Sub-Chronic Stress Exacerbates the Pro-Thrombotic Phenotype in BDNFVal/Met Mice: Gene-Environment Interaction in the Modulation of Arterial Thrombosis

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
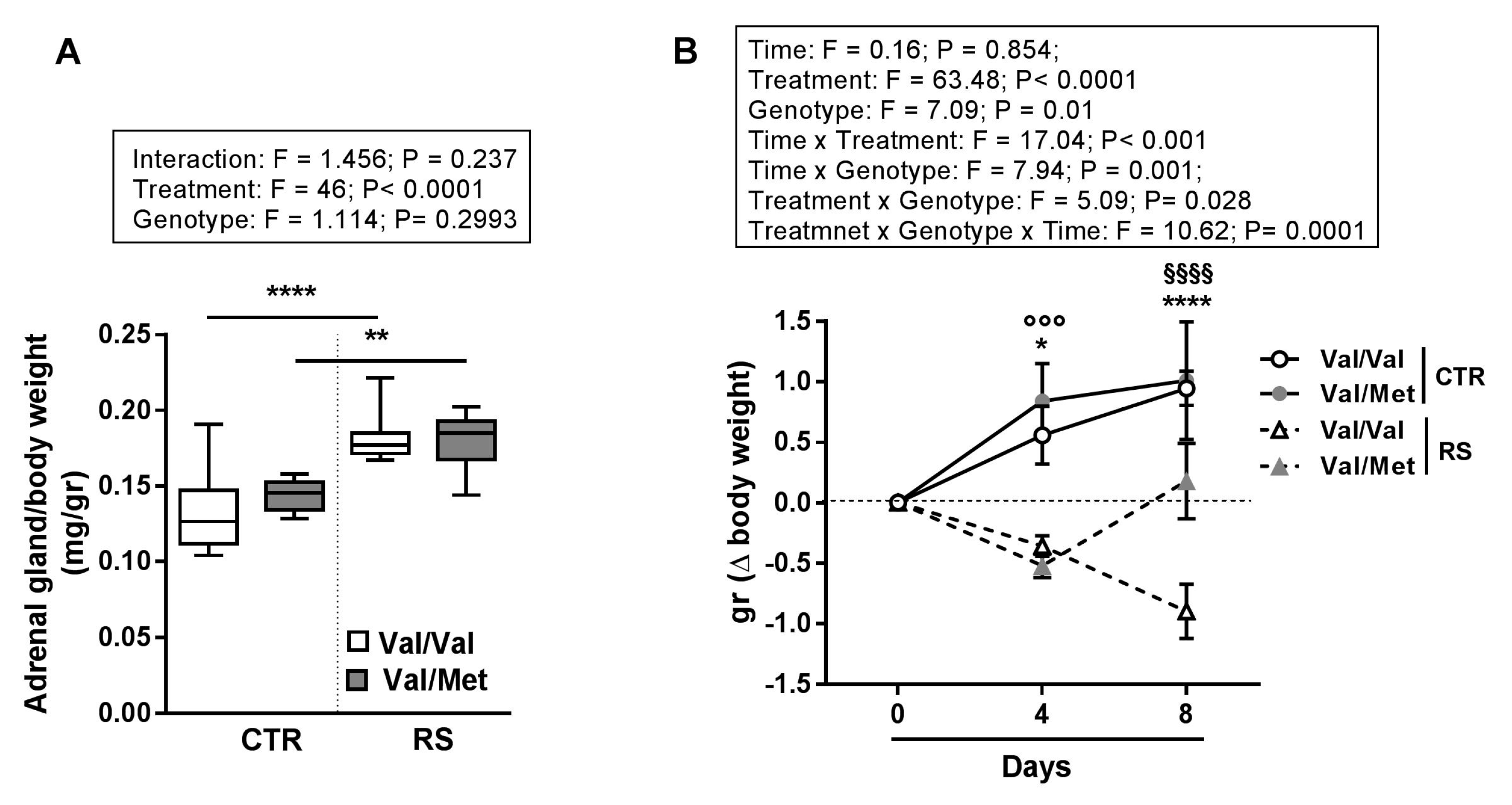
2.1. Sub-Chronic Restraint Stress (RS) Induced Activation of the Hypothalamic-Pituitary-Adrenal Axis and Altered Body Weight
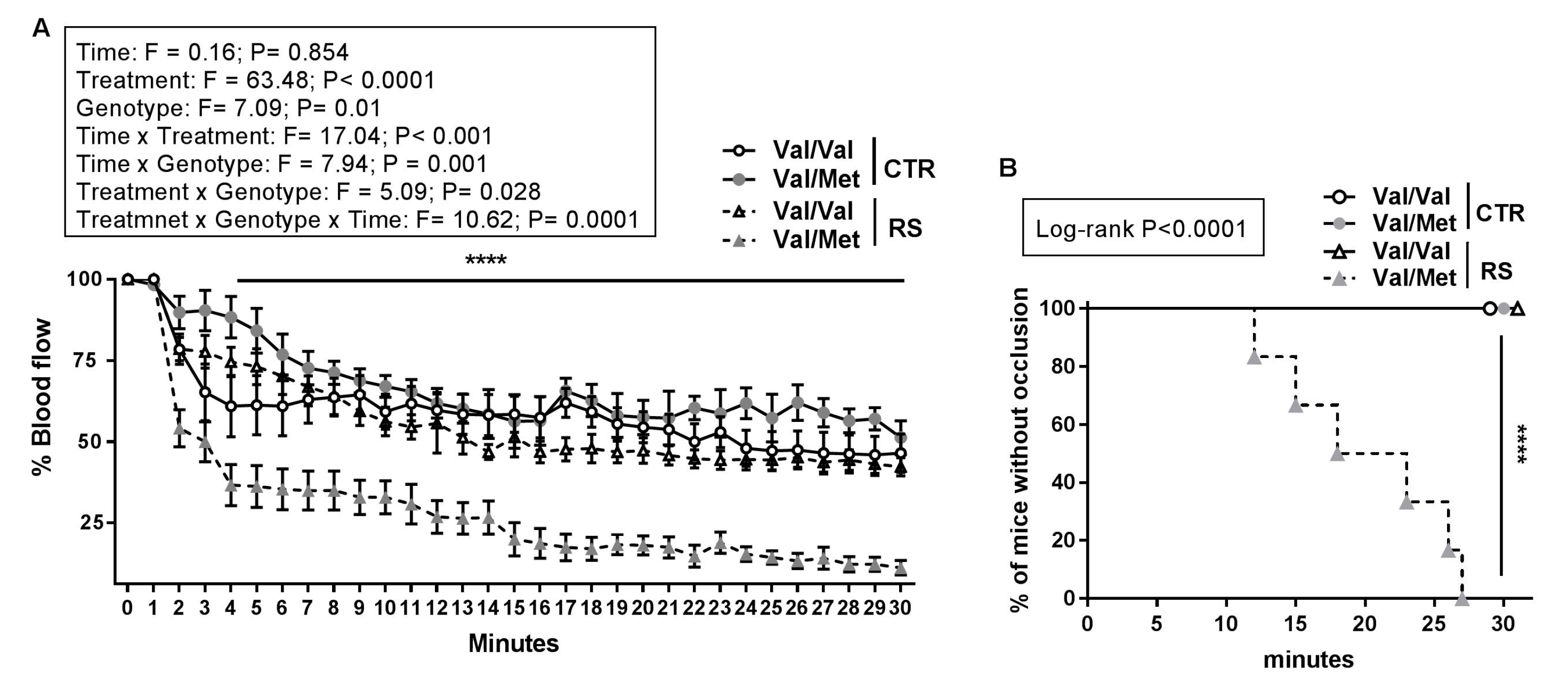
2.2. RS Predisposed BDNFVal/Met Mice to Thrombosis
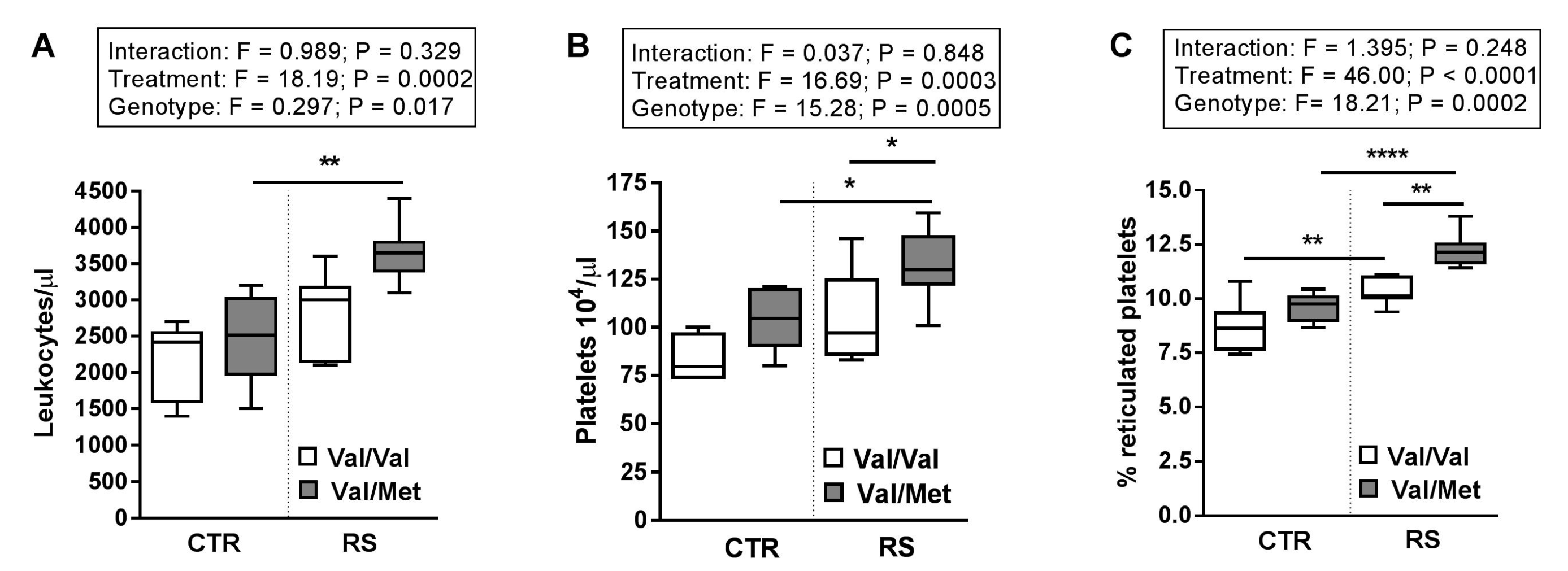
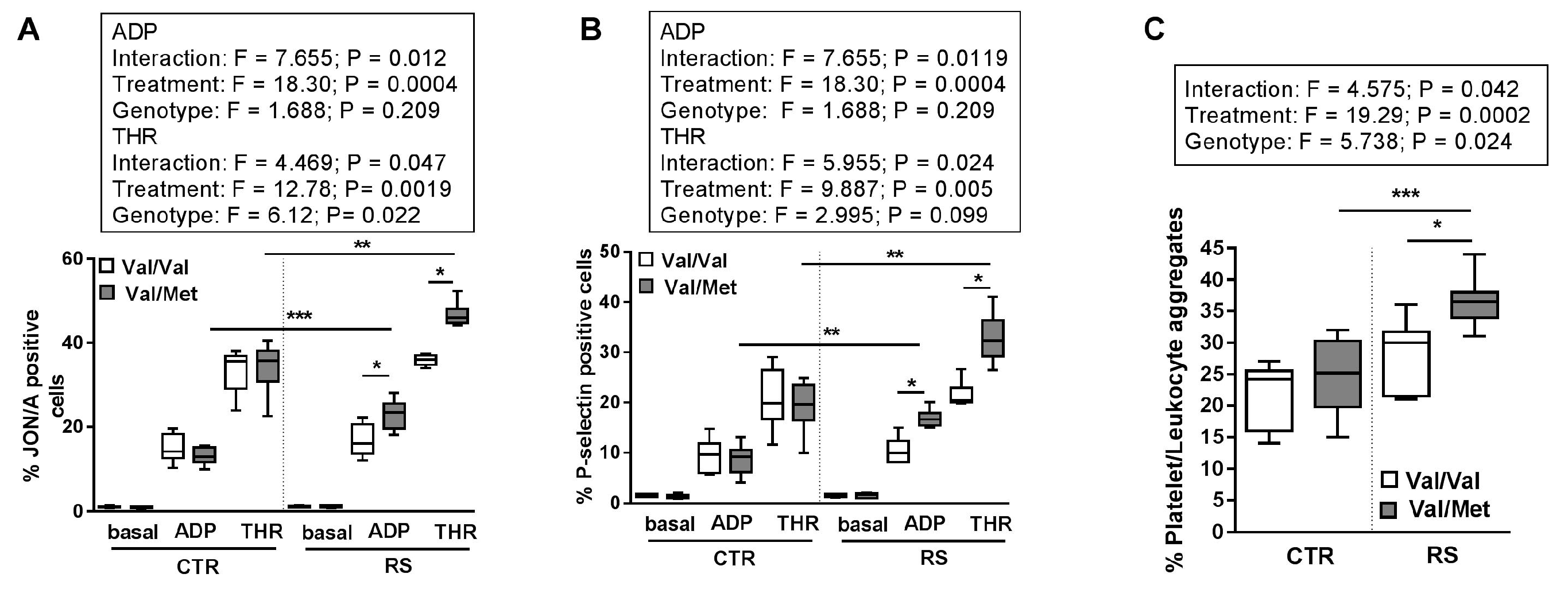
2.3. RS Increased Circulating Blood Cell Number and Platelet Activation State
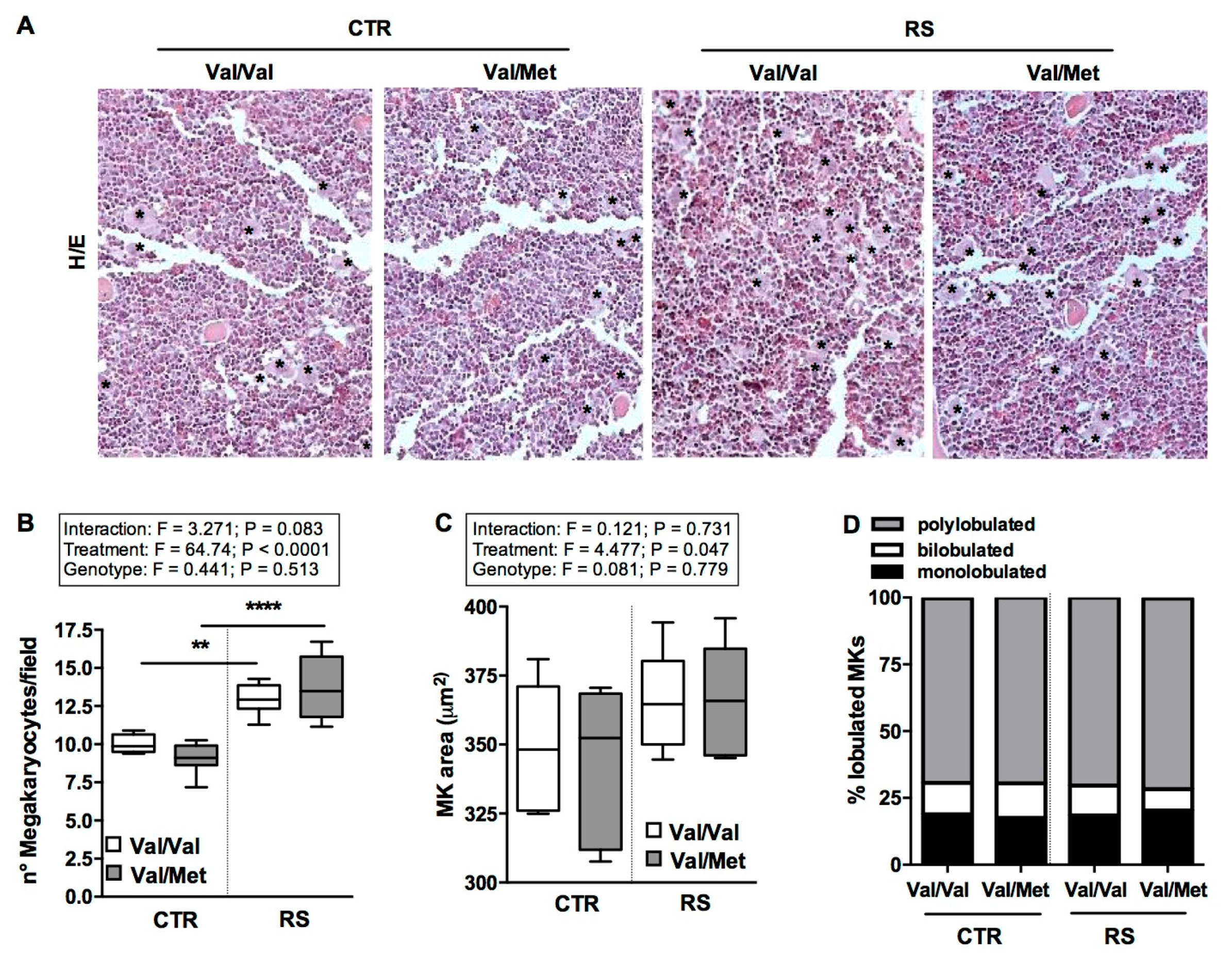
2.4. RS Increased Bone-Marrow Megakaryocytes Number without Affecting Their Maturation State
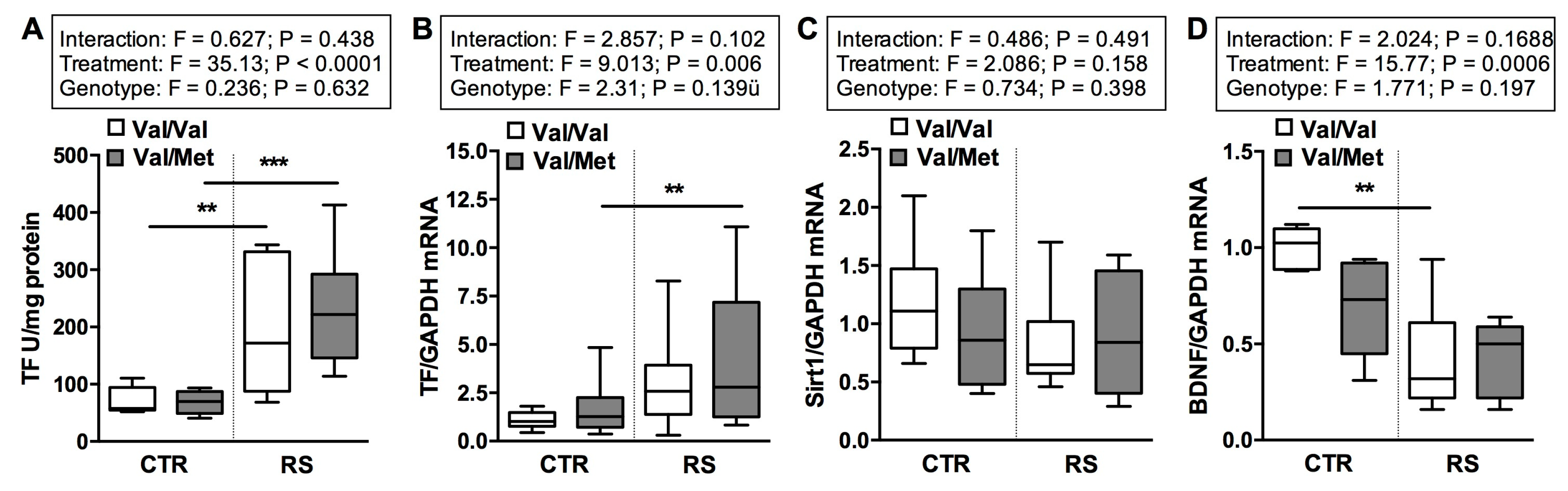
2.5. RS Altered the Expression of Tissue Factor and BDNF but Not the Expression of Sirt1
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Restraint Stress (RS) Procedure
4.3. Arterial Thrombosis Model
4.4. Whole Blood Counts
4.5. Platelet–Leukocyte Aggregate Analysis
4.6. Platelet Studies
4.7. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.8. Measurement of TF Activity in Aortic Tissue
4.9. Bone Marrow Histology and MK Analyses
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BDNF | Brain-Derived Neurotrophic Factor |
| CNS | Central Nervous System |
| CVD | Cardiovascular Disease |
| GxE | Gene-environment Interaction |
| HPA | Hypothalamic–Pituitary–Adrenal Axis |
| RS | Restraint Stress |
References
- Cohen, B.E.; Edmondson, D.; Kronish, I.M. State of the Art Review: Depression, Stress, Anxiety, and Cardiovascular Disease. Am. J. Hypertens. 2015, 28, 1295–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S. Physiology and Neurobiology of Stress and Adaptation: Central Role of the Brain. Physiol. Rev. 2007, 87, 873–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duman, R.S.; Monteggia, L.M. A neurotrophic model for stress-related mood disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Du, C.; Shen, M.; Zhao, G.; Xu, Y.; Yang, K.; Wang, X.; Li, F.; Zeng, D.; Chen, F.; et