Infectious Bronchitis Virus Variants: Molecular Analysis and Pathogenicity Investigation


1
Department of Veterinary Medicine, National Taiwan University, Taipei 10617, Taiwan
2
Research Center for Nanotechnology and Infectious Diseases, Taipei 11529, Taiwan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(10), 2030; https://doi.org/10.3390/ijms18102030
Submission received: 12 August 2017
/
Revised: 16 September 2017
/
Accepted: 18 September 2017
/
Published: 22 September 2017
(This article belongs to the Special Issue Molecular Research of Emerging Viruses: Viral Evolution, Diagnostics and Pathogenesis and Therapeutics)
Abstract
:Infectious bronchitis virus (IBV) variants constantly emerge and pose economic threats to poultry farms worldwide. Numerous studies on the molecular and pathogenic characterization of IBV variants have been performed between 2007 and 2017, which we have reviewed herein. We noted that viral genetic mutations and recombination events commonly gave rise to distinct IBV genotypes, serotypes and pathotypes. In addition to characterizing the S1 genes, full viral genomic sequencing, comprehensive antigenicity, and pathogenicity studies on emerging variants have advanced our understanding of IBV infections, which is valuable for developing countermeasures against IBV field outbreaks. This review of IBV variants provides practical value for understanding their phylogenetic relationships and epidemiology from both regional and worldwide viewpoints.
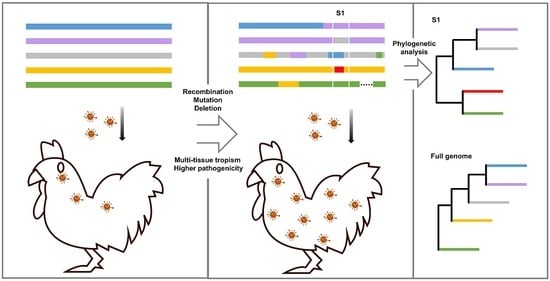
1. Introduction
Infectious bronchitis virus (IBV) is an economically important pathogen in the poultry industry. Infectious bronchitis, resulting from IBV, is an acute, highly contagious infection accompanied by upper respiratory symptoms and urinary and reproductive system failures, resulting in low production rates, egg drop, and death in chickens. Since its first documentation in 1931 in the United States [1], IBV has spread to all continents that host chicken farming. Vaccination is the most common method to prevent IBV infections in chickens, and commercial vaccines include live attenuated vaccines and inactivated vaccines. However, the cross protections of the vaccines are limited, as novel serotypes are continuously emerging, and immune failures are reported frequently. The Massachusetts (Mass) and 4/91 (also known as 793B) types are widely used around the world, while local strains have been selected for vaccine development in individual regions.
IBV is a gammacoronavirus belonging to the Coronaviridae family, and its virion contains a single copy of a positive-sense, single-stranded 27.6-kb RNA. The IBV genome encodes four structural proteins, polyproteins 1a and 1ab, and several accessory proteins. The coding regions of the polyproteins cover two-thirds of the viral genome and are subjected to further processing into 15 non-structural proteins (nsp2–16). Accessory proteins include 3a, 3b, 5a, and 5b, but these may be missing in some IBV strains. The structural proteins are spike (S), envelope (E), membrane (M) and nucleocapsid (N). The S protein, especially the S1 subunit that is formed by post-translational cleavage [2,3], has proven to be critical for antigenic neutralization, hemagglutination and cell tropism determination [4,5,6]. Most previously published IBV molecular characterizations have focused on the analysis of the S gene, which may not be sufficient to explain the observed changes in the serotypes and pathotypes of IBV variants. Several studies have noted that viral replication and immune escape may be modulated by non-structural and accessory viral proteins in IBV and other coronaviruses, such as mouse hepatitis virus and severe acute respiratory syndrome coronavirus (SARS-CoV) [7,8]. Additionally, cytotoxic T lymphocyte epitopes in the N protein can protect chickens from IBV infection [9,10]. These observations emphasize the importance of studying the full-length IBV genome and its associations with serological and pathogenic characteristics. To provide a comprehensive outlook on circulating IBV variants, we have reviewed numerous studies reporting IBV variants between 2007 and 2017 from different continents, and summarized the findings from molecular analyses, epidemiological data, and antigenicity/pathogenicity investigations (Table 1).
2. IBV Classification
Typically, IBV can be classified by nucleic acid-based or antibody-based methods, which provides genotyping and serotyping results, respectively. The cross neutralization test, a conventional method for distinguishing viral serotypes, analyzes the neutralizing activity of antisera prepared in specific-pathogen-free (SPF) chickens. Archetti and Horsfall Jr [11] demonstrated the calculation of the relatedness value (R-value) obtained from neutralization tests. The hemagglutination inhibition test is also used in serotyping IBV, but disadvantageously, the virus must be treated with neuraminidase to yield hemagglutination activity [12]. The enzyme-linked immunosorbent assay (ELISA), in contrast, cannot discriminate between different serotypes, as the cross-reactive antibody interferes with the detecting signal, particularly when the coating antigen is derived from whole virions. To this end, several serotyping ELISAs incorporating the specificity of monoclonal antibodies have been reported [13,14]. Collectively, performing serotyping procedures with embryonic eggs or chickens is not cost-effective; therefore, genotyping, which is relatively simple to perform, has become increasingly popular with the development of reverse transcriptase polymerase chain reaction (RT-PCR) techniques, gene sequencing technology, and bioinformatics. Most laboratories use these tools in a combined method to amplify the S1 gene of IBV variants, and then analyze the sequences using Basic Local Alignment Search Tool (BLAST) or phylogenetic analyses. In addition, using genomic sequence comparison, recombination events can be detected among two or more nucleotide sequences, and the breakpoints can be identified by the Recombination Analysis Tool (RAT) or SimPlot [15]. While the S1 sequence is reportedly strongly correlated with the protective R value [16], changes in serotype can result from only a few changes in the amino acids of the S1 protein [17]. However, there was no unifying nomenclature on IBV genotypes, leading to distinctive and confusing names based primarily on the viruses’ geographic origin. To provide a more unified IBV genetic classification, Valastro et al. [18] put forward a method that defines IBV strains into six genotypes comprising 32 distinct viral lineages based on the complete S1 gene. In the Table 1, the genotype and lineage of each strain according to this classification method were indicated together with the traditional sorting name. IBV pathotypes can vary greatly among strains, and clinical symptoms caused by infection can be diverse due to various chicken breeds, environmental circumstances and immunity levels. Although IBV demonstrates clear tropism by producing lesions in the respiratory system, kidney, reproductive system, and alimentary tract, the viral pathotypes observed in field outbreaks are usually more complicated due to co-infections with other pathogens [19]. Thus, pathogenicity results generated from experimental infection using SPF chickens with good laboratory practice are more reliable. The protectotype, a direct indicator of the protective efficacy against a virus, is a practical typing method for vaccine evaluation. However, the large number of experimental animals and labor required to determine the pathotypes or protectotypes of IBV are costly.
3. Evolution Mechanism of IBV
According to modern evolutionary hypotheses [47], genetic diversity and selection processes are the two basic steps of evolution. Spontaneous mistakes (nucleotide substitutions, deletions or insertions, and recombination) made during viral replication provide the materials for selection, eventually resulting in virus evolution. The mutation rate of RNA virus is 10−4 to 10−5 substitutions per nucleotide (nt) per round of replication. Even though a 3′ to 5′ exoribonuclease (ExoN) in coronavirus nsp14 was proven to have RNA proofreading function, the mutation rate remains significantly higher than the likes of DNA genome organisms, such as E. coli, which has a mutation rate of only 10−9 to 10−11 substitutions per nt per round of replication [48,49,50]. However, the proofreading function of nsp14 may also play an important role in coronavirus replication, to reach a balance between fidelity and diversity, facilitating adaptation to environmental selection pressure. Details regarding IBV selection regulation were fully elucidated by Toro et al. [51], and selection forces may be immune responses induced by several types of vaccines, the microenvironment of infected hosts, or physical and biosafety conditions. In chickens, immune responses can be ceased by infection with an immunosuppressive virus, such as chicken anemia virus, infectious bursal disease virus, and Marek’s disease virus. Additionally, viral immunity levels and specific antibodies may differ across farms or countries based on their imposed immunization schedules. Host microenvironments can have slight differences, even though in individual hosts, distinct tissues have distinct cell receptors, temperatures, pH values, enzymes, and concentrations that can alter the predominant virus genotype by selective pressure [52,53]. Physical conditions aiding virus spread include environmental temperature, humidity, and wind. The distance of a chicken farm from an infectious source and the all-in-all-out system play important roles in biosafety conditions. While numerous IBV variants have emerged because of errors during genomic replication, only a few variants became endemic in specific regions under these described selection factors, while the others inflicted no long-term effects. This theory can also explain why some strains are first observed in one county, disappear, and then reappear as an endemic in other countries several years later. Zhao et al. [54], confirmed that every IBV-encoding gene undergoes positive selection, and analyzed the evolution rate of IBV in China, finding that the E gene evolves at the fastest rate among all the structural protein coding genes, with a substitution rate of 10−6 nt substitutions per site per year. The N gene evolves the slowest, with 10−5 nt substitutions per site per year. These selective pressures have forced IBV to rapidly evolve in the past several decades, and variants having distinct genotypes, serotypes, and pathogenic types have been continually reported.
