
Plant Resistance Inducers against Pathogens in Solanaceae Species—From Molecular Mechanisms to Field Application


Abstract
:
1. Introduction and Background
2. Plant Resistance Inducers (PRIs) in Solanaceae Species
2.1. Non-Biological PRIs
2.1.1. 2,6-Dichloro isonicotinic acid (INA), Benzothiadiazole (BTH), and 3-Chloro-1-methyl-1H-pyrazole-5-carboxylic acid (CMPA)
2.1.2. β-Aminobutyric Acid (BABA)
2.1.3. Inorganic Salts
2.2. Bio-Based PRIs
2.2.1. Intrinsic Compounds Used as PRIs
2.2.2. Bacterial Elicitors
2.2.3. Fungal Elicitors
2.2.4. Plant Extracts (Botanicals)
3. Induced Resistance Mechanisms in Solanaceae
3.1. Genes Underlying Induced Resistance
3.2. “-Omics” Studies on the Effect of PRIs in Solanaceae
4. Transgenerational Effects of Induced Resistance
5. Induced Resistance in Field and Practice
6. PRIs Tested in Solanaceous Species in Field Conditions
7. Conclusions and Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Beauverie, J. Essais d’immunisation des végétaux contre les maladies cryptogamiques. CR Acad. Sci. Ser. III 1901, 133, 107–110. [Google Scholar]
- Walters, D.R.; Fountaine, J.M. Practical Application of Induced Resistance to Plant Diseases: An Appraisal of Effectiveness under Field Conditions; Cambridge University Press: Cambridge, UK, 2009; Volume 147, p. 13. [Google Scholar]
- Walters, D.R.; Ratsep, J.; Havis, N.D. Controlling crop diseases using induced resistance: Challenges for the future. J. Exp. Bot. 2013, 64, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Gozzo, F.; Faoro, F. Systemic acquired resistance (50 years after discovery): Moving from the lab to the field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed]
- White, R. Acetylsalicylic acid (aspirin) induces resistance to tobacco mosaic virus in tobacco. Virology 1979, 99, 410–412. [Google Scholar] [CrossRef]
- Walters, D.; Newton, A.C.; Lyon, G. Induced Resistance for Plant Defence: A Sustainable Approach to Crop Protection; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Walters, D.; Heil, M. Costs and trade-offs associated with induced resistance. Physiol. Mol. Plant Pathol. 2007, 71, 3–17. [Google Scholar] [CrossRef]
- Van Hulten, M.; Pelser, M.; van Loon, L.; Pieterse, C.M.; Ton, J. Costs and benefits of priming for defense in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 5602–5607. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U.; Pieterse, C.M.; Mauch-Mani, B. Priming in plant-pathogen interactions. Trends Plant Sci. 2002, 7, 210–216. [Google Scholar] [CrossRef]
- Bengtsson, T.; Weighill, D.; Proux-Wéra, E.; Levander, F.; Resjö, S.; Burra, D.D.; Moushib, L.I.; Hedley, P.E.; Liljeroth, E.; Jacobson, D. Proteomics and transcriptomics of the BABA-induced resistance response in potato using a novel functional annotation approach. BMC Genom. 2014, 15, 315. [Google Scholar] [CrossRef] [PubMed]
- Camañes, G.; Scalschi, L.; Vicedo, B.; González-Bosch, C.; García-Agustín, P. An untargeted global metabolomic analysis reveals the biochemical changes underlying basal resistance and priming in Solanum lycopersicum, and identifies 1-methyltryptophan as a metabolite involved in plant responses to Botrytis cinerea and Pseudomonas syringae. Plant J. 2015, 84, 125–139. [Google Scholar] [PubMed]
- Balmer, A.; Pastor, V.; Gamir, J.; Flors, V.; Mauch-Mani, B. The ”prime-ome“: Towards a holistic approach to priming. Trends Plant Sci. 2015, 20, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Holeski, L.M.; Jander, G.; Agrawal, A.A. Transgenerational defense induction and epigenetic inheritance in plants. Trends Ecol. Evol. 2012, 27, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Pastor, V.; Luna, E.; Mauch-Mani, B.; Ton, J.; Flors, V. Primed plants do not forget. Environ. Exp. Bot. 2013, 94, 46–56. [Google Scholar] [CrossRef]
- Fry, W. Phytophthora infestans: The plant (and R gene) destroyer. Mol. Plant Pathol. 2008, 9, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Davidse, L.; Looijen, D.; Turkensteen, L.; van der Wal, D. Occurrence of metalaxyl-resistant ttrains of Phytophthora infestans in Dutch potato fields. Eur. J. Plant Pathol. 1981, 87, 65–68. [Google Scholar] [CrossRef]
- Parra, G.; Ristaino, J.B. Resistance to mefenoxam and metalaxyl among field isolates of Phytophthora capsici causing Phytophthora blight of bell pepper. Plant Dis. 2001, 85, 1069–1075. [Google Scholar] [CrossRef]
- Kromann, P.; Pérez, W.G.; Taipe, A.; Schulte-Geldermann, E.; Sharma, B.P.; Andrade-Piedra, J.L.; Forbes, G.A. Use of phosphonate to manage foliar potato late blight in developing countries. Plant Dis. 2012, 96, 1008–1015. [Google Scholar] [CrossRef]
- Oostendorp, M.; Kunz, W.; Dietrich, B.; Staub, T. Induced disease resistance in plants by chemicals. Eur. J. Plant Pathol. 2001, 107, 19–28. [Google Scholar] [CrossRef]
- Bektas, Y.; Eulgem, T. Synthetic plant defense elicitors. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Abreha, K.B.; Alexandersson, E.; Vossen, J.H.; Anderson, P.; Andreasson, E. Inoculation of transgenic resistant potato by Phytophthora infestans affects host plant choice of a generalist moth. PLoS ONE 2015, 10, e0129815. [Google Scholar] [CrossRef] [PubMed]
- Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar]
- Consortium, T.G. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [Green Version]
- Mongae, A.; Moleleki, L. The effect of β-aminobutyric acid (BABA) on root knot nematode and soft rot pathogen disease complexes in Solanum tuberosum plants. Eur. J. Plant Pathol. 2015, 142, 117–124. [Google Scholar] [CrossRef]
- Sós-Hegedűs, A.; Juhász, Z.; Poór, P.; Kondrák, M.; Antal, F.; Tari, I.; Mauch-Mani, B.; Bánfalvi, Z. Soil drench treatment with β-aminobutyric acid increases drought tolerance of potato. PLoS ONE 2014, 9, e114297. [Google Scholar]
- Hong, J.K.; Hwang, B.K.; Kim, C.H. Induction of local and systemic resistance to Colletotrichum coccodes in pepper plants by dl-β-Amino-n-Butyric Acid. J. Phytopathol. 1999, 147, 193–198. [Google Scholar] [CrossRef]
- Lee, Y.K.; Hong, J.K.; Hippe-Sanwald, S.; Hwang, B.K. Histological and ultrastructural comparisons of compatible, incompatible and dl-β-amino-n-butyric acid-induced resistance responses of pepper stems to Phytophthora capsici. Physiol. Mol. Plant Pathol. 2000, 57, 269–280. [Google Scholar] [CrossRef]
- Sunwoo, J.Y.; Lee, Y.K.; Hwang, B.K. Induced resistance against Phytophthora capsici in pepper plants in response to dl-β-amino-n-butyric acid. Eur. J. Plant Pathol. 1996, 102, 663–670. [Google Scholar] [CrossRef]