al. Sympathetic stimulation facilitates thrombopoiesis by promoting megakaryocyte adhesion, migration, and proplatelet formation. Blood 2016, 127, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, L.; Ieraci, A.; Amadio, P.; Popoli, M.; Tremoli, E.; Barbieri, S.S. Apocynin Prevents Abnormal Megakaryopoiesis and Platelet Activation Induced by Chronic Stress. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Thrall, G.; Lane, D.; Carroll, D.; Lip, G.Y. A systematic review of the effects of acute psychological stress and physical activity on haemorheology, coagulation, fibrinolysis and platelet reactivity: Implications for the pathogenesis of acute coronary syndromes. Thromb. Res. 2007, 120, 819–847. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, A.; Hawken, S.; Ounpuu, S.; Sliwa, K.; Zubaid, M.; Almahmeed, W.A.; Blackett, K.N.; Sitthi-amorn, C.; Sato, H.; Yusuf, S.; et al. Association of psychosocial risk factors with risk of acute myocardial infarction in 11119 cases and 13648 controls from 52 countries (the INTERHEART study): Case-control study. Lancet 2004, 364, 953–962. [Google Scholar] [CrossRef]
- Stämpfli, S.F.; Camici, G.G.; Keller, S.; Rozenberg, I.; Arras, M.; Schuler, B.; Gassmann, M.; Garcia, I.; Lüscher, T.F.; Tanner, F.C. Restraint stress enhances arterial thrombosis in vivo—Role of the sympathetic nervous system. Stress 2014, 17, 126–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebner, K.; Singewald, N. Individual differences in stress susceptibility and stress inhibitory mechanisms. Curr. Opin. Behav. Sci. 2017, 14, 54–64. [Google Scholar] [CrossRef]
- Mallei, A.; Ieraci, A.; Popoli, M. Chronic Social Defeat Stress Differentially Regulates the Expression of BDNF transcripts and Epigenetic Modifying Enzymes in Susceptible and Resilient Mice. World J. Biol. Psychiatry 2018, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Belsky, J.; Jonassaint, C.; Pluess, M.; Stanton, M.; Brummett, B.; Williams, R. Vulnerability genes or plasticity genes? Mol. Psychiatry 2009, 14, 746–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondy, B. Common genetic factors for depression and cardiovascular disease. Dialogues Clin. Neurosci. 2007, 9, 19–28. [Google Scholar] [PubMed]
