Endovascular Interventions Permit Isolation of Endothelial Colony-Forming Cells from Peripheral Blood


Abstract
:
1. Introduction
2. Results
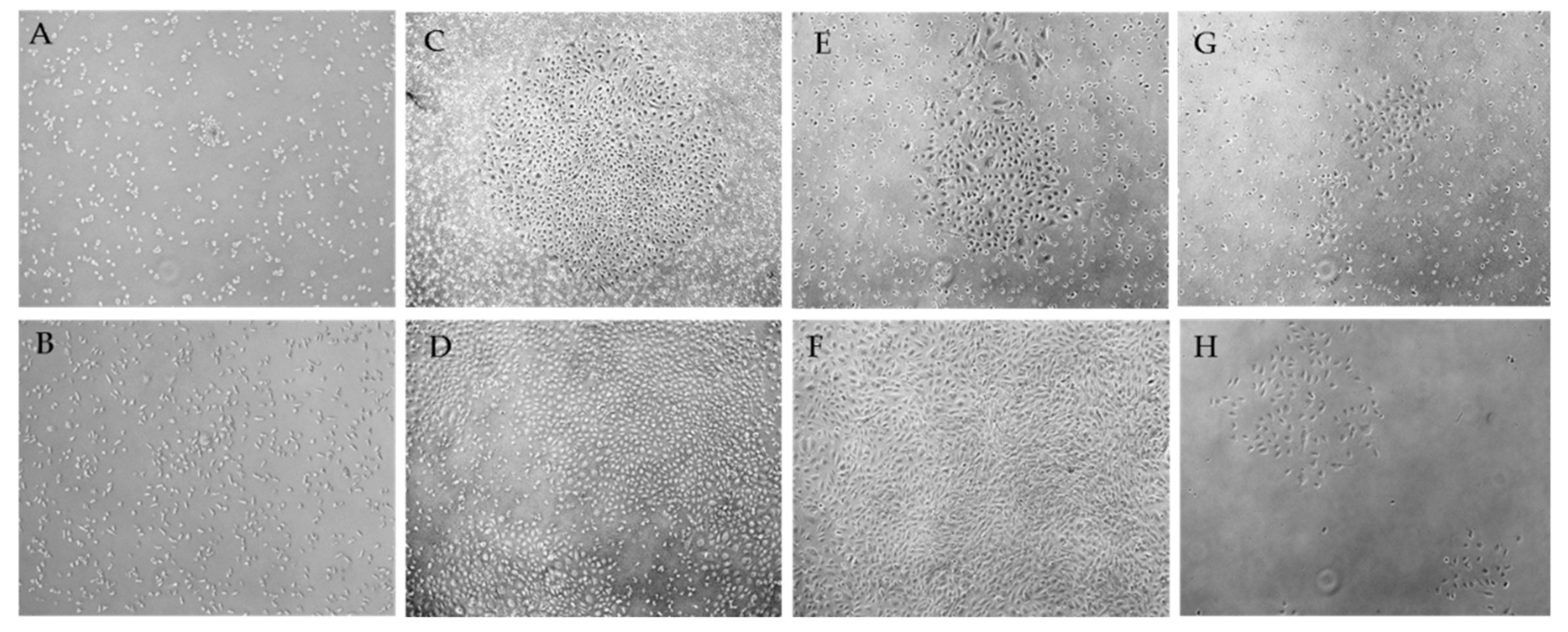
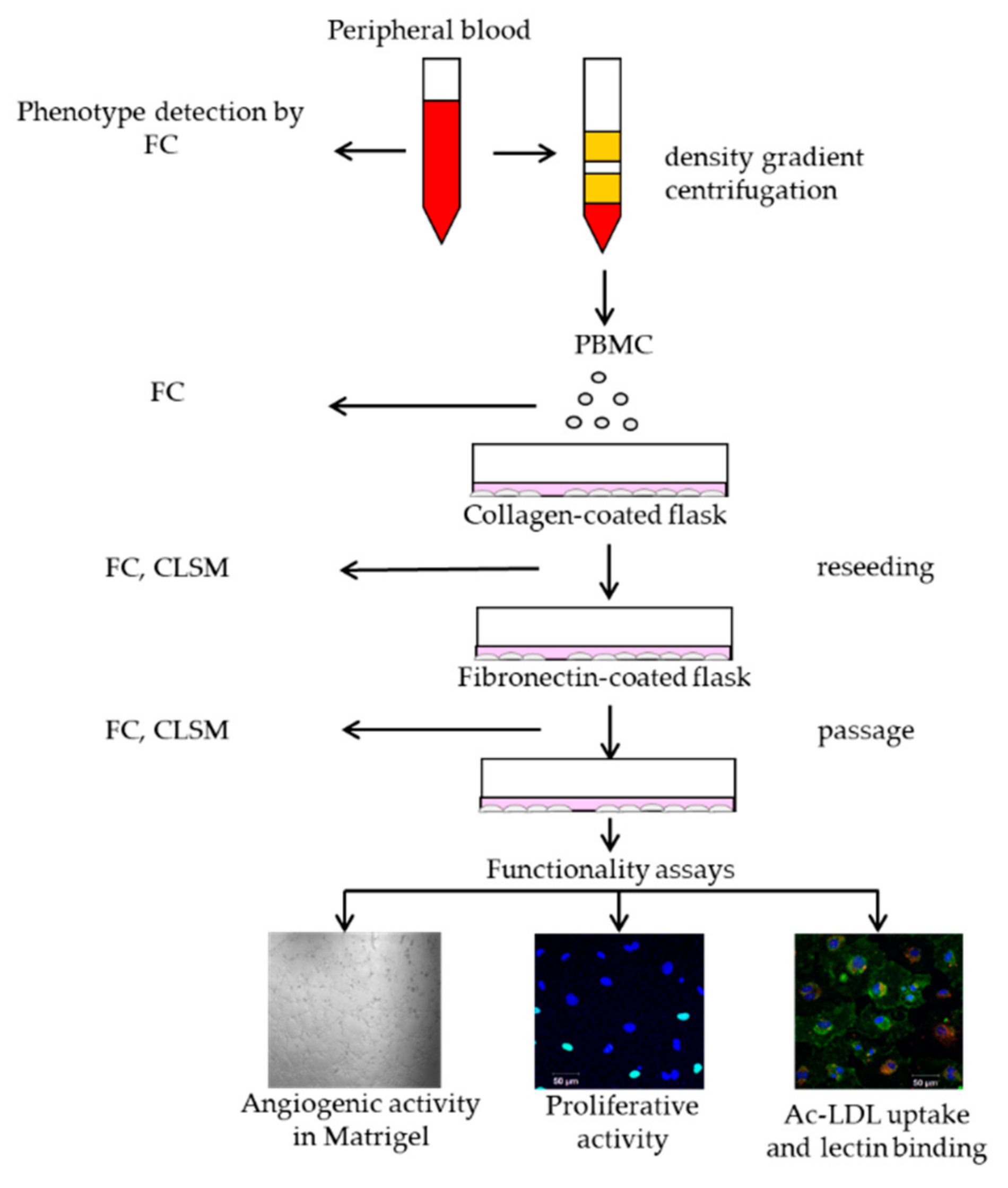
2.1. Morphological Patterns of Obtained Cell Cultures
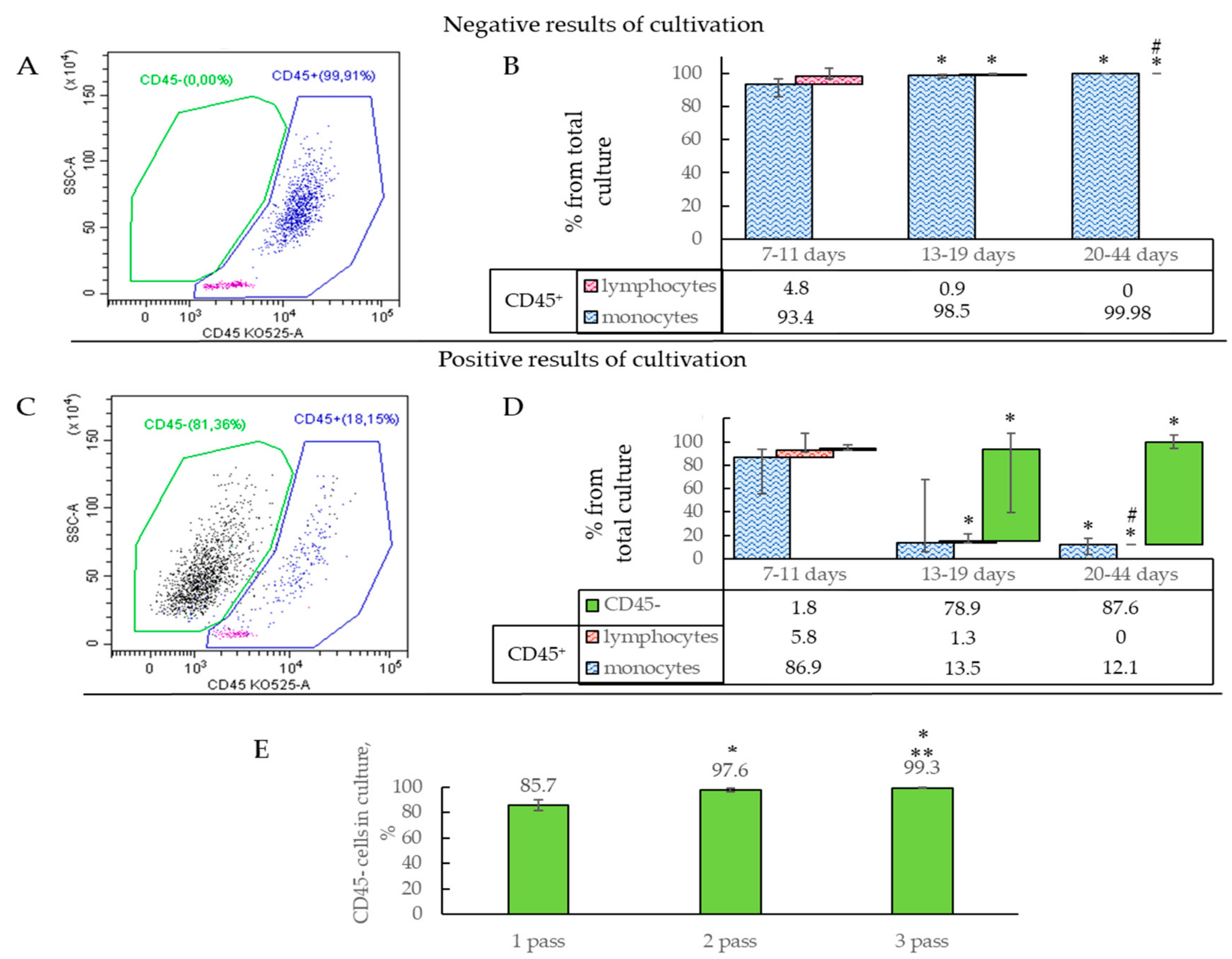
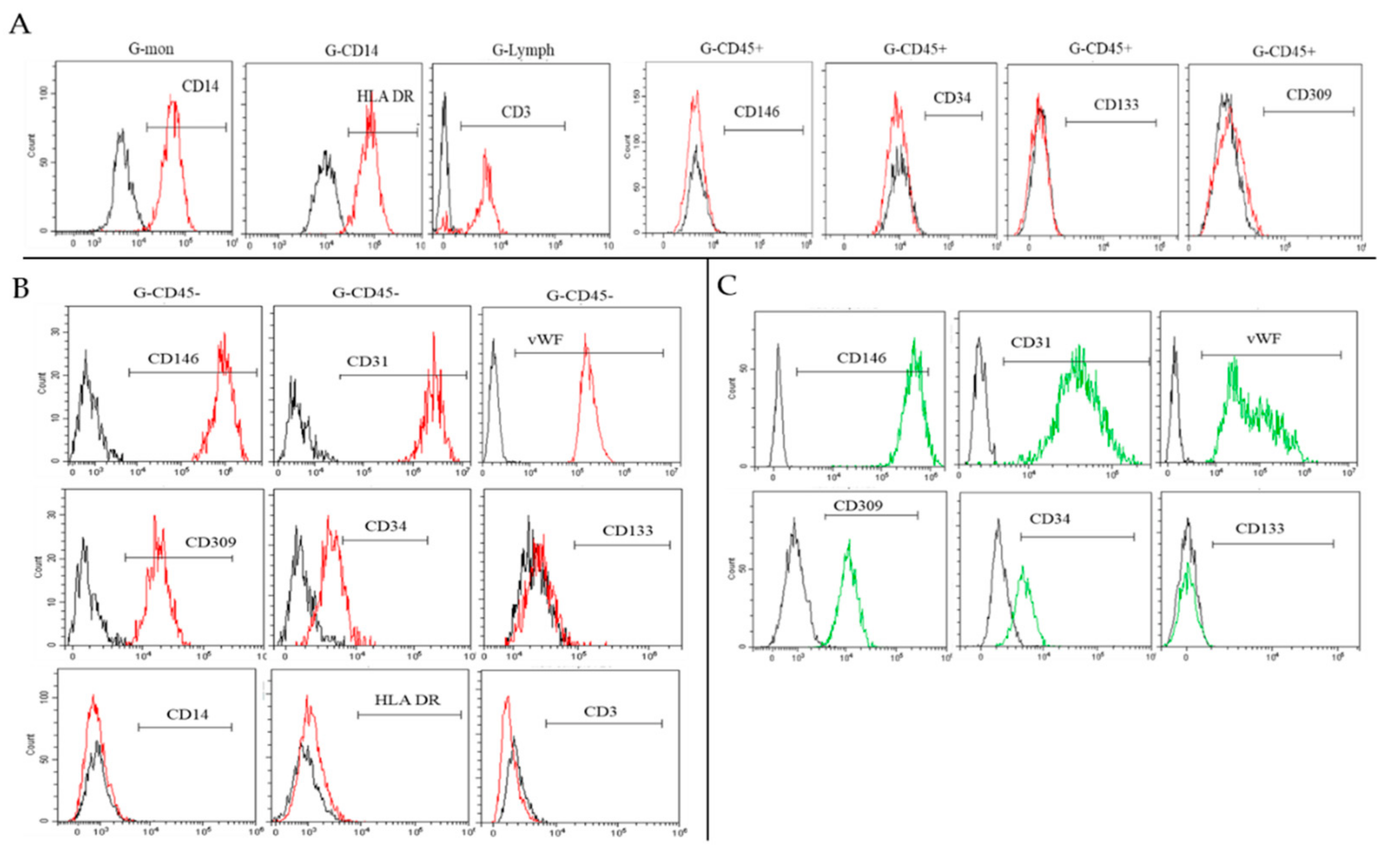
2.2. Characterization of Isolated Cells
2.3. Functional Properties of the Selected ECFC Populations
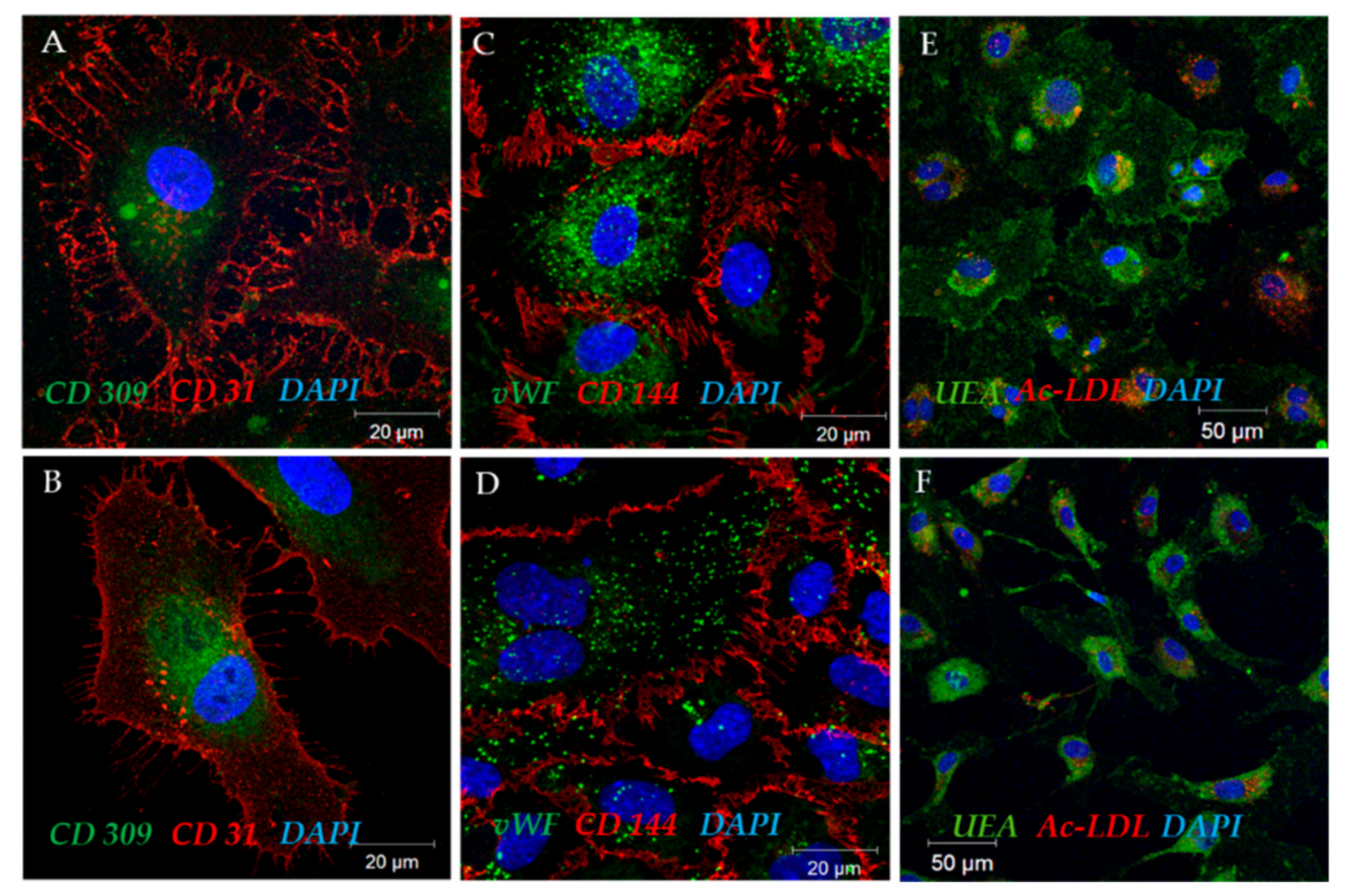
2.3.1. Uptake of Acetylated Low-Density Lipoproteins and Lectin Binding
2.3.2. ECFCs Angiogenic Activity In Vitro
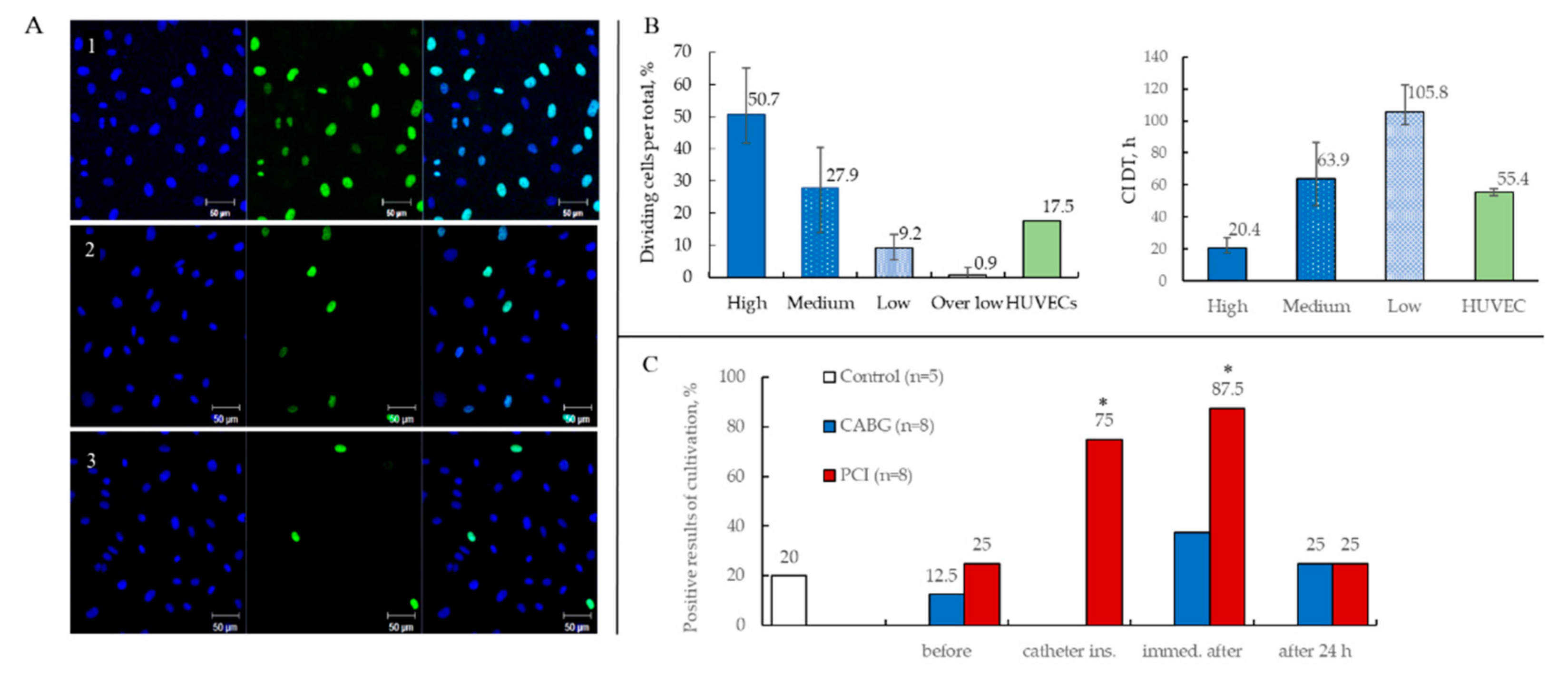
2.3.3. Proliferative Activity of ECFCs
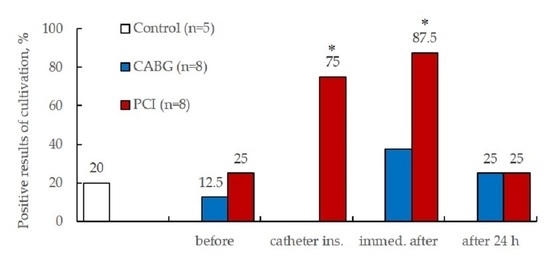
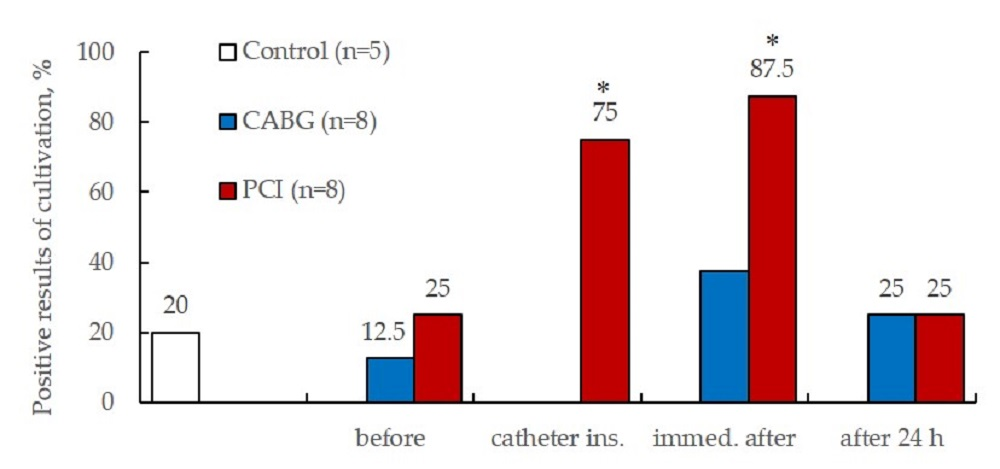
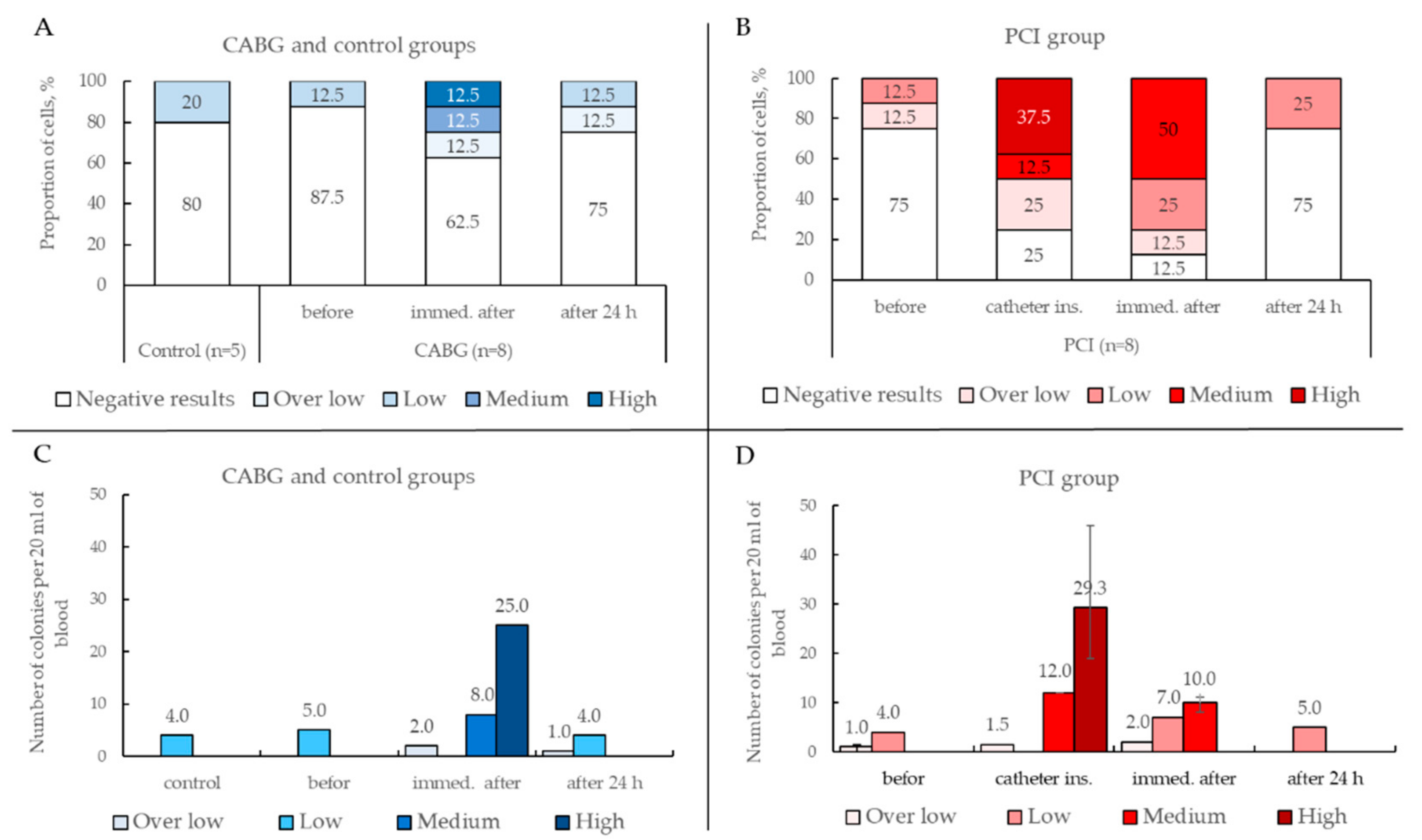
2.3.4. Results of ECFCs Culture in Different Patient Groups
2.4. Detection of Various EPC Phenotypes and ECFC Precursors in PB and PBMCs
3. Discussion
4. Materials and Methods
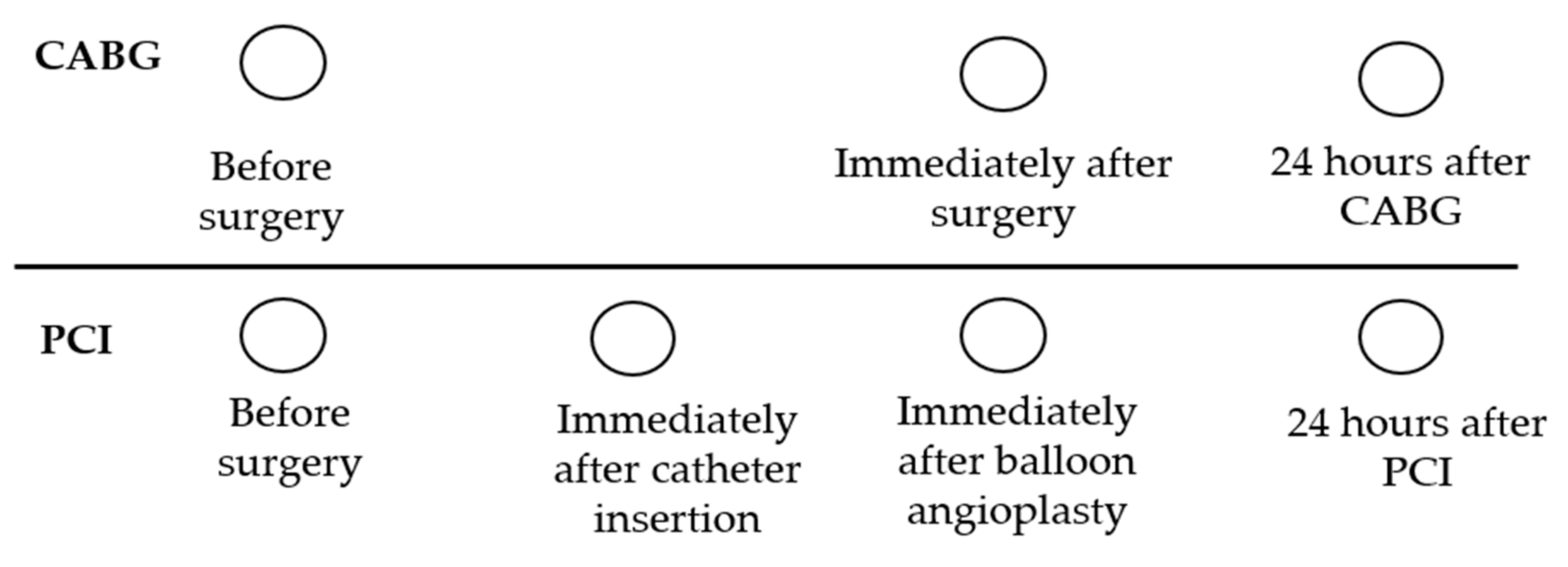
4.1. Study Population and Design
4.2. Cell Phenotyping
4.2.1. Flow Cytometry
- CD3, CD14, HLADR, CD45
- CD34, KDR, CD146, CD133, CD31, CD45
4.2.2. Laser Scanning Microscopy
4.2.3. Proliferative Activity Assessment
4.2.4. Ability of Cells to Absorb Acetylated Low-Density Lipoproteins (Ac-LDL) and Lectin Binding
4.2.5. Angiogenic Activity of Cells In Vitro
4.2.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ECFCs | Endothelial colony-forming cells |
| PB | Peripheral blood |
| PCI | Percutaneous coronary intervention |
| CABG | Coronary artery bypass grafting |
| CAD | Coronary artery disease |
| EPC | Endothelial progenitor cell |
