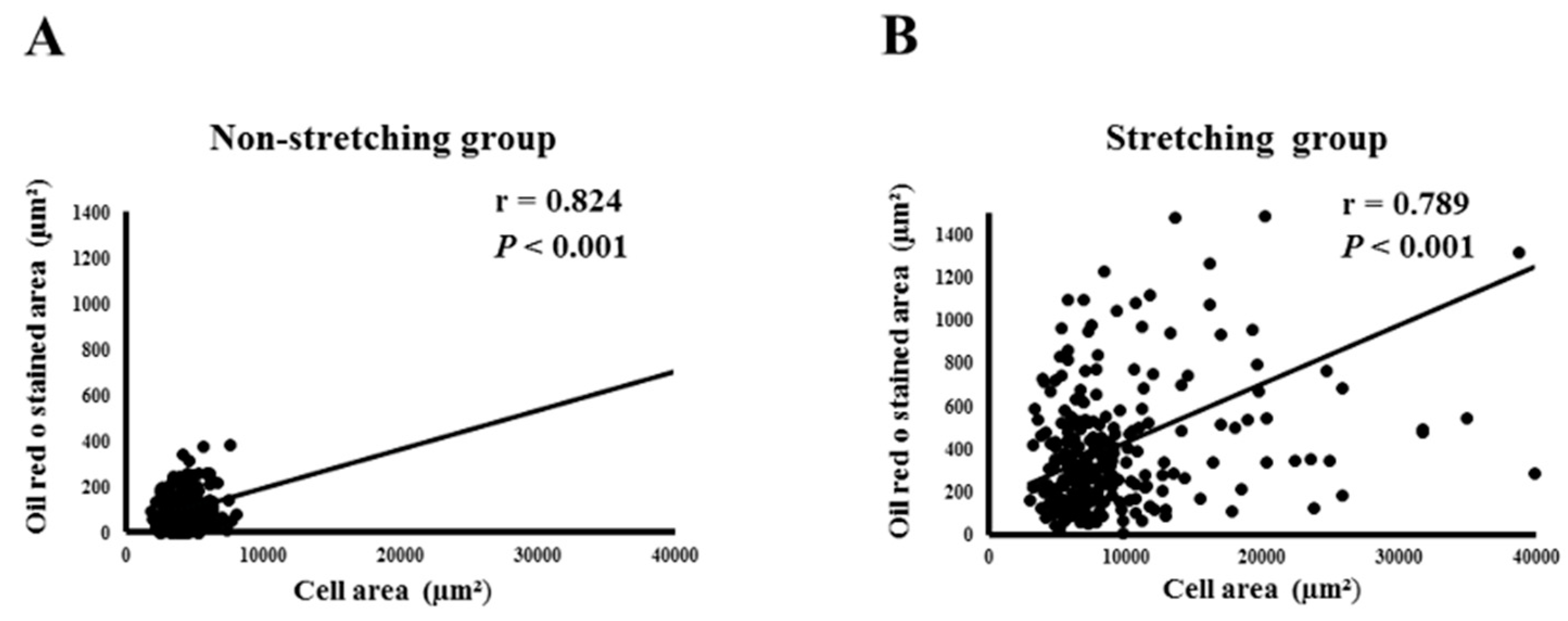
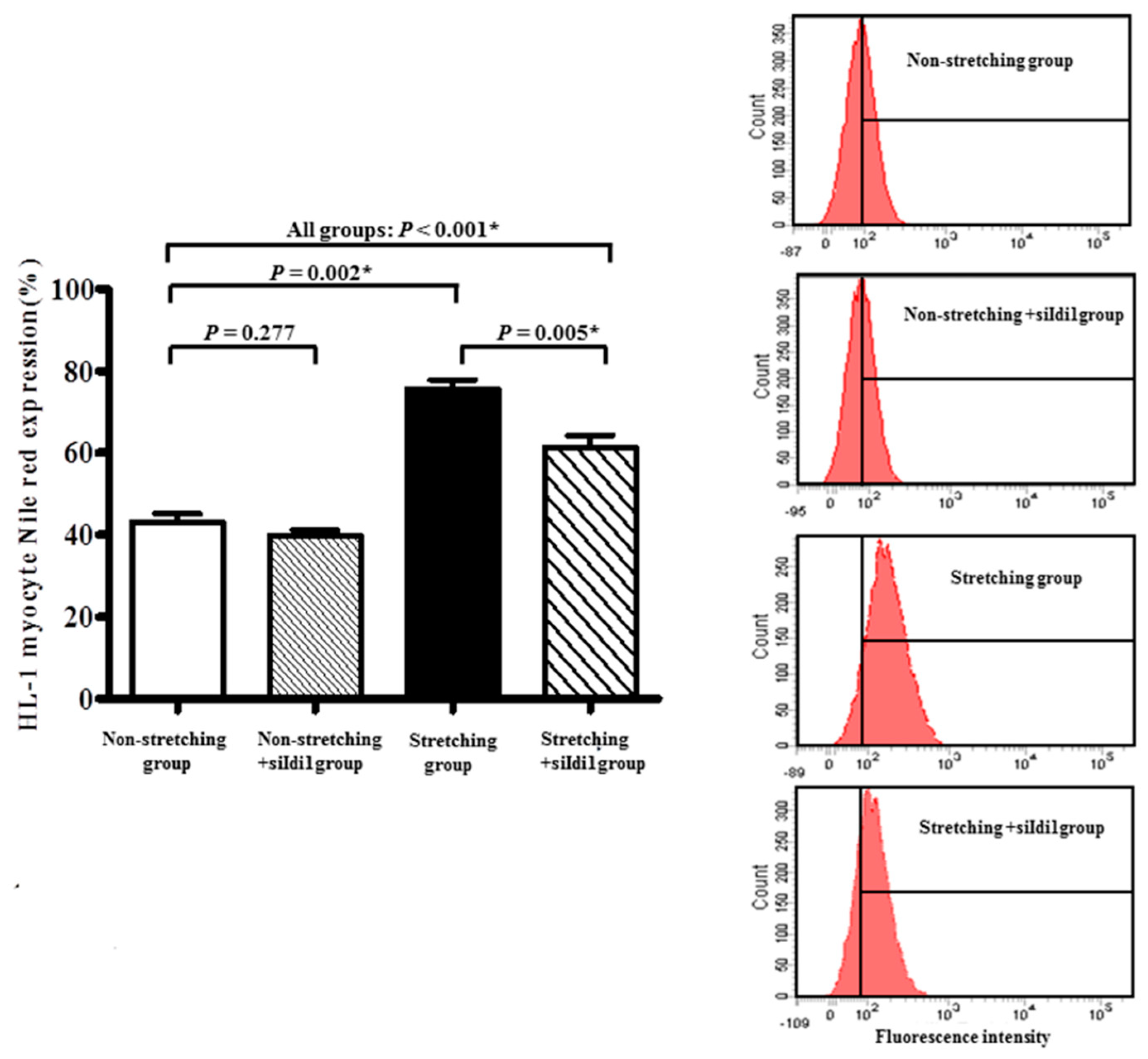
2.1. Morphological Changes and Lipid Expression after Mechanical Stretching of HL-1 Atrial Myocytes
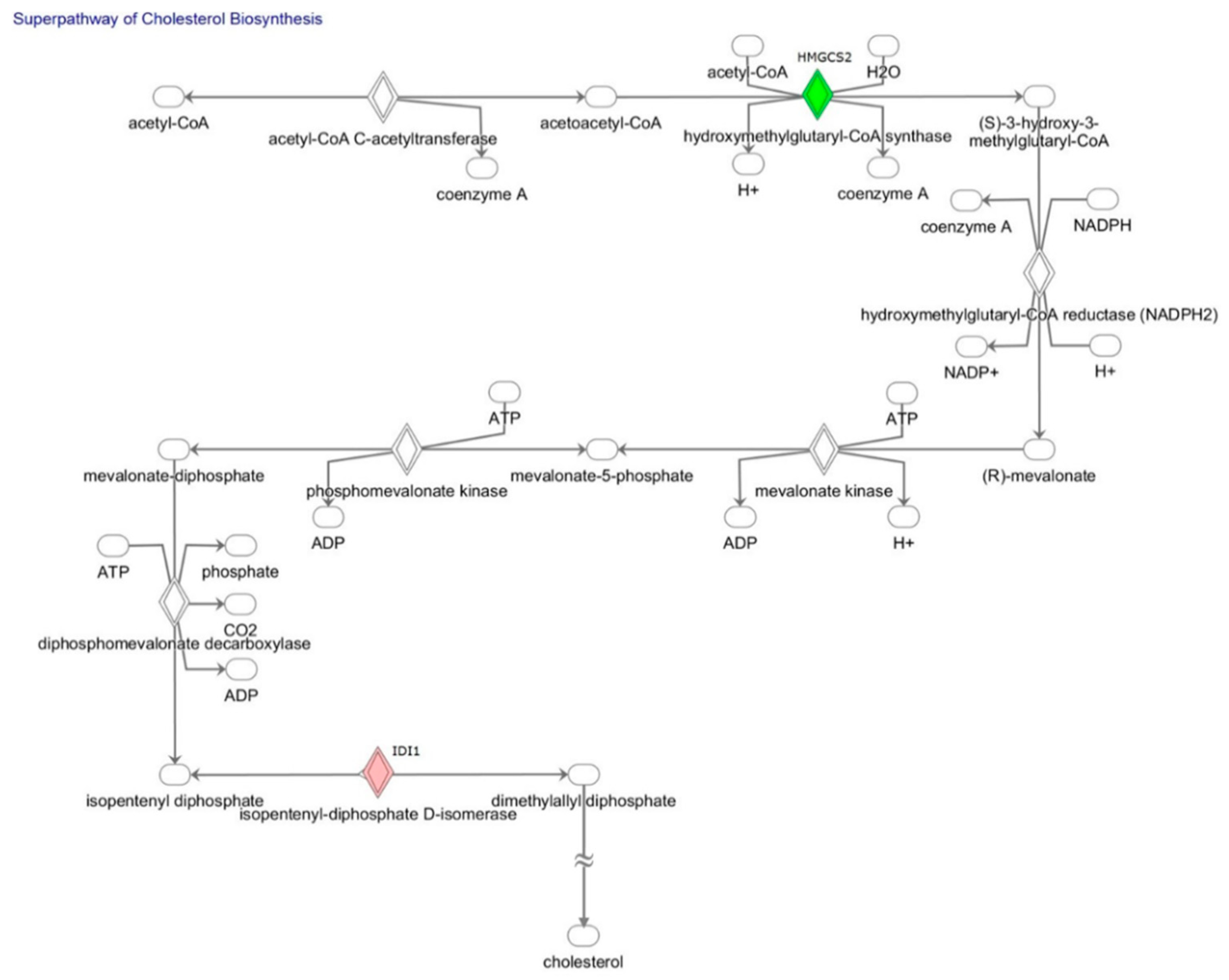
2.2. Gene Expression Analyses of Fatty Acid Metabolism, Lipoprotein Signaling, and Cholesterol Metabolism: Comparison of Stretched and Non-Stretched HL-1 Atrial Myocytes
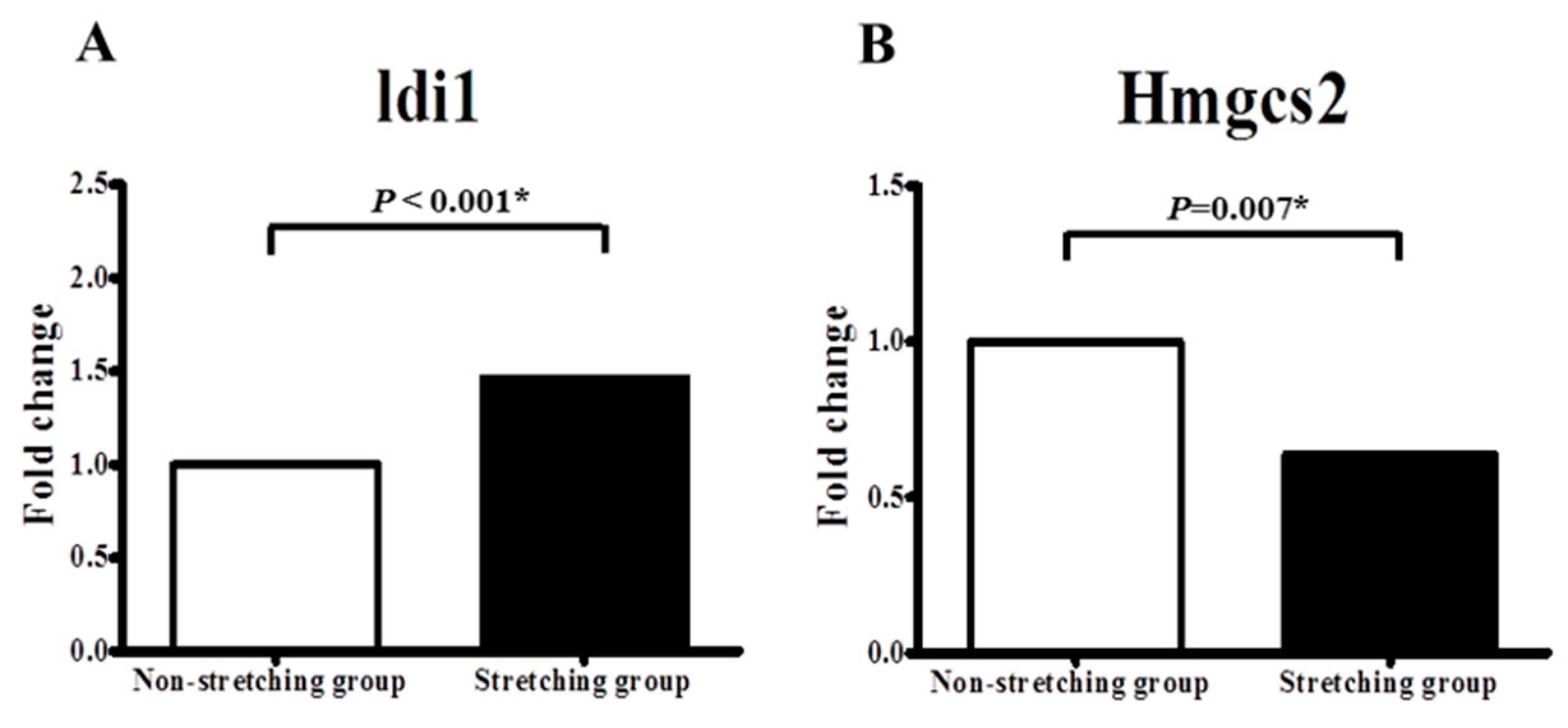
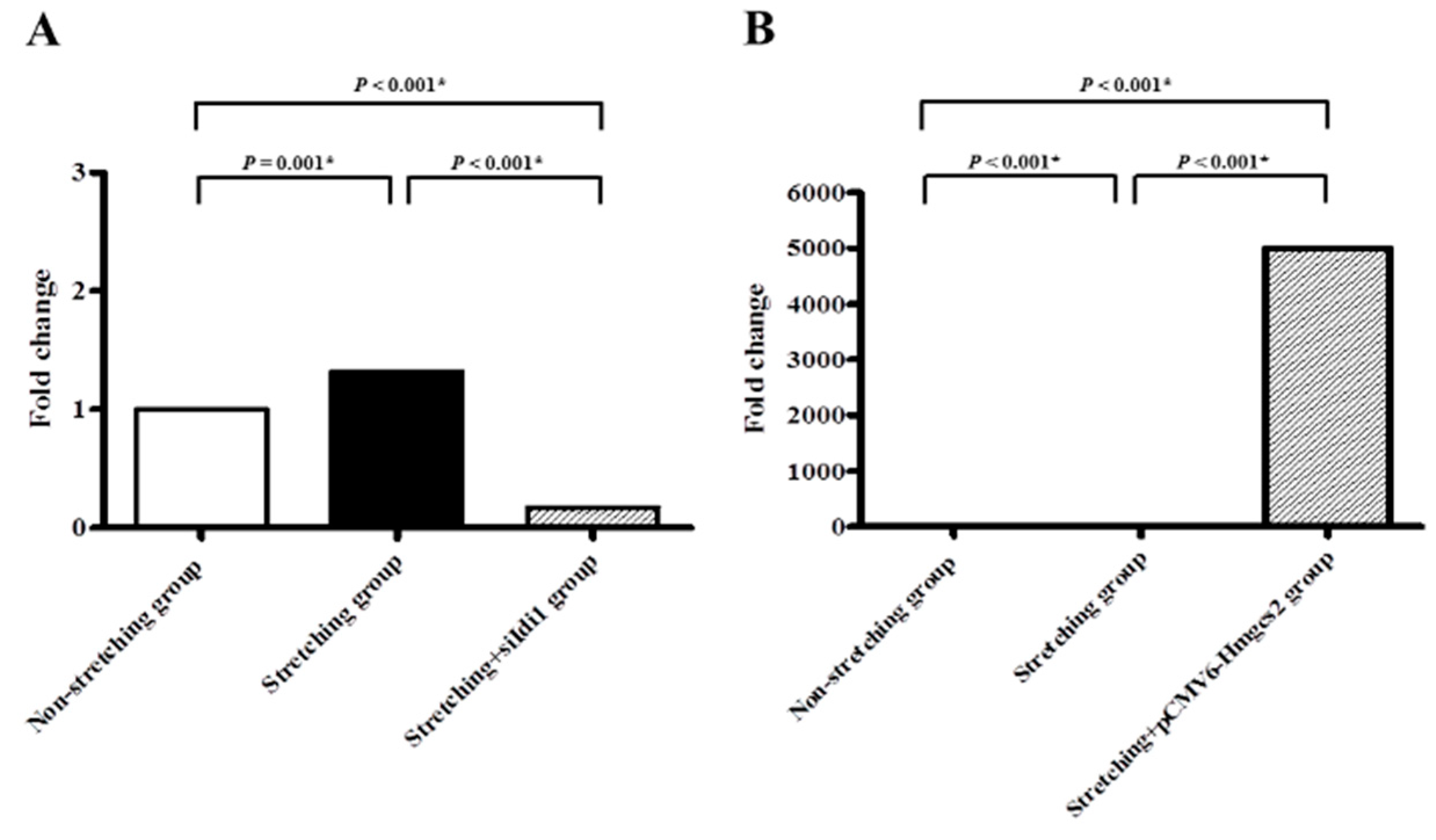
2.3. Quantitative PCR Validation of Idi1 and Hmgcs2 mRNAs in the Stretched vs. Non-Stretched HL-1 Atrial Myocytes
2.4. The Effect on Lipid Expression of Silencing the Idi1 Gene and Genetic Modification of the Hmgcs2 Gene, after Mechanical Stretching of HL-1 Atrial Myocytes
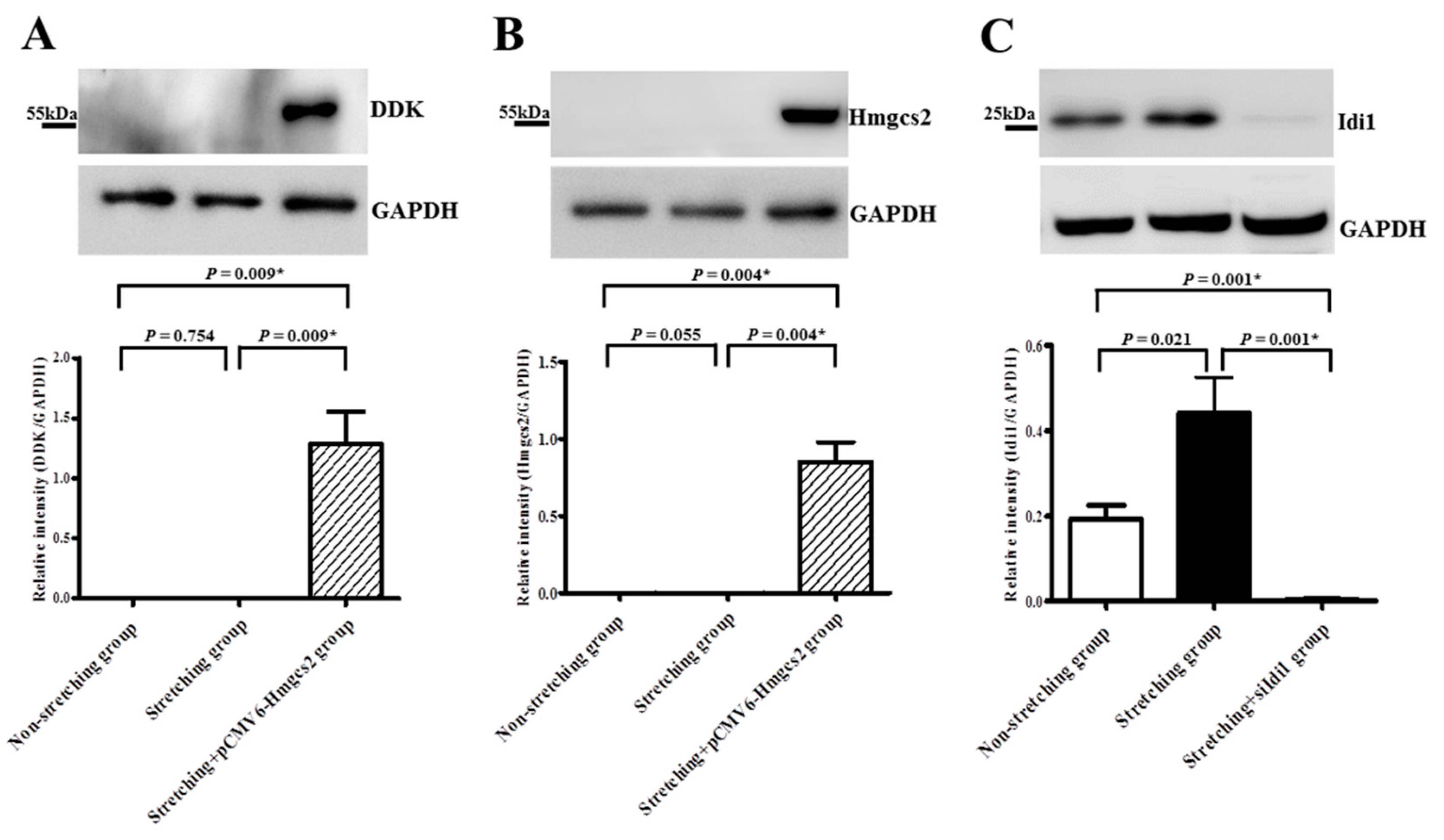
2.5. Immunoblot Analysis Confirmed Upregulation of Idi1 and Downregulation of Hmgcs2 by Stretching at the Translational Level
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Mechanical Stretching of Atrial Myocytes
4.2. Immunofluorescence Staining
4.3. Oil Red O Staining
4.4. Measurement of Intracellular Lipid by Flow Cytometry
4.5. PCR Assay and Data Processing
4.6. Quantitative Determination of RNAs by Real-Time PCR
