
Soy-Leaf Extract Exerts Atheroprotective Effects via Modulation of Krüppel-Like Factor 2 and Adhesion Molecules

 ,
,
Abstract
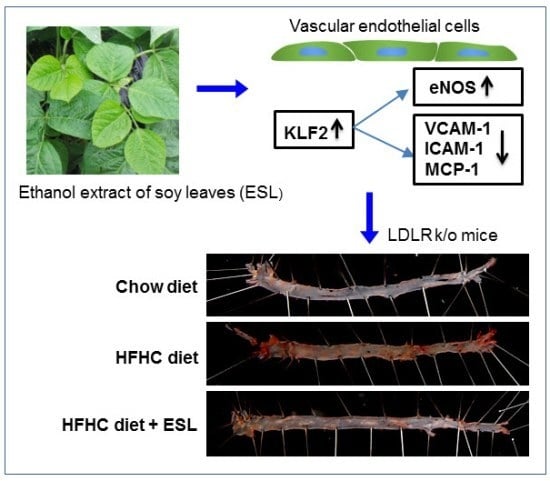
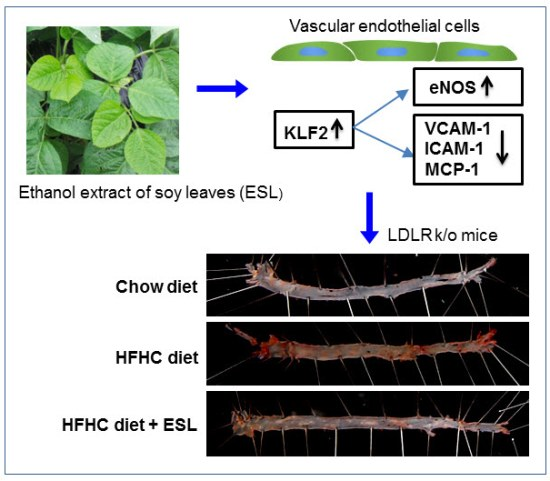
:
1. Introduction
2. Results and Discussion
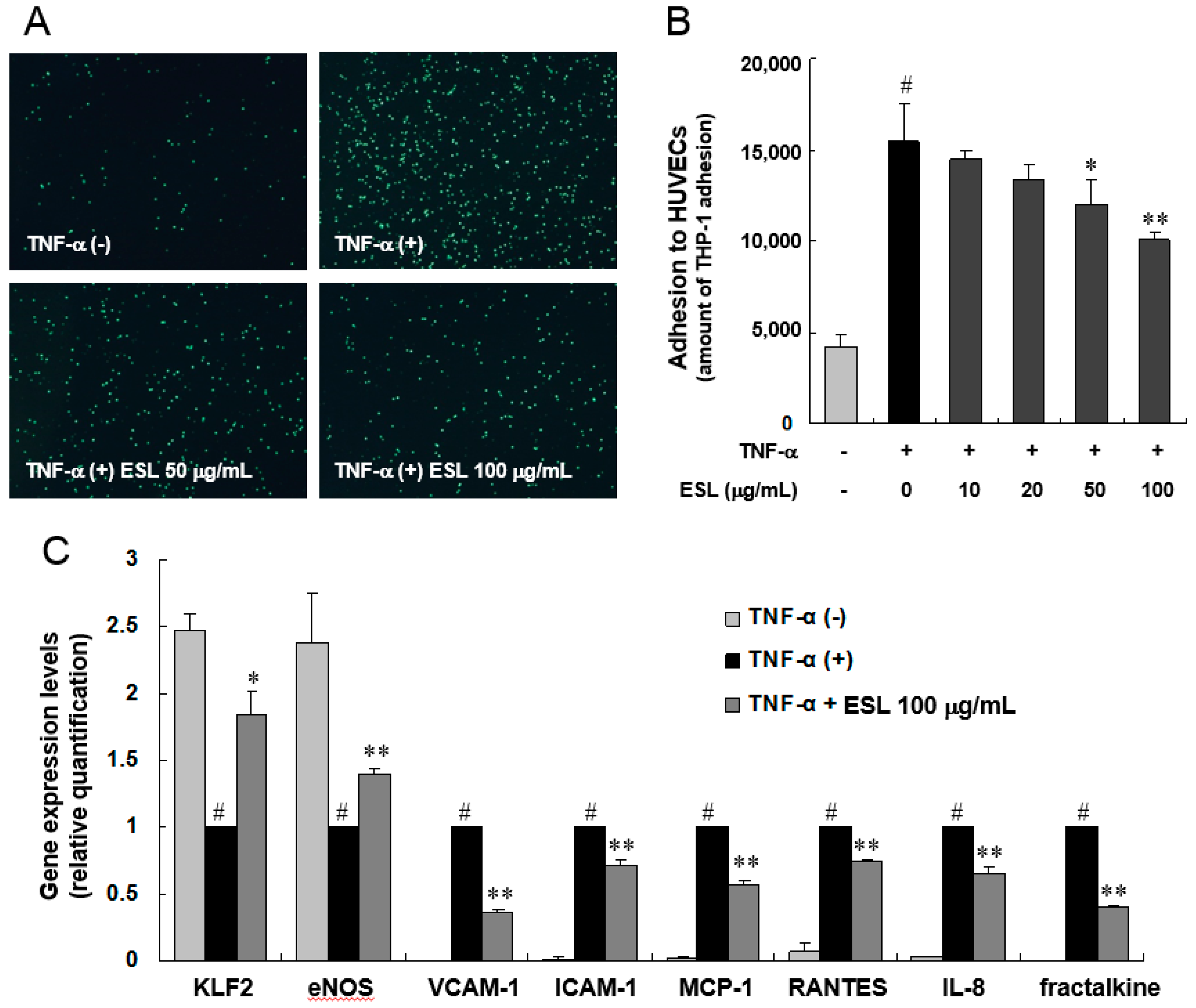
2.1. ESL Inhibited the Adhesion of Monocytes to Activated HUVECs and Expression of Adhesion Molecules
2.2. ESL Antagonized TNF-α-Induced Suppression of KLF2 and eNOS Expression
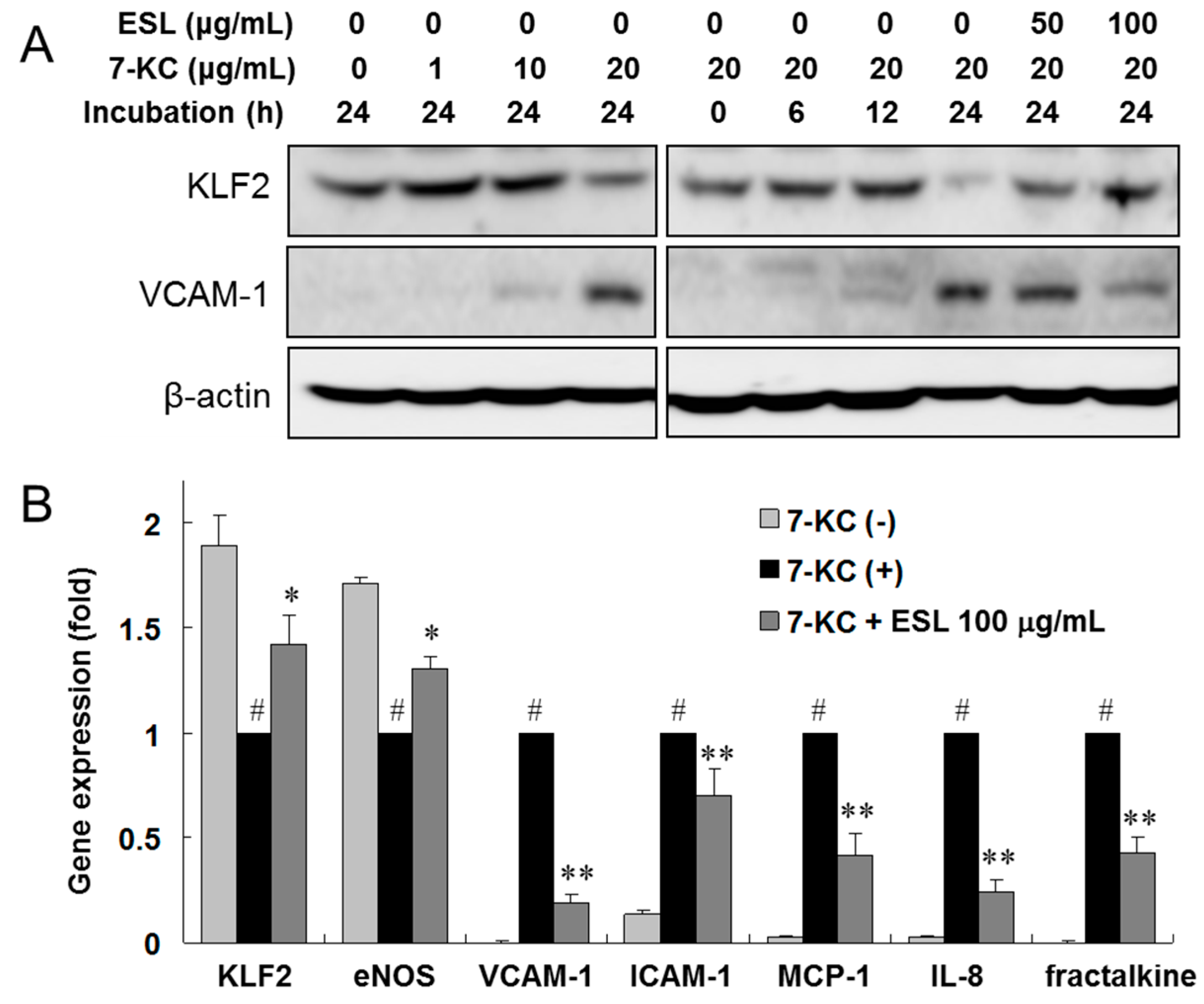
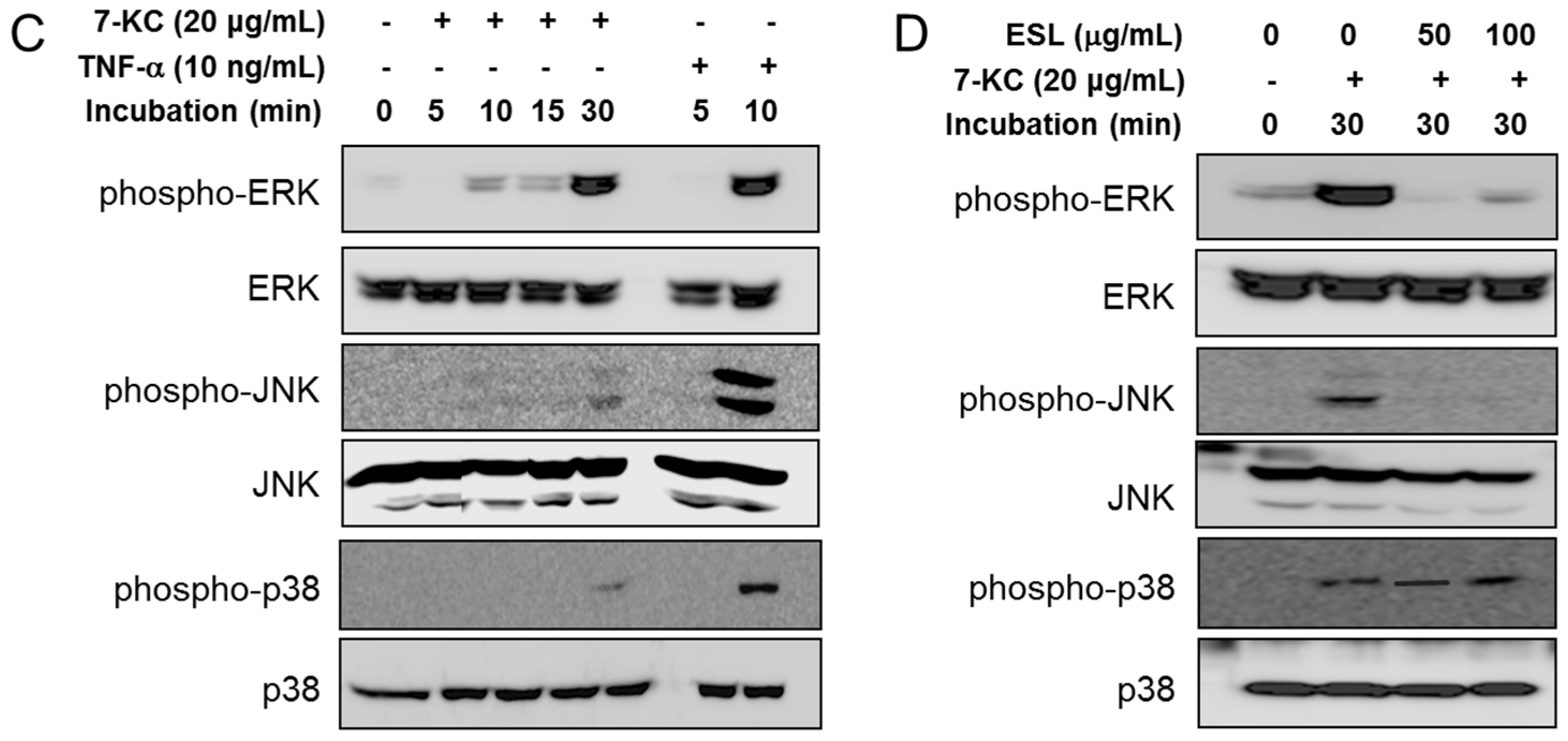
2.3. ESL Antagonized 7-KC-Induced Suppression of KLF2 and eNOS Expression
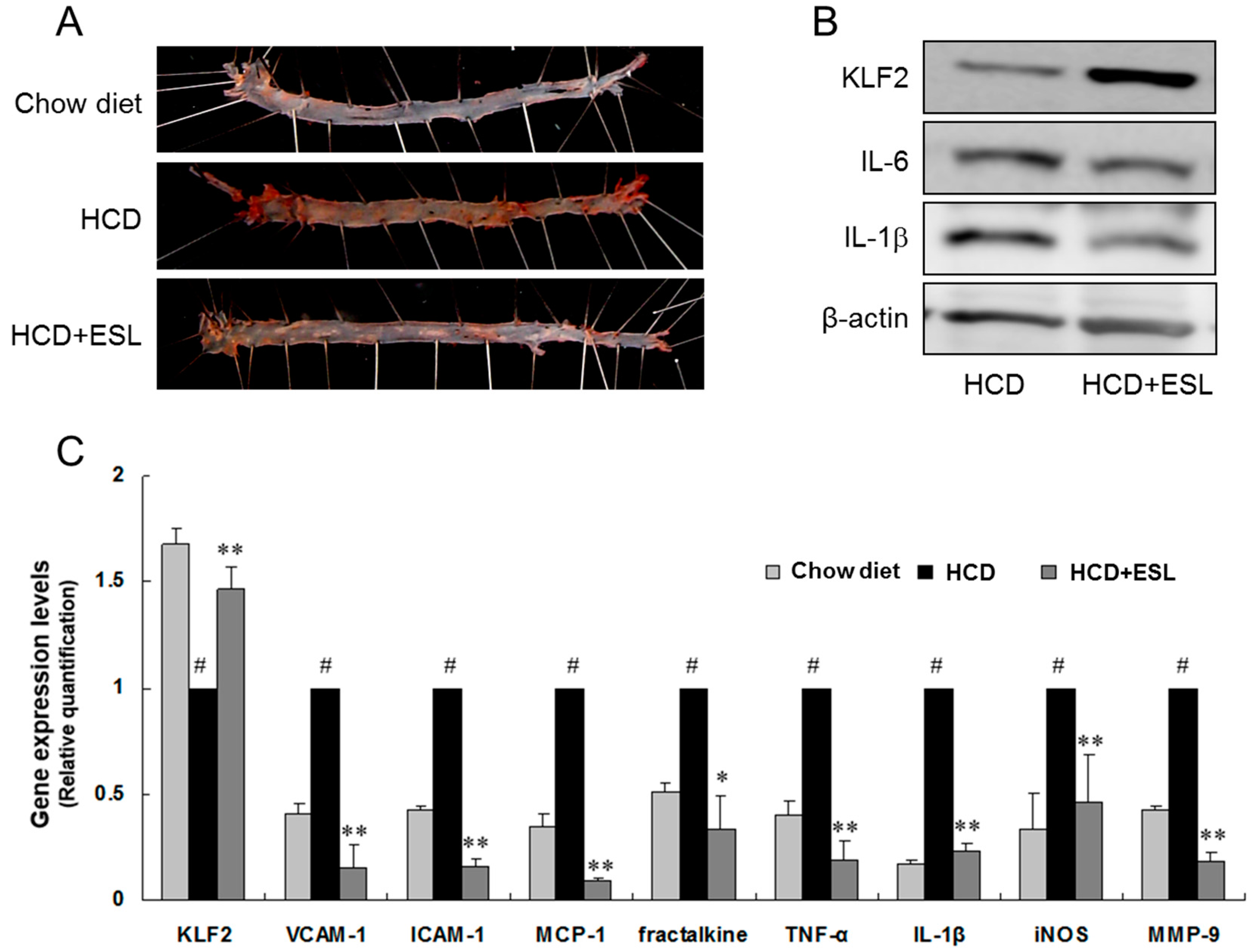
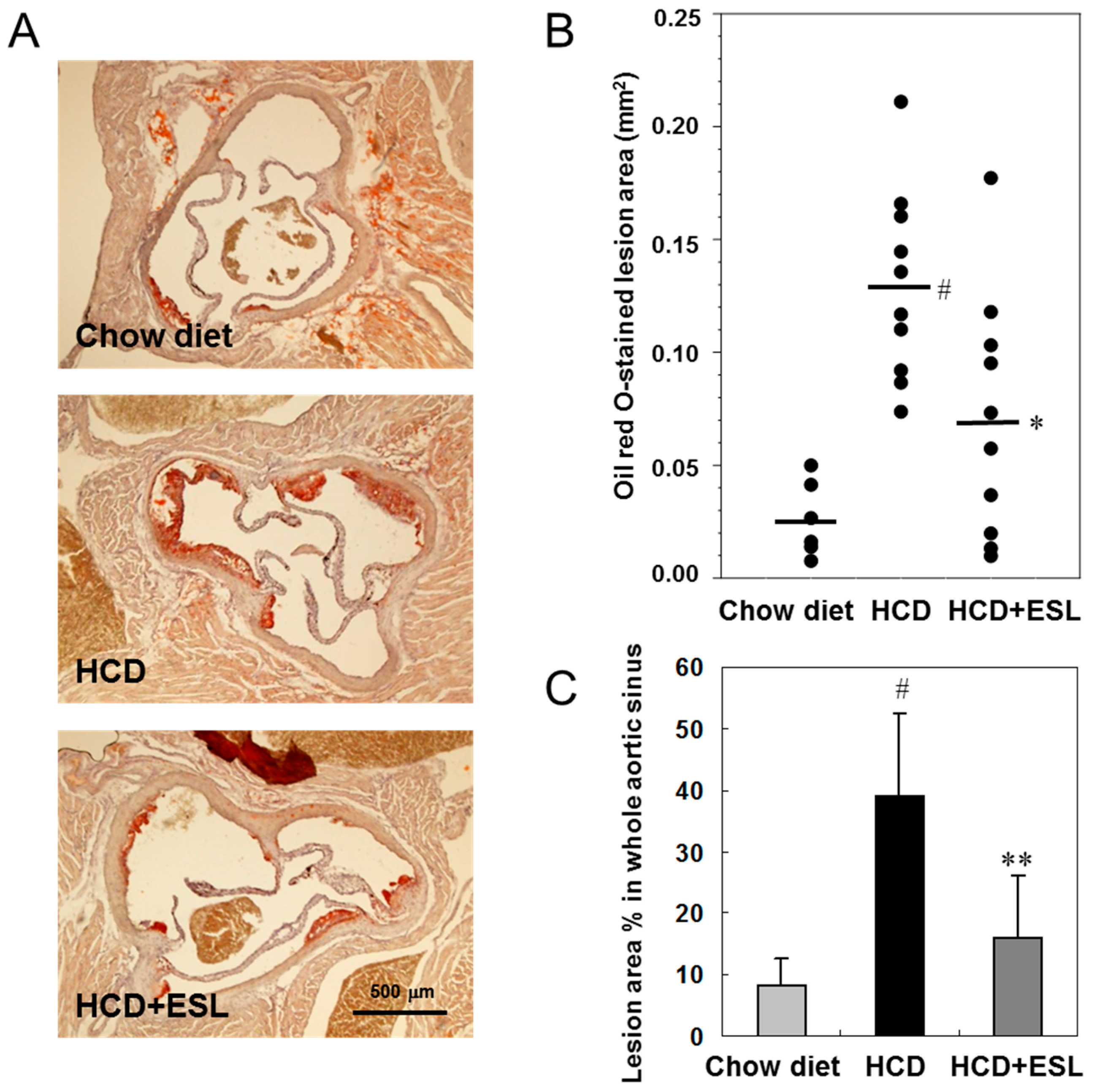
2.4. ESL Reduced the Development of Atherosclerosis in LDLR−/− Mice
3. Materials and Methods
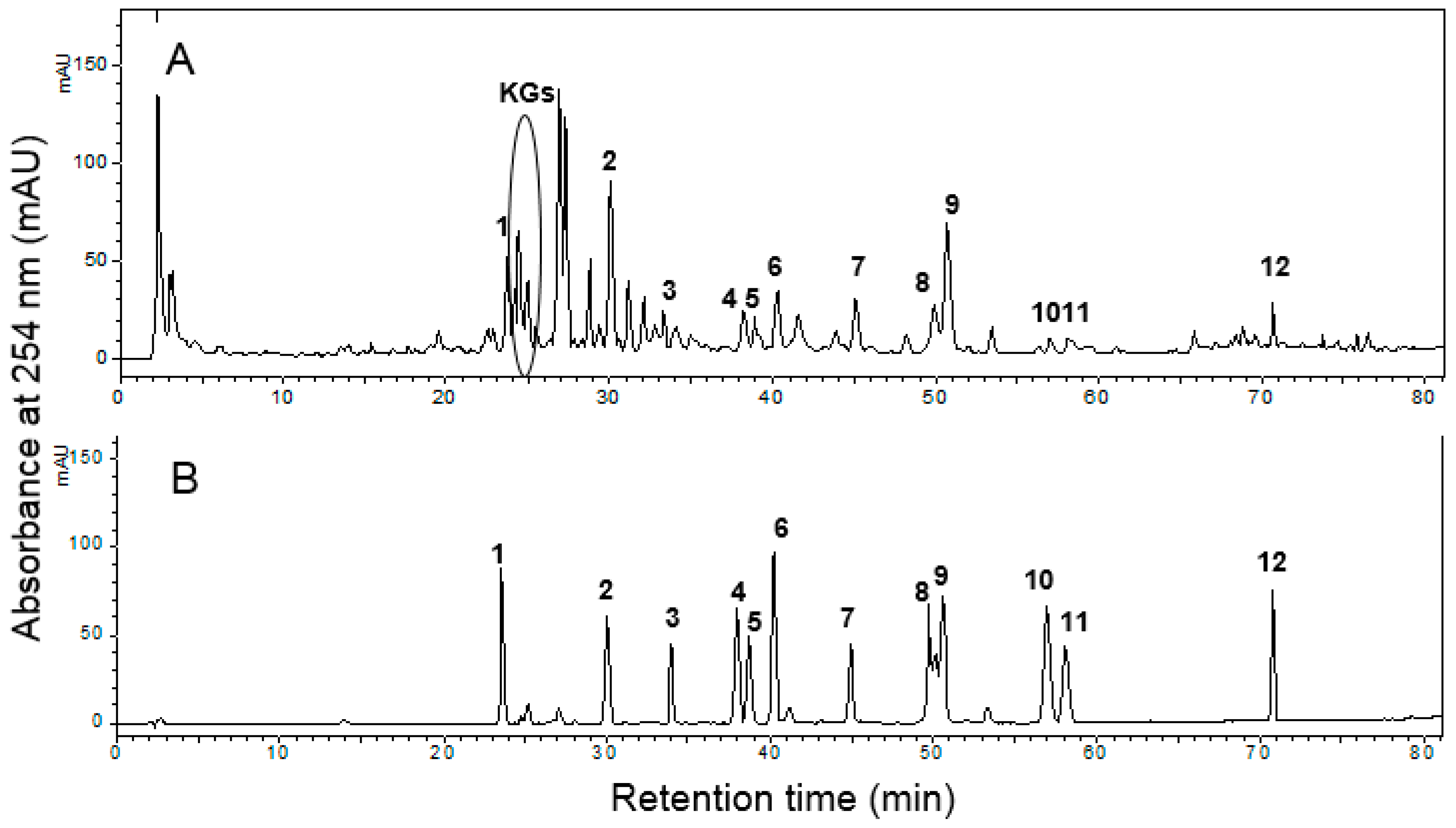
3.1. Preparation of ESL and HPLC Analysis
3.2. Materials and Cell Culture
3.3. Animal Experiments
3.4. Measurement of Fatty Streak Lesion Area
3.5. Cell Adhesion Assay
3.6. Immunofluorescence Analysis
3.7. Real-Time qRT-PCR Analysis
3.8. Western Blot Analysis
3.9. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hansson, G.K. Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, M.W.; Lin, Z.; Fisch, S.; Jain, M.K. An emerging role for Krűppel-like factors in vascular biology. Trends Cardiovasc. Med. 2004, 14, 241–246. [Google Scholar] [CrossRef] [PubMed]