| PBMC | PB mononuclear cell |
| SSC | Side Scatter |
| FSC | Forward Scatter |
| CD | Cluster of differentiation |
| vWF | von Willebrand factor |
| HUVEC | Human umbilical vein endothelial cell |
| HAEC | Human Aortic Endothelial Cells |
| Ac-LDL | Acetylated low density lipoproteins |
| EF | Ejection fraction |
| HD | Hypertonic disease |
| CHF | Сhronic heart failure |
| CPB | Coronary pump bypass |
| PBS | Phosphate buffered saline |
| FC | Flow Cytometry |
| CLSM | Confocal Laser Scanning Microscopy |
| DAPI | 4′,6-diamidino-2-phenylindole |
References
- Ingram, D.A.; Mead, L.E.; Tanaka, H.; Meade, V.; Fenoglio, A.; Mortell, K.; Pollok, K.; Ferkowicz, M.J.; Gilley, D.; Yoder, M.C. Identification of a novel hierarchy of endothelial progenitor cells using human peripheral and umbilical cord blood. Blood 2004, 104, 2752–2760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hur, J.; Yoon, C.H.; Kim, H.S.; Choi, J.H.; Kang, H.J.; Hwang, K.K.; Oh, B.H.; Lee, M.M.; Park, Y.B. Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Barclay, G.R.; Tura, O.; Samuel, K.; Hadoke, P.W.; Mills, N.L.; Newby, D.E.; Turner, M.L. Systematic assessment in an animal model of the angiogenic potential of different human cell sources for therapeutic revascularization. Stem Cell Res. Ther. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, R.J.; O’Neill, C.L.; Sweeney, M.; Guduric-Fuchs, J.; Gardiner, T.A.; Simpson, D.A.; Stitt, A.W. Molecular analysis of endothelial progenitor cell (EPC) subtypes reveals two distinct cell populations with different identities. BMC Med. Genom. 