4. Asia
IBV in Taiwan was first documented in 1958, and it can be divided into two groups, Taiwan group I (TW-I) and Taiwan group II (TW-II). Both groups are distinct from other strains around the world in terms of their genotypes and serotypes [55,56]. Except for the nucleocapsid gene, the structural and non-structural genes of Taiwan isolates are more closely related to China strains than US strains [57]. While most Chinese neighbors have been affected by Chinese QX-like IBV strains, this has not occurred in Taiwan. However, the Chinese CK/CH/LDL/97I (Q1-like) recombinant IBV strain was reported in Taiwan by Chen et al. [58]. The surveillance program conducted in poultry slaughterhouses showed IBV prevalence rates at 17% during 2005–2006 and 39% in 2013, respectively, from which several variants derived from recombination among Taiwan, Chinese Q1, Mass and Japan types were detected [59,60]. The Q1-like and Japan-like IBV strains revealed high lethality and possessed multi-organ tropism in experimentally infected chickens (unpublished data). In addition to recombination events, point mutations in a TW-I strain were found in the IBV 3575/08, resulting in increased pathogenicity and serotype and immunotype changes, suggesting that IBV in Taiwan is continuously evolving [34].
Because China is a large country and large populations of chickens are maintained at high densities in large-scale factories or backyards, IBV strains have numerous opportunities to spread and recombine with each other. In China, IBV was first observed in the early 1980s, and IBV outbreaks have since been frequently reported. According to the phylogenetic analysis of new isolates based on the S1 gene, the predominant strains in China are the QX-like strains [61]. The IBV QX strain was first isolated in China in 1996 and was characterized by proventriculus swelling in affected chickens [62]. Until 2009, the QX-like strain was found to be the second most prevalent strain in China [63]. In fact, the proportion of QX-like isolates has increased from 20% in the 2000s to over 60% since 2007 [54]. Han et al. [64] reported that 54.1% (119/220) of IBV isolates in China were of the LX4 type, which is a well-known QX-like type IBV. The LDT3 and 4/91 (also known as 793B) types have also frequently been isolated in China recently [65,66,67]. The N-terminal sequence of the S1 protein of the 4/91 type is reportedly more variable than those of other types, such as Mass and QX [68].
Commercial vaccines used against IBV in China include the attenuated live strains H120, LDT3, 4/91 and some other inactivated vaccines, such as M41. Mass type is the most widely inoculated vaccine and also circulates in China, as its isolation rates range from 6.94% to 8.64%, according to previous studies [23,64]. Because of recombination with other strains, some Mass type isolates can be cross-neutralized by H120 antiserum and are completely protected against by the H120 vaccination. However, two isolated Mass type recombinant strains, CK/CH/LDL/110931 and CK/CH/LHB/130573, are serologically different from the H120 vaccine and thus cannot be protected against [23]. Several amino acid point mutations in the S1 subunit may be responsible for this immune escape [64].
With plentiful local and vaccine strains simultaneously circulating in China, recombination has become a common phenomenon, and recombination events within vaccine strains [23,69,70] and other local strains [70,71] are widely found. For example, the CK/CH/LGX/130530 strain has a breakpoint in nsp14, and its 5′-terminal portion is from H120, while its 3′-terminal portion was donated by the tl/CH/LDT3/03-like virus. The lower virulence of CK/CH/LGX/130530 also provided evidence that non-structural proteins play an important role in virulence determination [29]. The Sczy3 strain is a recombinant strain that was derived from the major parental LX4 strain and the minor parental H120 strain [32]. The nephropathogenic strain SAIBK, which also recombined from the major parent SC021202 and H120, was determined to be the 4/91 serotype [33]. The CK/CH/LSD/100408 strain has sequences from both LX4 and tl/CH/LDT3/03 type strains in different parts of its S1 gene [61]. A newly emerged genotype was classified as lineage GI-28 since it was found to be genetically different from other IBV strains. The representative strains are CK/CH/LGX/111119 and CK/CH/2010/JT-1, which are recombinant strains originated from LX4 or partridge/GD/S14/2003 [20,21].
Taiwan groups (TW-I and TW-II) of IBV strains have been isolated in China and the emergence of TW types strains has increased since 2009 [26,65,72,73]. Xu et al. [26] reported a highly virulent recombinant created from S1 genes from the QX and TW-I strains, named the GD strain, that can induce severe respiratory symptoms, renal lesions, and mortality in approximately 40% of cases. The GD strain was classified as the TW genotype, but it can be completely protected by the QX-like IBV strain JS [27]. Another TW-I and LX4 strain recombinant (CK/CH/LDL/140520) that causes cystic oviducts as well as nephritis and respiratory distress in one-day-old chickens was identified by Gao et al. [28]. Moreover, the TW-II-like strain CK/CH/LHB/100801 was found to have emerged in chicken flocks in China in 2011, but this strain exhibits mutations and deletions from the strain isolated in Taiwan [69].
In Japan, IBV can be divided into three genetic groups, JP-I, JP-II and JP-III. In addition to these groups, some 4/91 type strains were isolated and confirmed to be variants [74,75]. JP/Wakayama/2003 is a 4/91 type strain that can be cross-neutralized by serum from 4/91, JP-I (GN strain) and H120, with R-values higher than 79. Another 4/91 type strain, JP/Iwate/2005, is completely protected by the 4/91 vaccine and the JP-II strain (TM-86w), even though its R-value with TM-86w was low [35]. Notably, all these 4/91 type isolates are more virulent than the vaccine strain. A novel genotype strain, JP/Ibaraki/168-1/2009, was distinctly isolated with other Japanese strains but was similar to the TC07-2 strain that was isolated in south China based on the hypervariable S1 region, and serology tests also confirmed this strain as a novel serotype. This novel strain cannot be neutralized by Japan type strains or the H120, Gray and 4/91 strains [76].
Two groups of IBVs exist in Korea, named Korean group I and Korean group II. Korean group I is more closely related to the Mass type, while Korean group II is a distinctive branch. Among Korean group II viruses, three subgroups were found, QX-like, KM91-like and New Cluster 1 (NC1) [77]. All of the new isolates that emerged from 2005 to 2010 were nephropathogenic and clustered into Korean group II. Among these, K716/05 was found to be a KM91 and QX recombinant strain in the S1 gene [77]. A KM91-like backbone recombinant, SNU8067, was reported to inhibit hierarchal ovarian follicle formation and oviduct maturation. Recombination with the Mass type vaccine strain has also been documented [36], and the NC1 strain is continuously evolving and accumulating point mutations [78].
In Thailand, while the vaccine strains derived from M41, H120, Ma5, Connecticut (Conn), 4/91 and the local DLD strain were widely used, three distinct groups including the unique Thailand type QX-like, and Mass type are now in circulation [79]. Promkuntod et al. [80] reported that 62.5% (15/24) of Thailand isolates are QX-like variants, demonstrating that QX-like strains have been predominant since 2009.
In India, Patel et al. [81] isolated a strain similar, but genetically distinct, to the Mass type vaccine M41 strain. The first 4/91 type strain in India was isolated by Sumi et al. [82], which may be a new vaccine strain variant based on its genomic sequence.
Ganapathy et al. [83] demonstrated that six IBV genotypes were detected in the Middle East from 2009–2014: 4/91, IS/1494/06, Mass, IS/885/00, Q1 and D274. In total, 32.87% and 18.87% of the S1 genes of the 4/91 and Mass type strains, respectively, were more than 99% homologous with vaccine strains, indicating the genetic mutation of vaccine strains. The authors of this study also noted that 4/91 field strain isolates identified after 2012 were differently clustered with former isolates. Other researchers have reported the appearance of some China-like strains in the Middle East, such as strains similar to CK/CH/Guangdong/Xindadi/0903 [84] and CK/CH/LDL/97I [85] based on their S1 sequences. Interestingly, the LDL/97I-like strains that were first isolated in the Middle East exhibited more extensive tissue tropism, as they were detectable in trachea, kidneys, ovarian tissue, and cecal tonsils, than the original LDL/97I strain that was limited to the respiratory system and kidneys.
5. Africa
In Egypt, IBV isolates are mainly divided into the Egy/Var I, Egy/Var II and Mass type groups [38]. The Egy/Var I and Egy/Var II variants, which recombined from the original Egyptian variant and the Israeli strain, are grouped with Middle Eastern IBV strains. In addition, these variants showed high virulence in one-day-old SPF chickens, with 50% mortality. Furthermore, another study on Egy/Var type isolates identified a deletion at position 63, a substitution at I69A/S, and an additional N-glycosylation site in the S1 protein [11]. The first QX-like strains were confirmed in Zimbabwe in 2011 [86], and Italy 02 type strains were first detected in Africa (Morocco) by Fellahi et al. [87] in 2014. At that time, the proportion of IBV Italy 02 type strains was 32%, and they quickly became the second most prevalent genotype in Morocco. However, Moroccan Italy 02 isolates were slightly different from European Italy 02 strains based on their S1 genes, according to phylogenetic tree analysis.