4.7. RNA Interference and In Vitro siRNA Transfection
4.8. Plasmid Transfection
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Iung, B.; Gohlke-Bärwolf, C.; Tornos, P.; Tribouilloy, C.; Hall, R.; Butchart, E.; Vahanian, A.; Working Group on Valvular Heart Disease. Recommendations on the management of the asymptomatic patient with valvular heart disease. Eur. Heart J. 2002, 23, 1253–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, C.M. Timing of surgery in mitral regurgitation. Heart 2003, 89, 100–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts-Thomson, K.C.; Stevenson, I.; Kistler, P.M.; Haqqani, H.M.; Spence, S.J.; Goldblatt, J.C.; Sanders, P.; Kalman, J.M. The role of chronic atrial stretch and atrial fibrillation on posterior left atrial wall conduction. Heart Rhythm 2009, 6, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- De Jong, A.M.; Maass, A.H.; Oberdorf-Maass, S.U.; Van Veldhuisen, D.J.; Van Gilst, W.H.; Van Gelder, I.C. Mechanisms of atrial structural changes caused by stretch occurring before and during early atrial fibrillation. Cardiovasc. Res. 2011, 89, 754–765. [Google Scholar] [CrossRef] [PubMed]
- Goette, A.; Kalman, J.M.; Aguinaga, L.; Akar, J.; Cabrera, J.A.; Chen, S.A.; Chugh, S.S.; Corradi, D.; D’Avila, A.; Dobrev, D.; et al. EHRA/HRS/APHRS/SOLAECE expert consensus on Atrial cardiomyopathies: Definition, characterisation, and clinical implication. J. Arrhythm. 2016, 32, 247–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchhof, P.; Benussi, S.; Kotecha, D.; Ahlsson, A.; Atar, D.; Casadei, B.; Castella, M.; Diener, H.C.; Heidbuchel, H.; Hendriks, J.; et al. 2016 ESC Guidelines for the management of atrial fibrillation developed in collaboration with EACTS. Eur. Heart J. 2016, 37, 2893–2962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grygiel-Górniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications—A review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Finck, B.N. The PPAR regulatory system in cardiac physiology and disease. Cardiovasc. Res. 2007, 73, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, J.; Moller, D.E. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Sundseth, S.S.; Jones, S.A.; Brown, P.J.; Wisely, G.B.; Koble, C.S.; Devchand, P.; Whali, W.; Willson, T.M.; Lenhard, J.M.; et al. Fatty acids and eicosanoids regulate gene expression through direct interactions with peroxisome proliferator- activated receptors alpha and gamma. Proc. Natl. Acad. Sci. USA 1997, 94, 4318–4323. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, D.M.; Rinaldo, P.; Rhead, W.J.; Tian, L.; Millington, D.S.; Vockley, J.; Hamm, D.A.; Brix, A.E.; Lindsey, J.R.; Pinkert, C.A.; et al. Targeted disruption of mouse long-chain acyl-CoA dehydrogenase gene reveals crucial roles for fatty acid oxidation. Proc. Natl. Acad. Sci. USA 1998, 95, 15592–15597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, H.C.; Kovacs, A.; Ford, D.A.; Hsu, F.F.; Garcia, R.; Herrero, P.; Saffitz, J.E.; Schaffer, J.E. A novel mouse model of lipotoxic cardiomyopathy. J. Clin. Investig. 2001, 107, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.C.; Chang, J.P.; Chang, T.H.; Hsu, S.D.; Huang, H.D.; Ho, W.C.; Wang, F.S.; Hsiao, C.C.; Liu, W.H. Unraveling regulatory mechanisms of atrial remodeling of mitral regurgitation pigs by gene expression profiling analysis: Role of type I angiotensin II receptor antagonist. Transl. Res. 2015, 165, 599–620. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Chang, J.P.; Lin, Y.S.; Pan, K.L.; Ho, W.C.; Liu, W.H.; Chang, T.H.; Huang, Y.K.; Fang, C.Y.; Chen, C.J. Deciphering the gene expression profile of peroxisome proliferator-activated receptor signaling pathway in the left atria of patients with mitral regurgitation. J. Transl. Med. 2016, 14, 157. [Google Scholar] [CrossRef] [PubMed]
- Kijima, K.; Matsubara, H.; Murasawa, S.; Maruyama, K.; Mori, Y.; Ohkubo, N.; Komuro, I.; Yazaki, Y.; Iwasaka, T.; Inada, M. Mechanical stretch induces enhanced expression of angiotensin II receptor subtypes in neonatal rat cardiac myocytes. Circ. Res. 1996, 79, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.P.; Chen, M.C.; Liu, W.H.; Lin, Y.S.; Huang, Y.K.; Pan, K.L.; Ho, W.C.; Fang, C.Y.; Chen, C.J.; Chen, H.C. Mitochondrial apoptotic pathway activation in the atria of heart failure patients due to mitral and tricuspid regurgitation. Exp. Mol. Pathol. 2015, 99, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.P.; Chen, M.C.; Liu, W.H.; Yang, C.H.; Chen, C.J.; Chen, Y.L.; Pan, K.L.; Tsai, T.H.; Chang, H.W. Atrial myocardial nox2 containing NADPH oxidase activity contribution to oxidative stress in mitral regurgitation: Potential mechanism for atrial remodeling. Cardiovasc. Pathol. 