2010, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moccia, F.; Poletto, V. May the remodeling of the Ca2+ toolkit in endothelial progenitor cells derived from cancer patients suggest alternative targets for anti-angiogenic treatment? BBA-Mol. Cell Res. 2015, 1853, 1958–1973. [Google Scholar] [CrossRef]
- Powell, TM; Paul, JD; Hill, JM; Thompson, M.; Benjamin, M.; Rodrigo, M.; McCoy, JP; Read, EJ; Khuu, HM; Leitman, SF; Finkel, T.; Cannon, RO. Granulocyte colony-stimulating factor mobilizes functional endothelial progenitor cells in patients with coronary artery disease. In Arterioscler. Thromb. Vasc. Biol.; 2005; Volume 25, pp. 296–301. [Google Scholar] [CrossRef]
- Tura, O.; Crawford, J.; Barclay, G.R.; Samuel, K.; Hadoke, P.W.F.; Roddie, H.; Davies, J.; Turner, M.L. Granulocyte colony-stimulating factor (G-CSF) depresses angiogenesis in vivo and in vitro: Implications for sourcing cells for vascular regeneration therapy. J. Thromb. Haemost. 2010, 8, 1614–1623. [Google Scholar] [CrossRef] [PubMed]
- Sobrino, T.; Hurtado, O.; Moro, M.A.; Rodríguez-Yáñez, M.; Castellanos, M.; Brea, D.; Moldes, O.; Blanco, M.; Arenillas, J.F.; Leira, R.; et al. The increase of circulating endothelial progenitor cells after acute ischemic stroke is associated with good outcome. Stroke 2007, 38, 2759–2764. [Google Scholar] [CrossRef] [PubMed]
- Adams, V.; Lenk, K.; Linke, A.; Lenz, D.; Erbs, S.; Sandri, M.; Tarnok, A.; Gielen, S.; Emmrich, F.; Schuler, G.; et al. Increase of circulating endothelial progenitor cells in patients with coronary artery disease after exercise-induced ischemia. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Tura, O.; Skinner, E.M.; Barclay, G.R.; Samuel, K.; Gallagher, R.C.; Brittan, M.; Hadoke, P.W.; Newby, D.E.; Turner, M.L.; Mills, N.L. Late outgrowth endothelial cells resemble mature endothelial cells and are not derived from bone marrow. Stem Cells 2013, 31, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Massa, M.; Campanelli, R.; Bonetti, E.; Ferrario, M.; Marinoni, B.; Rosti, V. Rapid and large increase of the frequency of circulating endothelial colony-forming cells (ECFCs) generating late outgrowth endothelial cells in patients with acute myocardial infarction. Exp. Hematol. 2009, 37, 8–9. [Google Scholar] [CrossRef] [PubMed]
- Güven, H.; Shepherd, R.M.; Bach, R.G.; Capoccia, B.J.; Link, D.C. The number of endothelial progenitor cell colonies in the blood is increased in patients with angiographically significant coronary artery disease. J. Am. Coll. Cardiol. 2006, 48, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Hou, D.; Thompson, M.A.; Baysden, S.E.; Shelley, W.C.; Ingram, D.A.; March, K.L.; Yoder, M.C. Acute myocardial infarction in swine rapidly and selectively releases highly proliferative endothelial colony forming cells (ECFCs) into circulation. Cell Transpl. 2007, 16, 887–897. [Google Scholar] [CrossRef]
- Lin, Y.; Weisdorf, D.J.; Solovey, A.; Hebbel, R.P. Origins of circulating endothelial cells and endothelial outgrowth from blood. J. Clin. Investig. 2000, 105, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Walker, L.; Afentoulis, M.; Anderson, D.A.; Jauron-Mills, L.; Corless, C.L.; Fleming, W.H. Transplanted human bone marrow contributes to vascular endothelium. Proc. Natl. Acad. Sci. USA 2004, 101, 