- Cohen-Cory, S.; Kidane, A.H.; Shirkey, N.J.; Marshak, S. Brain-derived neurotrophic factor and the development of structural neuronal connectivity. Dev. Neurobiol. 2010, 70, 271–288. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Nagappan, G.; Lu, Y. BDNF and synaptic plasticity, cognitive function, and dysfunction. Handb. Exp. Pharmacol. 2014, 220, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. Brain-derived neurotrophic factor as a biomarker for mood disorders: An historical overview and future directions. Psychiatry Clin. Neurosci. 2010, 64, 341–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naert, G.; Zussy, C.; Tran Van Ba, C.; Chevallier, N.; Tang, Y.P.; Maurice, T.; Givalois, L. Involvement of Endogenous Brain-Derived Neurotrophic Factor in Hypothalamic-Pituitary-Adrenal Axis Activity. J. Neuroendocrinol. 2015, 27, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Bath, K.G.; Schilit, A.; Lee, F.S. Stress effects on BDNF expression: Effects of age, sex, and form of stress. Neuroscience 2013, 239, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Ieraci, A.; Mallei, A.; Musazzi, L.; Popoli, M. Physical exercise and acute restraint stress differentially modulate hippocampal brain-derived neurotrophic factor transcripts and epigenetic mechanisms in mice. Hippocampus 2015, 25, 1380–1392. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Jing, D.; Bath, K.G.; Ieraci, A.; Khan, T.; Siao, C.-J.; Herrera, D.G.; Toth, M.; Yang, C.; McEwen, B.S.; et al. Genetic Variant BDNF (Val66Met) Polymorphism Alters Anxiety-Related Behavior. Science 2006, 314, 140–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hempstead, B.L. Brain-Derived Neurotrophic Factor: Three Ligands, Many Actions. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 9–19. [Google Scholar] [PubMed]