- SenBanerjee, S.; Lin, Z.; Atkins, G.B.; Greif, D.M.; Rao, R.M.; Kumar, A.; Feinberg, M.W.; Chen, Z.; Simon, D.I.; Luscinskas, F.W.; et al. KLF2 is a novel transcriptional regulator of endothelial proinflammatory activation. J. Exp. Med. 2004, 199, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.B.; Jain, M.K. Role of Krűppel-like transcription factors in endothelial biology. Circ. Res. 2007, 100, 1686–1695. [Google Scholar] [CrossRef] [PubMed]
- Das, H.; Kumar, A.; Lin, Z.; Patino, W.D.; Hwang, P.M.; Feinberg, M.W.; Majumder, P.K.; Jain, M.K. Kruppel-like factor 2 (KLF2) regulates proinflammatory activation of monocytes. Proc. Natl. Acad. Sci. USA 2006, 103, 6653–6658. [Google Scholar] [CrossRef] [PubMed]
- Parmar, K.M.; Nambudiri, V.; Dai, G.; Larman, H.B.; Gimbrone, M.A., Jr.; García-Cardeña, G. Statins exert endothelial atheroprotective effects via the KLF2 transcription factor. J. Biol. Chem. 2005, 280, 26714–26719. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.K.; Ridker, P.M. Anti-inflammatory effects of statins: Clinical evidence and basic mechanisms. Nat. Rev. Drug Discov. 2005, 4, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Tuomisto, T.T.; Lumivuori, H.; Kansanen, E.; Häkkinen, S.K.; Turunen, M.P.; van Thienen, J.V.; Horrevoets, A.J.; Levonen, A.L.; Ylä-Herttuala, S. Simvastatin has an anti-inflammatory effect on macrophages via upregulation of an atheroprotective transcription factor, Kruppel-like factor 2. Cardiovasc. Res. 2008, 78, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, M.; Rabkin, E.; Sugiyama, S.; Voglic, S.J.; Fukumoto, Y.; Furukawa, Y.; Shiomi, M.; Schoen, F.J.; Libby, P. An HMG-CoA reductase inhibitor, cerivastatin, suppresses growth of macrophages expressing matrix metalloproteinases and tissue factor in vivo and in vitro. Circulation 2001, 103, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Dekker, R.J.; van Soest, S.; Fontijn, R.D.; Salamanca, S.; de Groot, P.G.; VanBavel, E.; Pannekoek, H.; Horrevoets, A.J. Prolonged fluid shear stress induces a distinct set of endothelial cell genes, most specifically lung Kruppel-like factor (KLF2). Blood 2002, 100, 1689–1698. [Google Scholar] [CrossRef] [PubMed]
- Atkins, G.B.; Wang, Y.; Mahabeleshwar, G.H.; Shi, H.; Gao, H.; Kawanami, D.; Natesan, V.; Lin, Z.; Simon, D.I.; Jain, M.K. Hemizygous deficiency of Krűppel-like factor 2 augments experimental atherosclerosis. Circ. Res. 2008, 103, 690–693. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Jessup, W. Oxysterols and atherosclerosis. Atherosclerosis 1999, 142, 1–28. [Google Scholar] [CrossRef]
- Merz-Demlow, B.E.; Duncan, A.M.; Wangen, K.E.; Xu, X.; Merz-Demlow, B.E.; Duncan, A.M.; Wangen, K.E.; Xu, X.; Carr, T.P.; Phipps, W.R.; et al. Soy isoflavones improve plasma lipids in normocholesterolemic, premenopausal women. Am. J. Clin. Nutr. 2000, 71, 1462–1469. [Google Scholar] [PubMed]