2011, 20, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Kolwicz, S.C., Jr.; Tian, R. Glucose metabolism and cardiac hypertrophy. Cardiovasc. Res. 2011, 90, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wende, A.R.; Brahma, M.K.; McGinnis, G.R.; Young, M.E. Metabolic origins of heart failure. JACC Basic Transl. Sci. 2017, 2, 297–310. [Google Scholar] [CrossRef] [PubMed]
- Lionetti, V.; Stanley, W.C.; Recchia, F.A. Modulating fatty acid oxidation in heart failure. Cardiovasc. Res. 2011, 90, 202–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kada, K.; Yasui, K.; Naruse, K.; Kamiya, K.; Kodama, I.; Toyama, J. Orientation change of cardiocytes induced by cyclic stretch stimulation: Time dependency and involvement of protein kinases. J. Mol. Cell Cardiol. 1999, 31, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Frawley, L.E.; Orr-Weaver, T.L. Polyploidy. Curr. Biol. 2015, 25, R353–R358. [Google Scholar] [CrossRef] [PubMed]
- Mazein, A.; Watterson, S.; Hsieh, W.Y.; Griffiths, W.J.; Ghazal, P. A comprehensive machine-readable view of the mammalian cholesterol biosynthesis pathway. Biochem. Pharmacol. 