- Reuveni, R.; Dor, G.; Reuveni, M. Local and systemic control of powdery mildew (Leveillula taurica) on pepper plants by foliar spray of mono-potassium phosphate. Crop Prot. 1998, 17, 703–709. [Google Scholar] [CrossRef]
- Förster, H.; Adaskaveg, J.; Kim, D.; Stanghellini, M. Effect of phosphite on tomato and pepper plants and on susceptibility of pepper to Phytophthora root and crown rot in hydroponic culture. Plant Dis. 1998, 82, 1165–1170. [Google Scholar] [CrossRef]
- Choi, H.K.; Song, G.C.; Yi, H.-S.; Ryu, C.-M. Field evaluation of the bacterial volatile derivative 3-pentanol in priming for induced resistance in pepper. J. Chem. Ecol. 2014, 40, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Elad, Y.; David, D.R.; Harel, Y.M.; Borenshtein, M.; Kalifa, H.B.; Silber, A.; Graber, E.R. Induction of systemic resistance in plants by biochar, a soil-applied carbon sequestering agent. Phytopathology 2010, 100, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Buchenauer, H. Enhanced control of bacterial wilt of tomato by dl-3-aminobutyric acid and the fluorescent Pseudomonas isolate CW2. J. Plant Dis. Prot. 2008, 115, 199–207. [Google Scholar] [CrossRef]
- Baysal, Ö.; Gürsoy, Y.Z.; Örnek, H.; Duru, A. Induction of oxidants in tomato leaves treated with dl-β-Amino butyric acid (BABA) and infected with Clavibacter michiganensis ssp. michiganensis. Eur. J. Plant Pathol. 2005, 112, 361–369. [Google Scholar] [CrossRef]
- Thaler, J.S.; Fidantsef, A.L.; Duffey, S.S.; Bostock, R.M. Trade-offs in plant defense against pathogens and herbivores: A field demonstration of chemical elicitors of induced resistance. J. Chem. Ecol. 1999, 25, 1597–1609. [Google Scholar] [CrossRef]
- Hassan, M.A.; Abo-Elyousr, K.A. Activation of tomato plant defence responses against bacterial wilt caused by Ralstonia solanacearum using dl-3-aminobutyric acid (BABA). Eur. J. Plant Pathol. 2013, 136, 145–157. [Google Scholar] [CrossRef]
- Worrall, D.; Holroyd, G.H.; Moore, J.P.; Glowacz, M.; Croft, P.; Taylor, J.E.; Paul, N.D.; Roberts, M.R. Treating seeds with activators of plant defence generates long-lasting priming of resistance to pests and pathogens. New Phytol. 2012, 193, 770–778. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y. Local and systemic control of Phytophthora infestans in tomato plants by dl-3-amino-n-butanoic acids. Phytopathology 1994, 84, 55–59. [Google Scholar] [CrossRef]
- Mehari, Z.H.; Elad, Y.; Rav-David, D.; Graber, E.R.; Harel, Y.M. Induced systemic resistance in tomato (Solanum lycopersicum) against Botrytis cinerea by biochar amendment involves jasmonic acid signaling. Plant Soil 2015, 395, 31–44. [Google Scholar] [CrossRef]
- Mejía-Teniente, L.; Durán-Flores, F.D.D.; Chapa-Oliver, A.M.; Torres-Pacheco, I.; Cruz-Hernández, A.; González-Chavira, M.M.; Ocampo-Velázquez, R.V.; Guevara-González, R.G. Oxidative and molecular responses in Capsicum annuum L. after hydrogen peroxide, salicylic acid and chitosan foliar applications. Int. J. Mol. Sci. 2013, 14, 10178–10196. [Google Scholar] [CrossRef] [PubMed]
- Sharaf, E.F.; Farrag, A.A. Induced resistance in tomato plants by IAA against Fusarium oxysporum lycopersici. Pol. J. Microbiol. 2004, 53, 111–116. [Google Scholar] [PubMed]
- Zheng, Y.; Sheng, J.; Zhao, R.; Zhang, J.; Lv, S.; Liu, L.; Shen, L. Preharvest l-arginine treatment induced postharvest disease resistance to Botrysis cinerea in tomato fruits. J. Agric. Food Chem. 2011, 59, 6543–6549. [Google Scholar] [CrossRef] [PubMed]
- Scalschi, L.; Vicedo, B.; Camañes, G.; Fernandez-Crespo, E.; Lapeña, L.; González-Bosch, C.; García-Agustín, P. Hexanoic acid is a resistance inducer that protects tomato plants against Pseudomonas syringae by priming the jasmonic acid and salicylic acid pathways. Mol. Plant Pathol. 2013, 14, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Vicedo, B.; Flors, V.; de la O Leyva, M.; Finiti, I.; Kravchuk, Z.; Real, M.D.; García-Agustín, P.; González-Bosch, C. Hexanoic acid-induced resistance against Botrytis cinerea in tomato plants. Mol. Plant Microbe Interact. 2009, 22, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Fontanilla, M.; Montes, M.; de Prado, R. Effects of the foliar-applied protein “Harpin (Ea)” (messenger) on tomatoes infected with Phytophthora infestans. Commun. Agric. Appl. Biol. Sci. 2004, 70, 41–45. [Google Scholar]
- Kiirika, L.M.; Stahl, F.; Wydra, K. Phenotypic and molecular characterization of resistance induction by single and combined application of chitosan and silicon in tomato against Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2013, 81, 1–12. [Google Scholar] [CrossRef]
- Mandal, S.; Kar, I.; Mukherjee, A.K.; Acharya, P. Elicitor-induced defense responses in Solanum lycopersicum against Ralstonia solanacearum. Sci. World J. 2013, 2013, 561056. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Feng, G.; Chen, K. Defense responses of harvested tomato fruit to burdock fructooligosaccharide, a novel potential elicitor. Postharvest Biol. Technol. 2009, 52, 110–116. [Google Scholar] [CrossRef]
- Großkinsky, D.; Edelsbrunner, K.; Pfeifhofer, H.; van der Graaff, E.; Roitsch, T. Cis- and trans-zeatin differentially modulate plant immunity. Plant Signal. Behav. 2013, 8, e24798. [Google Scholar] [CrossRef] [PubMed]
- Király, L.; Künstler, A.; Höller, K.; Fattinger, M.; Juhász, C.; Müller, M.; Gullner, G.; Zechmann, B. Sulfate supply influences compartment specific glutathione metabolism and confers enhanced resistance to Tobacco mosaic virus during a hypersensitive response. Plant Physiol. Biochem. 2012, 59, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y. 3-Aminobutyric acid induces systemic resistance against Peronospore tabacina. Physiol. Mol. Plant Pathol. 1994, 44, 273–288. [Google Scholar] [CrossRef]
- Lazzarato, L.; Trebbi, G.; Pagnucco, C.; Franchin, C.; Torrigiani, P.; Betti, L. Exogenous spermidine, arsenic and β-aminobutyric acid modulate tobacco resistance to tobacco mosaic virus, and affect local and systemic glucosylsalicylic acid levels and arginine decarboxylase gene expression in tobacco leaves. J. Plant Physiol. 2009, 166, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, J.; Orober, M.; Buchenauer, H. β-Aminobutyric acid-mediated enhancement of resistance in tobacco to tobacco mosaic virus depends on the accumulation of salicylic acid. Physiol. Mol. Plant Pathol. 2000, 56, 95–106. [Google Scholar] [CrossRef]