- Ieraci, A.; Madaio, A.I.; Mallei, A.; Lee, F.S.; Popoli, M. Brain-Derived Neurotrophic Factor Val66Met Human Polymorphism Impairs the Beneficial Exercise-Induced Neurobiological Changes in Mice. Neuropsychopharmacology 2016, 41, 3070–3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallei, A.; Baj, G.; Ieraci, A.; Corna, S.; Musazzi, L.; Lee, F.S.; Tongiorgi, E.; Popoli, M. Expression and Dendritic Trafficking of BDNF-6 Splice Variant are Impaired in Knock-In Mice Carrying Human BDNF Val66Met Polymorphism. Int. J. Neuropsychopharmacol. 2015, 18, pyv069. [Google Scholar] [CrossRef] [PubMed]
- Bukh, J.D.; Bock, C.; Vinberg, M.; Werge, T.; Gether, U.; Vedel Kessing, L. Interaction between genetic polymorphisms and stressful life events in first episode depression. J. Affect Disord. 2009, 119, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Egan, M.F.; Kojima, M.; Callicott, J.H.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef]
- Bozzini, S.; Gambelli, P.; Boiocchi, C.; Schirinzi, S.; Falcone, R.; Buzzi, P.; Storti, C.; Falcone, C. Coronary artery disease and depression: Possible role of brain-derived neurotrophic factor and serotonin transporter gene polymorphisms. Int. J. Mol. Med. 2009, 24, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Su, G.B.; Duan, C.H.; Wang, J.H.; Liu, H.M.; Feng, N.; Wang, Q.X.; Liu, X.E.; Zhang, J. Brain-derived neurotrophic factor gene polymorphisms are associated with coronary artery disease-related depression and antidepressant response. Mol. Med. Rep. 2014, 10, 3247–3253. [Google Scholar] [CrossRef] [PubMed]
- Amadio, P.; Colombo, G.I.; Tarantino, E.; Gianellini, S.; Ieraci, A.; Brioschi, M.; Banfi, C.; Werba, J.P.; Parolari, A.; Lee, F.S.; et al. BDNFVal66met polymorphism: A potential bridge between depression and thrombosis. Eur. Heart J. 2017, 38, 1426–1435. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, D.D.; Wang, Y.; Liu, T.; Lee, F.S.; Chen, Z.Y. Variant brain-derived neurotrophic factor Val66Met polymorphism alters vulnerability to stress and response to antidepressants. J. Neurosci. 2012, 32, 4092–4101. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, S.S.; Amadio, P.; Gianellini, S.; Tarantino, E.; Zacchi, E.; Veglia, F.; Howe, L.R.; Weksler, B.B.; Mussoni, L.; Tremoli, E. Cyclooxygenase-2-derived prostacyclin regulates arterial thrombus formation by suppressing tissue factor in a sirtuin-1-dependent-manner. Circulation 2012, 126, 1373–1384. [Google Scholar] [CrossRef] [PubMed]
- Breitenstein, A.; Stein, S.; Holy, E.W.; Camici, G.G.; Lohmann, C.; Akhmedov, A.; Spescha, R.; Elliott, P.J.; Westphal, C.H.; Matter, C.M.; et al. Sirt1 inhibition promotes in vivo arterial thrombosis and tissue factor expression in stimulated cells. Cardiovasc. Res. 2011, 89, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Svensson, T.; Kitlinski, M.; Engström, G.; Melander, O. A genetic risk score for CAD, psychological stress, and their interaction as predictors of CAD, fatal MI, non-fatal MI and cardiovascular death. PLoS ONE 2017, 12, e0176029. [Google Scholar] [CrossRef] [PubMed]
- Alexander, N.; Osinsky, R.; Schmitz, A.; Mueller, E.; Kuepper, Y.; Hennig, J. The BDNF Val66Met polymorphism affects HPA-axis reactivity to acute stress. Psychoneuroendocrinology 2010, 35, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Takeshita, K.; Yamamoto, K.; Kikuchi, R.; Nakayama, T.; Nomura, M.; Cheng, X.W.; Egashira, K.; Matsushita, T.; Nakamura, H.; et al. Stress augments insulin resistance and prothrombotic state: Role of visceral adipose-derived monocyte chemoattractant protein-1. Diabetes 2012, 61, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Shimokawa, T.; Yi, H.; Isobe, K.; Kojima, T.; Loskutoff, D.J.; Saito, H. Aging and obesity augment the stress-induced expression of tissue factor gene in the mouse. Blood 2002, 100, 4011–4018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidt, T.; Sager, H.B.; Courties, G.; Dutta, P.; Iwamoto, Y.; Zaltsman, A.; von Zur Muhlen, C.; Bode, C.; Fricchione, G.L.; Denninger, J.; et al. Chronic variable stress activates hematopoietic stem cells. Nat. Med. 2014, 20, 754–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, P.T.; Hjemdahl, P.; Olsson, G.; Egberg, N.; Hornstra, G. Altered