2013, 86, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthelot, K.; Estevez, Y.; Deffieux, A.; Peruch, F. Isopentenyl diphosphate isomerase: A checkpoint to isoprenoid biosynthesis. Biochimie 2012, 94, 1621–1634. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.W.; Chou, H.C.; Lin, S.T.; Chen, Y.H.; Chang, Y.J.; Chen, L.; Chan, H.L. Cardioprotective effects of quercetin in cardiomyocyte under ischemia/reperfusion injury. Evid. Based Complement. Altern. Med. 2013, 2013, 364519. [Google Scholar] [CrossRef] [PubMed]
- Vilà-Brau, A.; De Sousa-Coelho, A.L.; Mayordomo, C.; Haro, D.; Marrero, P.F. Human HMGCS2 regulates mitochondrial fatty acid oxidation and FGF21 expression in HepG2 cell line. J. Biol. Chem. 2011, 286, 20423–20430. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Fold Change (Stretched/Non-Stretched) | p Value |
|---|---|---|
| Upregulated (fold change > 1.5 and/or p < 0.05) | ||
| Olr1 | 2.8307 | |
| Nr1h4 | 2.4049 | |
| Fabp2 | 2.2812 | |
| Prkag3 | 2.2569 | |
| Slc27a5 | 1.7197 | |
| Fabp6 | 1.5525 | |
| Idi1 | 1.4388 | <0.04 |
| Downregulated (fold change < 0.67) | ||
| Apoa4 | 0.3939 | |
| Hmgcs2 | 0.4057 | |
| Acsm4 | 0.4868 | |
| Apol8 | 0.5564 | |
| Fabp1 | 0.5858 | |
| Srebf1 | 0.5859 | |
| Acox2 | 0.649 | |
