Inhibiting Skp2 E3 Ligase Suppresses Bleomycin-Induced Pulmonary Fibrosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. BLM-Induced Lung Fibrosis Was Suppressed in Skp2-Deficient Mice
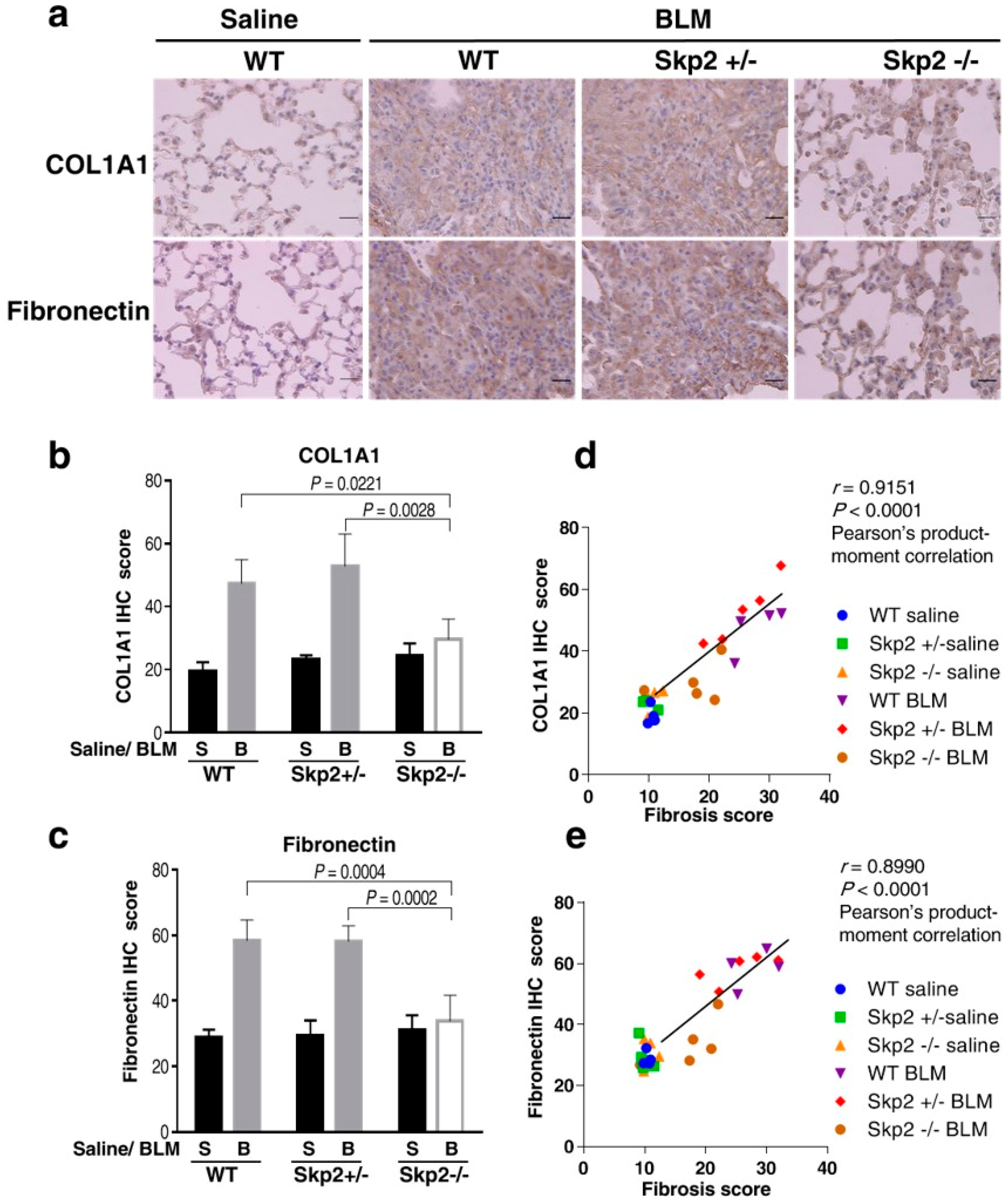
2.2. Skp2-Deficiency Suppressed the Accumulation of Fibrosis Markers in the BLM Model
2.3. Effects of Skp2-Deficiency on Bronchoalveolar Lavage Fluid Cells in the BLM Mouse Model
2.4. The Effects of Skp2-Deficiency on p27 Levels in BLM-Induced Lung Fibrosis
2.5. Skp2-Deficiency May Suppress the BLM-Induced Increase in Mesenchymal Fibroblasts
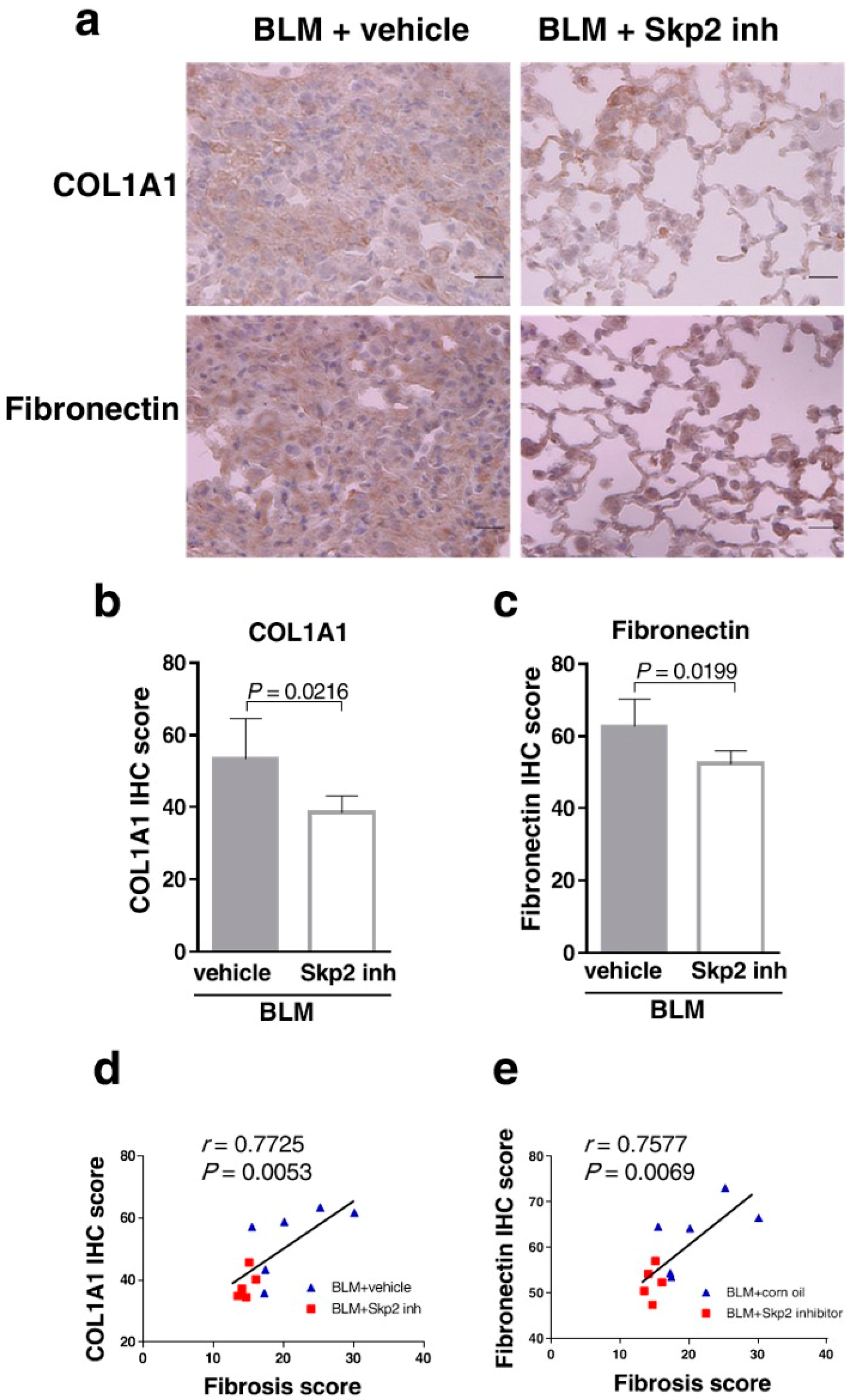
2.6. The Skp2 Inhibitor SZL-P1-41 Suppressed BLM-Induced Lung Fibrosis
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Murine Model of Pulmonary Fibrosis and SZL-P1-41 Treatment
4.3. Bronchoalveolar Lavage (BAL)
4.4. Histochemical Staining
4.5. Immunohistochemistry
4.6. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AECI | type I alveolar epithelial cells |
| AECII | type II alveolar epithelial cells |
| ANOVA | analysis of variance |
| BALF | bronchoalveolar lavage fluid |
| BLM | bleomycin |
| CDK | cyclin dependent kinase |
| COL1A1 | type 1 alpha 1 collagen |
| DAB | 3,3′-Diaminobenzidine |
| ECM | extracellular matrix |
| ER | endoplasmic reticulum |
| FGF | fibroblast growth factor |
| HE | haematoxylin-eosin |
| HRP | horseradish peroxidase |
| MT | Masson trichrome |
| PBS | phosphate buffered saline |
| PDGF | platelet-derived growth factor receptor |
| SCF | Skp1/Cullin/F-box |
| SFTPC | surfactant protein C |
| Skp2 | S-phase kinase-associated protein 2 |
| TUNEL | TdT-mediated dUTP nick end labeling |
| TGF-β | transforming growth factor |
| VEGF | vascular endothelial growth factor |
| WT | wild type |
References
- Wolters, P.J.; Collard, H.R.; Jones, K.D. Pathogenesis of idiopathic pulmonary fibrosis. Annu. Rev. Pathol. 2014, 9, 157–179. [Google Scholar] [CrossRef] [PubMed]