platelet function during mental stress and adrenaline infusion in humans: Evidence for an increased aggregability in vivo as measured by filtragometry. Clin. Sci. 1989, 76, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Patterson, S.M.; Krantz, D.S.; Gottdiener, J.S.; Hecht, G.; Vargot, S.; Goldstein, D.S. Prothrombotic effects of environmental stress: Changes in platelet function, hematocrit, and total plasma protein. Psychosom. Med. 1995, 57, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Cheng, Y.-W.; Yang, F.; Sun, P.-W.; Zhu, C.-J.; Zhu, L.; Zhang, G.-X. Chronic Stress Facilitates the Development of Deep Venous Thrombosis. Oxid. Med. Cell. Longev. 2015, 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Frodl, T.; Schüle, C.; Schmitt, G.; Born, C.; Baghai, T.; Zill, P.; Bottlender, R.; Rupprecht, R.; Bondy, B.; Reiser, M.; et al. Association of the brain-derived neurotrophic factor Val66Met polymorphism with reduced hippocampal volumes in major depression. Arch. Gen. Psychiatry 2007, 64, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Montag, C.; Reuter, M.; Newport, B.; Elger, C.; Weber, B. The BDNF Val66Met polymorphism affects amygdala activity in response to emotional stimuli: Evidence from a genetic imaging study. Neuroimage 2008, 42, 1554–1559. [Google Scholar] [CrossRef] [PubMed]
- Geuze, E.; Vermetten, E.; Bremner, J.D. MR-based in vivo hippocampal volumetrics: 2. Findings in neuropsychiatric disorders. Mol. Psychiatry 2005, 10, 160–184. [Google Scholar] [CrossRef] [PubMed]
- Ressler, K.J. Amygdala activity, fear, and anxiety: Modulation by stress. Biol. Psychiatry 2010, 67, 1117–1119. [Google Scholar] [CrossRef] [PubMed]
- Tawakol, A.; Ishai, A.; Takx, R.A.; Figueroa, A.L.; Ali, A.; Kaiser, Y.; Truong, Q.A.; Solomon, C.J.; Calcagno, C.; Mani, V.; et al. Relation between resting amygdalar activity and cardiovascular events: A longitudinal and cohort study. Lancet 2017, 389, 834–845. [Google Scholar] [CrossRef]