6. Europe
QX strains were first detected in Europe in the Netherlands between 2003 and 2004, and their proportionality has since sharply increased. Over the past decade, QX-like strains, characterized as European QX because of some nucleotide substitutions, have been detected in the United Kingdom, Finland, Hungary, Russia, Slovenia, Spain, Sweden and Switzerland [44,88,89,90,91,92,93,94]. Pohjola et al. [90] found a unique 117LDKG120 sequence that appeared to be a recombination between a QX strain and an unknown IBV strain in Finland, a country in which IBV is not vaccinated against. Abro et al. [88] dissected a 2010 isolated Swedish QX strain, CK/SWE/0658946/10, and found several regions, especially amino acids 1650–2850, 3735–3940 and 4915–5285 located in the 1ab gene, that underwent strong positive selective pressure and had numerous non-synonymous substitutions. In Russia, three QX-like IBVs were found that contained recombinant S1 genes from the vaccine strains H120, 4/91, and D274 [92].
The Q1 strain was initially isolated in China by Yu et al. [95] between 1996 and 1998 in young layers exhibiting proventriculitis and respiratory symptoms, but no kidney lesions. This strain did not circulate outside Asia but was detected in 15-day-old broiler chickens in Italy in 2011 [96]. Furthermore, Q1-like isolates in Italy targeted the proventriculus and kidney [42]. Another study analyzed the full-length genome of the Q1-like strain γCoV/Ck/Italy/I2022/13, which was isolated in Italy in 2013, and noted multiple recombination events [43]. Evidence clearly shows the circulation of variant Q1 strains in Italy.
As a country that does not vaccinate against IBV, Finland has been exempt from IBV clinical cases since the 1970s. However, several vaccine-like pathogenic strains, including the D274 and 4/91 strains, were found in a 2011 outbreak, indicating the variation of vaccine strains [90]. Finland is now considering improving their vaccination process with live attenuated vaccines.
According to Krapez et al. [91], the predominant IBV strains in Slovenia from 1990 to 2005 were 624/I type strains (9/15, 60%), which can be distinctly clustered with strains of earlier isolation periods and geographical origin, based on the S1 gene.
7. America
Many types of commercial vaccine strains have originated from the United States, such as the Mass, Conn, and Arkansas (Ark) types. A nephropathogenic IBV strain, DMV/1639/11, isolated from Delmarva in 2011 was similar to the strain responsible for the Pennsylvania IBV outbreak from 1997 to 2000. When the Mass vaccine type was combined with the Conn or Ark types in a laboratory setting via the eye-drop method, the virus shedding rate and renal lesions in broiler chickens was decreased after DMV/1639/11 challenge. However, in the commercial setting of live vaccine spraying, trachea and kidney protection was not observed [97]. While the Cal99 strain usually causes only respiratory symptoms, a Cal99 variant was found to be nephropathogenic and could spread to respiratory and gastrointestinal tracts, and the bursa [40]. A novel IBV genotype in Georgia was determined to have S1 gene sequences from both the Australian-isolated N1-62 strain and the Ark DPI strain [98].
The IBV strains in Canada can be divided into four groups: Canadian variants, vaccine-like, US variant-like, non-Canadian, and non-US. While the current Mass and Conn type vaccines can provide satisfactory protection results to most of the groups, the emergence and circulation of the 4/91 type indicates the need for a 4/91 type vaccine [99].
Brazil has reported the most IBV isolates in North America. Brazilian IBV strains form a unique Brazilian cluster that includes three subclusters, Brazil 01, 02 and 03 [100], but their genetic variation was low for a long time [101]. The D207, Mass, Conn, and Ark serotypes have also been found in Brazil [102], and the 4/91 strain was first documented by Villarreal et al. [100]. Analysis of IBV-positive rates in different tissues showed the highest presence of IBV in the digestive system (43.5%), followed by the respiratory system (37.7%) [103]. The proventriculus type Q1-like strain was found in other South American countries, such as Argentina and Uruguay [104].
8. Oceania
IBV evolution is independent in Australia because of its geographical isolation. IBV strains in Australia can be classified into three groups based on the S1 gene: Australian group 1, containing vaccine and vaccine-like strains, group 2, emerging from the 1980s to the 1990s, and group 3, containing newly isolated recombinant strains from groups 1 and 2. In 2016, however, two group 2 strains were isolated that had been undetectable for two decades [45,46]. Quinteros et al. [46] proposed the existence of an unknown IBV parental strain circulating in the field, which explains how the 1980s strains underwent recombination to fit new environments. Former studies have found that the open reading frames (ORFs) of Australian group 2 IBV strains were analogous to those of turkey coronaviruses in that they lack the 3a, 3b, 5a and 5b genes. However, while the recently isolated group 2 strain had the 3b and 5b genes, it contained genetic mutations that could influence transcription [105].
9. Conclusions
Among the studies reporting the IBV variants reviewed herein, mutations and recombination can take place in both the structural and the non-structural proteins, although the exact link of these variations to pathogenicity alternation is still unknown. Most of the mutations and recombination events have been detected in the S gene (54%, 15 out of 28 references reviewed) and then in the polyprotein 1a and 1ab (35.7%, 10 out of 28 references reviewed). In particular, the nsp 2–6 were more variable in these studies, consistent with previous observations [54].
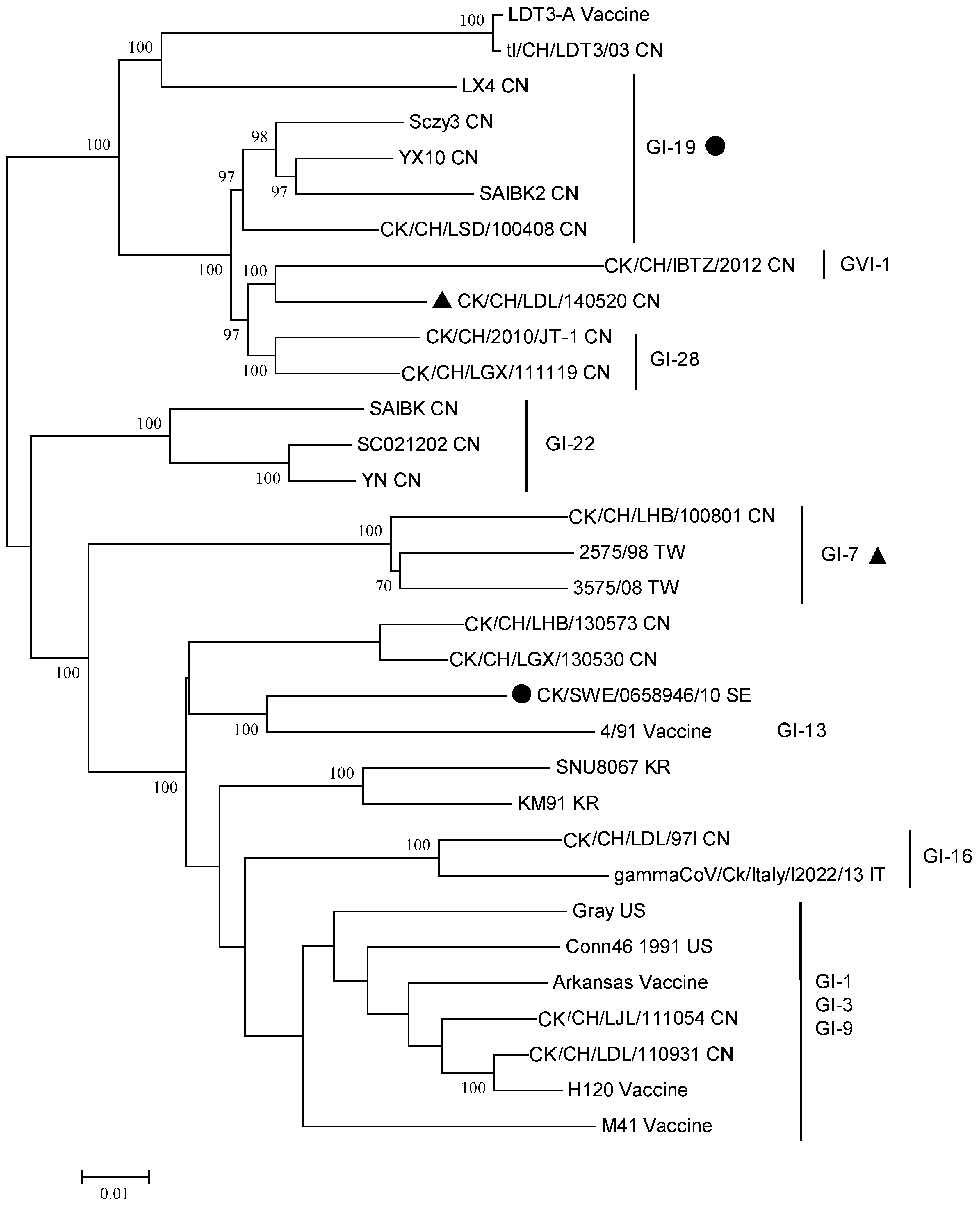
Phylogeny of the reference strains and the IBV variants reviewed in this article is shown in Figure 1, based on classification scheme described by Valastro et al. [18]. The strain name, GenBank accession number and geographic origin are listed in the Supplementary Table S1. Interestingly, some IBV variants are not consistently classified in the same groups or lineages when analyzed using the full-length genomes (Figure 2). For instance, the S1 gene of the CK/CH/LGX/111119 belongs to the GI-7 lineage, however, the full genome of this variant appears more related to the GVI-1 or GI-28 lineage, suggesting the genome may comprise segments originated from other lineages, e.g., inter-lineage recombination. Similar findings are also observed in the strain CK/SWE/0658946/10. Thus, examining the full-length genomes of novel IBV strains and understanding their contextual sequences are of equal importance.