- Nagarajan, S. Mechanisms of anti-atherosclerotic functions of soy-based diets. J. Nutr. Biochem. 2010, 21, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Rostagno, M.A.; Palma, M.; Barroso, C.G. Ultrasound-assisted extraction of soy isoflavones. J. Chromatogr. A 2003, 1012, 119–128. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, M.; Sciarappa, W.J.; Simon, J.E. LC/UV/ESI-MS analysis of isoflavones in Edamame and Tofu soybeans. J. Agric. Food Chem. 2004, 52, 2763–2769. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.M.; Chen, R.Y.; Leung, L.K.; Chan, F.L.; Huang, Y.; Chen, Z.Y. Difference in flavonoid and isoflavone profile between soybean and soy leaf. Biomed. Pharmacother. 2002, 56, 289–295. [Google Scholar] [CrossRef]
- Li, H.; Ji, H.S.; Kang, J.H.; Shin, D.H.; Park, H.Y.; Choi, M.S.; Lee, C.H.; Lee, I.K.; Yun, B.S.; Jeong, T.S. Soy leaf containing kaempferol glycosides and pheophorbides improves glucose homeostasis by enhancing pancreatic β-cell function and suppressing hepatic lipid accumulation in db/db mice. J. Agric. Food Chem. 2015, 63, 7198–7210. [Google Scholar] [CrossRef] [PubMed]
- Yuk, H.J.; Lee, J.H.; Curtis-Long, M.J.; Lee, J.W.; Kim, Y.S.; Ryu, H.W.; Park, C.G.; Jeong, T.S.; Park, K.H. The most abundant polyphenol of soy leaves, coumestrol, displays potent α-glucosidase inhibitory activity. Food Chem. 2011, 126, 1057–1063. [Google Scholar] [CrossRef]