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Caporali, A.; Emanueli, C. Cardiovascular actions of neurotrophins. Physiol. Rev. 2009, 89, 279–308. [Google Scholar] [CrossRef] [PubMed]
- Donovan, M.J.; Lin, M.I.; Wiegn, P.; Ringstedt, T.; Kraemer, R.; Hahn, R.; Wang, S.; Ibañez, C.F.; Rafii, S.; Hempstead, B.L. Brain derived neurotrophic factor is an endothelial cell survival factor required for intramyocardial vessel stabilization. Development 2000, 127, 4531–4540. [Google Scholar] [PubMed]
- Kermani, P.; Hempstead, B. Brain-derived neurotrophic factor: A newly described mediator of angiogenesis. Trends Cardiovasc. Med. 2007, 17, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Lee, D.H.; Kang, S.S. Effects of chronic restraint stress on body weight, food intake, and hypothalamic gene expressions in mice. Endocrinol. Metab. 2013, 28, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, B.; Bariohay, B.; Moyse, E.; Jean, A. Brain-derived neurotrophic factor (BDNF) and food intake regulation: A minireview. Auton. Neurosci. 2006, 126–127, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Rios, M. BDNF and the central control of feeding: Accidental bystander or essential player? Trends Neurosci. 2013, 36, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Lyons, W.E.; Mamounas, L.A.; Ricaurte, G.A.; Coppola, V.; Reid, S.W.; Bora, S.H.; Wihler, C.; Koliatsos, V.E.; Tessarollo, L. Brain-derived neurotrophic factor-deficient mice develop aggressiveness and hyperphagia in conjunction with brain serotonergic abnormalities. Proc. Natl. Acad. Sci. USA 1999, 96, 15239–15244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppola, V.; Tessarollo, L. Control of hyperphagia prevents obesity in BDNF heterozygous mice. Neuroreport 2004, 15, 2665–2668. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Yeo, G.S.; Cox, J.J.; Morton, J.; Adlam, A.L.; Keogh, J.M.; Yanovski, J.A.; El Gharbawy, A.; Han, J.C.; Tung, Y.C.; et al. Hyperphagia, severe obesity, impaired cognitive function, and hyperactivity associated with functional loss of one copy of the brain-derived neurotrophic factor (BDNF) gene. Diabetes 2006, 55, 3366–3371. [Google Scholar] [CrossRef] [PubMed]