In studies correlating genotypes, serotypes and pathogenic types, genotyping and serotyping were consistent in most cases. In addition, according to Jackwood et al. [106] and Lee et al. [107], various serotypes defined by neutralization were more than 10% genetically different, with some exceptions [23,26,27,34,35]. Cavanagh et al. [3] noted that only a few amino acid substitutions, resulting in 2% sequence divergence in the S1 gene, can alter the serotype, and determining the pathogenic type according to genotype is risky. Evidence indicates that non-structural proteins, accessory proteins, and nucleocapsid proteins can influence viral replication or host-antigen interactions in different phases, thereby influencing virulence. Examples include inhibition of the signal transducer and activator of transcription 1 (STAT 1) signaling pathway, interferon production [108,109], and modulation of viral transcriptional and translational levels [110]. For protectotype identification, determining serotypes by virus neutralization is not sufficient, since systemic antibody, mucosal, and cellular immune responses must be considered together to evaluate immune protection. When investigating the pathogenicity of IBV variants, a tendency for the virus to infect more extensive tissues was found. IBV was first identified as a respiratory system pathogen, and then became pathogenic against the renal and reproductive systems, and the proventriculus. Among these, proventriculus infection greatly decreases the feed conversion rate, and infection of the reproductive system, such as the oviduct, results in the so-called false layer syndrome, resulting in large economic losses in the poultry industry.
Over the past decade, IBV variants have continuously emerged, and most have been isolated from chicken flocks receiving primary or boost immunizations. Different efficacies of the same types of vaccine immunizations created in both the laboratory and in commercial settings serve as a warning [97] that an immune strategy urgently needs to be refined to choose appropriate vaccines and effective immune routes. Many of the IBV variants described in this review have not emerged via accumulated point mutations, but rather by recombination with other existing strains. While the existing strains may have been discovered decades ago and have not circulated in their original regions since that time, they have not vanished and are still evolving, seeking an appropriate environment to cause outbreaks. This worldwide recombination may result from the continuous expansion of global poultry product trade and the migration of people and wild birds, carrying IBV antigens all around the world, and thus provides evidence that environmental selection deeply influences IBV variation. Facing this complex situation, we must admit the importance of developing potent IBV vaccines to ensure biosecurity and global cooperation. This review of IBV variants has practical value for understanding their phylogenetic relationships and epidemiology from both regional and worldwide viewpoints.
Supplementary Materials
Supplementary materials can be found at www.mdpi.com/1422-0067/18/10/2030/s1.
Acknowledgments
This work was supported by the Ministry of Science and Technology in Taiwan (103-2321-B-002-066, 104-2321-B-002-023, 105-2321-B-002-007, 106-2311-B-002-030-MY3).
Author Contributions
Shu-Yi Lin wrote the manuscript, Hui-Wen Chen edited and approved the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schalk, A.; Hawn, M. An apparently new respiratory disease of baby chicks. J. Am. Vet. Med. Assoc. 1931, 78, 413–422. [Google Scholar]
- Cavanagh, D.; Davis, P.J.; Mockett, A.P.A. Amino-Acids within Hypervariable Region-1 of Avian Coronavirus Ibv (Massachusetts Serotype) Spike Glycoprotein Are Associated with Neutralization Epitopes. Virus Res. 1988, 11, 141–150. [Google Scholar] [CrossRef]
- Cavanagh, D.; Elus, M.M.; Cook, J.K. Relationship between sequence variation in the S1 spike protein of infectious bronchitis virus and the extent of cross-protection in vivo. Avian Pathol. 1997, 26, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Davis, P.J. Coronavirus IBV: Removal of spike glycopolypeptide S1 by urea abolishes infectivity and haemagglutination but not attachment to cells. J. Gen. Virol. 1986, 67, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Promkuntod, N.; Van Eijndhoven, R.; De Vrieze, G.; Gröne, A.; Verheije, M. Mapping of the receptor-binding domain and amino acids critical for attachment in the spike protein of avian coronavirus infectious bronchitis virus. Virology 2014, 448, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Casais, R.; Dove, B.; Cavanagh, D.; Britton, P. Recombinant avian infectious bronchitis virus expressing a heterologous spike gene demonstrates that the spike protein is a determinant of cell tropism. J. Virol. 2003, 77, 9084–9089. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cai, H.; Pan, J.A.; Xiang, N.; Tien, P.; Ahola, T.; Guo, D. Functional screen reveals SARS coronavirus nonstructural protein nsp14 as a novel cap N7 methyltransferase. Proc. Natl. Acad. Sci. USA 2009, 106, 3484–3489. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Chen, G.; Guo, B.; Cheng, G.; Tang, H. PLP2, a potent deubiquitinase from murine hepatitis virus, strongly inhibits cellular type I interferon production. Cell Res. 2008, 18, 1105–1113. [Google Scholar] [PubMed]
- Seo, S.H.; Wang, L.; Smith, R.; Collisson, E.W. The carboxyl-terminal 120-residue polypeptide of infectious bronchitis virus nucleocapsid induces cytotoxic T lymphocytes and protects chickens from acute infection. J. Virol. 1997, 71, 7889–7894. [Google Scholar] [PubMed]
- Yang, T.; Wang, H.-N.; Wang, X.; Tang, J.-N.; Lu, D.; Zhang, Y.-F.; Guo, Z.-C.; Li, Y.-L.; Gao, R.; Kang, R.-M. The protective immune response against infectious bronchitis virus induced by multi-epitope based peptide vaccines. Biosci. Biotechnol. Biochem. 2009, 73, 1500–1504. [Google Scholar] [CrossRef] [PubMed]
- Archetti, I.; Horsfall, F.L., Jr. Persistent antigenic variation of influenza A viruses after incomplete neutralization in ovo with heterologous immune serum. J. Exp. Med. 1950, 92, 441. [Google Scholar] [CrossRef] [PubMed]
- Ruano, M.; El-Attrache, J.; Villegas, P. A rapid-plate hemagglutination assay for the detection of infectious bronchitis virus. Avian Dis. 2000, 44, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Karaca, K.; Naqi, S. A monoclonal antibody blocking ELISA to detect serotype-specific infectious bronchitis virus antibodies. Vet. Microbiol. 1993, 34, 249–257. [Google Scholar] [CrossRef]
- Chen, H.W.; Wang, C.H.; Cheng, I.C. A type-specific blocking ELISA for the detection of infectious bronchitis virus antibody. J. Virol. Methods 2011, 173, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Evaluation of methods for detecting recombination from DNA sequences: Computer simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef] [PubMed]
- Ladman, B.S.; Loupos, A.B.; Gelb, J., Jr. Infectious bronchitis virus S1 gene sequence comparison is a better predictor of challenge of immunity in chickens than serotyping by virus neutralization. Avian Pathol. 2006, 35, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Davis, P.; Cook, J.K.; Li, D.; Kant, A.; Koch, G. Location of the amino acid differences in the S1 spike glycoprotein subunit of closely related serotypes of infectious bronchitis virus. Avian Pathol. 1992, 21, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Naqi, S. Infectious Bronchitis; John Wiley & Sons: Hoboken, NJ, USA, 2003; Volume 11, pp. 101–119. [Google Scholar]