- Li, J.G.; Liu, H.X.; Cao, J.; Chen, L.F.; Gu, C.; Allen, C.; Guo, J.H. PopW of Ralstonia solanacearum, a new two-domain harpin targeting the plant cell wall. Mol. Plant Pathol. 2010, 11, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yang, X.; Qiu, D.; Guo, L.; Zeng, H.; Mao, J.; Gao, Q. PeaT1-induced systemic acquired resistance in tobacco follows salicylic acid-dependent pathway. Mol. Biol. Rep. 2011, 38, 2549–2556. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Feng, G.; Chen, K. Burdock fructooligosaccharide induces resistance to tobacco mosaic virus in tobacco seedlings. Physiol. Mol. Plant Pathol. 2009, 74, 34–40. [Google Scholar] [CrossRef]
- Monjil, M.S.; Shibata, Y.; Takemoto, D.; Kawakita, K. Bis-aryl methanone compound is a candidate of nitric oxide producing elicitor and induces resistance in Nicotiana benthamiana against Phytophthora infestans. Nitric Oxide 2013, 29, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Lobato, M.; Altamiranda, E.G.; Daleo, G.; Huarte, M.; Guevara, M.; Andreu, A. BABA effects on the behaviour of potato cultivars infected by Phytophthora infestans and Fusarium solani. Eur. J. Plant Pathol. 2009, 123, 47–56. [Google Scholar] [CrossRef]
- Altamiranda, E.A.G.; Andreu, A.B.; Daleo, G.R.; Olivieri, F.P. Effect of β-aminobutyric acid (BABA) on protection against Phytophthora infestans throughout the potato crop cycle. Australas. Plant Pathol. 2008, 37, 421–427. [Google Scholar] [CrossRef]
- Andreu, A.B.; Guevara, M.G.; Wolski, E.A.; Daleol, G.R.; Caldiz, D.O. Enhancement of natural disease resistance in potatoes by chemicals. Pest Manag. Sci. 2006, 62, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Si-Ammour, A.; Mauch-Mani, B.; Mauch, F. Quantification of induced resistance against Phytophthora species expressing GFP as a vital marker: β-Aminobutyric acid but not BTH protects potato and Arabidopsis from infection. Mol. Plant Pathol. 2003, 4, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Liljeroth, E.; Bengtsson, T.; Wiik, L.; Andreasson, E. Induced resistance in potato to Phytphthora infestans-effects of BABA in greenhouse and field tests with different potato varieties. Eur. J. Plant Pathol. 2010, 127, 171–183. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Singh, P.; Chen, C.-W.; Thomas, J.; Weber, J.; Mauch-Mani, B.; Zimmerli, L. Priming for enhanced defence responses by specific inhibition of the Arabidopsis response to coronatine. Plant J. 2011, 65, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, T.; Holefors, A.; Witzell, J.; Andreasson, E.; Liljeroth, E. Activation of defence responses to Phytophthora infestans in potato by BABA. Plant Pathol. 2014, 63, 193–202. [Google Scholar] [CrossRef]
- Cohen, Y.R. β-Aminobutyric acid-induced resistance against plant pathogens. Plant Dis. 2002, 86, 448–457. [Google Scholar] [CrossRef]
- Eschen-Lippold, L.; Altmann, S.; Rosahl, S. dl-β-aminobutyric acid-induced resistance of potato against Phytophthora infestans requires salicylic acid but not oxylipins. Mol. Plant Microbe Interact. 2010, 23, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Floryszak-Wieczorek, J.; Arasimowicz-Jelonek, M.; Milczarek, G.; Janus, L.; Pawlak-Sprada, S.; Abramowski, D.; Deckert, J.; Billert, H. Nitric oxide-mediated stress imprint in potato as an effect of exposure to a priming agent. Mol. Plant Microbe Interact. 2012, 25, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Baider, A.; Cohen, Y. Synergistic interaction between BABA and mancozeb in controlling Phytophthora infestans in potato and tomato and Pseudoperonospora cubensis in cucumber. Phytoparasitica 2003, 31, 399–409. [Google Scholar] [CrossRef]
- Czajkowski, R.; Pérombelon, M.; Jafra, S.; Lojkowska, E.; Potrykus, M.; van der Wolf, J.; Sledz, W. Detection, identification and differentiation of Pectobacterium and Dickeya species causing potato blackleg and tuber soft rot: A review. Ann. Appl. Biol. 2015, 166, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Moushib, L.I.; Witzell, J.; Lenman, M.; Liljeroth, E.; Andreasson, E. Sugar beet extract induces defence against Phytophthora infestans in potato plants. Eur. J. Plant Pathol. 2013, 136, 261–271. [Google Scholar] [CrossRef]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J.; Drzewiecka, K.; Chmielowska-Bąk, J.; Abramowski, D.; Izbiańska, K. Aluminum induces cross-resistance of potato to Phytophthora infestans. Planta 2014, 239, 679–694. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, L.; Lu, G.; Zhan, X.-B.; Lin, C.-C.; Zheng, Z.-Y. Curdlan β-1,3-Glucooligosaccharides Induce the Defense Responses against Phytophthora infestans Infection of Potato (Solanum tuberosum L. cv. McCain G1) Leaf Cells. PLoS ONE 2014, 9, e97197. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y.; Gisi, U.; Mosinger, E. Systemic resistance of potato plants against Phytophthora infestans induced by unsaturated fatty acids. Physiol. Mol. Plant Pathol. 1991, 38, 255–263. [Google Scholar] [CrossRef]
- Burra, D.D.; Berkowitz, O.; Hedley, P.E.; Morris, J.; Resjö, S.; Levander, F.; Liljeroth, E.; Andreasson, E.; Alexandersson, E. Phosphite-induced changes of the transcriptome and secretome in Solanum tuberosum leading to resistance against Phytophthora infestans. BMC Plant Biol. 2014, 14, 254. [Google Scholar] [CrossRef] [PubMed]
- Liljeroth, E.; Lankinen, Å.; Wiik, L.; Burra, D.D.; Alexandersson, E.; Andreasson, E. Potassium phosphite combined with reduced doses of fungicides provides efficient protection against potato late blight in large-scale field trials. Crop Prot. 2016, 86, 42–55. [Google Scholar] [CrossRef]
- Lim, S.; Borza, T.; Peters, R.D.; Coffin, R.H.; Al-Mughrabi, K.I.; Pinto, D.M.; Wang-Pruski, G. Proteomics analysis suggests broad functional changes in potato leaves triggered by phosphites and a complex indirect mode of action against Phytophthora infestans. J. Proteom. 2013, 93, 207–223. [Google Scholar] [CrossRef] [PubMed]
- Lobato, M.C.; Machinandiarena, M.F.; Tambascio, C.; Dosio, G.A.; Caldiz, D.O.; Daleo, G.R.; Andreu, A.B.; Olivieri, F.P. Effect of foliar applications of phosphite on post-harvest potato tubers. Eur. J. Plant Pathol. 2011, 130, 155–163. [Google Scholar] [CrossRef]
- Lobato, M.C.; Olivieri, F.P.; Gonzalez Altamiranda, E.A.; Wolski, E.A.; Daleo, G.R.; Caldiz, D.O.; Andreu, A.B. Phosphite compounds reduce disease severity in potato seed tubers and foliage. Eur. J. Plant Pathol. 2008, 122, 349–358. [Google Scholar] [CrossRef]