- Ho, H.M.; Chen, R.; Huang, Y.; Chen, Z.Y. Vascular effects of a soy leaves (Glycine max) extract and kaempferol glycosides in isolated rat carotid arteries. Planta Med. 2002, 68, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.M.; Leung, L.K.; Chan, F.L.; Huang, Y.; Chen, Z.Y. Soy leaf lowers the ratio of non-HDL to HDL cholesterol in hamsters. J. Agric. Food Chem. 2003, 51, 4554–4558. [Google Scholar] [CrossRef] [PubMed]
- Kim, U.H.; Yoon, J.H.; Li, H.; Kang, J.H.; Ji, H.S.; Park, K.H.; Shin, D.H.; Park, H.Y.; Jeong, T.S. Pterocarpan-enriched soy leaf extract ameliorates insulin sensitivity and pancreatic β-cell proliferation in type 2 diabetic mice. Molecules 2014, 19, 18493–18510. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Jeon, S.M.; Park, K.H.; Lee, W.S.; Jeong, T.S.; McGregor, R.A.; Choi, M.S. Does Glycine max leaves or Garcinia cambogia promote weight-loss or lower cholesterol in overweight individuals: A randomized control trial. Nutr. J. 2011, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.S.; Ryu, R.; Seo, Y.R.; Jeong, T.S.; Shin, D.H.; Park, Y.B.; Kim, S.R.; Jung, U.J. The beneficial effect of soybean (Glycine max (L.) Merr.) leaf extracts in adults with prediabetes: A randomized placebo controlled trial. Food Funct. 2014, 5, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Tsuda, Y.; Sakono, M.; Imaizumi, K. Dietary soy protein isolate, compared with casein, reduces atherosclerotic lesion area in apolipoprotein E-deficient mice. J. Nutr. 1998, 128, 1884–1889. [Google Scholar] [PubMed]
- Potter, S.M. Overview of proposed mechanisms for the hypocholesterolemic effect of soy. J. Nutr. 1995, 125, 606S–611S. [Google Scholar] [PubMed]