- Chen, Y.; Jiang, L.; Zhao, W.; Liu, L.; Zhao, Y.; Shao, Y.; Li, H.; Han, Z.; Liu, S. Identification and molecular characterization of a novel serotype infectious bronchitis virus (GI-28) in China. Vet. Microbiol. 2017, 198, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhang, M.; Tian, X.; Shao, H.; Qian, K.; Ye, J.; Qin, A. Identification of a novel recombinant virulent avian infectious bronchitis virus. Vet. Microbiol. 2017, 199, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, X.; Xu, P.; Zhou, L.; Zhang, Z.; Wang, H. Genome sequence and origin analyses of the recombinant novel IBV virulent isolate SAIBK2. Virus Genes 2016, 52, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, T.; Han, Z.; Liang, S.; Xu, Y.; Xu, Q.; Chen, Y.; Zhao, Y.; Shao, Y.; Li, H.; et al. Molecular and antigenic characteristics of Massachusetts genotype infectious bronchitis coronavirus in China. Vet. Microbiol. 2015, 181, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Hu, Y.X.; Jin, J.H.; Zhao, Y.; Zhao, J.; Zhang, G.Z. Pathogenicity of virulent infectious bronchitis virus isolate YN on hen ovary and oviduct. Vet. Microbiol. 2016, 193, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Hu, Y.; Ma, Z.; Yu, Q.; Zhao, J.; Liu, X.; Zhang, G. Virulent avian infectious bronchitis virus, People’s Republic of China. Emerg. Infect. Dis. 2012, 18, 1994–2001. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Liu, X.Y.; Zhao, Y.; Chen, Y.; Zhao, J.; Zhang, G.Z. Characterization and analysis of an infectious bronchitis virus strain isolated from southern China in 2013. Virol. J. 2016, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.H.; Chen, Y.; Zhao, J.; Xu, G.; Zhao, Y.; Zhang, G.Z. Pathogenicity of a TW-Like Strain of Infectious Bronchitis Virus and Evaluation of the Protection Induced against It by a QX-Like Strain. Front. Microbiol. 2016, 7, 1653. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Wang, Q.; Zhao, W.; Chen, Y.; Zhang, T.; Han, Z.; Xu, Q.; Kong, X.; Liu, S. Serotype, antigenicity, and pathogenicity of a naturally recombinant TW I genotype infectious bronchitis coronavirus in China. Vet. Microbiol. 2016, 191, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Zhang, T.; Xu, Q.; Gao, M.; Chen, Y.; Wang, Q.; Zhao, Y.; Shao, Y.; Li, H.; Kong, X.; et al. Altered pathogenicity of a tl/CH/LDT3/03 genotype infectious bronchitis coronavirus due to natural recombination in the 5′- 17kb region of the genome. Virus Res. 2016, 213, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Xu, Q.; Han, Z.; Liu, X.; Li, H.; Guo, H.; Sun, N.; Shao, Y.; Kong, X. Origin and characteristics of the recombinant novel avian infectious bronchitis coronavirus isolate ck/CH/LJL/111054. Infect. Genet. Evol. 2014, 23, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Tang, M.; Jiang, Y.; Chen, X.; Shen, X.; Li, J.; Dai, Y.; Zou, J. Complete genome sequence of a novel infectious bronchitis virus strain circulating in China with a distinct S gene. Virus Genes 2014, 49, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zou, N.; Wang, F.; Guo, M.; Liu, P.; Wen, X.; Cao, S.; Huang, Y. Analysis of a QX-like avian infectious bronchitis virus genome identified recombination in the region containing the ORF 5a, ORF 5b, and nucleocapsid protein gene sequences. Virus Genes 2013, 46, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.T.; Ma, B.C. Complete genome sequence of a nephropathogenic infectious bronchitis virus strain isolated in china. Genome Announc. 2013, 1, e00815-13. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Li, Y.T.; Chen, Y.T.; Chen, T.C.; Hu, C.J.; Chen, H.W. Identification of an infectious bronchitis coronavirus strain exhibiting a classical genotype but altered antigenicity, pathogenicity, and innate immunity profile. Sci. Rep. 2016, 6, 37725. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, Y.; Horiuchi, T.; Harada, M.; Tanimura, C.; Seki, Y.; Kuroda, Y.; Yagyu, K.; Nakamura, S.; Suzuki, S. Isolation of 4/91 type of infectious bronchitis virus as a new variant in Japan and efficacy of vaccination against 4/91 type field isolate. Avian Dis. 2008, 52, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.M.; Kwon, H.J.; Kim, I.H.; Mo, M.L.; Kim, J.H. Comparative genomics of Korean infectious bronchitis viruses (IBVs) and an animal model to evaluate pathogenicity of IBVs to the reproductive organs. Viruses 2012, 4, 2670–2683. [Google Scholar] [CrossRef] [PubMed]
- Khataby, K.; Kichou, F.; Loutfi, C.; Ennaji, M.M. Assessment of pathogenicity and tissue distribution of infectious bronchitis virus strains (Italy 02 genotype) isolated from moroccan broiler chickens. BMC Vet. Res. 2016, 12, 94. [Google Scholar] [CrossRef] [PubMed]
- Zanaty, A.; Arafa, A.S.; Hagag, N.; El-Kady, M. Genotyping and pathotyping of diversified strains of infectious bronchitis viruses circulating in Egypt. World J. Virol. 2016, 5, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Zanaty, A.; Naguib, M.M.; El-Husseiny, M.H.; Mady, W.; Hagag, N.; Arafa, A.S. The sequence of the full spike S1 glycoprotein of infectious bronchitis virus circulating in Egypt reveals evidence of intra-genotypic recombination. Arch. Virol. 2016, 161, 3583–3587. [Google Scholar] [CrossRef] [PubMed]
- Franca, M.; Woolcock, P.R.; Yu, M.; Jackwood, M.W.; Shivaprasad, H.L. Nephritis associated with infectious bronchitis virus Cal99 variant in game chickens. Avian Dis. 2011, 55, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Chacon, J.L.; Assayag, M.S., Jr.; Revolledo, L.; Astolfi-Ferreira, C.S.; Vejarano, M.P.; Jones, R.C.; Piantino Ferreira, A.J. Pathogenicity and molecular characteristics of infectious bronchitis virus (IBV) strains isolated from broilers showing diarrhoea and respiratory disease. Brit. Poult. Sci. 2014, 55, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Bonci, M.; Bano, L.; Bano, L.; Valastro, V.; Vascellari, M.; Capua, I.; Terregino, C. Diagnostic and clinical observation on the infectious bronchitis virus strain Q1 in Italy. Vet. Ital. 2013, 49, 347–355. [Google Scholar] [PubMed]
- Franzo, G.; Listorti, V.; Naylor, C.J.; Lupini, C.; Laconi, A.; Felice, V.; Drigo, M.; Catelli, E.; Cecchinato, M. Molecular investigation of a full-length genome of a Q1-like IBV strain isolated in Italy in 2013. Virus Res. 2015, 210, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, K.; Wilkins, M.; Forrester, A.; Lemiere, S.; Cserep, T.; McMullin, P.; Jones, R.C. QX-like infectious bronchitis virus isolated from cases of proventriculitis in commercial broilers in England. Vet. Rec. 2012, 171, 597. [Google Scholar] [CrossRef] [PubMed]
- Hewson, K.A.; Noormohammadi, A.H.; Devlin, J.M.; Browning, G.F.; Schultz, B.K.; Ignjatovic, J. Evaluation of a novel strain of infectious bronchitis virus emerged as a result of spike gene recombination between two highly diverged parent strains. Avian Pathol. 2014, 43, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Quinteros, J.A.; Lee, S.W.; Markham, P.F.; Noormohammadi, A.H.; Hartley, C.A.; Legione, A.R.; Coppo, M.J.; Vaz, P.K.; Browning, G.F. Full genome analysis of Australian infectious bronchitis viruses suggests frequent recombination events between vaccine strains and multiple phylogenetically distant avian coronaviruses of unknown origin. Vet. Microbiol. 2016, 197, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Mayr, E. Toward a New Philosophy of Biology: Observations of an Evolutionist; Harvard University Press: Cambridge, MA, USA, 1988. [Google Scholar]
- Smith, E.C.; Blanc, H.; Vignuzzi, M.; Denison, M.R. Coronaviruses lacking exoribonuclease activity are susceptible to lethal mutagenesis: Evidence for proofreading and potential therapeutics. PLoS Pathog. 2013, 9, e1003565. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.R.; Graham, R.L.; Donaldson, E.F.; Eckerle, L.D.; Baric, R.S. Coronaviruses: An RNA proofreading machine regulates replication fidelity and diversity. RNA Biol. 2011, 8, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Minskaia, E.; Hertzig, T.; Gorbalenya, A.E.; Campanacci, V.; Cambillau, C.; Canard, B.; Ziebuhr, J. Discovery of an RNA virus 3′→5′ exoribonuclease that is critically involved in coronavirus RNA synthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 5108–5113. [Google Scholar] [CrossRef] [PubMed]