- Machinandiarena, M.F.; Lobato, M.C.; Feldman, M.L.; Daleo, G.R.; Andreu, A.B. Potassium phosphite primes defense responses in potato against Phytophthora infestans. J. Plant Physiol. 2012, 169, 1417–1424. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.S.; Olsen, N.; Woodell, L.; Porter, L.D.; Clayson, S. Post-harvest applications of zoxamide and phosphite for control of potato tuber rots caused by oomycetes at harvest. Am. J. Potato Res. 2006, 83, 269–278. [Google Scholar] [CrossRef]
- Beckers, G.J.; Conrath, U. Priming for stress resistance: From the lab to the field. Curr. Opin. Plant Biol. 2007, 10, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Van Kan, J.A.; Cozijnsen, T.; Danhash, N.; de Wit, P.J. Induction of tomato stress protein mRNAs by ethephon, 2,6-dichloroisonicotinic acid and salicylate. Plant Mol. Biol. 1995, 27, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Tally, A.; Oostendorp, M.; Lawton, K.; Staub, T.; Bassi, B.; Agrawal, A.; Tuzun, S.; Bent, E. Commercial development of elicitors of induced resistance to pathogens. In Induced Plant Defenses Against Pathogens And Herbivores: Biochemistry, Ecology, And Agriculture; Agrawal, A.A., Tuzun, S., Bent, E., Eds.; APS press: St. Paul, MN, USA, 1999; pp. 357–369. [Google Scholar]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.-Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809. [Google Scholar] [CrossRef] [PubMed]
- Harel, Y.M.; Mehari, Z.H.; Rav-David, D.; Elad, Y. Systemic resistance to gray mold induced in tomato by benzothiadiazole and Trichoderma harzianum T39. Phytopathology 2014, 104, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M.; Nishioka, M.; Nakashita, H.; Yamaguchi, I.; Yoshida, S. Pyrazolecarboxylic acid derivative induces systemic acquired resistance in tobacco. Biosci. Biotechnol. Biochem. 2003, 67, 2614–2620. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Bruns, C.; Butz, A.; Finckh, M. Effects of fertilizers and plant strengtheners on the susceptibility of tomatoes to single and mixed isolates of Phytophthora infestans. Eur. J. Plant Pathol. 2012, 133, 739–751. [Google Scholar] [CrossRef]
- Šašek, V.; Nováková, M.; Dobrev, P.I.; Valentová, O.; Burketová, L. β-Aminobutyric acid protects Brassica napus plants from infection by Leptosphaeria maculans. Resistance induction or a direct antifungal effect? Eur. J. Plant Pathol. 2012, 133, 279–289. [Google Scholar] [CrossRef]
- Deliopoulos, T.; Kettlewell, P.S.; Hare, M.C. Fungal disease suppression by inorganic salts: A review. Crop Prot. 2010, 29, 1059–1075. [Google Scholar] [CrossRef]
- McDonald, A.E.; Grant, B.R.; Plaxton, W.C. Phosphite (phosphorous acid): Its relevance in the environment and agriculture and influence on plant phosphate starvation response. J. Plant Nutr. 2001, 24, 1505–1519. [Google Scholar] [CrossRef]
- Thao, H.T.B.; Yamakawa, T. Phosphite (phosphorous acid): Fungicide, fertilizer or bio-stimulator? Soil Sci. Plant Nutr. 2009, 55, 228–234. [Google Scholar] [CrossRef]
- Olivieri, F.; Feldman, M.; Machinandiarena, M.; Lobato, M.; Caldiz, D.; Daleo, G.; Andreu, A. Phosphite applications induce molecular modifications in potato tuber periderm and cortex that enhance resistance to pathogens. Crop Prot. 2012, 32, 1–6. [Google Scholar] [CrossRef]
- Lobato, M.; Olivieri, F.; Daleo, G.; Andreu, A. Antimicrobial activity of phosphites against different potato pathogens. J. Plant Dis. Prot. 2010, 117, 102–109. [Google Scholar] [CrossRef]
- Bashan, B.; Levy, Y.; Cohen, Y. Variation in sensitivity of Phytophthora infestans to fosetyl-Al. Plant Pathol. 1990, 39, 134–140. [Google Scholar] [CrossRef]
- Berkowitz, O.; Jost, R.; Kollehn, D.O.; Fenske, R.; Finnegan, P.M.; O’Brien, P.A.; Hardy, G.E.S.J.; Lambers, H. Acclimation responses of Arabidopsis thaliana to sustained phosphite treatments. J. Exp. Bot. 2013, 64, 1731–1743. [Google Scholar] [CrossRef] [PubMed]
- Danova-Alt, R.; Dijkema, C.; de Waard, P.; Koeck, M. Transport and compartmentation of phosphite in higher plant cells—Kinetic and 31P nuclear magnetic resonance studies. Plant Cell Environ. 2008, 31, 1510–1521. [Google Scholar] [CrossRef] [PubMed]
- Burketova, L.; Trda, L.; Ott, P.G.; Valentova, O. Bio-based resistance inducers for sustainable plant protection against pathogens. Biotechnol. Adv. 2015, 33, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Wiesel, L.; Newton, A.C.; Elliott, I.; Booty, D.; Gilroy, E.M.; Birch, P.R.; Hein, I. Molecular effects of resistance elicitors from biological origin and their potential for crop protection. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Scheler, C.; Durner, J.; Astier, J. Nitric oxide and reactive oxygen species in plant biotic interactions. Curr. Opin. Plant Biol. 2013, 16, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Monjil, M.S.; Takemoto, D.; Kawakita, K. Defense induced by a bis-aryl methanone compound leads to resistance in potato against Phytophthora infestans. J. Gen. Plant Pathol. 2014, 80, 38–49. [Google Scholar] [CrossRef]
- Leyva, M.; Vicedo, B.; Finiti, I.; Flors, V.; del Amo, G.; Real, M.; García-Agustín, P.; González-Bosch, C. Preventive and post-infection control of Botrytis cinerea in tomato plants by hexanoic acid. Plant Pathol. 2008, 57, 1038–1046. [Google Scholar] [CrossRef]
- Veluchamy, S.; Hind, S.R.; Dunham, D.M.; Martin, G.B.; Panthee, D.R. Natural variation for responsiveness to flg22, flgII-28, and csp22 and Pseudomonas syringae pv. tomato in heirloom tomatoes. PLoS ONE 2014, 9, e106119. [Google Scholar]
- Desender, S.; Klarzynski, O.; Potin, P.; Barzic, M.R.; Andrivon, D.; Val, F. Lipopolysaccharides of Pectobacterium atrosepticum and Pseudomonas corrugata induce different defence response patterns in tobacco, tomato, and potato. Plant Biol. 2006, 8, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, T.; Holefors, A.; Liljeroth, E.; Hultberg, M.; Andreasson, E. Biosurfactants have the potential to induce defence against Phytophthora infestans in Potato. Potato Res. 2015, 58, 83–90. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Ortíz, H.; Benavides-Mendoza, A.; Flores-Olivas, A.; Ledezma-Pérez, A. Use of the interpolyelectrolyte complexes of poly(acrylic acid)-chitosan as inductors of tolerance against pathogenic fungi in Tomato (Lycopersicon esculentum Mill. var. Floradade). Macromol. Biosci. 2003, 3, 566–570. [Google Scholar] [CrossRef]