- Majewska, E.; Paleolog, E.; Baj, Z.; Kralisz, U.; Feldmann, M.; Tchórzewski, H. Role of tyrosine kinase enzymes in TNF-α and IL-1 induced expression of ICAM-1 and VCAM-1 on human umbilical vein endothelial cells. Scand. J. Immunol. 1997, 45, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Bu, D.X.; Tarrio, M.; Grabie, N.; Zhang, Y.; Yamazaki, H.; Stavrakis, G.; Maganto-Garcia, E.; Pepper-Cunningham, Z.; Jarolim, P.; Aikawa, M.; et al. Statin-induced kruppel-like factor 2 expression in human and mouse T cells reduces inflammatory and pathogenic responses. J. Clin. Investig. 2010, 120, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Sen-Banerjee, S.; Mir, S.; Lin, Z.; Hamik, A.; Atkins, G.B.; Das, H.; Banerjee, P.; Kumar, A.; Jain, M.K. Kruppel-like factor 2 as a novel mediator of statin effects in endothelial cells. Circulation 2005, 112, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Shimozawa, M.; Naito, Y.; Manabe, H.; Uchiyama, K.; Kuroda, M.; Katada, K.; Yoshida, N.; Yoshikawa, T. 7-Ketocholesterol enhances the expression of adhesion molecules on human aortic endothelial cells by increasing the production of reactive oxygen species. Redox Rep. 2004, 9, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, B.W.; Kim, J.H.; Jeong, T.S.; Kim, M.J.; Lee, W.S.; Park, K.H. LDL-antioxidant pterocarpans from roots of Glycine max (L.) Merr. J. Agric. Food Chem. 2006, 54, 2057–2063. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Seo, W.D.; Jeong, S.H.; Jeong, T.S.; Lww, W.S.; Park, K.H. Human acyl-CoA: Cholesterol acyltransferase inhibitory effect of flavonoids from roots of Glycine max (L.) Merr. Agric. Chem. Biotechnol. 2006, 49, 57–61. [Google Scholar]
- Xiao, H.B.; Jun, F.; Lu, X.Y.; Chen, X.J.; Chao, T.; Sun, Z.L. Protective effects of kaempferol against endothelial damage by an improvement in nitric oxide production and a decrease in asymmetric dimethylarginine level. Eur. J. Pharmacol. 2009, 616, 213–222. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Number) | Primer Sequence (Forward 5′–3′) | Primer Sequence (Reverse 5′–3’) |
|---|---|---|
| hCCL5 (NM_002985) | CGGGAGTACATCAACTCTTTGGA | CAAGCTAGGACAAGAGCAAGCA |
| heNOS (NM_000603) | CTCATGGGCACGGTGATG | ACCACGTCATACTCATCCATACAC |
| hFractalkine (NM_002996) | CGCAGCATATTCAGGAAGCT | TCCTTGTCCATGTCCTGCTT |
| hICAM-1 (NM_000201) | AGAGGTTGAACCCCACAGTC | TCTGGCTTCGTCAGAATCAC |
| hIL-8 (NM_000584) | CCTAGATATTGCACGGGAGA | GTGGAACAAGGACTTGTGGA |
| hKLF2 (NM_016270) | CCTCCTTGACGAGTTTTGTTTTTC | AAGGCATCACAAGCCTCGAT |
| hMCP-1 (NM_002982) | GCTCAGCCAGATGCAATCAA | CTTGGCCACAATGGTCTTGA |
| hVCAM-1 (NM_001078) | GTTGAAGGATGCGGGAGTAT | TTCATGTTGGCTTTTCTTGC |
| hActin (NM_001101) | GGCACCACACCTTCTACAAT | TCTGGGCATCCTTCACAGCT |
| mFractalkine (NM_009142) | TCACGTGCAGCAAGATGACA | TCCTTGACCCATTGCTCCTT |
| mICAM-1 (NM_010493) | GTGATGCTCAGGTATCCATC | GAATACACGGTGATGGTAGC |