16891–16896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, D.A.; Mead, L.E.; Moore, D.B.; Woodard, W.; Fenoglio, A.; Yoder, M.C. Vessel wall-derived endothelial cells rapidly proliferate because they contain a complete hierarchy of endothelial progenitor cells. Blood 2005, 105, 2783–2786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacic, J.C.; Boehm, M. Resident vascular progenitor cells: An emerging role for non-terminally differentiated vessel-resident cells in vascular biology. Stem Cell Res. 2009, 2, 2–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zengin, E.; Chalajour, F.; Gehling, U.M.; Ito, W.D.; Treede, H.; Lauke, H.; Weil, J.; Reichenspurner, H.; Kilic, N.; Ergün, S. Vascular wall resident progenitor cells: A source for postnatal vasculogenesis. Development 2006, 133, 1543–1551. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.G.; Bai, Y.; James, J.R.; Shlapak, DP.; Ju, S. Transplanted endothelial progenitor cells improve ischemia muscle regeneration in mice by diffusion tensor MR imaging. Stem Cells Int. 2016. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Tagawa, S.; Nakanishi, C.; Mori, M.; Yoshimuta, T.; Yoshida, S.; Shimojima, M.; Yokawa, J.; Kawashiri, M.; Yamagishi, M.; Hayashi, K. Determination of early and late endothelial progenitor cells in peripheral circulation and their clinical association with coronary artery disease. Int. J. Vasc. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, M.; Dohle, E.; Katerla, D.; Kirkpatrick, C.J.; Fuchs, S. Enrichment of Outgrowth Endothelial Cells in High and Low Colony-Forming Cultures from Peripheral Blood Progenitors. Tissue Eng. Part C Methods 2010, 16, 