- Toro, H.; van Santen, V.L.; Jackwood, M.W. Genetic diversity and selection regulates evolution of infectious bronchitis virus. Avian Dis. 2012, 56, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, R.A.; Van Santen, V.L.; Toro, H. Host intraspatial selection of infectious bronchitis virus populations. Avian Dis. 2010, 54, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Shahwan, K.; Hesse, M.; Mork, A.K.; Herrler, G.; Winter, C. Sialic acid binding properties of soluble coronavirus spike (S1) proteins: Differences between infectious bronchitis virus and transmissible gastroenteritis virus. Viruses 2013, 5, 1924–1933. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, H.; Zhao, J.; Zhong, Q.; Jin, J.H.; Zhang, G.Z. Evolution of infectious bronchitis virus in China over the past two decades. J. Gen. Virol. 2016, 97, 1566–1574. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Tsai, C.T. Genetic grouping for the isolates of avian infectious bronchitis virus in Taiwan. Arch. Virol. 1996, 141, 1677–1688. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Huang, Y.C. Relationship between serotypes and genotypes based on the hypervariable region of the S1 gene of infectious bronchitis virus. Arch. Virol. 2000, 145, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.M.; Wang, C.H.; Hou, M.H.; Huang, Y.P.; Kao, H.W.; Su, H.L. Evolution of infectious bronchitis virus in Taiwan: Characterisation of RNA recombination in the nucleocapsid gene. Vet. Microbiol. 2010, 144, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-W.; Huang, Y.-P.; Wang, C.-H. Identification of Taiwan and China-like recombinant avian infectious bronchitis viruses in Taiwan. Virus Res. 2009, 140, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.T.; Tsai, H.F.; Wang, C.H. Detection of infectious bronchitis virus strains similar to Japan in Taiwan. J. Vet. Med. Sci. 2016, 78, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.W.; Huang, Y.P.; Wang, C.H. Identification of intertypic recombinant infectious bronchitis viruses from slaughtered chickens. Poult. Sci. 2010, 89, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Shao, Y.; Sun, C.; Han, Z.; Liu, X.; Guo, H.; Liu, X.; Kong, X.; Liu, S. Genetic diversity of avian infectious bronchitis coronavirus in recent years in China. Avian Dis. 2012, 56, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Yudong, W.; YongLin, W.; Zichun, Z.; GenChe, F.; Yihai, J.; Xiange, L.; Jiang, D.; Shushuang, W. Isolation and identification of glandular stomach type IBV (QX IBV) in chickens. Chin. J. Anim. Quarantine 1998, 15, 1–3. [Google Scholar]
- Sun, C.; Han, Z.; Ma, H.; Zhang, Q.; Yan, B.; Shao, Y.; Xu, J.; Kong, X.; Liu, S. Phylogenetic analysis of infectious bronchitis coronaviruses newly isolated in China, and pathogenicity and evaluation of protection induced by Massachusetts serotype H120 vaccine against QX-like strains. Avian Pathol. 2011, 40, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Sun, C.; Yan, B.; Zhang, X.; Wang, Y.; Li, C.; Zhang, Q.; Ma, Y.; Shao, Y.; Liu, Q.; et al. A 15-year analysis of molecular epidemiology of avian infectious bronchitis coronavirus in China. Infect. Genet. Evol. 2011, 11, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Xue, Y.; Wang, F.; Chen, F.; Shu, D.; Xie, Q. Analysis of S1 gene of avian infectious bronchitis virus isolated in southern China during 2011–2012. Virus Genes 2014, 49, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xue, C.; Chen, F.; Qin, J.; Xie, Q.; Bi, Y.; Cao, Y. Isolation and genetic analysis revealed no predominant new strains of avian infectious bronchitis virus circulating in South China during 2004–2008. Vet. Microbiol. 2010, 143, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Xie, J.; Chen, F.; Shu, D.; Zuo, K.; Xue, C.; Qin, J.; Li, H.; Bi, Y.; Ma, J. Phylogenetic distribution and predominant genotype of the avian infectious bronchitis virus in China during 2008–2009. Virol. J. 2011, 8, 184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, Y.; Wang, H.; Zeng, F.; Yang, X.; Zhang, Y.; Zhang, A. Molecular detection and Smoothing spline clustering of the IBV strains detected in China during 2011–2012. Virus Res. 2016, 211, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Mo, M.L.; Li, M.; Huang, B.C.; Fan, W.S.; Wei, P.; Wei, T.C.; Cheng, Q.Y.; Wei, Z.J.; Lang, Y.H. Molecular characterization of major structural protein genes of avian coronavirus infectious bronchitis virus isolates in southern china. Viruses 2013, 5, 3007–3020. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; He, X.; Yao, K.C.; Du, L.J.; Liu, P.; Yan, Q.G.; Wen, Y.P.; Cao, S.J.; Han, X.F.; Huang, Y. Phylogenetic and antigenic analysis of avian infectious bronchitis virus in southwestern China, 2012–2016. Infect. Genet. Evol. 2016, 45, 11–19. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Li, M.; Wei, P.; Mo, M.L.; Wei, T.C.; Li, K.R. Complete genome sequence of an infectious bronchitis virus chimera between cocirculating heterotypic strains. J. Virol. 2012, 86, 13887–13888. [Google Scholar] [CrossRef] [PubMed]
- Zou, N.L.; Zhao, F.F.; Wang, Y.P.; Liu, P.; Cao, S.J.; Wen, X.T.; Huang, Y. Genetic analysis revealed LX4 genotype strains of avian infectious bronchitis virus became predominant in recent years in Sichuan area, China. Virus Genes 2010, 41, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Qin, J.; Chen, F.; Xie, Q.; Bi, Y.; Cao, Y.; Xue, C. Phylogenetic analysis of the S1 glycoprotein gene of infectious bronchitis viruses isolated in China during 2009–2010. Virus Genes 2012, 44, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Yamaguchi, S.; Imada, T. Existence of avian infectious bronchitis virus with a European-prevalent 4/91 genotype in Japan. J. Vet. Med. Sci. 2008, 70, 1341–1344. [Google Scholar]
- Shimazaki, Y.; Watanabe, Y.; Harada, M.; Seki, Y.; Kuroda, Y.; Fukuda, M.; Honda, E.; Suzuki, S.; Nakamura, S. Genetic analysis of the S1 gene of 4/91 type infectious bronchitis virus isolated in Japan. J. Vet. Med. Sci. 2009, 71, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Mase, M.; Kawanishi, N.; Ootani, Y.; Murayama, K.; Karino, A.; Inoue, T.; Kawakami, J. A novel genotype of avian infectious bronchitis virus isolated in Japan in 2009. J. Vet. Med. Sci. 2010, 72, 1265–1268. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.-H.; Lee, H.-J.; Lee, D.-H.; Lee, Y.-N.; Park, J.-K.; Youn, H.-N.; Kim, M.-S.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S. An emerging recombinant cluster of nephropathogenic strains of avian infectious bronchitis virus in Korea. Infect. Genet. Evol. 2011, 11, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.M.; Kwon, H.J.; Choi, K.S.; Kim, J.H. Comparative genomics of QX-like infectious bronchitis viruses in Korea. Arch. Virol. 2017, 162, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Pohuang, T.; Chansiripornchai, N.; Tawatsin, A.; Sasipreeyajan, J. Sequence analysis of S1 genes of infectious bronchitis virus isolated in Thailand during 2008–2009: Identification of natural recombination in the field isolates. Virus Genes 2011, 43, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Promkuntod, N.; Thongmee, S.; Yoidam, S. Analysis of the S1 gene of the avian infectious bronchitis virus (IBV) reveals changes in the IBV genetic groups circulating in southern Thailand. Res. Vet. Sci. 2015, 100, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.H.; Bhimani, M.P.; Bhanderi, B.B.; Jhala, M.K. Isolation and molecular characterization of nephropathic infectious bronchitis virus isolates of Gujarat state, India. Virus Disease 2015, 26, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Sumi, V.; Singh, S.D.; Dhama, K.; Gowthaman, V.; Barathidasan, R.; Sukumar, K. Isolation and molecular characterization of infectious bronchitis virus from recent outbreaks in broiler flocks reveals emergence of novel strain in India. Trop. Anim. Health Prod. 2012, 44, 1791–1795. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, K.; Ball, C.; Forrester, A. Genotypes of infectious bronchitis viruses circulating in the Middle East between 2009 and 2014. Virus Res. 2015, 210, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Seger, W.; GhalyanchiLangeroudi, A.; Karimi, V.; Madadgar, O.; Marandi, M.V.; Hashemzadeh, M. Genotyping of infectious bronchitis viruses from broiler farms in Iraq during 2014–2015. Arch. Virol. 2016, 161, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Ababneh, M.; Dalab, A.E.; Alsaad, S.; Al-Zghoul, M. Presence of Infectious Bronchitis Virus Strain CK/CH/LDL/97I in the Middle East. ISRN Vet. Sci. 2012, 2012, 201721. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Monne, I.; Terregino, C.; Cattoli, G.; Hodobo, C.T.; Gadaga, B.; Makaya, P.V.; Mdlongwa, E.; Swiswa, S. QX-like infectious bronchitis virus in Africa. Vet. Rec. 2011, 169, 589. [Google Scholar] [CrossRef] [PubMed]