- Guo, W.; Yin, H.; Ye, Z.; Zhao, X.; Yuan, J.; Du, Y. A comparison study on the interactions of two oligosaccharides with tobacco cells by time-resolved fluorometric method. Carbohydr. Polym. 2012, 90, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Thuerig, B.; Binder, A.; Boller, T.; Guyer, U.; Jiménez, S.; Rentsch, C.; Tamm, L. An aqueous extract of the dry mycelium of Penicillium chrysogenum induces resistance in several crops under controlled and field conditions. Eur. J. Plant Pathol. 2006, 114, 185–197. [Google Scholar] [CrossRef]
- Lyon, G. Agents that can elicit induced resistance. In Induced Resistance for Plant Defence. A Sustainable Approach to Crop Protection; Blackwell Publishing Ltd.: Oxford, UK, 2007; pp. 9–29. [Google Scholar]
- Ahn, I.-P.; Kim, S.; Lee, Y.-H. Vitamin B1 functions as an activator of plant disease resistance. Plant Physiol. 2005, 138, 1505–1515. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wei, F.; Wang, L.; Liu, H.; Zhu, X.; Liang, Y. Riboflavin activates defense responses in tobacco and induces resistance against Phytophthora parasitica and Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2010, 74, 330–336. [Google Scholar] [CrossRef]
- Rodewald, J.; Trognitz, B. Solanum resistance genes against Phytophthora infestans and their corresponding avirulence genes. Mol. Plant Pathol. 2013, 14, 740–757. [Google Scholar] [CrossRef] [PubMed]
- Ercolano, M.; Sanseverino, W.; Carli, P.; Ferriello, F.; Frusciante, L. Genetic and genomic approaches for R-gene mediated disease resistance in tomato: Retrospects and prospects. Plant Cell Rep. 2012, 31, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Pajerowska, K.M.; Parker, J.E.; Gebhardt, C. Potato homologs of Arabidopsis thaliana genes functional in defense signaling—Identification, genetic mapping, and molecular cloning. Mol. Plant Microbe Interact. 2005, 18, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Mitton, F.M.; Pinedo, M.L.; de la Canal, L. Phloem sap of tomato plants contains a DIR1 putative ortholog. J. Plant Physiol. 2009, 166, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Frades, I.; Abreha, K.B.; Proux-Wéra, E.; Lankinen, Å.; Andreasson, E.; Alexandersson, E. A novel workflow correlating RNA-seq data to resistance levels of wild Solanum species and potato clones to Phytophthora infestans. Front. Plant Sci. 2015, 6, 718. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U.; Linke, C.; Jeblick, W.; Geigenberger, P.; Quick, W.P.; Neuhaus, H.E. Enhanced resistance to Phytophthora infestans and Alternaria solani in leaves and tubers, respectively, of potato plants with decreased activity of the plastidic ATP/ADP transporter. Planta 2003, 217, 75–83. [Google Scholar] [PubMed]
- Hwang, I.S.; Choi, D.S.; Kim, N.H.; Kim, D.S.; Hwang, B.K. Pathogenesis-related protein 4b interacts with leucine-rich repeat protein 1 to suppress PR4b-triggered cell death and defense response in pepper. Plant J. 2014, 77, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Kim, N.H.; Hwang, B.K. The Capsicum annuum class IV chitinase ChitIV interacts with receptor-like cytoplasmic protein kinase PIK1 to accelerate PIK1-triggered cell death and defence responses. J. Exp. Bot. 2015, 66, 1987–1999. [Google Scholar] [CrossRef] [PubMed]
- Pajerowska-Mukhtar, K.M.; Mukhtar, M.S.; Guex, N.; Halim, V.A.; Rosahl, S.; Somssich, I.E.; Gebhardt, C. Natural variation of potato allene oxide synthase 2 causes differential levels of jasmonates and pathogen resistance in Arabidopsis. Planta 2008, 228, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Angulo, C.; Leyva, M.D.L.O.; Finiti, I.; López-Cruz, J.; Fernández-Crespo, E.; García-Agustín, P.; González-Bosch, C. Role of dioxygenase α-DOX2 and SA in basal response and in hexanoic acid-induced resistance of tomato (Solanum lycopersicum) plants against Botrytis cinerea. J. Plant Physiol. 2015, 175, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Manosalva, P.M.; Park, S.-W.; Forouhar, F.; Tong, L.; Fry, W.E.; Klessig, D.F. Methyl esterase 1 (StMES1) is required for systemic acquired resistance in potato. Mol. Plant Microbe Interact. 2010, 23, 1151–1163. [Google Scholar] [CrossRef] [PubMed]
- Oka, K.; Amano, Y.; Katou, S.; Seo, S.; Kawazu, K.; Mochizuki, A.; Kuchitsu, K.; Mitsuhara, I. Tobacco MAP kinase phosphatase (NtMKP1) negatively regulates wound response and induced resistance against necrotrophic pathogens and lepidopteran herbivores. Mol. Plant Microbe Interact. 2013, 26, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Sarowar, S.; Kim, Y.J.; Kim, K.D.; Hwang, B.K.; Ok, S.H.; Shin, J.S. Overexpression of lipid transfer protein (LTP) genes enhances resistance to plant pathogens and LTP functions in long-distance systemic signaling in tobacco. Plant Cell Rep. 2009, 28, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Manosalva, P.; Manohar, M.; Kogel, K.-H.; Kang, H.-G.; Klessig, D.F. The GHKL ATPase MORC1 Modulates Species-Specific Plant Immunity in Solanaceae. Mol. Plant Microbe Interact. 2015, 28, 927–942. [Google Scholar] [CrossRef] [PubMed]
- Hoeberichts, F.A.; Davoine, C.; Vandorpe, M.; Morsa, S.; Ksas, B.; Stassen, C.; Triantaphylidès, C.; van Breusegem, F. Cryptogein-induced transcriptional reprogramming in tobacco is light dependent. Plant Physiol. 2013, 163, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Amelot, N.; Carrouche, A.; Danoun, S.; Bourque, S.; Haiech, J.; Pugin, A.; Ranjeva, R.; Grima-Pettenati, J.; Mazars, C.; Briere, C. Cryptogein, a fungal elicitor, remodels the phenylpropanoid metabolism of tobacco cell suspension cultures in a calcium-dependent manner. Plant Cell Environ. 2011, 34, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Dokládal, L.; Obořil, M.; Stejskal, K.; Zdráhal, Z.; Ptáčková, N.; Chaloupková, R.; Damborský, J.; Kašparovský, T.; Jeandroz, S.; Žd'árská, M. Physiological and proteomic approaches to evaluate the role of sterol binding in elicitin-induced resistance. J. Exp. Bot. 2012, 63, 2203–2215. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Z.; Li, X.; Huang, L.; Hong, Y.; Zhang, Y.; Zhang, H.; Li, D.; Song, F. Elicitin-like proteins Oli-D1 and Oli-D2 from Pythium oligandrum trigger hypersensitive response in Nicotiana benthamiana and induce resistance against Botrytis cinerea in tomato. Mol. Plant Pathol. 2015, 16, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tian, Z.; Liu, J.; Xie, C. Comparative cDNA-AFLP analysis reveals that dl-β-amino-butyric acid induces resistance through early activation of the host-defense genes in potato. Physiol. Plant. 2009, 136, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Arasimowicz-Jelonek, M.; Kosmala, A.; Janus, Ł.; Abramowski, D.; Floryszak-Wieczorek, J. The proteome response of potato leaves to priming agents and S-nitrosoglutathione. Plant Sci. 2013, 198, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; Jakab, G.; Toquin, V.; Flors, V.; Iavicoli, A.; Maeder, M.N.; Metraux, J.