877–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, M.S.K.; Ng, W.K.; Chan, J.K.Y. Concise Review: Endothelial Progenitor Cells in Regenerative Medicine: Applications and Challenges. Stem Cells Transl. Med. 2016, 5, 530–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glynn, J.J.; Hinds, M.T. Endothelial outgrowth cells: Function and performance in vascular grafts. Tissue Eng. Part B Rev. 2014, 20, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.L.; Mund, J.A.; Ingram, D.A.; Case, J. Identification of endothelial cells and progenitor cell subsets in human peripheral blood. Curr. Protoc. Cytom. 2010, 9, 9–33. [Google Scholar] [CrossRef] [PubMed]
- Prokopi, M.; Pula, G.; Mayr, U.; Devue, C.; Gallagher, J.; Xiao, Q.; Boulanger, C.M.; Westwood, N.; Urbich, C.; Willeit, J.; et al. Proteomic analysis reveals presence of platelet microparticles in endothelial progenitor cell cultures. Blood 2009, 114, 723–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fina, L.; Molgaard, H.V.; Robertson, D.; Bradley, N.J.; Monaghan, P.; Delia, D.; Sutherland, D.R.; Baker, M.A.; Greaves, M.F. Expression of the CD34 gene in vascular endothelial cells. Blood 1990, 75, 2417–2426. [Google Scholar] [PubMed]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise review: Evidence for CD34 as a common marker for diverse progenitors. Stem Cells 2014, 32, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Siemerink, M.J.; Klaassen, I.; Vogels, I.M.; Griffioen, A.W.; Van Noorden, C.J.; Schlingemann, R.O. CD34 marks angiogenic tip cells in human vascular endothelial cell cultures. Angiogenesis 2012, 15, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreras, C.; Cole, C.L.; Urban, K.; Jayson, G.C.; Avizienyte, E. Segregation of late outgrowth endothelial cells into functional endothelial CD34− and progenitor-like CD34+ cell populations. Angiogenesis 2015, 18, 47–68. [Google Scholar] [CrossRef] [PubMed]