- Fellahi, S.; Ducatez, M.; El Harrak, M.; Guerin, J.L.; Touil, N.; Sebbar, G.; Bouaiti el, A.; Khataby, K.; Ennaji, M.M.; El-Houadfi, M. Prevalence and molecular characterization of avian infectious bronchitis virus in poultry flocks in Morocco from 2010 to 2014 and first detection of Italy 02 in Africa. Avian Pathol. 2015, 44, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Abro, S.H.; Renstrom, L.H.; Ullman, K.; Belak, S.; Baule, C. Characterization and analysis of the full-length genome of a strain of the European QX-like genotype of infectious bronchitis virus. Arch. Virol. 2012, 157, 1211–1215. [Google Scholar] [CrossRef] [PubMed]
- Irvine, R.M.; Cox, W.J.; Ceeraz, V.; Reid, S.M.; Ellis, R.J.; Jones, R.M.; Errington, J.; Wood, A.M.; McVicar, C.; Clark, M.I. Detection of IBV QX in commercial broiler flocks in the UK. Vet. Rec. 2010, 167, 877–879. [Google Scholar] [CrossRef] [PubMed]
- Pohjola, L.K.; Ek-Kommonen, S.C.; Tammiranta, N.E.; Kaukonen, E.S.; Rossow, L.M.; Huovilainen, T.A. Emergence of avian infectious bronchitis in a non-vaccinating country. Avian Pathol. 2014, 43, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Krapez, U.; Slavec, B.; Barlic-Maganja, D.; Rojs, O.Z. Molecular analysis of infectious bronchitis viruses isolated in Slovenia between 1990 and 2005: A retrospective study. Virus Genes 2010, 41, 414–416. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, E.V.; Bochkov, Y.A.; Shcherbakova, L.O.; Nikonova, Z.B.; Zinyakov, N.G.; Elatkin, N.P.; Mudrak, N.S.; Borisov, A.V.; Drygin, V.V. Molecular characterization of infectious bronchitis virus isolates from Russia and neighbouring countries: Identification of intertypic recombination in the S1 gene. Avian Pathol. 2011, 40, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.; Franzo, G.; Massi, P.; Tosi, G.; Blanco, A.; Antilles, N.; Biarnes, M.; Majo, N.; Nofrarias, M.; Dolz, R.; et al. A novel variant of the infectious bronchitis virus resulting from recombination events in Italy and Spain. Avian Pathol. 2017, 46, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Valastro, V.; Monne, I.; Fasolato, M.; Cecchettin, K.; Parker, D.; Terregino, C.; Cattoli, G. QX-type infectious bronchitis virus in commercial flocks in the UK. Vet. Rec. 2010, 167, 865–866. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Jiang, Y.; Low, S.; Wang, Z.; Nam, S.J.; Liu, W.; Kwang, J. Characterization of three infectious bronchitis virus isolates from China associated with proventriculus in vaccinated chickens. Avian Dis. 2001, 45, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Terregino, C.; Mazzacan, E.; Castaldello, I.; Capua, I.; Bonci, M. Detection of Chinese Q1 strain of infectious bronchitis virus in Europe. Vet. Rec. 2011, 169, 212–213. [Google Scholar] [CrossRef] [PubMed]
- Gelb, J., Jr.; Ladman, B.S.; Pope, C.R.; Ruano, J.M.; Brannick, E.M.; Bautista, D.A.; Coughlin, C.M.; Preskenis, L.A. Characterization of nephropathogenic infectious bronchitis virus DMV/1639/11 recovered from Delmarva broiler chickens in 2011. Avian Dis. 2013, 57, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.B.; Resurreccion, R.S. Genotyping of newly isolated infectious bronchitis virus isolates from northeastern Georgia. Avian Dis. 2010, 54, 1144–1151. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.A.; Brash, M.L.; Hoyland, S.K.; Coventry, J.M.; Sandrock, C.; Guerin, M.T.; Ojkic, D. Genotyping of infectious bronchitis viruses identified in Canada between 2000 and 2013. Avian Pathol. 2014, 43, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, L.Y.; Sandri, T.L.; Souza, S.P.; Richtzenhain, L.J.; de Wit, J.J.; Brandao, P.E. Molecular epidemiology of avian infectious bronchitis in Brazil from 2007 to 2008 in breeders, broilers, and layers. Avian Dis. 2010, 54, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Chacon, J.L.; Rodrigues, J.N.; Assayag Junior, M.S.; Peloso, C.; Pedroso, A.C.; Ferreira, A.J. Epidemiological survey and molecular characterization of avian infectious bronchitis virus in Brazil between 2003 and 2009. Avian Pathol. 2011, 40, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Felippe, P.A.; da Silva, L.H.; Santos, M.M.; Spilki, F.R.; Arns, C.W. Genetic diversity of avian infectious bronchitis virus isolated from domestic chicken flocks and coronaviruses from feral pigeons in Brazil between 2003 and 2009. Avian Dis. 2010, 54, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Balestrin, E.; Fraga, A.P.; Ikuta, N.; Canal, C.W.; Fonseca, A.S.; Lunge, V.R. Infectious bronchitis virus in different avian physiological systems-a field study in Brazilian poultry flocks. Poult. Sci. 2014, 93, 1922–1929. [Google Scholar] [CrossRef] [PubMed]
- Marandino, A.; Pereda, A.; Tomas, G.; Hernandez, M.; Iraola, G.; Craig, M.I.; Hernandez, D.; Banda, A.; Villegas, P.; Panzera, Y.; et al. Phylodynamic analysis of avian infectious bronchitis virus in South America. J. Gen. Virol. 2015, 96, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Hewson, K.A.; Ignjatovic, J.; Browning, G.F.; Devlin, J.M.; Noormohammadi, A.H. Infectious bronchitis viruses with naturally occurring genomic rearrangement and gene deletion. Arch. Virol. 2011, 156, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; Hilt, D.A.; Williams, S.M.; Woolcock, P.; Cardona, C.; O’Connor, R. Molecular and serologic characterization, pathogenicity, and protection studies with infectious bronchitis virus field isolates from California. Avian Dis. 2007, 51, 527–533. [Google Scholar] [CrossRef]
- Lee, C.-W.; Hilt, D.A.; Jackwood, M.W. Typing of field isolates of infectious bronchitis virus based on the sequence of the hypervariable region in the S1 gene. J. Vet. Diagn. Investig. 2003, 15, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Kint, J.; Dickhout, A.; Kutter, J.; Maier, H.J.; Britton, P.; Koumans, J.; Pijlman, G.P.; Fros, J.J.; Wiegertjes, G.F.; Forlenza, M. Infectious bronchitis coronavirus inhibits STAT1 signaling and requires accessory proteins for resistance to type I interferon activity. J. Virol. 2015, 89, 12047–12057. [Google Scholar] [CrossRef] [PubMed]
- Kint, J.; Langereis, M.A.; Maier, H.J.; Britton, P.; van Kuppeveld, F.J.; Koumans, J.; Wiegertjes, G.F.; Forlenza, M. Infectious bronchitis coronavirus limits interferon production by inducing a host shutoff that requires accessory protein 5b. J. Virol. 2016, 90, 7519–7528. [Google Scholar] [CrossRef] [PubMed]
- Kint, J.; Fernandez-Gutierrez, M.; Maier, H.J.; Britton, P.; Langereis, M.A.; Koumans, J.; Wiegertjes, G.F.; Forlenza, M. Activation of the chicken type I interferon response by infectious bronchitis coronavirus. J. Virol. 2015, 89, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
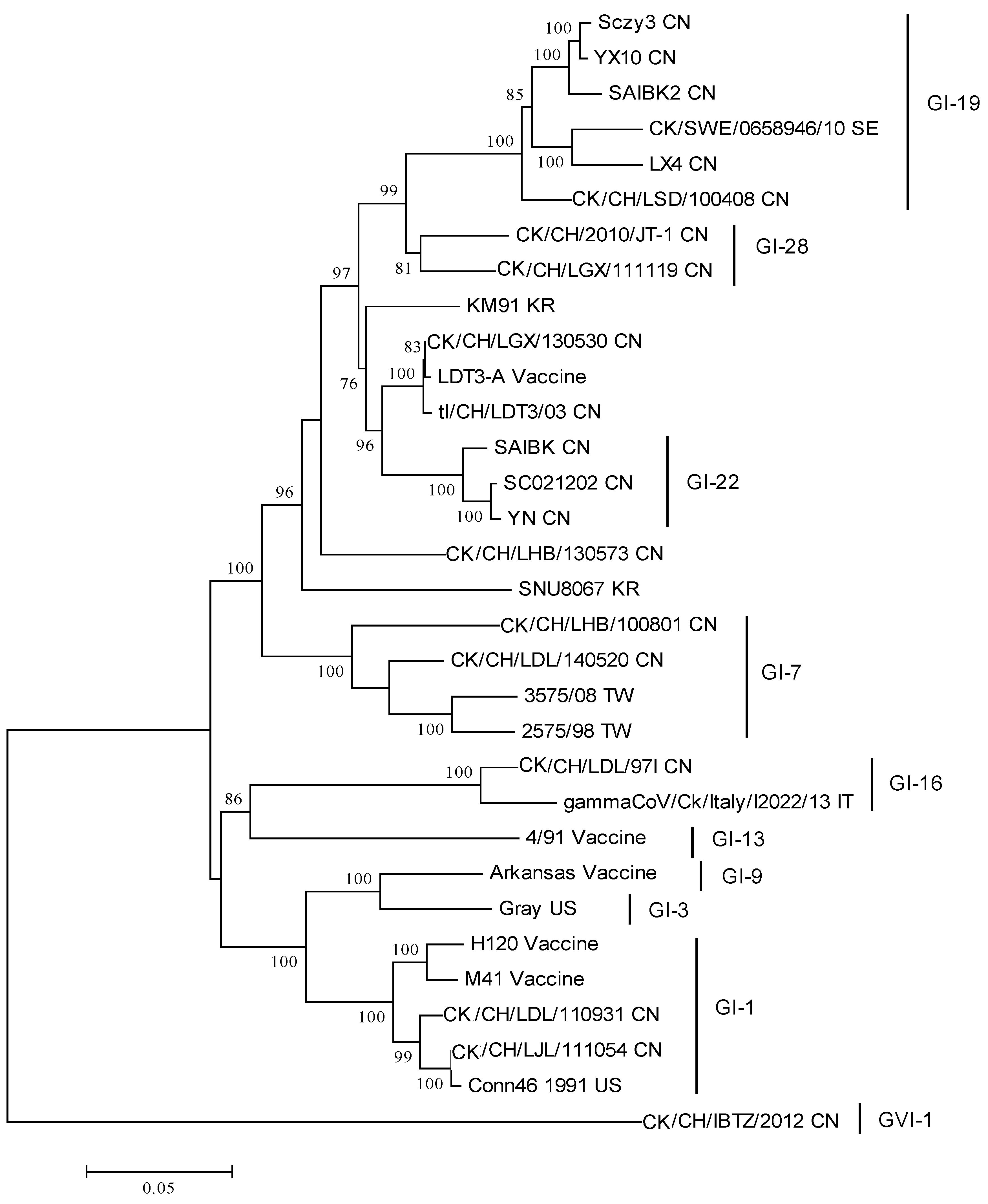
Figure 1.