-P.; Mauch-Mani, B. Dissecting the β-aminobutyric acid-induced priming phenomenon in Arabidopsis. Plant Cell 2005, 17, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.; van Hulten, M.; Zhang, Y.; Berkowitz, O.; López, A.; Pétriacq, P.; Sellwood, M.A.; Chen, B.; Burrell, M.; van de Meene, A. Plant perception of β-aminobutyric acid is mediated by an aspartyl-tRNA synthetase. Nat. Chem. Biol. 2014, 10, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Massoud, K.; Barchietto, T.; Le Rudulier, T.; Pallandre, L.; Didierlaurent, L.; Garmier, M.; Ambard-Bretteville, F.; Seng, J.-M.; Saindrenan, P. Dissecting phosphite-induced priming in Arabidopsis infected with Hyaloperonospora arabidopsidis. Plant Physiol. 2012, 159, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Ghareeb, H.; Bozsó, Z.; Ott, P.G.; Repenning, C.; Stahl, F.; Wydra, K. Transcriptome of silicon-induced resistance against Ralstonia solanacearum in the silicon non-accumulator tomato implicates priming effect. Physiol. Mol. Plant Pathol. 2011, 75, 83–89. [Google Scholar] [CrossRef]
- Rosli, H.G.; Zheng, Y.; Pombo, M.A.; Zhong, S.; Bombarely, A.; Fei, Z.; Collmer, A.; Martin, G.B. Transcriptomics-based screen for genes induced by flagellin and repressed by pathogen effectors identifies a cell wall-associated kinase involved in plant immunity. Genome Biol. 2013, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Chen, K.; Zhang, P. Transcriptome profile analysis of resistance induced by burdock fructooligosaccharide in tobacco. J. Plant Physiol. 2012, 169, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Finiti, I.; O Leyva, M.; Vicedo, B.; Gómez-Pastor, R.; López-Cruz, J.; García-Agustín, P.; Real, M.D.; González-Bosch, C. Hexanoic acid protects tomato plants against Botrytis cinerea by priming defence responses and reducing oxidative stress. Mol. Plant Pathol. 2014, 15, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Lepère, G.; Jay, F.; Wang, J.; Bapaume, L.; Wang, Y.; Abraham, A.-L.; Penterman, J.; Fischer, R.L.; Voinnet, O. Dynamics and biological relevance of DNA demethylation in Arabidopsis antibacterial defense. Proc. Natl. Acad. Sci. USA 2013, 110, 2389–2394. [Google Scholar] [CrossRef] [PubMed]
- Van Oosten, M.J.; Bressan, R.A.; Zhu, J.-K.; Bohnert, H.J.; Chinnusamy, V. The role of the epigenome in gene expression control and the epimark changes in response to the environment. Crit. Rev. Plant Sci. 2014, 33, 64–87. [Google Scholar] [CrossRef]
- Zhu, Q.-H.; Shan, W.-X.; Ayliffe, M.; Wang, M.-B. Epigenetic mechanisms: An emerging player in plant-microbe interactions. Mol. Plant Microbe Interact. 2015, 29, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Johannes, F.; Porcher, E.; Teixeira, F.K.; Saliba-Colombani, V.; Simon, M.; Agier, N.; Bulski, A.; Albuisson, J.; Heredia, F.; Audigier, P. Assessing the impact of transgenerational epigenetic variation on complex traits. PLoS Genet. 2009, 5, e1000530. [Google Scholar] [CrossRef] [PubMed]
- Reinders, J.; Wulff, B.B.; Mirouze, M.; Marí-Ordóñez, A.; Dapp, M.; Rozhon, W.; Bucher, E.; Theiler, G.; Paszkowski, J. Compromised stability of DNA methylation and transposon immobilization in mosaic Arabidopsis epigenomes. Genes Dev. 2009, 23, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.K.; Heredia, F.; Sarazin, A.; Roudier, F.; Boccara, M.; Ciaudo, C.; Cruaud, C.; Poulain, J.; Berdasco, M.; Fraga, M.F. A role for RNAi in the selective correction of DNA methylation defects. Science 2009, 323, 1600–1604. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, A.; Daniel, X.; Flors, V.; Luna, E.; Hohn, B.; Mauch-Mani, B. Descendants of Primed Arabidopsis Plants Exhibit Resistance to Biotic Stress. Plant Physiol. 2012, 158, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D. Acquired resistance to tobacco mosaic virus transmitted to the progeny of hypersensitive tobacco. Virology 1983, 124, 161–163. [Google Scholar] [CrossRef]
- Kathiria, P.; Sidler, C.; Golubov, A.; Kalischuk, M.; Kawchuk, L.M.; Kovalchuk, I. Tobacco mosaic virus infection results in an increase in recombination frequency and resistance to viral, bacterial, and fungal pathogens in the progeny of infected tobacco plants. Plant Physiol. 2010, 153, 1859–1870. [Google Scholar] [CrossRef] [PubMed]
- Lankinen, Å.; Abreha, K.B.; Alexandersson, E.; Andersson, S.; Andreasson, E. Nongenetic Inheritance of Induced Resistance in a Wild Annual Plant. Phytopathology 2016, 106, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Mandal, R.; Kathiria, P.; Psychogios, N.; Bouatra, S.; Krishnamurthy, R.; Wishart, D.; Kovalchuk, I. Progeny of tobacco mosaic virus-infected Nicotiana tabacum plants exhibit trans-generational changes in metabolic profiles. Biocatal. Agric. Biotechnol. 2012, 1, 115–123. [Google Scholar]
- Herrera, C.M.; Bazaga, P. Untangling individual variation in natural populations: Ecological, genetic and epigenetic correlates of long-term inequality in herbivory. Mol. Ecol. 2011, 20, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Leimar, O.; McNamara, J.M. The evolution of transgenerational integration of information in heterogeneous environments. Am. Nat. 2015, 185, E55–E69. [Google Scholar] [CrossRef] [PubMed]
- English, S.; Pen, I.; Shea, N.; Uller, T. The information value of non-genetic inheritance in plants and animals. PLoS ONE 2015, 10, e0116996. [Google Scholar]
- Nguyen, D.; D’Agostino, N.; Tytgat, T.O.; Sun, P.; Lortzing, T.; Visser, E.J.; Cristescu, S.M.; Steppuhn, A.; Mariani, C.; Dam, N.M. Drought and flooding have distinct effects on herbivore-induced responses and resistance in Solanum dulcamara. Plant Cell Environ. 2016, 39, 1485–1499. [Google Scholar] [CrossRef] [PubMed]
- Cruickshank, I.; Mandryk, M. The effect of stem infestation of Tobacco with Peronospora tabacina Adam, on foliage reaction to blue mold. J. Aust. Inst. Agric. Sci. 1960, 26, 369–372. [Google Scholar]
- Vallad, G.E.; Goodman, R.M. Systemic acquired resistance and induced systemic resistance in conventional agriculture. Crop Sci. 2004, 44, 1920–1934. [Google Scholar] [CrossRef]
- Lankinen, Å.; Abreha, K.B.; Bakhshandeh, M.; Masini, L.; Ali, A.; Andreasson, E. Plant immunity is seldom activated in natural populations and agricultural fields. In Proceedings of the Wild Plant Pathosystems 2016, Helsinki, Filand, 29–31 August 2016; p. 45.