| Acsl6 | 0.6512 | |
| Gk | 0.6589 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Hmgcs2 | CAGGAAACTTCGCTCACACC | GGAGCAGGAGGGATTGTAGA |
| Idi1 | CGAGCGATTGGATATGCTG | AATGTCTGATCTGACCTAGAACACAG |
| Gusb | GATGTGGTCTGTGGCCAAT | TGTGGGTGATCAGCGTCTT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, C.-Y.; Chen, M.-C.; Chang, T.-H.; Wu, C.-C.; Chang, J.-P.; Huang, H.-D.; Ho, W.-C.; Wang, Y.-Z.; Pan, K.-L.; Lin, Y.-S.; et al. Idi1 and Hmgcs2 Are Affected by Stretch in HL-1 Atrial Myocytes. Int. J. Mol. Sci. 2018, 19, 4094. https://doi.org/10.3390/ijms19124094
Fang C-Y, Chen M-C, Chang T-H, Wu C-C, Chang J-P, Huang H-D, Ho W-C, Wang Y-Z, Pan K-L, Lin Y-S, et al. Idi1 and Hmgcs2 Are Affected by Stretch in HL-1 Atrial Myocytes. International Journal of Molecular Sciences. 2018; 19(12):4094. https://doi.org/10.3390/ijms19124094
Chicago/Turabian StyleFang, Chih-Yuan, Mien-Cheng Chen, Tzu-Hao Chang, Chia-Chen Wu, Jen-Ping Chang, Hsien-Da Huang, Wan-Chun Ho, Yi-Zhen Wang, Kuo-Li Pan, Yu-Sheng Lin, and et al. 2018. "Idi1 and Hmgcs2 Are Affected by Stretch in HL-1 Atrial Myocytes" International Journal of Molecular Sciences 19, no. 12: 4094. https://doi.org/10.3390/ijms19124094