Phylogenetic analysis of the complete S1 gene of the IBV variants reviewed in this study and reference strains. The phylogenetic tree was constructed using MEGA version 6 by the neighbor-joining method (bootstrapping for 1000 replicates, bootstrap value >70%). CN: China; IT: Italy; KR: Korea; SE: Sweden; TW: Taiwan; US: United States.
Figure 1.
Phylogenetic analysis of the complete S1 gene of the IBV variants reviewed in this study and reference strains. The phylogenetic tree was constructed using MEGA version 6 by the neighbor-joining method (bootstrapping for 1000 replicates, bootstrap value >70%). CN: China; IT: Italy; KR: Korea; SE: Sweden; TW: Taiwan; US: United States.

Figure 2.
Phylogenetic analysis of the full-length genome of the IBV variants reviewed in this study and reference strains. The strains with a symbol belong to the lineage with the same symbol when the S1 gene is analyzed. The phylogenetic tree was constructed using MEGA version 6 by the neighbor-joining method (bootstrapping for 1000 replicates, bootstrap value >70%). CN: China; IT: Italy; KR: Korea; SE: Sweden; TW: Taiwan; US: United States.
Figure 2.
Phylogenetic analysis of the full-length genome of the IBV variants reviewed in this study and reference strains. The strains with a symbol belong to the lineage with the same symbol when the S1 gene is analyzed. The phylogenetic tree was constructed using MEGA version 6 by the neighbor-joining method (bootstrapping for 1000 replicates, bootstrap value >70%). CN: China; IT: Italy; KR: Korea; SE: Sweden; TW: Taiwan; US: United States.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Genotypes, serotypes and pathotypes of the described infectious bronchitis virus (IBV) variants.
Table 1.
Genotypes, serotypes and pathotypes of the described infectious bronchitis virus (IBV) variants.
| IBV Strain | Genotype | Lineage * | Geographic Origin | Immunization History | Arose From | Serotype or Protectotype | Viral Distribution | Mortality | References |
|---|---|---|---|---|---|---|---|---|---|
| CK/CH/LGX/111119 | Novel | GI-28 | China | H120 and 4/91 vaccinated yellow broiler | LX4, LDL/091022 and an unknown strain | Novel, different from Mass, 4/91, LX4, LDL/97I, and TW-I | Kidney, bronchi, cecal tonsils, oviduct | 30% in one-day-old specific-pathogen-free (SPF) white leghorn chickens with 106 50% embryo infective dose (EID50) | [20] |
| CK/CH/2010/JT-1 | Novel | GI-28 | China | H120 and 4/91 vaccinated broiler | QX, LSC/ 99I, LDT3/03 and 4/91 | Novel, different from Mass and 4/91 | Tracheal, lung, kidney | 43.75% in three-day-old SPF white leghorn chickens with 105 EID50 | [21] |
| SAIBK2 | Grouped in the most predominant genotype in China with YN, Sczy3, etc. | GI-19 | China | H120 vaccinated layer chickens | YX10, YN, and Mass | N.D.** | Kidney | 50% in one-day-old SPF chickens with 105 EID50 | [22] |
| CK/CH/LDL/110931 CK/CH/LHB/130573 | Mass | GI-1 | China | H120 vaccinated | H120-like vaccine strain and other types of viruses | Novel, different from H120, Conn and LDT3 | N.D. | N.D. | [23] |
| YN | Novel | GI-22 | China | H120 vaccinated broiler | N.D. | N.D. | Tracheal, lung, kidney, bursa, ovary, oviduct | 65% in 30-day-old SPF chickens with 105 EID50 40.5% in 21-day-old commercial laying hens with 106.5 EID50 | [24,25] |
| GD | TW-like | GI-7 | China | Mass-type vaccinated broiler | QX-like (S2, E, M and N) and TW-like (S1) | QX | Tracheal, lung, kidney, bursa, spleen, proventriculus | 40% in 21-day-old SPF chickens with 106 EID50 | [26,27] |
| CK/CH/LDL/140520 | TW-1 | GI-7 | China | H120 vaccinated | LX4 and Taiwan group 1 (TW-1) | Different from Mass, LDT3, 4/91, and Conn | Tracheal, kidney, oviduct | 30% in one-day-old SPF chickens with 105.5 EID50 | [28] |
| CK/CH/LGX/130530 | LDT3 | Unknown | China | H120 and Ma5 vaccinated broiler | H120 (nsp1-14) and LDT3/03 (nsp14-N) | LDT3-A vaccine provide protection while H120 not | Tracheal, kidney | 0% in one-day-old SPF chickens with 105 EID50 | [29] |
| CK/CH/LJL/111054 | Conn | GI-1 | China | H120 vaccinated layer | Mass type and Conn type | N.D. | N.D. | 0% in one-day-old SPF chickens with 105.5 EID50 | [30] |
| CK/CH/IBTZ/2012 | N.D. | GVI-1 | China | H120 vaccinated layer | LX4 and an unknown strain in S gene | N.D. | N.D. | N.D. | [31] |
| Sczy3 | LX4 | GI-19 | China | Unknown | LX4 (major) and H120 (minor) | N.D. | N.D. | N.D. | [32] |
| SAIBK | Novel | GI-22 | China | Unknown | SC021202 (major) and H120 | 4/91-like | [33] | ||
| 3575/08 | TW-I | GI-7 | Taiwan | TW-I vaccinated broiler | TW-I strain with point mutations | Novel, distinct with Mass or other local types | Trachea, proventriculus, kidney, bursa, oviduct | 83.3% in one-day-old SPF chickens with 106 EID50 | [34] |
| JP/Wakayama/2003 JP/Iwate/2005 | 4/91 | GI-13 | Japan | Without 4/91 immunization | N.D. | 4/91, JP/Iwate/2005 can be protected by 4/91 and JP-II strain. | Respiratory tract | 15.4% in one-day-old SPF chickens with 104 EID50 of JP/Wakayama/2003 | [35] |
| SNU8067 | Korean group I | GI-15 | South Korea | Unknown | Korean group II strain KM91 (except S1, 3a,3b) and Korean group I (S1) | KM91 and M41 inactivate vaccine protect 70% from oviduct lesion | Oviduct | N.D. | [36] |
| IBV/MN IBV/RA IBV/TU | Italy 02 | Unknown | Morocco | Unknown | N.D. | N.D. | Respiratory sign without renal lesion | 0% in one-day-old SPF chickens with 103.5 EID50 | [37] |
| IBV-EG/1212B-2012 IBV-EG/IBV1-2011 | Egy/Var II Egy/Var I | Unknown | Egypt | Unknown | Egyptian variant strains or Middle East strains with Israeli strains | N.D. | Tracheal, kidney | 50% to 40% in one-day-old SPF chickens with 105 EID50 | [38,39] |
| IBV/Cal99 variant/07 | Respiratory pathogenic Cal99 genotype | GI-9 | US | Game chickens without IBV vaccination | N.D. | N.D. | Trachea, lung, kidney, salivary glands, small intestine | N.D. | [40] |
| IBV/Brazil/2005/USP-10 | Brazilian type | Unknown | Brazil | Mass type vaccinated broiler | N.D. | N.D. | Respiratory tissue, kidney, intestine and testis | None in 26-day-old SPF chickens and male Ross broilers | [41] |
| γCoV/CK/Italy/I2022/13 IBV/FARM 1 IBV/FARM 5 | Q1 | GI-16 | Italy | H120 vaccinated broiler | Q1 and unknown minor parent strains | Q1 type | Trachea, lungs, kidneys, proventriculus | N.D., Ranging from 4.1% to 9.8% in broiler outbreak | [42,43] |
| KG3P | QX-like | GI-19 | England | 793B vaccinated Broiler | N.D. | N.D. | Trachea, lungs, kidneys, proventriculus | 16.7% in SPF chickens with 104 TOC50 | [44] |
| N1/08 | Australian group 2 | GIII-1 | Australia | Unknown | Australian group 2 and group 3 strains in S | N.D. | Trachea, cecal tonsils | None in 14-day-old SPF chickens with 105.5 EID50 | [45,46] |
* Classification based on the S1 gene, according to Valastro et al. [18]; ** N.D.: Not done.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lin, S.-Y.; Chen, H.-W. Infectious Bronchitis Virus Variants: Molecular Analysis and Pathogenicity Investigation. Int. J. Mol. Sci. 2017, 18, 2030. https://doi.org/10.3390/ijms18102030
AMA Style
Lin S-Y, Chen H-W. Infectious Bronchitis Virus Variants: Molecular Analysis and Pathogenicity Investigation. International Journal of Molecular Sciences. 2017; 18(10):2030. https://doi.org/10.3390/ijms18102030
Chicago/Turabian StyleLin, Shu-Yi, and Hui-Wen Chen. 2017. "Infectious Bronchitis Virus Variants: Molecular Analysis and Pathogenicity Investigation" International Journal of Molecular Sciences 18, no. 10: 2030. https://doi.org/10.3390/ijms18102030
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.