- Alexandersson, E.; Jacobson, D.; Vivier, M.A.; Weckwerth, W.; Andreasson, E. Field-omics—Understanding large-scale molecular data from field crops. Front. Plant Sci. 2014, 5, 286. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.S.; Yang, J.W.; Ryu, C.M. ISR meets SAR outside: Additive action of the endophyte Bacillus pumilus INR7 and the chemical inducer, benzothiadiazole, on induced resistance against bacterial spot in field-grown pepper. Front. Plant Sci. 2013, 4, 122. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-H.; Vallad, G.E.; Zhang, S.; Wen, A.; Balogh, B.; Figueiredo, J.F.L.; Behlau, F.; Jones, J.B.; Momol, M.T.; Olson, S.M. Effect of application frequency and reduced rates of acibenzolar-S-methyl on the field efficacy of induced resistance against bacterial spot on tomato. Plant Dis. 2012, 96, 221–227. [Google Scholar] [CrossRef]
- Cooke, L.R.; Little, G. The effect of foliar application of phosphonate formulations on the susceptibility of potato tubers to late blight. Pest Manag. Sci. 2002, 58, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Mayton, H.; Myers, K.; Fry, W.E. Potato late blight in tubers—The role of foliar phosphonate applications in suppressing pre-harvest tuber infections. Crop Prot. 2008, 27, 943–950. [Google Scholar] [CrossRef]
- Wang-Pruski, G.; Lim, S.; Coffin, R.H.; Peters, R.D.; Platt, H.W.; Al-Mughrabi, K.I.; Pinto, D.; Veenhuis-Mac-Neill, S.; Hardy, W.; Macdonald, I.; et al. Foliar application of phosphorous acid for control of potato late blight. Can. J. Plant Pathol. 2015, 32, 420. [Google Scholar]
- Mulugeta, T.; Andreasson, E.; Liljeroth, E.; Aregay, B.; Alexandersson, E. The effect of potassium phosphite in field-grown potato and tomato in Ethiopia (manuscript in preparation).
- Taylor, R.J.; Pasche, J.S.; Gudmestad, N.C. Effect of application method and rate on residual efficacy of mefenoxam and phosphorous acid fungicides in the control of pink rot of potato. Plant Dis. 2011, 95, 997–1006. [Google Scholar] [CrossRef]
- Johnson, S.B. Postharvest applications of phosphorous acid for control of Phytophthora infestans on potatoes. In Proceedings of the Twelfth EuroBlight Workshop, Arras, France, 3–6 May 2010; pp. 3–6.
- Cohen, Y.; Coffey, M.D. Systemic fungicides and the control of oomycetes. Annu. Rev. Phytopathol. 1986, 24, 311–338. [Google Scholar] [CrossRef]
- Tamm, L.; Thürig, B.; Fliessbach, A.; Goltlieb, A.; Karavani, S.; Cohen, Y. Elicitors and soil management to induce resistance against fungal plant diseases. Wagening. J. Life Sci. 2011, 58, 131–137. [Google Scholar] [CrossRef]
- Van Leeuwen, H.; Kliebenstein, D.J.; West, M.A.; Kim, K.; van Poecke, R.; Katagiri, F.; Michelmore, R.W.; Doerge, R.W.; Clair, D.A.S. Natural variation among Arabidopsis thaliana accessions for transcriptome response to exogenous salicylic acid. Plant Cell 2007, 19, 2099–2110. [Google Scholar] [CrossRef] [PubMed]
- Dugé de Bernonville, T.; Marolleau, B.; Staub, J.; Gaucher, M.; Brisset, M.-N.L. Using molecular tools to decipher the complex world of plant resistance inducers: An apple case study. J. Agric. Food Chem. 2014, 62, 11403–11411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima-Silva, V.; Rosado, A.; Amorim-Silva, V.; Muñoz-Mérida, A.; Pons, C.; Bombarely, A.; Trelles, O.; Fernández-Muñoz, R.; Granell, A.; Valpuesta, V. Genetic and genome-wide transcriptomic analyses identify co-regulation of oxidative response and hormone transcript abundance with vitamin C content in tomato fruit. BMC Genom. 2012, 13, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]



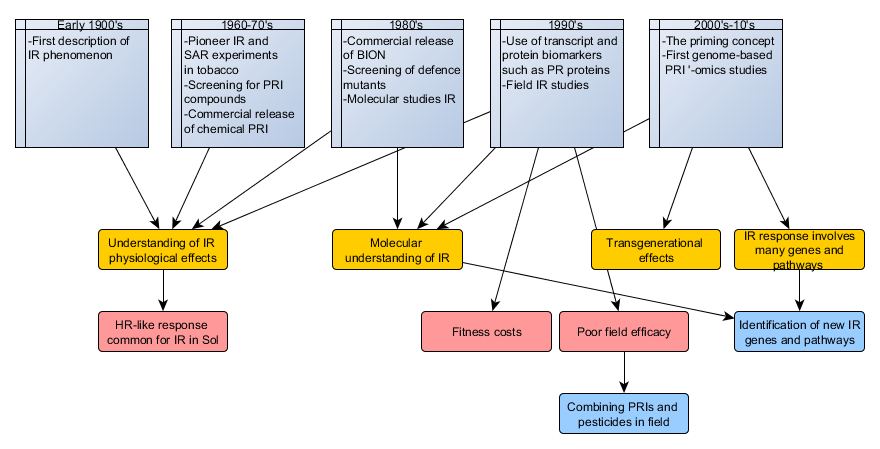{kind=link}
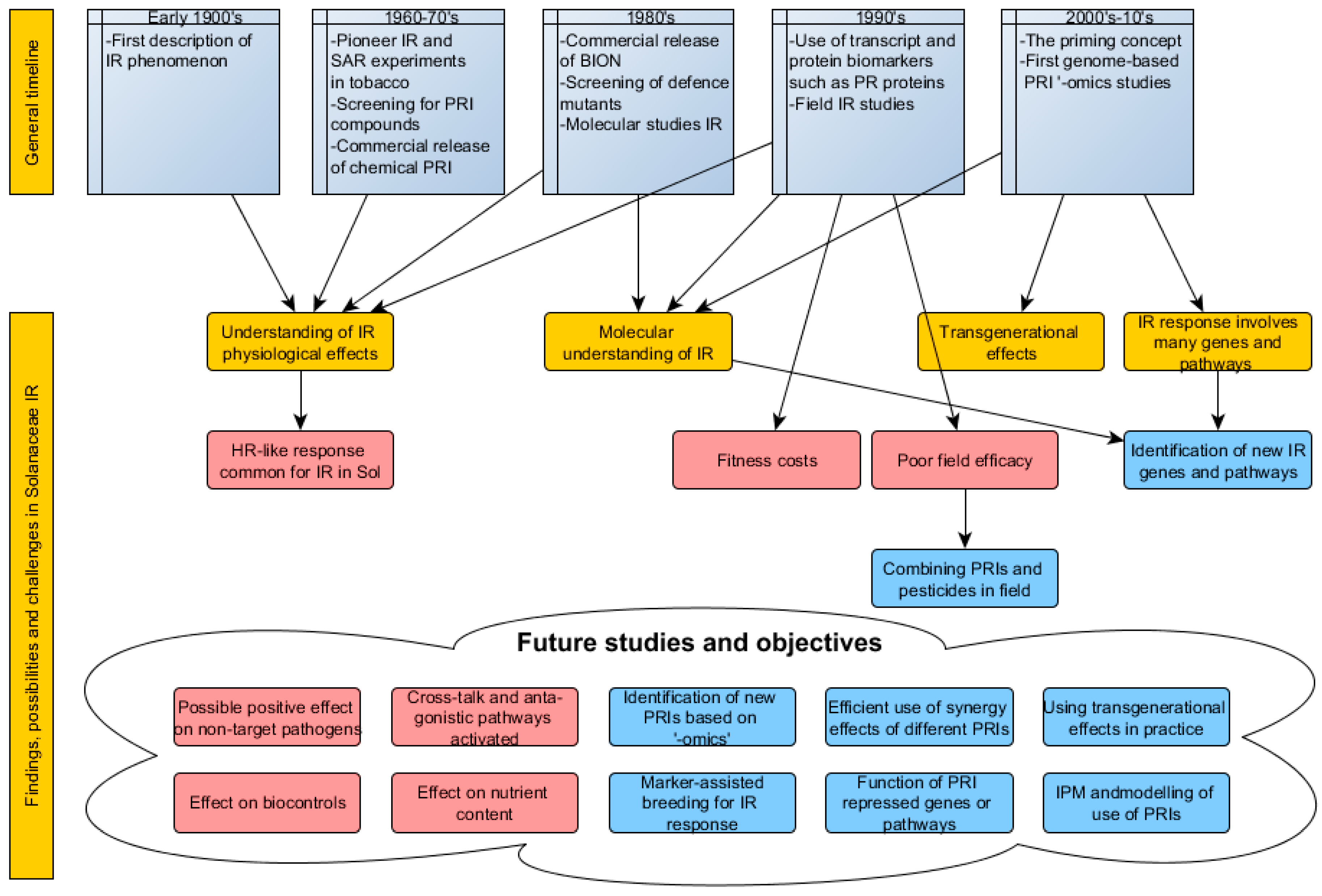{kind=link}
{kind=link}
| Solanaceae Species | Plant | Inducer | Protection against | Organism | Reference |
|---|---|---|---|---|---|
| Capsicum annuum | Pepper | BABA | Colletotrichum coccodes | Fungus | [27] |
| Capsicum annuum | Pepper | BABA | Phytophthora capsici | Oomycete | [28,29] |