- Bonacina, F.; Barbieri, S.S.; Cutuli, L.; Amadio, P.; Doni, A.; Sironi, M.; Tartari, S.; Mantovani, A.; Bottazzi, B.; Garlanda, C.; et al. Vascular pentraxin 3 controls arterial thrombosis by targeting collagen and fibrinogen induced platelets aggregation. Biochim. Biophys. Acta 2016, 1862, 1182–1190. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, S.S.; Petrucci, G.; Tarantino, E.; Amadio, P.; Rocca, B.; Pesce, M.; Machlus, K.R.; Ranelletti, F.O.; Gianellini, S.; Weksler, B.; et al. Abnormal megakaryopoiesis and platelet function in cyclooxygenase-2-deficient mice. Thromb. Haemost. 2015, 114, 1218–1229. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, S.S.; Amadio, P.; Gianellini, S.; Zacchi, E.; Weksler, B.B.; Tremoli, E. Tobacco smoke regulates the expression and activity of microsomal prostaglandin E synthase-1: Role of prostacyclin and NADPH-oxidase. FASEB J. 2011, 25, 3731–3740. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandrini, L.; Ieraci, A.; Amadio, P.; Veglia, F.; Popoli, M.; Lee, F.S.; Tremoli, E.; Barbieri, S.S. Sub-Chronic Stress Exacerbates the Pro-Thrombotic Phenotype in BDNFVal/Met Mice: Gene-Environment Interaction in the Modulation of Arterial Thrombosis. Int. J. Mol. Sci. 2018, 19, 3235. https://doi.org/10.3390/ijms19103235
Sandrini L, Ieraci A, Amadio P, Veglia F, Popoli M, Lee FS, Tremoli E, Barbieri SS. Sub-Chronic Stress Exacerbates the Pro-Thrombotic Phenotype in BDNFVal/Met Mice: Gene-Environment Interaction in the Modulation of Arterial Thrombosis. International Journal of Molecular Sciences. 2018; 19(10):3235. https://doi.org/10.3390/ijms19103235
Chicago/Turabian StyleSandrini, Leonardo, Alessandro Ieraci, Patrizia Amadio, Fabrizio Veglia, Maurizio Popoli, Francis S. Lee, Elena Tremoli, and Silvia Stella Barbieri. 2018. "Sub-Chronic Stress Exacerbates the Pro-Thrombotic Phenotype in BDNFVal/Met Mice: Gene-Environment Interaction in the Modulation of Arterial Thrombosis" International Journal of Molecular Sciences 19, no. 10: 3235. https://doi.org/10.3390/ijms19103235