| mIL-1β (NM_008361) | ATGAGGACATGAGCACCTTC | CATTGAGGTGGAGAGCTTTC |
| miNOS (NM_010927) | GGCAGCCTGTGAGACCTTTG | TGCATTGGAAGTGAAGCGTTT |
| mKLF2 (NM_008452) | CGCCACACATACTTGCAGCTA | GTGTACGCAGATGCGCCTTT |
| mMCP-1 (NM_011333) | TGCTGACCCCAAGAAGGAAT | TGCTTGAGGTGGTTGTGGAA |
| mMMP-9 (NM_013599) | CAGCCAACTATGACCAGGAT | TCGCTGGTACAGGAAGAGTA |
| mTNF-α (NM_013693) | CTCAGATCATCTTCTCAAAATTCGAGTGACA | CTTCACAGAGCAATGACTCCAAAGT |
| mVCAM-1 (NM_011693) | CTGTTTGCAGTCTCTCAAGC | CCAAACACTTGACTGTGACC |
| mGAPDH (NM_008084) | ACATCATCCCTGCATCCACT | AGATCCACGACGGACACATT |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.-M.; Li, H.; Cho, M.-H.; Baek, S.-H.; Lee, C.-H.; Park, H.-Y.; Jeong, T.-S. Soy-Leaf Extract Exerts Atheroprotective Effects via Modulation of Krüppel-Like Factor 2 and Adhesion Molecules. Int. J. Mol. Sci. 2017, 18, 373. https://doi.org/10.3390/ijms18020373
Han J-M, Li H, Cho M-H, Baek S-H, Lee C-H, Park H-Y, Jeong T-S. Soy-Leaf Extract Exerts Atheroprotective Effects via Modulation of Krüppel-Like Factor 2 and Adhesion Molecules. International Journal of Molecular Sciences. 2017; 18(2):373. https://doi.org/10.3390/ijms18020373
Chicago/Turabian StyleHan, Jong-Min, Hua Li, Moon-Hee Cho, Seung-Hwa Baek, Chul-Ho Lee, Ho-Yong Park, and Tae-Sook Jeong. 2017. "Soy-Leaf Extract Exerts Atheroprotective Effects via Modulation of Krüppel-Like Factor 2 and Adhesion Molecules" International Journal of Molecular Sciences 18, no. 2: 373. https://doi.org/10.3390/ijms18020373