- Siemerink, M.J.; Augustinm, A.J.; Schlingemann, R.O. Mechanisms of ocular angiogenesis and its molecular mediators. Dev. Ophthalmol. 2010, 46, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, H.; Golding, M.; Fruttiger, M.; Ruhrberg, C.; Lundkvist, A.; Abramsson, A.; Jeltsch, M.; Mitchell, C.; Alitalo, K.; Shima, D.; et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J. Cell Biol. 2003, 161, 1163–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellstrom, M.; Phng, L.K.; Hofmann, J.J.; Wallgard, E.; Coultas, L.; Lindblom, P.; Alva, J.; Nilsson, A.K.; Karlsson, L.; Gaiano, N.; et al. Dll4 signalling through Notch1 regulates formation of tip cells during angiogenesis. Nature 2007, 445, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Van Beijnum, J.R.; van der Linden, E.; Griffioen, A.W. Angiogenic profiling and comparison of immortalized endothelial cells for functional genomics. Exp. Cell Res. 2008, 314, 264–272. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


| Results | CD34+ | CD34+CD45− | CD34+CD45−CD133+ | CD34+CD45−CD146+ CD34+CD45−CD146+CD133− CD34+CD45−CD309+ | ||||
|---|---|---|---|---|---|---|---|---|
| PBMCs | Whole PB | PBMCs | Whole PB | PBMCs | Whole PB | PBMCs | Whole PB | |
| Negative (n = 35) | 0.05 0.03–0.08 | 0.04 0.03–0.05 | 85.9 73.3–93.8 | 74.2 65.3–86.5 | 59.1 55.7–62.5 | 50.0 44.4–70.0 | 0 0–0 | 0 0–0 |