- Ley, B.; Collard, H.R.; King, T.E., Jr. Clinical course and prediction of survival in idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2011, 183, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.-F.; Flaherty, K.R.; Lasky, J.A.; et al. An official ATS/ERS/JRS/ALAT statement: Idiopathic pulmonary fibrosis: Evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med. 2011, 183, 788–824. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, K.B.; Samet, J.M.; Stidley, C.A.; Colby, T.V.; Waldron, J.A. Cigarette smoking: A risk factor for idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 1997, 155, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Taskar, V.S.; Coultas, D.B. Is idiopathic pulmonary fibrosis an environmental disease? Proc. Am. Thorac. Soc. 2006, 3, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, J.; Rubio, G.A.; Limper, A.H.; Williams, K.; Elliot, S.J.; Ninou, I.; Aidinis, V.; Tzouvelekis, A.; Glassberg, M.K. Exploring animal models that resemble idiopathic pulmonary fibrosis. Front. Med. 2017, 4, 118. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Fukasawa, H.; Kitagawa, K.; Uchida, C.; Hattori, T.; Isobe, T.; Oda, T.; Misaki, T.; Ohashi, N.; Nakayama, K.; et al. Renal damage in obstructive nephropathy is decreased in Skp2-deficient mice. Am. J. Pathol. 2007, 171, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Skaar, J.R.; Pagan, J.K.; Pagano, M. Mechanisms and function of substrate recruitment by F-box proteins. Nat. Rev. Mol. Cell Biol. 2013, 14, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, P.; Inuzuka, H.; Wei, W. Roles of F-box proteins in cancer. Nat. Rev. Cancer 2014, 14, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Carrano, A.C.; Eytan, E.; Hershko, A.; Pagano, M. SKP2 is required for ubiquitin-mediated degradation of the CDK inhibitor p27. Nat. Cell Biol. 1999, 1, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, K.; Kitagawa, M. The SCF-type E3 ubiquitin ligases as cancer targets. Curr. Cancer Drug Targets 2016, 16, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Ohashi, N.; Kitagawa, M. Roles of the Skp2/p27 axis in the progression of chronic nephropathy. Cell. Mol. Life Sci. 2013, 70, 3277–3287. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 1988, 41, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.H.; Morrow, J.K.; Li, C.F.; Gao, Y.; Jin, G.; Moten, A.; Stagg, L.J.; Ladbury, J.E.; Cai, Z.; Xu, D.; et al. Pharmacological inactivation of Skp2 SCF ubiquitin ligase restricts cancer stem cell traits and cancer progression. Cell 2013, 154, 556–568. [Google Scholar] [CrossRef] [PubMed]
- King, T.E., Jr.; Pardo, A.; Selman, M. Idiopathic pulmonary fibrosis. Lancet 2011, 378, 1949–1961. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ishitsuka, Y.; Hayasaka, M.; Yamada, Y.; Miyata, K.; Endo, M.; Kondo, Y.; Moriuchi, H.; Irikura, M.; Tanaka, K.; et al. The exacerbating roles of CCAAT/enhancer-binding protein homologous protein (CHOP) in the development of bleomycin-induced pulmonary fibrosis and the preventive effects of tauroursodeoxycholic acid (TUDCA) against pulmonary fibrosis in mice. Pharmacol. Res. 2015, 99, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Qin, H.; Du, W.; Shen, Y.W.; Lee, W.H.; Riggs, A.D.; Liu, C.P. Inhibition of S-phase kinase-associated protein 2 (Skp2) reprograms and converts diabetogenic T cells to Foxp3+ regulatory T cells. Proc. Natl. Acad. Sci. USA 2012, 109, 9493–9498. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.M.S.; Gomes, S.T.M.; Ishak, R.; Vallinoto, A.C.R. Regulatory T Cell and forkhead box protein 3 as modulators of immune homeostasis. Front. Immunol. 2017, 8, 605. [Google Scholar] [CrossRef] [PubMed]
- Izbicki, G.; Segel, M.J.; Christensen, T.G.; Conner, M.W.; Breuer, R. Time course of bleomycin-induced lung fibrosis. Int. J. Exp. Pathol. 2002, 83, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Chung, H.S.; Shin, D.; Jung, K.H.; Lee, H.; Moon, J.; Bae, H. Adenovirus-mediated Foxp3 expression in lung epithelial cells reduces airway inflammation in ovalbumin and cockroach-induced asthma model. Exp. Mol. Med. 2016, 48, e259. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lim, H.S. Skp2 inhibitors: Novel anticancer strategies. Curr. Med. Chem. 2016, 23, 2363–2379. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Nagahama, H.; Minamishima, Y.A.; Matsumoto, M.; Nakamichi, I.; Kitagawa, K.; Shirane, M.; Tsunematsu, R.; Tsukiyama, T.; Ishida, N.; et al. Targeted disruption of Skp2 results in accumulation of cyclin E and p27Kip1, polyploidy and centrosome overduplication. EMBO J. 2000, 19, 2069–2081. [Google Scholar] [CrossRef] [PubMed]
- Goldner, J. A modification of the masson trichrome technique for routine laboratory purposes. Am. J. Pathol. 1938, 14, 237–243. [Google Scholar] [PubMed]
- Pandit, K.V.; Corcoran, D.; Yousef, H.; Yarlagadda, M.; Tzouvelekis, A.; Gibson, K.F.; Konishi, K.; Yousem, S.A.; Singh, M.; Handley, D.; et al. Inhibition and role of let-7d in idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2010, 182, 220–229. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikamo, M.; Kitagawa, K.; Sakai, S.; Uchida, C.; Ohhata, T.; Nishimoto, K.; Niida, H.; Suzuki, S.; Nakayama, K.I.; Inui, N.; et al. Inhibiting Skp2 E3 Ligase Suppresses Bleomycin-Induced Pulmonary Fibrosis. Int. J. Mol. Sci. 2018, 19, 474. https://doi.org/10.3390/ijms19020474
Mikamo M, Kitagawa K, Sakai S, Uchida C, Ohhata T, Nishimoto K, Niida H, Suzuki S, Nakayama KI, Inui N, et al. Inhibiting Skp2 E3 Ligase Suppresses Bleomycin-Induced Pulmonary Fibrosis. International Journal of Molecular Sciences. 2018; 19(2):474. https://doi.org/10.3390/ijms19020474
Chicago/Turabian StyleMikamo, Masashi, Kyoko Kitagawa, Satoshi Sakai, Chiharu Uchida, Tatsuya Ohhata, Koji Nishimoto, Hiroyuki Niida, Sayuri Suzuki, Keiichi I. Nakayama, Naoki Inui, and et al. 2018. "Inhibiting Skp2 E3 Ligase Suppresses Bleomycin-Induced Pulmonary Fibrosis" International Journal of Molecular Sciences 19, no. 2: 474. https://doi.org/10.3390/ijms19020474