| Capsicum annuum | Pepper | Monopotassium phosphate | Powdery mildew | Fungus | [30] |
| Capsicum annuum | Pepper | Phosphite | Phytophthora capsici | Oomycete | [31] |
| Capsicum annuum | Pepper | Bacterial VOCs | Xanthomonas axonopodis | Bacteria | [32] |
| Capsicum annuum | Pepper | Biochar | Leveillula taurica | Fungus | [33] |
| Solanum lycopersicum | Tomato | BABA | Clavibacter michiganensis | Bacteria | [34,35] |
| Solanum lycopersicum | Tomato | BTH | Pseudomonas syringae | Bacteria | [36] |
| Solanum lycopersicum | Tomato | BABA | Ralstonia solanacearum | Bacteria | [37] |
| Solanum lycopersicum | Tomato | BABA, seed treatment | Oidium neolycopersici | Fungus | [38] |
| Solanum lycopersicum | Tomato | BABA | Phytophthora infestans | Oomycete | [39] |
| Solanum lycopersicum | Tomato | BTH | Botrytis cinerea | Fungus | [40] |
| Solanum lycopersicum | Tomato | SA | Ralstonia solanacearum | Bacteria | [41] |
| Solanum lycopersicum | Tomato | IAA | Fusarium oxysporum | Fungus | [42] |
| Solanum lycopersicum | Tomato | L-arginine, post-harvest | Botrytis cinerea | Fungus | [43] |
| Solanum lycopersicum | Tomato | Hexanoic acid | Pseudomonas syringae | Bacteria | [44] |
| Solanum lycopersicum | Tomato | Hexanoic acid | Botrytis cinerea | Fungus | [45] |
| Solanum lycopersicum | Tomato | bacterial Harpin protein | Phytophthora infestans | Oomycete | [46] |
| Solanum lycopersicum | Tomato | Chitosan | Ralstonia solanacearum | Bacteria | [47,48] |
| Solanum lycopersicum | Tomato | Biochar | Botrytis cinerea | Fungus | [33] |
| Solanum lycopersicum | Tomato | fructooligosaccharide | Botrytis cinerea | Fungus | [49] |
| Nicotiana tabacum | Tobacco | Zeatin | Pseudomonas syringae | Bacteria | [50] |
| Nicotiana tabacum | Tobacco | Sulfur | Tobacco mosaic virus | Virus | [51] |
| Nicotiana tabacum | Tobacco | BABA | Peronospora tabacina | Oomycete | [52] |
| Nicotiana tabacum | Tobacco | BABA | Tobacco mosaic virus | Virus | [53,54] |
| Nicotiana tabacum | Tobacco | Bacterial harpin protein | Tobacco mosaic virus | Virus | [55] |
| Nicotiana tabacum | Tobacco | PeaT1 | Tobacco mosaic virus | Virus | [56] |
| Nicotiana tabacum | Tobacco | fructooligosaccharide | Tobacco mosaic virus | Virus | [57] |
| Nicotiana benthamiana | Tobacco relative | NUBS-4190 | Phytophthora infestans | Oomycete | [58] |
| Solanum tuberosum | Potato | BABA | Fusarium solani | Fungus | [59] |
| Solanum tuberosum | Potato | BABA | Phytophthora infestans | Oomycete | [39,59,60,61,62,63,64,65,66,67,68,69] |
| Solanum tuberosum | Potato | SA | Dickeya solani | Bacteria | [70] |
| Solanum tuberosum | Potato | SBE | Phytophthora infestans | Oomycete | [71] |
| Solanum tuberosum | Potato | Aluminium | Phytophthora infestans | Oomycete | [72] |
| Solanum tuberosum | Potato | Curdlan | Phytophthora infestans | Oomycete | [73] |
| Solanum tuberosum | Potato | Linoleic acid | Phytophthora infestans | Oomycete | [74] |
| Solanum tuberosum | Potato | Oleic acid | Phytophthora infestans | Oomycete | [74] |
| Solanum tuberosum | Potato | Phosphite | Phytophthora infestans | Oomycete | [75,76,77,78,79,80,81] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexandersson, E.; Mulugeta, T.; Lankinen, Å.; Liljeroth, E.; Andreasson, E. Plant Resistance Inducers against Pathogens in Solanaceae Species—From Molecular Mechanisms to Field Application. Int. J. Mol. Sci. 2016, 17, 1673. https://doi.org/10.3390/ijms17101673
Alexandersson E, Mulugeta T, Lankinen Å, Liljeroth E, Andreasson E. Plant Resistance Inducers against Pathogens in Solanaceae Species—From Molecular Mechanisms to Field Application. International Journal of Molecular Sciences. 2016; 17(10):1673. https://doi.org/10.3390/ijms17101673
Chicago/Turabian StyleAlexandersson, Erik, Tewodros Mulugeta, Åsa Lankinen, Erland Liljeroth, and Erik Andreasson. 2016. "Plant Resistance Inducers against Pathogens in Solanaceae Species—From Molecular Mechanisms to Field Application" International Journal of Molecular Sciences 17, no. 10: 1673. https://doi.org/10.3390/ijms17101673