| positive (n = 21) | 0.05 0.04–0.08 | 0.045 0.04–0.06 | 86.4 76.0–94.6 | 66.9 63.4–69.6 | 60.75 59.3–62.3 | 54.4 49.3–61.2 | 0 0–0 | 0 0–0 |
| CABG | PCI | p-Value | |
|---|---|---|---|
| Number of patients | 8 | 8 | |
| Age, years | 55.8 (50–64) | 61.0 (51–70) | |
| CAD | CAD | CAD | |
| PolyVD | 25% | 25% | |
| Angina | 75% | 75% | |
| Cardiosclerosis | 12.5% | 62.5 | 0.039 |
| EF > 50% | 100% | 100% | |
| HD III | 87.5% | 100% | |
| CHF I | 87.5% | 75% | |
| CHF II | 12.5% | 25% | |
| Type 2 diabetes | 0 | 25% | |
| Dyslipidemia | 25% | 0 | |
| Cerebral atherosclerosis | 50% | 37.5% | |
| CPB | 62.5% | - | |
| CPB time, min = | 81.4 (67–90) | - | |
| Aortic cross-clamp time, min | 55.4 (43–70) | - | |
| Duration of procedure | - | 38 min (20–80) | |
| balloon | - | 100% | |
| 1 stent/2/3 | - | 75/12.5/12.5 | |
| complications | 12.5% (ACVA) | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matveeva, V.; Khanova, M.; Sardin, E.; Antonova, L.; Barbarash, O. Endovascular Interventions Permit Isolation of Endothelial Colony-Forming Cells from Peripheral Blood. Int. J. Mol. Sci. 2018, 19, 3453. https://doi.org/10.3390/ijms19113453
Matveeva V, Khanova M, Sardin E, Antonova L, Barbarash O. Endovascular Interventions Permit Isolation of Endothelial Colony-Forming Cells from Peripheral Blood. International Journal of Molecular Sciences. 2018; 19(11):3453. https://doi.org/10.3390/ijms19113453
Chicago/Turabian StyleMatveeva, Vera, Mariam Khanova, Egor Sardin, Larisa Antonova, and Olga Barbarash. 2018. "Endovascular Interventions Permit Isolation of Endothelial Colony-Forming Cells from Peripheral Blood" International Journal of Molecular Sciences 19, no. 11: 3453. https://doi.org/10.3390/ijms19113453